Abstract
Carbon dioxide flux from the soil to the atmosphere is an important component of terrestrial C cycling, and accurate estimates of CO2-C fluxes are crucial for estimating C budgets. A field study was conducted (i) to examine the diurnal and seasonal soil CO2 flux pattern in spring maize fields on the Loess Plateau, and (ii) to determine the effects of soil characteristics affected by various cultivation practices on CO2 flux from the soil surface to the atmosphere. Soil surface CO2 flux was determined with an LI-8100 Automated Soil Flux System, and related environmental factors were also measured, including near-ground air temperature and relative humidity, soil moisture (0–15 cm), soil temperature (at depths of 5, 10, 15, and 20 cm), and leaf area index. Diurnal soil CO2 flux showed a single peak between 12-00 h and 16-00 h, and reached a minimum in the early morning, at about 4-00 h. During the crop's growing season, soil CO2 flux increased during the rapid vegetative growth stages, reached its maximum during the peak reproductive stages, and then declined as the plants senesced. Time series analysis showed that the temporal dynamics of the CO2 flux were more closely related to air temperature than to soil temperature; this may be because a substantial portion of the CO2 originated from surface residues. The time-averaged mean soil CO2 flux for different cultivation practices over the growing season was ranked as follows: plastic film mulching (3.980 µmol m−2s−1) > corn straw mulching (3.464 µmol m−2s−1) > supplementary irrigating (3.157 µmol m−2s−1) > rain-fed (2.371 µmol m−2s−1) > bare ground (1.934 µmol m−2s−1). Different cultivation practices affected plant and microbial activities, and soil hydrothermal conditions, and caused different patterns of soil surface CO2 flux in spring maize fields on the Loess Plateau.
Introduction
Increases in the emission of greenhouse gases such as carbon dioxide (CO2), methane (CH4), and nitrous oxide (N2O) from the soil surface to the atmosphere have been a worldwide concern for several decades. CO2 is recognized as a significant contributor to global warming and climatic change, accounting for 60% of global warming or total greenhouse effect (Rastogi et al., Citation2002). Measuring the soil CO2 flux is crucial for accurately evaluating the effects of soil-management practices on global warming and carbon cycling.
There is double the amount of C in the soil as in the atmosphere (Mielnick & Dugas, 1999; Maier & Kress, Citation2000), so it can be considered a major source and sink for atmospheric CO2 (Jensen et al., Citation1996). Soil CO2 flux or ‘soil respiration’ is one of the most important components of any ecosystem's C budget, which involves organic matter decomposition and mineralization, root respiration, and rhizosphere or faunal respiration (Carlisle et al., Citation2006).
Several factors are known to influence CO2 production and emissions rates from the soil surface. These factors include soil temperature, moisture content, mulching and irrigation practices, tillage systems and practice, the presence of organic matter and nutrients, soil aeration, microbial processes, and soil diffusivity (Edwards, Citation1975; Kern & Johnson, Citation1993 ; Ball et al., Citation1999; Mielnick & Dugas, 1999). An increase in soil temperature generally accelerates organic-matter decomposition, oxidation, microbial and root activity, and C mineralization processes. Consequently, this acceleration increases CO2 emission from the soil, which in turn reduces C storage. Both root and microbial sources of CO2 exhibit an exponential increase in activity as a function of temperature (Boone et al., Citation1998; Davidson et al., Citation1998). Seasonal changes in CO2 flux have been reported to follow seasonal temperature trends (Anderson, Citation1973; Rochette et al., Citation1991). On a shorter time scale, diurnal changes in soil CO2 flux have generally been observed to be related to soil temperature (Xu & Qi, Citation2001; Parkin & Kaspar, Citation2003; Yang & Cai, Citation2004; Han et al., Citation2007), with maximum CO2 fluxes occurring mid-afternoon, and minimum fluxes occurring in the early morning.
Soil moisture content affects soil respiration: higher moisture content (in the absence of waterlogging) often provides better conditions for microbial activity, increasing microbial oxygen consumption and CO2 production and emission from the soil (Buyanowski & Wagner, Citation1983). The relationship between soil CO2 emission and soil moisture content is not yet clearly understood. However, the effect of soil moisture content has been described by linear, quadratic, logarithmic, and parabolic equations (Davidson et al., Citation2000). Several studies have demonstrated that moisture content is an important factor in soil respiration (Edwards, Citation1975; Davidson et al., Citation1998, Citation2000; Han et al., Citation2007). Davidson et al. (Citation2000) showed that rates of soil respiration increased with increasing water content in both pasture and forest soils. They developed an empirical regression equation to describe soil respiration as a function of soil matric potential and concluded that CO2 flux increased with decreasing soil matric potential. Bottner (Citation1985) and Kieft et al. (Citation1987) suggested that high soil respiration rates may be a function of wetting-up effects, which have been shown to produce pulses of microbial activity and CO2 production.
