Abstract
A field study was conducted in Thailand under a rain-fed environment to determine the effect of four different cutting heights above ground level and two closing dates on dry matter (DM) production, yield components, and fodder quality of five different cultivars of Napier grass (Pennisetum purpureum Schumach), all treatments cut with the same frequency of approximately 4 weeks. Eighteen quantitative trait measurements were used to investigate the interactions between cultivar diversity and cutting regimes. Principal-component analysis (PCA) showed a clustering of two distinct cultivars and another group of cultivars in one cluster without obvious structure. There was also a clear clustering of the control (0 cm cutting height) across all cultivars whereas other cutting heights affected growth differently according to cultivar. The optimal cutting regime for obtaining high DM yield depended on the genetic background and did not relate to a reduction in the number of vegetative buds for the cultivars with a basal shooting pattern. For the cultivars with higher DM yield arising from high stubble height, the DM was differently distributed into leaf and stem material. Before cuttings, the tiller number was reduced in the control plots but not in the plots with more lenient cutting height. Average tiller yield increased with increasing cutting height to reach a maximum at 20-cm cutting height. Average harvested culm length was constant for the four cutting heights. The variation in a number of plant traits, arising from cultivar diversity, can be altered by agronomic practices, thereby causing potentially contradictory results. Defining specific interactions of cultivar-by-cutting height treatments and analysing these with PCA proved to be a useful approach for visualising clusters from multiple measurements. The proposed approach for analysing data could serve as a model for other trials with similar interactions between cultivar diversity and agronomic treatments.
Introduction
Thailand smallholder farmers raise most of their cattle in a cut-and-carry system, where the grass is cut and the herbage is used for stall feeding. Owing to sparse rainfall and low temperatures, the climate is not suitable for pasture production in the dry season, see . Hence, farmers often lack sufficient quality feed during the dry season, with subsequent reduction in milk production. To avoid this recurrent reduction, farmers are advised to use rice straw and rice straw concentrate to keep the cows producing milk in the dry season, but this is very costly. A cheap alternative is to increase the pasture production in the rainy season and store it for later use (Boonman, Citation1993). In order to optimise production in the rainy season, correct cutting must be implemented. Biomass regeneration can be influenced by several factors such as the amount of residual photosynthetic tissue, carbohydrate and other reserves, the recovery rate of root growth, nutrient and water uptake, and the quantity and activity of remaining meristems (Harris, Citation1978). Following defoliation the level of carbohydrate reserves fluctuate, which affects re-growth (Sprague & Sullivan, Citation1950; Ward & Blaser, Citation1961). Grasses native to cool climates accumulate fructosan whereas those adapted to warm climates accumulate sugars and starch (De Cugnac, Citation1931). The level of carbohydrates in temperate grasses is higher (8–9% of DM) than that of tropical grasses (0.5–4% of DM) and shows less seasonal variation (Noble & Lowe, Citation1974). Intense cutting (0 cm) compared with lenient cutting (30 cm) of 4 Pennisetum species was shown in a study by Wadi et al. (Citation2004) to reduce the carbohydrate level in the subterranean stem and this reduced the persistence.
Table I. Actual and long-term mean rainfall and temperature at Suwanvajokkasikit Research Station, Packchong during this study.
Pasture grasses are excellent fodder crops that are cheap to establish and provide high yields if managed appropriately. Pasture grasses in Thailand commonly consist of grasses such as Guinea grass (Panicum maximum Jacq.), para grass (Brachiaria mutica (Forssk) Stapf.), ruzi grass (Brachiaria ruzziiensis R. Germ and C.M. Evrad), green panic (Panicum maximum Jacq. var. trichoglume Robyns), and Napier grass (Tudsri & Sawasdipanit, Citation1993), which mature very quickly with corresponding decline in digestibility, voluntary intake, and protein content (Humphreys, Citation1991). If the grass is cut frequently it will increase the leafiness, which subsequently will increase the voluntary intake and the protein content. However, frequent and intense cutting have proved to decrease yield and persistency of the Napier grass sward (Chaparro et al., Citation1995) especially during a dry season (J⊘rgensen et al., Citation1997). Lenient-frequent cutting has proved best in long-term production of high-quality feed of many tropical species, but the optimal management of pasture grasses differs between species (growth habit), location (soil), and year (climate). Therefore, further research on defoliation and re-growth could substantially increase the pasture production in challenging environments.
