
Abstract
To optimize Agrobacterium-mediated transient transformation assay in mulberry (Morus alba L.), various infiltration methods, Agrobacterium tumefaciens (A. tumefaciens) strains, and bacterial concentrations were tested in mulberry seedlings. Compared with LBA4404, GV3101 harboring pBE2133 plasmids presented stronger GUS signals at 3 days post infiltration using syringe. Recombinant plasmids pBE2133:GFP and pBE2133:GFP:MaFT were successfully constructed. Transient expression of MaFT:GFP protein was found in leaves, petiole (cross section), and shoot apical meristem (SAM) of mulberry according to the GFP signal. Moreover, MaFT:GFP mRNA was also detected in leaves and SAM via RT-PCR and qRT-PCR. An efficient transient transformation system could be achieved in mulberry seedlings by syringe using A. tumefaciens GV3101 at the OD600 of 0.5. The movement of MaFT expression from leaves to SAM might trigger the precocious flowering of mulberry.
Graphical abstract
Transient MaFT expression could be achieved in mulberry by syringe using GV3101 (OD600 = 0.5). MaFT might trigger the precocious flowering of mulberry.
Mulberry (Morus alba L.) has been cultivated over 4000 years and its cultural significance has persisted throughout human history. Mulberries have been widely used in traditional Oriental medicine for several applications as anti-diabetic, anti-headache, anti-hypertensive, and diuretic agent.Citation1,2) Recently, the genome sequence has been drafted for the mulberry tree Morus notabilis in 2013.Citation3) Transient transformation is a powerful tool to analyze the function of genes in plants and is more efficient than stable transformation regarding to time and labor consuming.Citation4,5) Transient gene expression techniques, such as biolistic bombardment, protoplast transfection, and Agrobacterium-mediated infiltration (agro-infiltration), are available for model plants such as Arabidopsis, tobacco, and crop plants, including corn, potato, soybean, tomato, and wheat.Citation6–10) Agro-infiltration is a simple transformation process that exhibits highly efficient transformation and easy operation in several plant species including Arabidopsis, tobacco, grapevine, potato, switchgrass, and tomato.Citation9–14) In woody plants, several studies of transient gene expression by biolistic bombardment, protoplast transfection, and Agrobacterium co-cultivation in Populus have been reported,Citation15,16) while there is limited report on transient gene expression in mulberry.
The flowering locus T (FT) gene plays a crucial role in regulating the transition from the vegetative phase to the reproductive phase in plants.Citation17) FT gene encodes a small protein that has a similar structure to mammalian phosphatidy ethanolamine-binding protein.Citation18) FT genes of many plants have been isolated, and overexpression of FT homologous genes from Arabidopsis, rice, poplar, tomato, peas, cucurbits, sunflower, apple, and alfalfa have resulted in precocious flowering.Citation19–24) However, functional study of Morus alba flowering locus T (MaFT) gene in mulberry is very limited. Thus, the Agrobacterium-mediated transient expression system in mulberry was firstly optimized. The recombinant plasmids pBE2133:GFP and pBE2133:GFP:MaFT were then constructed to analyze the function of MaFT gene. In addition, the transient expression of MaFT:GFP was detected via fluorescence observation, reverse transcription polymerase chain reaction (RT-PCR), and quantitative RT-PCR (qRT-PCR), respectively.
Materials and methods
Plant and Agrobacterium tumefaciens strains
Mulberry breeds were cultivated in a Petri dish with water for 3 days at 4 °C and then incubated in a greenhouse (22 °C, 18 h light/6 h darkness, humidity: 46%) (Institute of Sericulture and Silk, Northwest A&F University). Upon the appearance of white roots, plants were transplanted into flowerpots (10 cm × 10 cm) with substrates. Then, flowerpots were placed in a growth chamber for 2 months (26 °C, 18 h light/6 h darkness). When mulberry seedlings were cultivated, no biological control was performed.
Agrobacterium tumefaciens (A. tumefaciens) strains, GV3101 and LBA4404, (State Key laboratory for Silkworm Genome Biology, Northwest A&F University) were grown on Luria broth (LB) medium supplemented with 50 mg/L rifampicin and 50 mg/L gentamicin (TaKaRa Bio Technology Co. Ltd, Dalian, China). Furthermore, 50 mg/L kanamycin (TaKaRa Bio Technology Co., Ltd, Dalian, China) was used to select bacterium harboring pBE2133 plasmid. The pBE2133 plasmids containing β-glucuronidase (GUS) gene were provided by Prof. Mitsuhara Ichiro (National Institute of Agrobiological Resources, Japan).