Agricultural management practices can change the characteristics of the soil surface, and hence influence the hydrothermal properties of the soil. For example, mulching can affect soil temperature and moisture content (Li et al., Citation1999; Ghosh et al., Citation2006), which can directly affect CO2 fluxes from the soil surface (Edwards, Citation1975; Davidson et al., Citation1998; Parkin & Kaspar, Citation2003; Amos et al., Citation2005; Han et al., Citation2007). Straw mulching (SM) systems can conserve soil water and reduce temperature because of decreased soil disturbance and increased residue accumulation at the soil surface (Lentz & Bjorneberg, Citation2003; Al-Kaisi & Yin, Citation2005). Soil mulching with plastic film, which results in reduced water loss and more evenly regulated soil temperatures, has been widely used in agriculture (Li et al., Citation2004a). Tiquia et al. (Citation2002) and Li et al. (Citation2004b) found that the CO2 concentration in the soil under mulch was much higher than in nonmulched areas (Phene & Sanders, Citation1976).
According to the literature, most studies examining the factors affecting soil CO2 flux have focused on a single time scale and few, if any, studies have considered various time scales. Most studies have concentrated on the study of soil CO2 flux in farmland exposed to only one cultivation practice; few studies have focused on farmlands with a variety of cultivation practices. The objectives of this study were: (i) to examine the diurnal and seasonal soil CO2 flux pattern in spring maize fields on the Loess Plateau; and (ii) to determine the effects of soil characteristics associated with different agricultural practices on CO2 flux from the soil surface.
Materials and methods
Site description
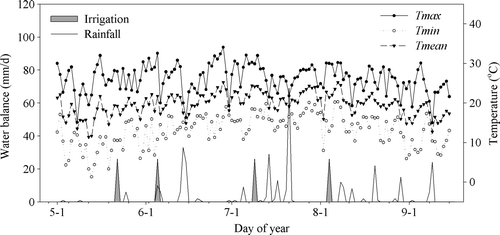
The Changwu experimental station (35.2°N, 107.8°E) is at an elevation of about 1206.5 m above sea level, and is located on loessial tableland. The loess on the tableland is more than 100 m thick. The soils are Cumuli-Ustic Isohumisols, according to the Chinese Soil Taxonomy (Gong et al., Citation1999), which contained 37% clay, 59% silt, and 4% sand with a pH-value of 8.4 and bulk density of 1.3 g cm−3. The organic matter, total nitrogen, available phosphorus, available potassium, and inorganic nitrogen were 11.8 g kg−1, 0.87 g kg−1, 14.4 mg kg−1, 144.6 mg kg−1, and 3.15 mg kg−1, respectively. The average annual precipitation is 578 mm, with 55% falling between July and September. The annual average temperature is 9.2 C; the annual maximum and minimum temperatures are 38.4 and −19.9 C, respectively. The common regional cropping system is a three-year rotation of winter wheat–winter wheat–spring maize. Rain-fed agriculture is the dominant production system. The temporal patterns of temperature, rainfall, and amount of irrigation during the maize growing season are presented in .
Figure 1. Main meteorological data and irrigation application during the maize-growing season in 2008.
Experimental design
Four field water management practices – supplementary irrigation (SI), rain-fed (RF) cultivation, corn straw mulching (SM), and plastic film mulching (FM) – were adopted in spring maize fields. The soil water supply for RF, SM, and FM cultivation relied on natural rainfall, while for the SI treatment sufficient moisture in the soil [70~85% of the field water capacity (FWC)] was maintained with tap water delivered by furrow irrigation. Soil water content of the 0–15-cm depth was determined using a time domain reflectometer (TDR) probe every 2–3 days. In the SI treatment, furrow irrigation was carried out when the soil water content fell below the critical value of 70% FWC. Over the growing season, the maize was irrigated four times (May 22, June 5, July 7, and August 4 in 2008). On each occasion, soil water to a depth of 25.6 mm was provided; this was equivalent to the difference between the lower and upper FWC limits. The four treatments were arranged in a completely randomized block design, with three replicates for each treatment. Bare ground (BG), with the same field-management regime as the rain-fed treatment but without maize, acted as the control. The size of each experimental plot was 50.7 m2 (7.8 m×6.5 m).
Ridge cultivation, a common maize-cultivation practice across the Loess Plateau, was adopted in all four of the treatments. First, 110 kg N ha−1 in the form of urea (N 46%), 50 kg P ha−1 in the form of calcium superphosphate (P2O5 12%), and 100 kg K ha−1 in the form of potassium sulfate (K2O 45%) were broadcast over the soil surface, which was then turned over to subsurface by plowing. Ridges were constructed by banking up soil from two sides to a height of 0.1 m from the base, producing a ridge 0.45 m wide at the top and leaving furrows 0.15 m wide at the base. In the plots mulched with plastic, film (0.7 m wide and 0.005 mm thick) was used to cover the soil surface of the ridges but not the furrows; the edges were secured under the soil in the bottom of the furrows. On April 18 2008, spring maize (Zea mays L. pioneer 335) was sown in 5-cm-deep holes spaced 0.2 m apart, using a human-powered hole-drilling machine, along the midline on the top of the ridge. Before backfilling, water was added as required, to encourage seedling emergence. On May 8, around the time of seedling emergence, corn straw cut into 0.1-m-long segments, was applied at a rate of 6000 kg ha−1 uniformly on both ridges and furrows in the SM plots. Additional nitrogen, in the form of urea, was applied at the jointing and tasselling stages, at rates of 80 kg N ha−1 and 90 kg N ha−1, respectively, following a nutrient-management plan aimed to achieve a final yield of 14 t ha−1. Maize cobs were harvested gradually, according to their ripeness, from 8 to 20 September. Weeds were controlled manually, as required, during the growing season.
