
Abstract
Acrolein is a reactive α,β-unsaturated aldehyde derived from lipid peroxides, which are produced in plants under a variety of stress. We investigated effects of acrolein on light-induced stomatal opening using Arabidopsis thaliana. Acrolein inhibited light-induced stomatal opening in a dose-dependent manner. Acrolein at 100 μM inhibited plasma membrane inward-rectifying potassium (Kin) channels in guard cells. Acrolein at 100 μM inhibited Kin channel KAT1 expressed in a heterologous system using Xenopus leaves oocytes. These results suggest that acrolein inhibits light-induced stomatal opening through inhibition of Kin channels in guard cells.
Graphical Abstract
A reactive α,β-unsaturated aldehyde, acrolein, inhibits Kin channel currents through inhibition of KAT1, resulting in inhibition of light-induced stomatal opening.
Reactive carbonyl species (RCS) are electrophilic α,β-unsaturated aldehydes and endogenous products through reactive oxygen species (ROS)-initiated peroxidation of polyunsaturated fatty acids in cell membranes in plants under a variety of stresses.Citation1–5) Light stress increases the concentration of acrolein from 4.8 to 11.7 μM in whole leaves of Nicotiana tabacum.Citation5) Acrolein (2-propenal) is one of the most reactive RCS which has potential to modify proteins.Citation6) Hence, acrolein accumulation seems to be closely involved in stress responses.
Plants control gas exchange and water loss through modulation of apertures of stomata, which are pores formed by a pair of guard cells in the epidermis. Guard cells respond to numerous biotic and abiotic stimuli, resulting in stomatal opening or closure.Citation7–9) Light-induced stomatal opening is accompanied by uptake of K+ into guard cells,Citation10) which is mediated by inward-rectifying K+ (Kin) channels in plasma membrane of guard cells.Citation11) A dominant negative mutation of KAT1, which is a Kin channel expressed in Arabidopsis thaliana guard cells, reduces guard cell Kin channel currents in A. thaliana, resulting in suppression of light-induced stomatal opening.Citation12) These studies indicate that well-functioning KAT1 is essential for stomatal opening.
The effects of acrolein in animals have been well studied,Citation13,14) but its effects in plants remain to be clarified.Citation15) It is unclear whether acrolein affects light-induced stomatal movement or ion channels in plants. In this study, we examined the effects of acrolein on light-induced stomatal opening and guard cell Kin channels in A. thaliana, and KAT1 expressed in Xenopus leaves oocytes.
Materials and methods
Plant materials and growth conditions
Arabidopsis (A. thaliana) ecotype Columbia was grown on soil containing 70% (v/v) vermiculite (Asahi-kogyo, Okayama, Japan) and 30% (v/v) Kureha soil (Kureha Chemical, Tokyo, Japan) in a growth chamber at 21 ± 2 °C and 60% relative humidity with a 16-h light period with 80 μmol m−2 s−1 photon flux density and 8 h of dark. Water containing 0.1% Hyponex (Hyponex Japan, Osaka, Japan) was applied to the plant growth tray two to three times a week.
Measurement of stomatal aperture
Stomatal apertures were measured as described previously,Citation16) with modification. Excised rosette leaves of four- to six-week-old were floated on an assay solution containing 5 mM KCl, 50 μM CaCl2, and 10 mM MES-Tris (pH 6.15). The rosette leaves were incubated for 2 h in the dark followed by addition of acrolein, and then were incubated in the light (80 μmol m−2 s−1). For measurement of stomatal apertures, the leaves were shredded in a commercial blender for 25 s and epidermal tissues were collected using nylon mesh. For each sample, 20 stomatal apertures were measured.