Optimization of Agrobacterium-mediated transient expression system
The pBE2133 (pBE2133:GUS) plasmids were, respectively, introduced into competent cells of A. tumefaciens strain GV3101 and LBA4404 via fast freezing method (5 μL purified plasmid per 100 μL of Agrobacterium competent cells) to analyze the transient transformation efficiency through agro-infiltration.Citation25) Firstly, competent cells of A. tumefaciens GV3101 and LBA4404 were obtained. Then, pBE2133:GUS plasmids were introduced into the competent cells to obtain recombinant cells. The recombinant cells were gently mixed, placed in an ice bath for 5 min and then quickly frozen in liquid nitrogen for 10 min followed by thawing in a 37 °C water bath for 5 min. After incubation for 2–4 h by gentle shaking at 28 °C, cells were plated on selective LB medium supplemented with 50 mg/L kanamycin, 50 mg/L gentamycin, and 50 mg/L rifamycin for 2 days at 28 °C.
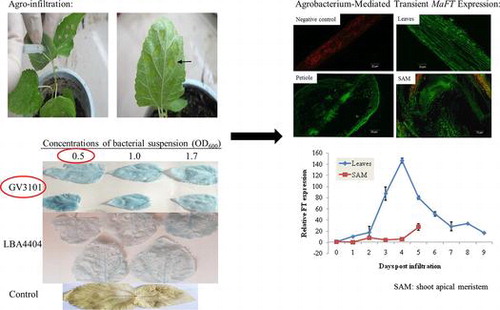
Two days prior to agro-infiltration, single colonies of A. tumefaciens GV3101 and LBA4404 harboring pBE2133:GUS plasmids were inoculated in 5 mL antibiotic-supplemented LB medium. The bacterial cultures were incubated for 2 days at 28 °C on a shaker at 250 rpm, and 1 mL of the culture was used to inoculate 100-mL LB medium supplemented with antibiotics. The new cultures were grown for another day at 28 °C with agitation. Bacterial pellets were obtained by centrifugation (4 °C, 3000 g) for 10 min and washed twice with 5-mL infiltration buffers (10 mM MgCl2 and 100 μM acetosyringone). The bacterial pellets were then resuspended in the infiltration buffer to obtain the desired concentrations (OD600: 0.5, 1.0, and 1.7). Prior to agro-infiltration, the bacterial suspensions were incubated at 4 and 27 °C for 3–4 and 1 h, respectively. The bacterial suspension was infiltrated by 1 mL syringe without a needleCitation26,27) and vacuum infiltrated with the abaxial surface of young leaves with thick mesophylls of mulberry seedlings, respectively. Leaves infiltrated with water were taken as controls. After infiltration, plants were grown under the standard growth conditions.
GUS staining
GUS staining was carried out as the methods previously described.Citation28) Briefly, leaves were collected at 3 days post infiltration, incubated in GUS staining solution at 37 °C for continuing 18 h, and washed with phosphate buffer saline. Finally, the leave tissues were dipped in 90 and 75% ethanol solution to remove chlorophylls until control leaves became white.
Plasmid construction and bacterial transformation
To conveniently observe MaFT transformation, green fluorescence protein (GFP) replaced GUS of pBE2133 plasmid as a reporter gene to construct a recombinant plasmid pBE2133:GFP. The GFP coding sequence was amplified with DNA polymerase from pAdTrack-CMV plasmid, which was obtained from Gongshe Yang (College of Animal Science and Technology, Northwest A&F University), using 5′-CGCGGATCCATGGFGAGCAAGGGCGAGGAGCTGT-3′ (forward) and 5′-TACATGAGCTCCTTGCTCAGGGCGGACTGGGTGCTC-3′ (reverse) containing BamH I (TaKaRa Bio Technology Co., Ltd, Dalian, China) and Sac I (TaKaRa Bio Technology Co., Ltd, Dalian, China) restriction sites (underlined in the sequence), respectively. The product was subcloned into a pMD18-T vector (TaKaRa Bio Technology Co., Ltd, Dalian, China). After digestion of recombinant pMD18-T:GFP vector, the BamH I-Sac I fragment was inserted into the pBE2133 plasmid at the entries of BamH I and Sac I to construct the recombinant plasmid pBE2133:GFP.
Then, MaFT gene was fused in the pBE2133:GFP plasmid to construct a new recombinant plasmid pBE2133:GFP:MaFT. The 522-bp coding sequence of MaFT (http://www.ncbi.nlm.nih.gov/nuccore/JX462956) was amplified using the following primers: 5′-GCTCTAGAATGCCTAGAGATAGAGACCCTATTG-3′ (forward) and 5′-CCGGATCCTTATCTCGGCCTTCTTGCACCAGTT-3′ (reverse), which included an Xba I (TaKaRa Bio Technology Co., Ltd, Dalian, China) and BamH I site (underlined in the sequence). The verified PCR products were cloned into the pMD18-T vector. After digestion of the recombinant pMD18-T:MaFT vector, the Xba I–BamH I fragment was moved into the pBE2133:GFP plasmid which was cleaved at the entries of BamH I and Xba I to construct the new plasmid pBE2133:GFP:MaFT.