This research was conducted to examine the possible interactions between cultivars and cutting regimes in terms of effect on DM and quality of the forage.
Materials and methods
Experimental site
The experiment was conducted at Suwanvajokkasikit Research Station at Pakchong (150 km north-east of Bangkok, 14° 38′ N, 101° 19′ E, 388 m. a.s.l.). The soil was a sandy loam classified as a moderate reddish brown lateritic with a pH of 6.5. The chemical content of the soil in the top 0–15 cm was 65 ppm available P (Bray II), 125 ppm K, and 2.1% organic content. The experiment was conducted in a rain-fed environment with a long-term annual precipitation of 1091 mm where the period from November until the end of February is regarded as the dry period (). Maximum temperatures can reach 35 °C in April and minimum temperatures are as low as 15 °C in December and January. Weather data during the research period are also given in .
Experimental design
The experimental design was a split-plot with three replications from 1994 to 1995. In order to obtain information on the effect of closing date defined here as the date for the final cut before the dry season, the experiment was modified at the end of 1995 to a split-split-plot design. Main plots comprised five Napier grass cultivars, namely Common Napier, Merkeron, Dwarf Napier, Taiwan A25, and Tangashima. The sub-plot-factor consisted of four different cutting-height treatments; i.e., 0 (control), 10, 20, and 30 cm above ground level. The sub-sub-plot factor was two different closing dates, namely dry season closing on November 15th, 1995 and January 15th, 1996. In this analysis, no data on the effect of closing date (sub-sub-plot) were used. The Napier grasses were grown in rectangular plots of 3 m × 5 m. They were vegetatively propagated and the planting material consisted of stem pieces with three nodes. Planting took place on the 16th of November, 1993 in rows 50 cm apart with 50 cm spacing within rows (4 plants m−2).
Treatments and data collection
Cutting started on the 30th of March, 1994 and was carried out at monthly intervals throughout 1994, giving a total of 9 cuts. In 1995, the grass was cut three times. For data collection, one square, 1.0 × 1.0 m, was placed at random positions in each plot and cut with shears at given cutting heights. For the cut on August 21st, 1995 one sample was collected in each sub-plot. For the cuts on September 25th, 1995 and November 10th, 1995 two separate samples was collected in each sub-plot. The fresh material from the sampling area was subdivided into leaves and stems at all cuts. The material from each harvest was dried at 60 °C for 3 days and DM yield per unit area was then calculated for each sub-plot (1994–1995). After each sampling, all plots were cut to the given height according to their treatment and material removed. Besides DM a number of quantitative data were recorded given a total of 18 different dataset, also referred to as traits (). Basal fertilisers were applied at the rate of 300 kg ha−1 NPK (15-15-15) before the start of the experiment. Subsequently, 60 kg ha−1 N were applied every three months during 1994 and 300 kg ha−1 NPK (15-15-15) before each cut from 1995 to 1996; i.e. a total application of 720 kg N ha−1. For further experimental details see J⊘rgensen (Citation1996) and Tudsri et al. (Citation2002).
Table II. Quantitative and qualitative measurements used in the principal-component analysis.
Data analysis
When assessing the phenotypic and qualitative variation, the five cultivars and four cutting regimes were defined as 20 specific interactions. The period from June 1995 to November 1995 was chosen for further analysis, since significant effects of the main factors were found in this period without significant interactions between the same factors in the analysis of variance (Tudsri et al., Citation2002). A number of agronomic data were recorded, which, together with the quantitative data for the same period, were used in this study to investigate the interactions between genetic diversity and agronomic treatments. For defining clusters and assessing interactions between the cultivars and the cutting heights, PCA was undertaken using the 18 quantitative and qualitative measurements () on the 20 specific interactions averaged across blocks (). Derived data were removed before the analysis, e.g. total DM yield. The statistical analyses were carried out using the R software environment (R Development Core Team, Citation2005).
Table III. Specific interactions defined by five cultivars and four cutting heights.