Determination of soil CO2 flux and related environmental factors
Soil surface CO2 flux was determined with an LI-8100 Automated Soil Flux System (LI-COR Inc.) over a 20.3-cm diameter collar installed on the soil surface at each survey site. A 20-cm (LI-8100-103) respiration survey chamber was used, in which the increase in CO2 concentration was recorded at 1 s intervals by the LI-8100's infrared gas analyser unit. This chamber has a carefully designed pressure vent to prevent pressure gradients and wind incursion from outside the chamber or air flow out of the chamber (Bain et al., Citation2005). A pressure/vacuum air-flow system expands and contracts bellows to raise and lower the chamber above the soil collar automatically, thus minimizing perturbations of the microclimate inside the chamber during the measurements. Soil collars were constructed by cutting 11.4-cm lengths of thin-walled poly(vinyl chloride) (PVC) pipe (inner diameter 20.3 cm, outer diameter 21.34 cm).
During the experiment, soil-surface CO2 efflux measurements were made at two locations in each plot at 10–15-day intervals during the crop's growing season. Thus, there were six replicate soil-surface CO2 efflux measurements for each treatment. Survey sites were located in the space between two maize seedlings in a row, and the two sites in each plot were located in different rows, 2–3 m apart. The soil collars were inserted to a depth of approximately 7 cm, extending 4.4 cm above the soil surface to allow placing of the chamber; this was done 24 h before the measurements were taken, to allow equilibrium to be reached, in order to eliminate any effects of disturbance on soil CO2 respiration. Collar insertion cuts through the litter and severs superficial soil root networks and may adversely affect root respiration (Wang et al. Citation2005), although it does minimize soil-surface disturbance and it reduces sudden flushing of CO2 from the soil and litter (Jabro et al., Citation2008). Despite these considerations, this method was deemed to be appropriate in this situation. During installation in the FM plots, the plastic film inside the collars was removed; in the SM plots, the straws were kept in situ. Meanwhile, living plants and other residue inside the collar were also pulled up by hand with their roots to eliminate aboveground plant respiration. After each measurement session, collars were collected for subsequent use. Measurements were generally conducted between 9-00 h and 11-00 h (local time) on clear days or at least two days after rains, throughout the experiment. The time span between chamber contact with the soil and the start of measurements (the deadband) was 20 s; this has previously been determined to be sufficient for pressure equilibration. The measurement time was set to 120 s in order to avoid important chamber CO2 concentration changes (Davidson et al., Citation2002).
During the soil CO2 flux measurements, air temperature and relative humidity inside the chamber were also recorded by the LI-8100 analyser unit. The leaf areas of three plants per plot were measured after determining the soil CO2 flux. Leaf area = leaf length×maximum width×k, where k is a shape factor, set to 0.75. The values of leaf area index (LAI) were then calculated on the basis of plant density (85 000 plants ha−1), i.e., LAI = Leaf area (m2 plant−1)×85 000 (plants ha−1)/10 000 (m2 ha−1). Soil moisture content (SWC) in the top 15 cm of the soil was measured volumetrically with a TDR probe at five locations in each plot at 2–3-day intervals. A set of mercury thermometers was installed side by side on the ridge at 5-cm depth increments, down to 20 cm in the soil, in each plot. The soil temperature was recorded daily at 8-00 h and 14-00 h, the coolest and warmest times of the day; the values were averaged to give the mean daytime temperature.
In order to examine the diurnal soil CO2 flux pattern in spring maize, soil CO2 flux, as well as temperature and moisture, were also measured simultaneously at 2-h intervals for 24 hours during August 23 to 24, at the time of the crop's late filling stage. Measurements were taken at 6-00 h, 8-00 h, 10-00 h, 12-00 h, 14-00 h, 16-00 h, 18-00 h, 20-00 h, 22-00 h, 0-00 h (the next day), 2-00 h, and 4-00 h. During this 24-hour period, the sky was clear and cloudless.