The plasmid pBE2133:GFP:MaFT and pBE2133:GFP (the control plasmid) were, respectively, introduced into competent cells of A. tumefaciens GV3101 via fast freezing method.Citation25) Then, the abaxial surface of young leaves with thick mesophylls of mulberry seedlings was infiltrated using the optimized methods. After infiltration, plants were grown under the standard growth conditions.
Detection of GFP
Leaves, petiole (cross section), and SAM of agro-infiltrated mulberry seedlings were collected at 3 days post infiltration and used for frozen section analysis. The GFP signals of these samples were observed and photographed under an inverted fluorescence microscope from Eppendorf (Hamburg, Germany).
RNA extraction and RT-PCR
Mulberry leaves (0, 1, 2, 3, 4, 5, 6, 7, 8, and 9 days after infiltration) and; SAM tissues (0, 1, 2, 3, 4, and 5 days after infiltration) were collected. Samples at 0 day after infiltration were considered as the controls. Mulberry leaves and SAM tissues (100 mg) were homogenized in liquid nitrogen, and total RNA was extracted using TRIZOL (Invitrogen) according to the described protocol. The reverse transcription was conducted at 42 °C for 1 h and 70 °C for 15 min with the moloney murine leukemia virus reverse transcriptase (M-MLV RT, Takara).
The following primer sequences were used: MaFT, 5′-ATGCCTAGAGATAGAGACCCTATTG-3′ (forward), 5′-CTTGCTCAGGGCGGACTGGGTGCTC-3′ (reverse); Actin, 5′-GTGGCCGTACAACTGGTATT-3′ (forward), 5′-GCCAGGTCAAGACGAAGAAT-3′ (reverse). The DNA amplification was conducted in 20 μL reaction volume: 1 μL cDNA, 50 μM dNTP, 100 nM primers, 1 mM MgCl2, 1 U Taq DNA polymerase, and 2 μL 10× Taq buffer. The parameters of PCR cycling were as follows: 35 cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for 30 s, extension at 72 °C for 3 min, and final extension at 72 °C for 10 min.
Quantitative RT-PCR
The cDNA samples from leaves (0–9 days post infiltration) and SAM (0–5 days post infiltration) were used for detecting expression of MaFT via qRT-PCR. The DNA SYBR Green real-time qPCR kit from Bio-TOYOBO Company was used for two-step amplification. The primer sequences of MaFT were listed as follows: 5′-GGATGACCTCAGGACCTTCTA-3′ (forward) and 5′-GTCACCAACCAATGCAAGTATTC-3′ (reverse). The Actin was also used as internal control. The qRT-PCR reaction was taken place in 10-μL system containing 5 μL 2× SYBR Real-time PCR Pre-mixture, 0.3 μL forward primer (10 μM), 0.3 μL reverse primer (10 μM), 2 μL cDNA, and 2.4 μL ddH2O. The PCR program was: 95 °C pre-denaturation for 2 min followed by 40 cycles of 95 °C denaturation for 15 s, 60 °C annealing for 20 s, and a final dissociation stage. Melting cure was generated between 65 and 95 °C. The comparative threshold (Ct) cycle method (2–ΔΔCt) was used for relative quantification and calculations.
Results
Assay optimization
Various infiltration methods were used to evaluate transformation efficiency in mulberry seedlings. Seedling leaves were infiltrated by syringe (Fig. (a) and (b)), while vacuum infiltration required detached leaves. Thus, following-up experiments that demanded undetached leaves were performed using syringe to infiltrate the leaves.
Fig. 1. Agro-infiltration was performed for mulberry seedling leaves.

Based on the GUS staining in the infiltrated areas, both A. tumefaciens strains LBA4404 and GV3101 were observed to be able to induce transformation (Fig. ). Compared with A. tumefaciens LBA4404, GV3101 presented stronger GUS signals at 3 days post infiltration. Meanwhile, no obvious differences of GUS signals were observed among the 3 bacterial concentrations (OD600: 0.5, 1.0, and 1.7) (Fig. ). Thus, the concentration of OD600 = 0.5 was selected for further experiments because it was the lowest concentration yielding consistent transient expression.
Fig. 2. Effects of Agrobacterium strains and concentrations of bacterial suspension (OD600: 0.5, 1.0, and 1.7) on GUS expression.