Patch-clamp measurement
Whole-cell Kin channel current measurements of A. thaliana guard cell protoplasts (GCPs) were performed as described previously.Citation16) The GCPs were prepared from rosette leaves of four- to six-week-old plants with the digestion solution containing 1.0% (w/v) Cellulase R10, 0.5% (w/v) Macerozyme R10, 0.5% (w/v) bovine serum albumin, 0.1% (w/v) kanamycin, 10 mM ascorbic acid, 0.1 mM KCl, 0.1 mM CaCl2, and 500 mM D-mannitol, (pH 5.5), with KOH. Whole-cell currents were recorded using a patch-clamp amplifier (model CEZ-2200; Nihon Kohden, Tokyo, Japan). No leak subtraction was applied for all current–voltage curves. For data analysis, a software, pCLAMP 10.3 (Molecular Devices, Sunnyvale, CA) was used. Pipette solution contained 30 mM KCl, 70 mM K-Glu, 2 mM MgCl2, 3.35 mM CaCl2, 6.7 mM EGTA, and 10 mM HEPES-Tris (pH 7.1). Bath solution contained 30 mM KCl, 2 mM MgCl2, 40 mM CaCl2, and 10 mM MES-Tris (pH 5.5). Osmolarity was adjusted to 500 mmol kg−1 (pipette solutions) and 485 mmol kg−1 (bath solutions) with D-sorbitol.
KAT1 gene cloning and construction of expression vector
A. thaliana KAT1 gene was cloned into pNB1u expression vector for heterologous expression in Xenopus laeves oocytes using an advanced uracil-excision-based cloning technique as described previouslyCitation17). A. thaliana cDNA was amplified by polymerase chain reaction (PCR) using PfuTurbo Cx Hotstart DNA polymerase (Agilent Technologies, La Jolla, CA). The primers used in PCR amplification were as follows 5′-GGCTTAAUATGTCGATCTCTTGGACTCG-3′ (KAT1_USER_F) and 5′-GGTTTAAUTCAATTTGATGAAAAATACAAATG-3′ (KAT1_USER_R). Circular pNB1u-KAT1 plasmid DNA was linearized by digestion with restriction enzyme NotI (New England Biolabs, Hertfordshire, UK). The Arabidopsis Genome Initiative number for KAT1 is AT5g46240.
Measurement of KAT1 currents in Xenopus leaves oocytes
For expression in oocytes, cRNAs were synthesized using a mMESSAGE mMACHINE® T7 transcription kit with cap analog (Ambion, Austin, TX) from linearized pNB1u-KAT1plasmid DNA as described previouslyCitation18) with modification. Xenopus oocytes were defolliculated using collagenase and microinjected with 50 ng of cRNAs using a microinjector (model Nanoject II; Drummond Scientific, Broomall, PA) after 3 d of incubation in ND96(+)containing 94 mM NaCl, 2 mM KCl, 1 mM CaCl2.2H2O, 1 mM MgCl2.6H2O, 5 mM HEPES (pH 7.5), and 50 μg/mL gentamicin at 18 °C. Two-electrode voltage clamp experiments were performed using Digidata 1440A and Axoclamp 900A amplifiers (Molecular Devices) at 22 °C. Data were processed with a software, Clampex 10.2 (Molecular Devices). The microelectrodes contained 3 M KCl. The bath solution contained 100 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, and 10 mM HEPES titrated to pH 7.5 with NaOH.
Statistical analysis
Data were analyzed by analysis of variance (ANOVA), and mean values for stomatal apertures and for ion channel currents were compared by Tukey test and Student’s t-test, respectively. We regarded differences at the level of P < 0.05 as significant.
Results
Inhibition by acrolein of light-induced stomatal opening
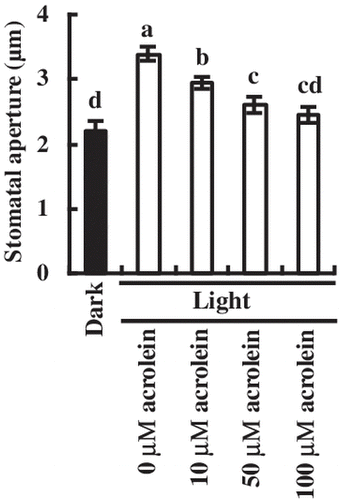
We observed stomatal movement in the absence and the presence of acrolein under light and dark conditions (Fig. ). Incubation for 3 h in the light caused stomatal opening (Fig. ). Application of acrolein at 10 μM, 50 μM, and 100 μM significantly inhibited light-induced stomatal opening by 13.2% (P < 0.002), 23.6% (P < 0.001), and 27.7% (P < 0.0006), respectively, indicating that acrolein inhibits light-induced stomatal opening in a dose-dependent manner (Fig. ).
Fig. 1. Inhibition by acrolein of light-induced stomatal opening in A. thaliana.