Results
Total cumulative DM production
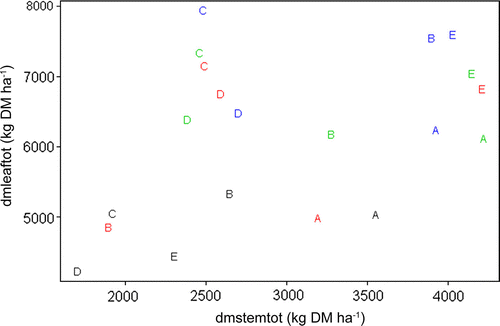
As a cumulative sum of the three cuttings included in this study the total DM production reached a maximum of 11 621 kg DM ha−1 for the cultivar Tangashima at 30-cm cutting height and a minimum of 5930 kg DM ha−1 for the cultivar Taiwan A25 at 0-cm cutting height (). All cultivars had reduced DM production with 0-cm cutting height compared with 30-cm cutting height. The reductions in DM production from the 0-cm cutting height were in the interval of 16–41% compared with the 30-cm cutting height. Ten-cm compared with 30-cm cutting height also negatively affected DM production for Common Napier and Merkeron and for unexplained reasons also gave lower DM production compared with 0-cm cutting height (). The maximum total DM yield from Tangashima (30 cm) compared with the DM production of Dwarf Napier (30 cm) originated from a higher stem yield (+ 1531 kg DM ha−1). The total leaf DM production was highest for Dwarf Napier cut at 30 cm with a maximum leaf DM production of 7927 kg DM ha−1 ().
Table IV. Cumulative DM production and leaf percentage from 20 specific interactions from 3 cutting dates (Aug–Nov) (Numeric code for specific interactions as defined in ).
Leaf percentage
The five cultivars clearly exhibited different mechanisms for incorporating increases in DM yield as a result of different cutting heights (), although there was no significant interaction between cutting height and cultivars for leaf percentage. Merkeron showed a smooth pattern, where increases in total DM yield are equally distributed into leaf and stem material, except for the 10-cm cutting height. Common Napier clustered into two clusters, suggesting that the optimal stubble height could be 20 cm for this cultivar. Dwarf Napier had a significantly higher leaf percentage than the other cultivars () and increases in total DM yield were mainly from increases in the leaf proportion (). Taiwan A25 had a slightly lower leaf percentage than Dwarf Napier and a stable leaf percentage across cutting heights. Tangashima clustered into two clusters, namely the control (0 cm) being separated from the cutting treatments suggesting that 10-cm cutting height is the absolute minimum for this cultivar for obtaining high DM yield.
Figure 1. Scatter plot of total dry matter yield (Kg DM ha −1) from leaf (dmleaftot) and stem (dmstemtot) proportion for five cultivars (A: Common Napier, B: Merkeron, C: Dwarf Napier, D: Taiwan A25, and E: Tangashima) at four cutting heights (Black: 0 cm, Red: 10 cm, Green: 20 cm, Blue: 30 cm).
The increase in harvested material from lenient cutting was from a larger average tiller yield, which cannot be associated with the increased plant height because the harvested culm length was almost equal for the four cutting heights (plant height minus cutting height = harvested culm length). The control plots (i.e., 0-cm cutting height) showed smaller average tiller yield although the tiller number was reduced (). The average tiller yield is a rough estimate of tiller size and the individual tiller yield is highly variable and depends on the tiller order as demonstrated by Ishii et al. (Citation2005).
Table V. Effect of cutting height on agronomic performance.
Tiller number
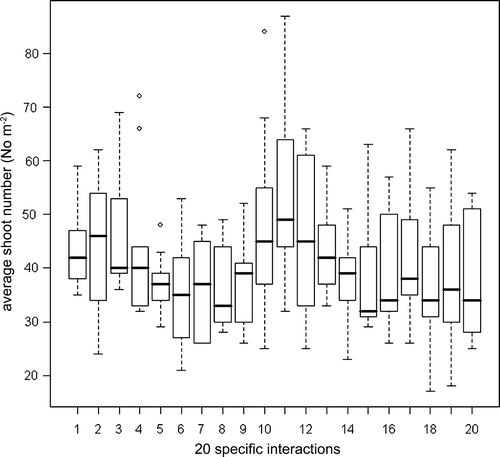
Tiller number measured before harvest did not differ between cultivars, while the tiller number was significantly negatively affected by severe cutting, i.e., in the control plots (). The tiller number also significantly changed among the cultivars in question during this study period, e.g. Dwarf Napier produced the minimum number of tillers prior to August cutting and maximum number of tillers prior to November cutting. All cultivars had the highest number of tillers prior to the dry season and the difference between the 3 cutting dates was significant ().