Results
Diurnal patterns of soil CO2 and related environmental factors in maize fields
For all four cultivation practices in spring maize fields on the Loess Plateau, the soil surface CO2 flux over the course of a typical experimental day showed a clear pattern, increasing during the morning, reaching a maximum around noon and decreasing thereafter (). The similarity between the diurnal pattern of soil respiration and the soil and air temperatures and moisture (the latter exhibiting a negative relationship) inside the LI-8100 chamber suggests that the diurnal variation in soil surface CO2 efflux was controlled mostly by soil temperature and near-ground air temperature and relative humidity. Although the patterns were similar, the time of day at which the maximum soil CO2 flux was recorded was different for the different cultivation practices. In BG, the soil CO2 flux reached its peak the earliest of all the treatments, between 10-00 h and 12-00 h; in SI, RF, and FM the maximum was reached around 14-00 h, and in SM between 14-00 h and 16-00 h. Thus there was a four-hour lag between the times of the earliest and latest maxima. Hence, crop-cultivation practices could postpone the soil CO2 flux peak significantly as a result of impacts on the microclimatic conditions, i.e. soil temperature, air temperature, and relative humidity near the soil surface and beneath the maize canopy. This was the result of shading as well as soil management (in the SM plots, for example). The minimum soil CO2 flux was recorded at 4-00 h, corresponding to the lowest soil temperature reading for all treatments. Mean soil CO2 flux, calculated by averaging diurnal flux observations in the same treatment, was only 1.87 µmol m−2 s−1 in BG, 4.22 µmol m−2 s−1 in FM, 4.19 µmol m−2 s−1 in SI, 3.95 µmol m−2 s−1 in SM, and 2.15 µmol m−2 s−1 in RF. Analysis of variance showed that soil CO2 fluxes in the BG and RF treatments were significantly (P<0.05) lower than in FM, SI, and SM, and there was no significant difference between BG and RF, and between FM, SI, and SM ().
Figure 2. Diurnal patterns of soil CO2 and related environmental factors in maize fields. (SI, RF, FM, SM, BG refer to supplementary irrigation, rain-fed, plastic film mulching, corn straw mulching, and bare ground, respectively.)
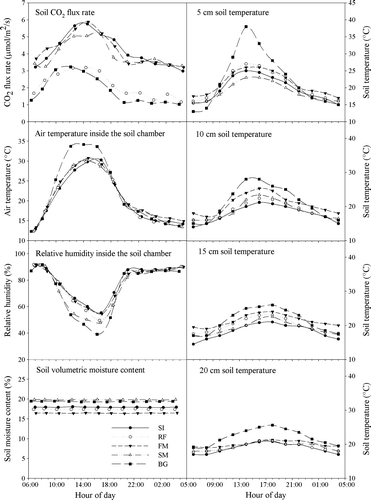
Air temperature and relative humidity inside the soil respiration chamber, representing the atmospheric conditions near the ground, exhibited similar, but opposite, diurnal patterns. The maximum temperature and minimum humidity occurred around 12-00~16-00 h, whilst the minimum temperature and maximum humidity occurred at 4-00 h, for all the treatments. With increasing depth, the diurnal soil temperature range narrowed, and the peak tended to occur later. However, a rather similar diurnal pattern for temperature occurred at different depths in the soil. In contrast to other environmental factors, there was no detectable diurnal variation in the soil moisture content. Therefore, soil temperature, near-ground air temperature, and relative humidity were the main factors regulating the diurnal variations in soil CO2 flux. Soil moisture content was not the critical factor affecting the variations in soil CO2 flux since it changed little over a single day, and it was generally sufficient to meet the requirements for microbial activity.
Seasonal variations of soil CO2 flux in spring maize fields
The soil CO2 flux in maize fields exhibited similar seasonal patterns for the four cultivation practices, closely mirroring crop growth (). Soil respiration was rather low at the very beginning of the season (May 14 – seedling stage), and then rose gradually as the crop developed, reaching a maximum around July 12 (maize tasselling and silking stage); it then declined gradually until the end of the season. Soil and air temperatures inside the respiratory chamber exhibited the same seasonal pattern, while the air relative humidity exhibited the inverse pattern.
Figure 3. Seasonal variations in soil CO2 flux and related environmental factors in maize fields. (SI, RF, FM, SM, BG refer to supplementary irrigation, rain-fed, plastic film mulching, corn straw mulching, and bare ground, respectively.)
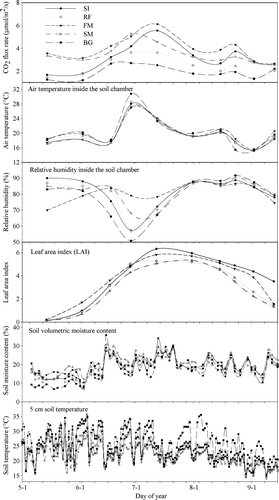
shows that the leaf area index (LAI) increased sharply from the jointing (June 2) to the tasselling (July 12) stage, and reached its maximum around the tasselling and silking stages (July 12); it then decreased dramatically during the ripening stage. The LAI pattern was in agreement with the variations in soil CO2 flux during the crop's growing season, indicating that soil CO2 exchanges were primarily regulated by seasonal crop growth activities. The amplitude of seasonal fluctuations in soil moisture content in the upper soil profile was quite large during the study period. Soil moisture was generally low at the crop seedling stage because high soil evaporation removed water from the upper soil profile; moisture levels increased after the jointing stage due to precipitation, and enhanced canopy shading, as well as root hydraulic lift. The general seasonal pattern of soil temperature was similar to that of atmospheric temperature, but it was also influenced by weather and maize canopy development.