The rosette leaves were incubated for 2 h in the dark (black bar) and then were incubated for 3 h in the light (80 μmol m−2 s−1) followed by addition of acrolein (white bars). Bars having same letters do not differ significantly at the 5% level (one-way ANOVA with Tukey test). Averages for three independent experiments (60 stomata per bar) are shown. Error bars represent SEM.
Suppression of Kin channel currents by acrolein in guard cells
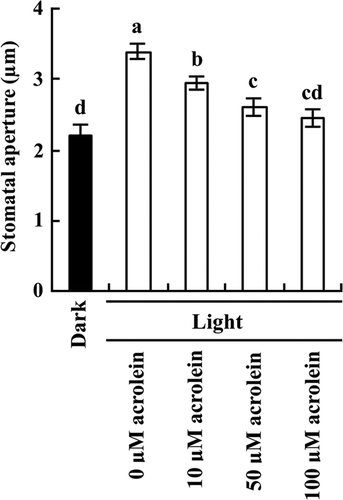
We measured Kin channel currents in A. thaliana GCPs untreated and treated with acrolein using a whole-cell patch clamp technique (Fig. ). The typical voltage-dependent Kin channel currents were observed for the acrolein-untreated guard cells (Fig. (A) and (C)). Treatment with 100 μM acrolein for 2 h significantly decreased Kin channel currents in GCPs by 73.4% (P < 0.001 at –180 mV; Fig. (B) and (C)).
Fig. 2. Inhibition by acrolein of Kin channel currents in GCPs of A. thaliana.
(A) and (B), Whole-cell recordings of Kin channel currents in GCPs treated with 0 μM acrolein (A) or with 100 μM acrolein (B). (C), Steady-state current-voltage relationship for acrolein inhibition of Kin channel currents in GCPs as recorded in A and B. The voltage protocol was stepped up from 0 mV to –180 mV in 20-mV decrements (holding potential, –40 mV). GCPs were treated with acrolein for 2 h before recordings. Each datum point was obtained from at least seven GCPs. Error bars represent SEM.
Suppression of inward currents by acrolein in KAT1-expressing Xenopus leaves oocytes
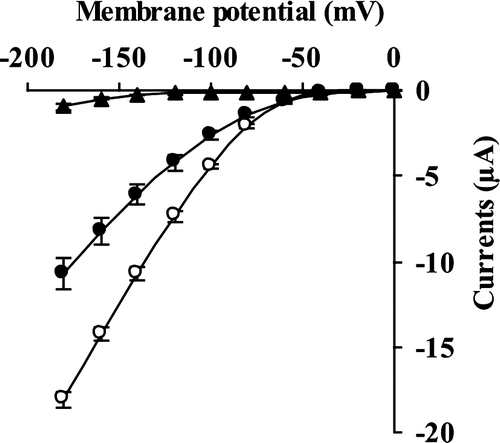
We measured inward currents in oocytes using the two-electrode voltage clamp technique (Fig. ). Compared with the water-injected oocytes, the KAT1-injected oocytes showed larger inward currents (Fig. ), which is in agreement with the previous result.Citation19) The inward currents were significantly reduced by treatment with 100 μM acrolein (P < 0.0001 at –180 mV; Fig. ).
Fig. 3. Suppression by acrolein of KAT1 channel currents in Xenopus leaves oocytes.
Inward currents were recorded from oocytes injected with 50 nl of water (▲) and 50 nl of water containing 2 ng KAT1 cRNA (○, ●). For acrolein treatment, the injected oocytes were incubated in ND96(+) supplemented with 100 μM acrolein (●) for 2 h before recording. The voltage protocol was stepped up from 0 mV to –180 mV in 20 mV decrements (holding potential, –40 mV) with a pulse duration of 3 s. Each datum point was obtained from at least eight oocytes. Error bars represent SEM.
Discussion
Acrolein (2-propenal) is one of the most reactive RCS because of its electrophilicity and consequently has the potential to modify proteins through reaction with nucleophiles, such as the sulfhydryl group of cysteine, imidazole group of histidine, and amino group of lysine.Citation6) RCS is generated from lipid peroxides of which production is initiated by ROS.Citation1,2,4) A variety of abiotic and biotic stress induces ROS production. Hence, acrolein can cause stress-induced damage via modification of critical target molecules.