Table VI. Number of tillers per m2 measured prior to 3 cutting dates, mean of 4 cutting heights.
Plant height
Plant height differed significantly among cultivars and as an average Dwarf Napier was the shortest cultivar whereas Common Napier and Tangashima were the tallest cultivars in this study period (). There was a clear interaction between cultivars and cutting dates; i.e., cultivars changed growth pattern throughout the season. Common Napier had a relative low plant height in August but was the tallest cultivar throughout the following period (). The cutting intensity significantly affected the plant height, with progressively higher plants being obtained with lenient cuttings (higher stubble), see .
Table VII. Plant height measured from ground to top (cm) prior to 3 cutting dates, mean of 4 cutting heights.
Shooting pattern
The shooting pattern following defoliation was not significantly affected by cutting height for the five cultivars. There was a tendency for Dwarf Napier to produce more shoots with higher stubble (), which can be explained by aerial tillering for this cultivar. The other four cultivars had a more basal shooting and erect growth pattern and from visual observations there were few remaining leaves left in the stubble after cutting. Cutting date significantly affected the number of shoots, with an increase at the latest cutting date prior to the dry season, when the cultivars started flowering. At this cutting date, the effect was not equal among cultivars and cutting heights, because large variation exists between cultivars for the ability to enter the reproductive stage (Wouw et al., Citation1999). Dwarf Napier-the cultivar with the highest tendency for flowering-also produced the highest number of shoots with a severe reduction in the control plot.
Figure 2. Box plot of average shoot number (No m −2) of 20 specific interactions at three dates measured approximately one week after cutting Each interaction code as defined in Table III.
Specific leaf area
The specific leaf area (SLA) was unaffected by the cutting intensity (). The SLA of Dwarf Napier was significantly higher than those of the other cultivars. Dwarf Napier had thinner leaves, which is associated with higher feeding value (Duru et al., Citation2004).
Table VIII. Specific leaf area (cm2 g−1) of five Napier grass cultivar at four cutting heights.
Correlation matrix
Several significant correlations occurred between the different agronomic and quantitative measurements (). The number of emerging shoots counted one week after cutting is consistently negatively correlated with plant height prior to cutting, suggesting that high plants could affect the following shooting (−0.14*; −0.11, and −0.10 for the three cuttings, respectively), although only significant at the 5% level after the first cutting date in August 1995 when plants were tallest (). A positive correlation occurred between plant height and stem DM yield (0.62***; 0.70***, and 0.68*** for the three cuttings, respectively). The number of shoots emerging after defoliation correlated significantly with the tiller number prior to the following cutting date (0.27* and 0.28*). The plant height measured prior to the first cutting did not correlate with the subsequent measurements of plant height, while the four other measurements of plant height were highly significantly correlated.
Table IX. Correlation matrix between different agronomic recordings and quantitative measurements (abbreviations as defined in ).
Principal-component Analysis (PCA)
In the PCA bi-plot, using the first two principal components (PRIN 1 and PRIN 2) as X and Y axes, respectively, the cultivars Common Napier (central right) and Dwarf Napier (upper left) form two distinct clusters, while the remaining cultivars formed a 3rd cluster (lower left) without obvious structure (). Of the different agronomic treatments, the 0-cm cutting height showed a clear cluster (left), while the rest were more scattered with a tendency to sequentially reflect the cutting heights. The eigenvalue (calculated for the 10 first principal components) indicated almost equal weights for the 2nd and 3rd components (). PRIN1 and PRIN2 explained 54% of the variation and thus explain the variation of approximately 10 characters. The loadings of the different characters on PRIN1 are reasonably uniform with a tendency to load the most on height and DM measurements and are likely to represent those, while on PRIN2 the loadings are more variable ().
Figure 3. Scatter plot for the first two principal components using 18 quantitative and qualitative measurements on the 20 interactions defined by cultivars (A: Common Napier, B: Merkeron, C: Dwarf Napier, D: Taiwan A25, and E: Tangashima) and cutting heights (Black: 0 cm, Red 10 cm, Green: 20 cm, and Blue: 30 cm).