Effects of cultivation practices on soil CO2 flux and environmental factors
Mean soil CO2 fluxes and means for the related environmental variables were calculated by averaging the corresponding observations over the growing season for the different cultivation practices, in order to evaluate their effects on CO2 flux in maize fields (). In the present study, soil CO2 fluxes were much higher when mulched – FM (3.980 µmol m−2 s−1) or SM (3.464 µmol m−2 s−1) – than when no mulch had been applied – SI (3.157 µmol m−2 s−1), RF (2.371 µmol m−2 s−1), and BG (1.934 µmol m−2 s−1). Active crop growth, increased soil respiration, and plant root activity may explain this, in part.
Table I. Mean soil CO2 fluxes and related environmental factors during the maize-growing season.
Mulching greatly increased soil respiration and affected the measured environmental factors in different ways. FM increased the topsoil temperature as well as the near-ground air temperature, and the improved soil conditions enhanced plant root activity as well as microbial respiration. However, it should be noted that CO2 efflux might be over-estimated by the recording method used here. Over-accumulated CO2 (high CO2 density in the soil atmosphere because of the plastic mulch preventing its escape) might have led to enhanced CO2 diffusion from the soil after the removal of film for measurement, despite the overnight equilibration; in addition, over-accumulated CO2 could restrict respiration under a plastic mulch. Unfortunately, such impacts could not be distinguished in the experiment. As regards SM, although the soil temperature decreased, high CO2 efflux was still observed; this is probably due to the enhanced soil fauna, organic catabolism of the straw, and relatively high soil water content. Soil respiration in the RF treatment was quite low due to low soil humidity, compared with the other treatments; this suggests that water deficit may have occurred during the crop's growing season, which could have inhibited microbial activity as well as crop growth. For SI, irrigation increased soil moisture content. Therefore, the enhanced soil respiration may be because the relatively high soil water content stimulated root growth and promoted soil organic carbon mineralization.
Environmental factors influencing surface CO2 flux in spring maize on the Loess Plateau
Correlations between the different environment factors and soil CO2 flux recorded over a 24-hour period indicated that diurnal soil respiration was significantly (P<0.01) correlated with air temperature and relative humidity inside the chamber/near the ground in all treatments (). Soil temperature at a depth of 5 cm was also significantly (P<0.01) correlated with soil CO2 flux for all treatments; soil temperature at depths below 5 cm was less well correlated with soil CO2 flux. This is probably because root respiration and microbial activities, which are linked to temperature in the uppermost topsoil (about 5 cm deep), account for a large proportion of the total CO2 production and emission from the soil. However, soil moisture content was weakly correlated with the diurnal variation in soil CO2 flux, because the tiny fluctuations over the course of a day would have little effect on microbial activity.
Table II. Correlations between soil CO2 flux and environmental factors based on diurnal fluctuations.
Correlations between the different environmental factors and soil CO2 flux recorded over the crop's growing season in each treatment indicated that seasonal soil respiration was correlated with air temperature and relative humidity inside the chamber/near the ground as well as with LAI, although the correlation coefficients varied according to cultivation practices (). In our study, air temperature in the respiration chamber exhibited the best correlation with CO2 flux, and the correlations were significant (P<0.05) for all treatments except BG. In addition, there were significant (P<0.05) correlations between LAI and soil CO2 flux rate for all treatments except SM. However, there were rather weak correlations between internal chamber air humidity (r 2=0.0904) or topsoil moisture content (r 2= − 0.0320) and seasonal CO2 flux for the FM treatment. This was largely because film mulching induced soil-moisture effects on CO2 flux by blocking the diffusion of CO2 and water vapor from soil pores to the atmosphere. In addition, correlation coefficients of soil temperature (5 cm) with CO2 flux were generally low in our study.
Table III. Correlations between soil CO2 flux and environmental factors based on seasonal variations.
The correlation between soil moisture in the topsoil and soil CO2 flux was significant (r 2=0.7986, P<0.001) in the BG treatment, and this correlation was stronger than those for other treatments. Relatively low correlation coefficients in the RF (r 2=0.5972) and SI (r 2=0.4765) treatments indicated that crop-growth effects (root respiration, induced microbial activity, and so on) might have overshadowed the impact of soil moisture. Furthermore, additional effects such as blocked diffusion of CO2 and water vapor from soil pores to the atmosphere by mulching and increased soil organic matter content derived from the straw mulch might have caused weak correlations between soil moisture and soil CO2 flux in the FM (r 2= − 0.0320) and in SM (r 2=0.1661) treatments.