In this study, acrolein at 10–100 μM inhibited light-induced stomatal opening (Fig. ). The acrolein concentration in whole leaves under normal conditions is in a range of 4–5 μM, and increases 150% in whole leaves under stress conditions,Citation5) indicating that the concentration of acrolein can reach the sub-mM level. Hence, in stressed plants, light-induced stomatal opening is likely to be inhibited by acrolein, resulting in suppression of carbon dioxide uptake and transpirational water loss.
Light-induced stomatal opening is accompanied by K+ uptake into guard cells,Citation10) which is mediated by Kin channels.Citation11) The dominant negative mutants of KAT1 show reduction of Kin channel currents and impairment of light-induced stomatal opening.Citation12) In this study, acrolein inhibited Kin channel currents (Fig. ) and KAT1 channel currents (Fig. ). These results suggest that inhibition by acrolein of Kin channel currents due to inhibition by acrolein of KAT1 channel currents results in inhibition by acrolein of light-induced stomatal opening in A. thaliana.
A variety of stimuli do induce not only ROS and RCS production but also reactive nitrogen species (RNS) production in guard cells. Treatment of RNS, such as nitric oxide (NO), as well as treatment of ROS, such as H2O2, decreases Kin channel currents in guard cellsCitation20,21) and inhibits stomatal opening.Citation22,23) These results suggest that RCS, ROS, and RNS affect light-induced stomatal opening through modulation of Kin channel activities in stressed plants. However, these reactive compounds have different lifetime and reactivity to proteins, which may make differences in extents and kinetics of inhibition of stomatal opening.
The phosphorylation of C-terminal regions of KAT1 by protein kinase C (PKC)Citation24) and stress-activated protein kinase SnRK2.6 (Snf1-related protein kinase 2.6)Citation25) modulates the activity of the KAT1 channel, indicating that the modification of C-terminal regions of KAT1 changes the activity of KAT1 channel. Hence, acrolein may also modulate the activity via modification of the C-terminal regions of KAT1.
It is concluded that acrolein inhibits light-induced stomatal opening, which is partially due to inhibition of Kin channels in guard cells.
Acknowledgments
We are grateful to Dr Hussam Hassan Nour-Eldin (University of Copenhagen) for supplying expression vector pNB1u.
Notes
Abbreviations: GCP, guard cell protoplast; Kin channel, inward-rectifying potassium channel; PCR, polymerase chain reaction; RCS, reactive carbonyl species; ROS, reactive oxygen species.
References
- Mueller MJ. Archetype signals in plants. Curr. Opin. Plant. Biol. 2004;7:441–448.10.1016/j.pbi.2004.04.001
- Mosblech A, Feussner I, Heilmann I. Oxylipins: structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009;47:511–517.
- Mano J. Reactive carbonyl species: their production from lipid peroxides, action in environmental stress, and the detoxification mechanism. Plant Physiol. Biochem. 2012;59:90–97.
- Yin L, Mano J, Wang S, Tsuji W, Tanaka K. The involvement of lipid peroxide-derived aldehydes in aluminum toxicity of tobacco roots. Plant Physiol. 2010;152:1406–1417.10.1104/pp.109.151449
- Mano J, Tokushige K, Mizoguchi H, Fujii H, Khorobrykh S. Accumulation of lipid peroxide-derived, toxic α,β-unsaturated aldehydes (E)-2-pentenal, acrolein and (E)-2-hexenal in leaves under photoinhibitory illumination. Plant Biotechnol. 2010;27:193–197.10.5511/plantbiotechnology.27.193
- Esterbauer H, Schauer RJ, Zollner H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991;11:81–128.10.1016/0891-5849(91)90192-6
- Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D. Guard cell signal transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001;52:627–658.