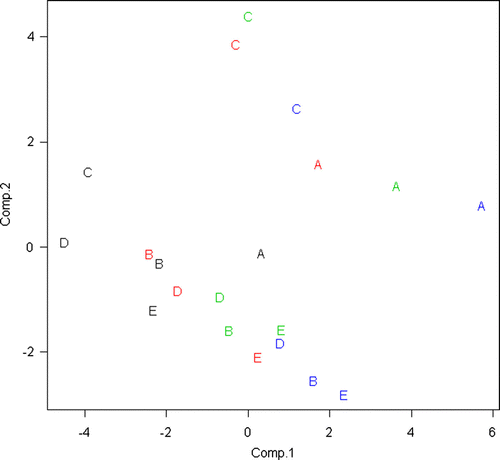
Figure 4. Eigen value of the 10 first principal components.
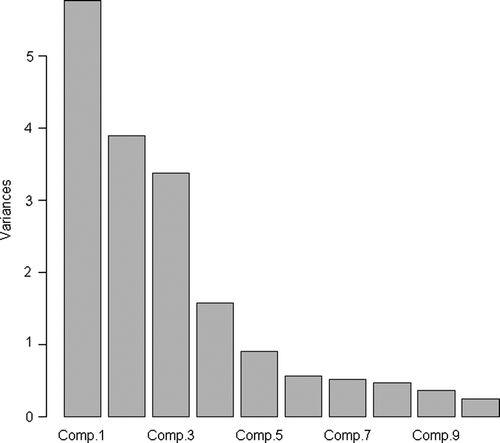
Table X. Loadings of 18 measurements on principal components 1 and 2.
Discussion
The total cumulative DM production was significantly different between the 5 cultivars and also between the 4 cutting heights and reached a maximum for Tangashima at 30-cm cutting height (11 621 kg DM ha−1) and a minimum for the cultivar Taiwan at 0-cm cutting height (5930 kg DM ha−1). The differences in DM production between cultivars were reduced with higher cutting height. Selecting the best cultivar and best management practise almost doubled the DM yield. The plant height measured before each cutting is highly positively correlated with stem DM production, and therefore also total DM production. This correlation is not useful for cultivar selection since the stem fraction has low digestibility (Ishii et al., Citation2005). For cultivar selection emphasis should be on high leaf DM production combined with high feeding quality, e.g. SLA, where Dwarf Napier has significant thinner leaves compared with the other cultivars in this study, not previously examined for different Napier grass cultivars. The SLA of Dwarf Napier was 212 cm2 g−1 as an average of the 4 cutting heights, which is markedly higher than that of Common Napier with an SLA of 162 cm2 g−1.
Defining specific interactions and analysing them with PCA proved to be a useful approach for visualising cultivar-by-cutting height interactions in Napier grass. From it is possible to compare the loading of genetic diversity (5 cultivars) with the loading of agronomic treatments (4 cutting heights) on a multiple set of measurements, defined here as traits. In this study the genetic diversity is separating the specific interactions more than the agronomic treatments. The cultivar × crop management interactions showed a clear clustering. Although, in this study, the clustering for some specific parameters could have been illustrated using more trivial methods, clustering of a larger dataset may not be easy to visualise without PCA. Analysis of variance on a specific parameter (total DM production of different cultivars), i.e. two dimensions, is unable to establish general patterns in large datasets with multiple dimensions. In this study, Dwarf Napier and Common Napier belong to different clusters, which is for a large part explained by the clear difference in plant height and leaf:stem ratio between the 2 cultivars. Plant height has, in another study by Wouw et al. (Citation1999), proved to be a good morphological character to distinguish accessions of Napier grass. The other cultivars (Merkeron, Taiwan A25, and Tangashima) clustered without obvious structure. There was also a clear clustering of the control (0-cm cutting height) across all cultivars, while the other treatments were grouping according to cutting height with some differences between cultivars. The PCA analysis could have been further improved with more data on quality parameters. For instance, the feeding quality is often negatively correlated with yield parameters. The PCA analysis could also be further expanded if more morphological data had been collected. The proposed approach for analysing data could serve as a model for other trials with similar interactions between cultivar diversity and agronomic treatments (Genotype-by-Management interactions) and thus assist the forage scientist in selecting the cultivar-management practice that is best adapted to the specific environment in question.