Discussion
Temporal scales have often been neglected in studies aimed at identifying the environmental factors that control soil surface CO2 efflux (Anderson, Citation1973; Rochette et al., Citation1991; Xu & Qi, Citation2001; Parkin & Kaspar, Citation2003; Yang & Cai, Citation2004; Han et al., Citation2007). This study has demonstrated that, in spring maize fields exposed to different cultivation practices, environmental factors regulating diurnal fluctuations in CO2 flux are quite different from those governing seasonal variations. Diurnal soil CO2 flux showed a single peak between 12-00 h and 16-00 h, and was lowest early in the morning, at about 4-00 h; this is in agreement with patterns recorded in forests (Xu & Qi, Citation2001), grassland (Cao et al., Citation2004), and farmland (Han et al., Citation2007). Furthermore, correlation analysis revealed that air temperature and relative humidity inside the chamber and soil temperature explained most of the diurnal fluctuations in CO2 flux, while other commonly cited environmental factors, such as soil moisture content, plant biomass, and soil properties, had little effect on CO2 emission from the soil surface because their diurnal variations were too small. Hence, we consider that soil temperature exerted the main control over diurnal patterns of soil CO2 flux.
Seasonal variations in soil CO2 flux have been observed in almost all ecosystems. Soil respiration rates are usually highest during summer and lowest in winter. The seasonal variation is driven largely by changes in temperature, moisture, and photosynthate production (Epron et al., Citation2001; Xu & Qi, Citation2001; Borken et al., Citation2002). The main factors controlling seasonal variations in soil respiration may depend on the type of ecosystem and the climate. In farmland ecosystems, seasonal soil respiration was greatly influenced by crop growth, which affects soil microclimate and structure, the quantity and quality of detritus supplied to the soil, and direct root respiration: there were interactions between environmental factors and plant growth. Hence, Davidson et al. (Citation1998) pointed out that it is perhaps impossible to distinguish statistically the independent effects of temperature and soil moisture within such a complex interaction. Generally, the main predictors of soil CO2 flux vary over crops’ growing seasons. In our study, temperature and soil moisture exerted most of the control over soil CO2 flux in the early and late maize-growth stages, but were greatly overshadowed by plant-growth effects during the crop peak season; this is consistent with previous studies (Liu, Citation1998; Han et al., Citation2007). The amplitude of the seasonal changes in soil respiration was positively correlated with the seasonal changes in LAI. Furthermore, seasonal increases in CO2 fluxes are closely related to increases in root production and biomass (Thomas et al., Citation2000). Liu et al. (Citation2005) also investigated soil respiration in soybean and maize rooting zones, and observed a similar pattern of soil respiration: rather low in the early stages, increasing as the root system develops, and finally declining in the late stages. Our study supports suggestions that air temperature and crop root respiration control soil surface CO2 flux, and that temperature effects decline with enhanced root respiration as the crop grows. We also observed that soil CO2 flux had a stronger temporal relationship with air temperature than with soil temperature, indicating that CO2 flux patterns at our site may be more closely related to environmental conditions near the soil surface.
Farmland ecosystems, one of the most important ecosystems worldwide in terms of C budget, have recently come under increasing scrutiny in the context of climate change because of their potential to affect net soil CO2 productivity and exchange between the soil and the atmosphere, as a result of deliberate human activities. Land-management practices, such as tillage, fertilizer application, and irrigation, can influence soil microclimate, which can directly or indirectly affect CO2 fluxes from the soil surface. For example, tillage accelerates soil CO2 emissions by improving soil aeration, disaggregating soil, increasing the contact between the soil and crop residues, and speeding up organic C decomposition (Al-Kaisi & Yin, Citation2005). In the temperate steppe ecosystem, N fertilization stimulates soil respiration primarily through enhancing plant growth, belowground C input, root activity, and respiration (Carreiro et al., Citation2000).
In this study, soil CO2 flux in spring maize fields on the Loess Plateau was influenced by different cultivation practices as a result of regulating plant growth activity, microbial habitat processes, and CO2 diffusion from the surface of the soil. In the SI treatment, mean diurnal and seasonal soil surface CO2 effluxes were much higher than in the RF treatment; it is likely that irrigation relieved the inhibition of plant growth caused by water deficit, stimulated plant growth and microbial activity, and hence increased soil CO2 flux ( Zhou et al., Citation2002). This result is consistent with previous studies of soybean (Yang & Cai, Citation2004).
Mulching, either with plastic film or straw, can significantly affect soil microclimate (soil temperature and moisture content) (Lentz & Bjorneberg, Citation2003; Ghosh et al., 2006) and, hence, soil respiration, but their effects are not necessarily the same. In addition to the hydrothermal effect, film mulching can block the diffusion of CO2 from the soil surface into the atmosphere. Few studies have examined soil CO2 flux in film-mulched fields. Here we measured the flux inside a PVC chamber, installed the day before to allow overnight equilibrium without the mulch. It was hoped that this would eliminate any blocking effect; however, the measurements were still much higher than for any of the other treatments. This was because film mulching probably improved the soil microclimate, i.e., increased soil temperature and moisture content, and hence accelerated soil respiration. Unlike film mulching, straw mulching decreased soil temperature; this may have decreased root and microbial activities, but increased the quantity and quality of respiration substrates (Lei et al., Citation2005), thus more than compensating for any reductions and resulting in a net increase in CO2 flux.
Acknowledgements
This work was supported by the National Basic Research Program of China (2009CB118604) and National Natural Science Foundation of China (90502006).