- Melotto M, Underwood W, Kocjan J, Nomura K, He SY. Plant stomata function in innate immunity against bacterial invasion. Cell. 2006;126:969–980.10.1016/j.cell.2006.06.054
- Shimazaki K, Doi M, Assmann SM, Kinoshita T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007;58:219–247.10.1146/annurev.arplant.57.032905.105434
- Humble GD, Raschke K. Stomatal opening quantitatively related to potassium transport. Plant Physiol. 1971;48:447–453.10.1104/pp.48.4.447
- Schroeder JI, Hedrich R, Fernandez JM. Potassium-selective single channels in guard cell protoplasts of Vicia faba. Nature. 1984;312:361–362.10.1038/312361a0
- Kwak JM, Murata Y, Baizabal-Aguirre VM, Merrill J, Wang M, Kemper A, Hawke SD, Tallman G, Schroeder JI. Dominant negative guard cell K+ channel mutants reduce inward-rectifying K+ currents and light-induced stomatal opening in Arabidopsis. Plant Physiol. 2001;127:473–485.10.1104/pp.010428
- O’Brien PJ, Siraki AG, Shangari N. Aldehyde sources, metabolism, molecular toxicity mechanisms, and possible effects on human health. Crit. Rev. Toxicol. 2005;35:609–662.10.1080/10408440591002183
- Gan JC, Oandasan A, Ansari GA. In vitro covalent modification of serum albumin by acrolein. Chemosphere. 1991;23:939–947.10.1016/0045-6535(91)90098-X
- Mano J, Miyatake F, Hiraoka E, Tamoi M. Evaluation of the toxicity of stress-related aldehydes to photosynthesis in chloroplasts. Planta. 2009;230:639–648.10.1007/s00425-009-0964-9
- Ye W, Muroyama D, Munemasa S, Nakamura Y, Mori IC, Murata Y. Calcium-dependent protein kinase CPK6 positively functions in induction by yeast elicitor of stomatal closure and inhibition by yeast elicitor of light-induced stomatal opening in Arabidopsis. Plant Physiol. 2013;163:591–599.10.1104/pp.113.224055
- Nour-Eldin HH, Hansen BG, Nørholm MHH, Jensen JK, Halkier BA. Advancing uracil-excision based cloning towards an ideal technique for cloning PCR fragments. Nucleic Acids Res. 2006;34:e122.10.1093/nar/gkl635
- Hoque TS, Okuma E, Uraji M, Furuichi T, Sasaki T, Hoque MA, Nakamura Y, Murata Y. Inhibitory effects of methylglyoxal on light-induced stomatal opening and inward K+ channel activity in Arabidopsis. Biosci. Biotechnol. Biochem. 2012;76:617–619.10.1271/bbb.110885
- Véry AA, Gaymard F, Bosseux C, Sentenac H, Thibaud JB. Expression of a cloned plant K+ channel in Xenopus oocytes: analysis of macroscopic currents. Plant J. 1995;7:321–332.10.1046/j.1365-313X.1995.7020321.x
- Garcia-Mata C, Gay R, Sokolovski S, Hills A, Lamattina L, Blatt MR. Nitric oxide regulates K+ and Cl– channels in guard cells through a subset of abscisic acid-evoked signaling pathways. Proc. Natl Acad. Sci. USA. 2003;100:11116–11121.10.1073/pnas.1434381100
- Xiao Z, Miao YC, An GY, Zhou Y, Shangguan ZP, Gao JF, Song CP. K+ channels inhibited by hydrogen peroxide mediate abscisic acid signaling in Vicia guard cells. Cell Res. 2001;11:195–202.
- Garcia-Mata C, Lamattina L. Abscisic acid (ABA) inhibits light-induced stomatal opening through calcium- and nitric oxide-mediated signaling pathways. Nitric Oxide. 2007;17:143–151.10.1016/j.niox.2007.08.001
- Yan J, Tsuichihara N, Etoh T, Iwai S. Reactive oxygen species and nitric oxide are involved in ABA inhibition of stomatal opening. Plant Cell Environ. 2007;30:1320–1325.10.1111/pce.2007.30.issue-10
- Sato A, Gambale F, Dreyer I, Uozumi N. Modulation of the Arabidopsis KAT1 channel by an activator of protein kinase C in Xenopus laevis oocytes. FEBS J. 2010;277:2318–2328.10.1111/j.1742-4658.2010.07647.x
- Sato A, Sato Y, Fukao Y, Fujiwara M, Umezawa T, Shinozaki K, Hibi T, Taniguchi M, Miyake H, Goto DB, Uozumi N. Threonine at position 306 of the KAT1 potassium channel is essential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6 protein kinase. Biochem. J. 2009;424:439–448.10.1042/BJ20091221