The effect of cutting height on total DM yield in Napier grass solely occurs after the first season; i.e., when vigour is reduced (Tudsri et al., Citation2002), which is in line with the study of Chaparro et al. (Citation1995). The beneficial effects of lenient cutting are related to the greater size and levels of residual plant constituents (Sprague & Sullivan, Citation1950; Ward & Blaser, Citation1961; Harris, Citation1978). It was expected that lenient cutting would have given a higher leaf percentage, since a major part of the stem was left in the ground at harvest (approximately 20% with a plant height of 150 cm and 30-cm cutting height). The lowest part of the stem was supposed to be the thickest; i.e., adding further to the proportion of stem material left in the ground. However, Descamps & de Brito (Citation2001) did not find significant variation in stem width in the median region compared with the basal region. Chaparro et al. (Citation1995) found higher leaf percentage with 46-cm cutting height compared with 10-cm cutting height, although cutting frequency affected leaf percentage more than did cutting height. In this study the lack of difference in leaf percentage between the 4 cutting heights can be explained by the short frequency between cuttings (4 weeks). In this study, the positive effects of lenient cutting on DM yield ensued from a combination of more and heavier tillers measured as average tiller weight. Should the plant height be deducted from the cutting height, the four cutting treatments have the same harvested culm length. Hence, the positive effect on total DM yield by changing from 10-cm to 30-cm cutting height could only be explained by thicker harvested tillers with possibly more and larger leaves attached to each.
In this experiment, there was no significant effect of cutting height on the number of shoots (growing points). Holt & McDaniel (Citation1963) showed that species with a basal shooting pattern were less affected than were species with aerial shoots. Dwarf Napier was the only cultivar with a high number of aerial growing points and also the only cultivar to have a tendency to higher shoot number with higher stubble. In the other cultivars, the residual leaf area or stored carbohydrates may be responsible for the positive effect of high stubble. For the erect cultivars, carbohydrate storage was presumably more important than were residual leaves since no leaves were left after defoliation. The residual leaves have little photosynthetic efficiency because of previous shading (Langer & Keoghan, Citation1970; Ludlow & Wilson, Citation1971).
For tropical pasture production the cultivar Dwarf Napier seems to be a relevant choice in cultivar selection. The DM production of Dwarf Napier is high although plant height is comparably low, which can be explained by a high number of relatively small tillers combined with a high portion of thinner leaf material compared with the other cultivars in this field experiment. Small tillers, high leaf percentage, and thin leaf blades are important parameters for increasing the digestibility of grass fodder (Duru et al., Citation2004; Ishii et al., Citation2005). The crop management should aim at maximising DM production and keeping a high feeding value without destroying the persistence of the grass sward, which in this experiment is achieved with a 30-cm cutting height with a constant 4-week cutting frequency.
Future work on Napier grass should include further breeding efforts, since the plant's diversity is assumed not to be fully exploited, as large diversity has been found in a study with 10 lines (Zewdu, Citation2005). For pasture production a combination of high DM yield should be combined with a high feeding value. Napier grass could also become important in the BIOHEAT industry as described by Samson et al (Citation2005), where the management and cultivar choice should focus mainly on maximising the DM production.
Acknowledgements
We thank the staff of Suwanvajokkasikit Research Station for their assistance. This experiment was supported by the National Research Council of Thailand and Kasetsart University Research and Development Institute. A grant from the Pedersholm Foundation was given to the first author for leading the writing of this manuscript. We also thank Dr Sean Mayes, Mr Florian Stadler, and several anonymous reviewers for critical comments on the manuscript.
References
- Boonman , J.G. 1993 . East Africa's Grasses and Fodders: Their Ecology and Husbandry , Boston : Kluwer Academic Publishers .
- Chaparro , C.J. , Sollenberger , L.E. , & Jones , C.S. Jr. 1995 . Defoliation effects on “Mott” elephantgrass productivity and leaf percentage . Agronomy Journal , 87 , 981 985 .
- De Cugnac , A. 1931 . Recherches sur les Glucides des Graminees . Annales des Sciences Naturelles de Botaniques , 13 : 1 – 129 . (In French)
- Deschamps , F.C. , & de Brito , C.J.F.A. 2001 . Forage quality and relative participation in dry matter of different fractions of elephant grass cultivars (Pennisetum purpureum Schumach.) . Revista Brasileira de Zootecnia , 30 , 1418 1423 (in Portuguese) .
- Duru , M. , Cruz , P. and Magda , D. 2004 . Using plant traits to compare sward structure and composition of grass species across environmental gradients . Applied Vegetation Science , 7 : 11 – 18 .