Related Research Data
References
- Al-Kaisi , M.M. and Yin , X. 2005 . Tillage and crop residue effects on soil carbon and carbon dioxide emission in corn-soybean rotations . Journal of Environmental Quality , 34 : 437 – 445 .
- Amos , B. , Arkebauer , T.J. and Doran , J.W. 2005 . Soil surface fluxes of greenhouse gases in an irrigated maize-based agroecosystem . Soil Science Society of America Journal , 69 : 387 – 395 .
- Anderson , J.M. 1973 . Carbon dioxide evolution from two temperate, deciduous woodland soils . Journal of Applied Ecology , 10 : 361 – 378 .
- Bain , W.G. , Hutyra , L. , Patterson , D.C. , Bright , A.V. , Daube , B.C. , Munger , J.W. and Wofsy , S.C. 2005 . Wind-induced error in the measurement of soil respiration using closed dynamic chambers . Agricultural and Forest Meteorology , 131 : 225 – 232 .
- Ball , B.C. , Scott , A. and Parker , J.P. 1999 . Field N2O, CO2 and CH4 fluxes in relation to tillage, compaction and soil quality in Scotland . Soil and Tillage Research , 53 : 29 – 39 .
- Boone , R.D. , Nadelhoffer , K.J. , Canary , J.D. and Kaye , J.P. 1998 . Root exerts a strong influence on the temperature sensitivity of soil respiration . Nature , 396 : 570 – 572 .
- Borken , W. , Muhs , A. and Beese , F. 2002 . Application of compost in spruce forests: Effects on soil respiration, basal respiration and microbial biomass . Forest Ecology and Management , 159 : 49 – 58 .
- Bottner , P. 1985 . Response of microbial biomass to alternate moist and dry conditions in a soil incubated with 14C- and 15N-labeled plant material . Soil Biology and Biochemistry , 17 : 329 – 337 .
- Buyanowski , G.A. and Wagner , G.H. 1983 . Annual cycles of carbon dioxide level in soil air . Soil Science Society of America Journal , 47 : 1139 – 1145 .
- Cao , G.M. , Tang , Y.H. , Mo , W.H. , Wang , Y.S. , Li , Y.N. and Zhao , X.Q. 2004 . Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau . Soil Biology and Biochemistry , 36 : 237 – 243 .
- Carlisle , E.A. , Steenwerth , K.L. and Smart , D.R. 2006 . Effects of land use on soil respiration: conversion of oak woodlands to vineyards . Journal of Environmental Quality , 35 : 1396 – 1404 .
- Carreiro , M.M. , Sinsabaugh , R.L. , Repert , D.A. and Parkhurst , D.F. 2000 . Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition . Ecology , 81 : 2359 – 2365 .
- Davidson , E.A. , Belk , E. and Boone , R.D. 1998 . Soil water content and temperature as independent of confounded factors controlling soil respiration in a temperate mixed hardwood forest . Global Change Biology , 4 : 217 – 227 .
- Davidson , E.A. , Verchot , L.V. , Cattanio , J.H. , Ackerman , I.L. and Carvalho , J.E.M. 2000 . Effects of soil water content on soil respiration in forests and cattle pasture of eastern Amazonia . Biochemistry , 48 : 53 – 69 .
- Davidson , E.A. , Savage , K. , Verchot , L.V. and Navarro , R. 2002 . Minimizing artifacts and biases in chamber-based measurements of soil respiration . Agricultural and Forest Meteorology , 113 : 21 – 37 .
- Edwards , N.T. 1975 . Effects of temperature and moisture on carbon dioxide evolution in a mixed deciduous forest floor . Soil Science Society of America Journal , 39 : 361 – 365 .
- Epron , D. , Le Dantec , V. , Dufrene , E. and Granier , A. 2001 . Seasonal dynamics of soil carbon dioxide efflux and simulated rhizosphere respiration in a beech forest . Tree Physiology , 21 : 145 – 152 .
- Ghosh , P.K. , Dayal , D. , Bandyopadhyay , K.K. and Mohanty , M. 2006 . Evaluation of straw and polythene mulch for enhancing productivity of irrigated summer groundnut . Field Crops Research , 99 : 76 – 86 .
- Gong , Z.T. 1999 . Chinese Soil Taxonomy . Beijing Science Press Publishing (in Chinese) .
- Han , G.X. , Zhou , G.S. , Xu , Z.Z. , Yang , Y. , Liu , J.L. and Shi , K.Q. 2007 . Biotic and abiotic factors controlling the spatial and temporal variation of soil respiration in an agricultural ecosystem . Soil Biology and Biochemistry , 39 : 418 – 425 .
- Jabro , J.D. , Sainju , U. , Stevens , W.B. and Evans , R.G. 2008 . Carbon dioxide flux as affected by tillage and irrigation in soil converted from perennial forages to annual crops . Journal of Environmental Management , 4 : 1478 – 1484 .
- Jensen , L.S. , Mueller , T. , Tate , R.K. , Riss , D.J. , Magid , J. and Nelson , N.E. 1996 . Soil surface CO2 flux as an index of soil respiration in situ: a comparison of two chamber methods . Soil Biology and Biochemistry , 28 : 1297 – 1306 .