- Harris , W. 1978 . “ Defoliation as a determinant of the growth, persistence and composition of pasture ” . In Plant Relations in Pastures , Edited by: Wilson , J.R. 67 – 85 . Melbourne, , Australia : CSIRO .
- Holt , E.C. and McDaniel , J.C. 1963 . Influence of clipping on yield regrowth and root development in grass, Paspalum dilatatum Poir, and Kleingrass, Paspalum coloratum L . Agronomy Journal , 55 : 561 – 564 .
- Humphreys , L.R. 1991 . Tropical Pasture Utilisation , Cambridge, , UK : Cambridge University Press .
- Ishii , Y. , Yamagushi , N. and Idota , S. 2005 . Dry matter production and in vitro dry matter digestibility of tillers among napiergrass (Pennesetum purpureum Schumach) varieties . Grassland Science , 51 : 153 – 163 .
- J⊘rgensen , S.T. 1996 . Effects of Cutting Height in Yield and Quality of Five Napier Cultivars . MSc Thesis, The Royal Veterinary and Agricultural University, Copenhagen, Denmark .
- J⊘rgensen , S.T. , Tudsri , S. , Pookpakdi , A. , & Stolen , O. 1997 . Effects of cutting height in yield and quality of five Napier cultivars , In : Proceedings of the 18th International Grassland Congress, Session 22: 5–6 . Saskatoon , Canada .
- Langer , R.H.M. and Keoghan , J.M. 1970 . Growth of lucerne following defoliation . Proceedings of the New Zealand Grassland Association , 32 : 98 – 107 .
- Ludlow , M.M. and Wilson , G.L. 1971 . Photosynthesis of tropical pasture plants. 2. Temperature and illuminance history . Australian Journal of Biological Sciences , 24 : 1065 – 1075 .
- Noble , A. and Lowe , K.F. 1974 . Alcohol-soluble carbohydrates in various tropical and temperate pasture species . Tropical Grasslands , 8 : 179 – 187 .
- R Development Core Team 2005 . R: A language and environment for statistical computing . R Foundation for Statistical Computing, Vienna, Austria, ISBN 3-900051-07-0 . URL: http://www.R-project.org (accessed on 30 March 2009) .
- Samson , S. , Mani , S. , Boddey , R. , Sokhansanj , S. , Quesada , D. , Urquiaga , S. , Reis , V. and Lem , C.H. 2005 . The Potential of C4 Perenial Grasses for Developing a Global BIOHEAT Industry . Critical Reviews in Plant Sciences , 24 : 461 – 495 .
- Sprague , V.G. and Sullivan , J.T. 1950 . Reserve Carbohydrates in Orchardgrass Clipped Periodically . Plant Physiology , 25 : 92 – 102 .
- Tudsri , S. , & Sawasdipanit , S. 1993 . Managerial approach to pasture production in Thailand . In : C.P. Chen & C. Satjipanon Strategies for forage-based livestock production in southeast asia 111 125 . Proceedings of the 3rd meeting of RWG on Grazing and Feed Resources of Southeast Asia, 31 January–6 February 1993 , Khoan Kaen , Thailand .
- Tudsri , S. , J⊘rgensen , S.T. , Riddach , P. and Pookpakdi , A. 2002 . Effect of cutting height and dry season closing date on yield and quality of five Napier grass cultivars in Thailand . Tropical Grasslands , 36 : 248 – 252 .
- Wadi , A. , Ishii , Y. and Idota , S. 2004 . Effect of Cutting Interval and Cutting Height on Dry Matter Yield and Overwintering Ability at the Established Year in Pennisetum Species . Plant Production Science , 7 : 88 – 96 .
- Ward , C.Y. and Blaser , R.E. 1961 . Carbohydrate food reserves and leaf area in regrowth of orchardgrass . Crop Science , 1 : 366 – 369 .
- Wouw , M. van de Hanson , J. , & Luethi , S. 1999 . Morphological and agronomic characterisation of a collection of napier grass (Pennisetum purpureum) and P. purpureum x P. glaucum . Tropical Grasslands , 33 , 150 158 .
- Zewdu , T. 2005 . Variation in growth, yield, chemical composition and in vitro dry matter digestibility of Napier grass accessions (Pennisetum purpureum) . Tropical Science , 45 : 67 – 73 .