- Kern , J.S. and Johnson , M.G. 1993 . Conservation tillage impacts on national soil and atmospheric carbon levels . Soil Science Society of America Journal , 57 : 200 – 210 .
- Kieft , T.L. , Soroker , E. and Firestone , M.K. 1987 . Microbial biomass to a rapid increase in water potential when dry soil is wetted . Biology and Biochemistry , 19 : 119 – 126 .
- Lei , H.J. , Li , B.G. , Bai , Y.L. , Huang , Y.X. , & Li , K.J. 2005 . Estimation of soil organic matter mineralization in the plough layer and CO2 emission under different farming practices in China's Huang-Huai-Hai Plain . Scientia Agricultura Sinica , 38 , 2035 2042 (in Chinese with English abstract).
- Lentz , R.D. and Bjorneberg , D.L. 2003 . Polyacrylamide and straw residue effects on irrigation furrow erosion and infiltration . Journal of Soil and Water Conservation , 58 : 312 – 319 .
- Li , F.M. , Guo , A.H. and Wei , H. 1999 . Effects of clear plastic film mulch on yield of spring wheat . Field Crops Research , 63 : 79 – 86 .
- Li , F.M. , Wang , J. , Xu , J.Z. and Xu , H.L. 2004a . Productivity and soil response to plastic film mulching durations for spring wheat on entisols in the semiarid Loess Plateau of China . Soil and Tillage Research , 78 : 9 – 20 .
- Li , F.M. , Song , Q.H. , Jjemba , P.K. and Shi , Y.C. 2004b . Dynamics of soil microbial biomass C and soil fertility in cropland mulched with plastic film in a semiarid agro-ecosystem . Soil Biology and Biochemistry , 36 : 1893 – 1902 .
- Liu , Q.H. , Huang , Y. , & Zheng , X.H. 2005 . Determination of upland soil respiration and its components with BaPS system . Acta Scientiae Circumstantiae , 25 , 1105 1111 (in Chinese with English abstract) .
- Liu , Y.F. 1998 . A tentative study of CO2 emission from farmland soil in Tibetan Plateau . Journal of Natural Resources , 13 , 181 186 (in Chinese with English abstract) .
- Maier , C.A. and Kress , L.W. 2000 . Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability . Canadian Journal of Forest Research , 30 : 347 – 359 .
- Mielnick , P.C. and Dugas , W.A. 2000 . Soil CO2 flux in a tallgrass prairie . Soil Biology and Biochemistry , 32 : 221 – 228 .
- Parkin , T.B. and Kaspar , T.C. 2003 . Temperature controls on diurnal carbon dioxide flux: implications for estimating soil carbon loss . Soil Science Society of America Journal , 67 : 1763 – 1772 .
- Phene , C.J. and Sanders , D.C. 1976 . High-frequency trickle irrigation and row spacing effects on yield and quality of potatoes . Agronomy Journal , 68 : 602 – 607 .
- Rastogi , M. , Singh , S. and Pathak , H. 2002 . Emission of carbon dioxide from soil . Current Science , 82 : 510 – 517 .
- Rochette , P. , Desjardins , R.L. and Pattey , E. 1991 . Spatial and temporal variability of soil respiration in agricultural fields . Canadian Journal of Soil Science , 71 : 189 – 196 .
- Thomas , S.M. , Cook , F.J. , Whitehead , D. and Adams , J.A. 2000 . Seasonal soil-surface carbon fluxes from the root systems of young Pinus radiata trees growing at ambient and elevated CO2 concentration . Global Change Biology , 6 : 393 – 406 .
- Tiquia , S.M. , Lloyd , J. , Herms , D.A. , Hoitink , H.A.J. and Michel , F.C. Jr. 2002 . Effects of mulching and fertilization on soil nutrients, microbial activity and rhizosphere bacterial community structure determined by analysis of TRFLPs of PCR-amplified 16S rRNA genes . Applied Soil Ecology , 21 : 31 – 48 .
- Wang , W.J. , Zu , Y.G. , Wang , H.M. , Hirano , T. , Takagi , K. , Sasa , K. and Koike , T. 2005 . Effect of collar insertion on soil respiration in a larch forest measured with a LI-6400 soil CO2 flux system . Journal of Forest Research , 10 : 57 – 60 .
- Xu , M. and Qi , Y. 2001 . Soil surface CO2 efflux and its spatial and temporal variations in a young ponderosa pine plantation in northern California . Global Change Biology , 7 : 667 – 677 .
- Yang , L.F. , & Cai , Z.C. 2004 . Diurnal variation of soil respiration in soil-potted with soybean (Glycine max L.) at various growth stages and its affecting factors . Acta Ecologica Sinica , 24 , 2955 2960 (in Chinese with English abstract) .
- Zhou , Z.T. , Cheng , S.K. , Liu , Y.F. , & Li , J.Y. 2002 . CO2 emission of soil under different land-use types in subtropical red soil hilly areas in China: preliminary exploration . Resources Science , 24 , 83 87 (in Chinese with English abstract) .