Transient expression of MaFT
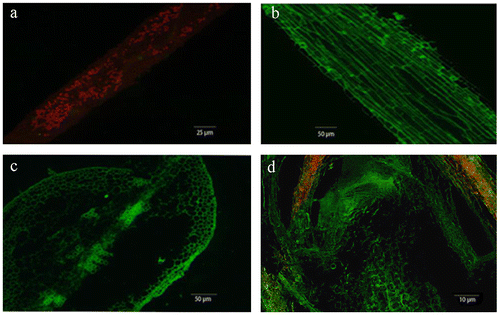
Three days post infiltration, no GFP signal was detected in mulberry leaves infiltrated with Agrobacterium harboring plasmid pBE2133:GFP (Fig. (a)), while the MaFT:GFP fusion protein was present in mulberry leaves infiltrated with Agrobacterium harboring plasmid pBE2133:GFP:MaFT (Fig. (b)). Strong MaFT:GFP signals were also detected in petiole (Fig. (c)) and provasculature at the base of the SAM (Fig. (d)).
Fig. 3. Transient expression of MaFT:GFP was observed under inverted fluorescence microscope at 3 days post infiltration.
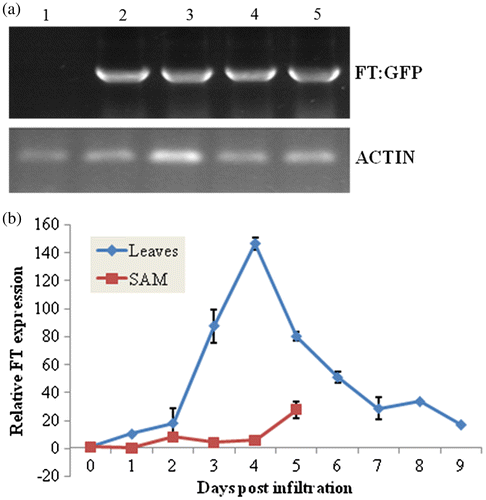
The transient expressions of MaFT:GFP in leaves and SAM were confirmed at mRNA level using RT-PCR (Fig. (a)). According to the qRT-PCR, the expression of MaFT in leaves reached peak level at 4 days post infiltration and declined to basal level at 9 days post infiltration (Fig. (b)). In SAM, the MaFT expression level showed a continuous increase within 5 days post infiltration (Fig. (b)).
Fig. 4. Detection and quantification of GFP:MaFT by RT-PCR and quantitative RT-PCR (qRT-PCR) in mulberry.
Discussion
Agrobacterium-mediated transient expression assays could be used as useful tools to analyze gene function for plants once these transient expression assays are optimized.Citation29) In this study, we successfully optimized Agrobacterium-mediated transient expression assay in mulberry with regarding to the factors including infiltration method, A. tumerfaciens strain, and bacterial concentration. Hosein et al. have found that developmental stage of the leaf could influence the efficiency of Agrobacterium transient transformation.Citation30) Therefore, fully expanded young leaves with thick mesophylls from 2-month-old seedlings were adopted in our study to ensure the quality of the leaf material. A. tumerfaciens is an alphaproteobacterium, which has the capacity to integrate transferred DNA (T-DNA) from tumur-inducing plasmid (Ti plasmid) into chromosomal DNA of its host plant cells.Citation31) Modified A. tumerfaciens strains GV3101 and LBA4404 have been widely used as an efficient delivery system for genetic engineering in plants by cloning a desired gene sequence into the T-DNA.Citation32,33) In this study, the transient transformation efficiencies of these two A. tumerfaciens strains (GV3101 and LBA4404) were investigated, and the results indicated that A. tumefaciens strain GV3101 showed higher transformation efficiency than LBA4404. Moreover, the effect of bacterial concentration on the efficiency of transient transformation was also investigated and no obvious differences were observed among different bacterial concentrations (OD600: 0.5, 1.0 and 1.7). The bacterial concentrations of OD600 ranging from 0.2 to 1.5 have been reported in transient expression assays.Citation34–36) In this study, the concentration at the OD600 of 0.5 was selected for further experiments because it was the lowest concentration yielding consistent transient expression.
According to the optimization analysis, the gene function of MaFT was investigated with A. tumefaciens strain GV3101 harboring plasmid pBE2133:MaFT:GFP to infiltrate the leaves of mulberry seedlings by syringe method. The transient expressed MaFT:GFP protein was traced via detecting fused GFP signals. The GFP signals were observed in petiole and provasculature at the base of the SAM, indicating that MaFT:GFP fusion proteins expressed in the leaves were transmitted from the phloem companion cells to the meristem. What’s more, the transient expressions of MaFT:GFP in leaves and SAM were confirmed at mRNA level by using RT-PCR and qRT-PCR. It has been reported that photosynthate, mRNA, small RNA, and proteins are present in the phloem translocation stream of the angiosperms.Citation37) Therefore, these results indicated that the movement of MaFT:GFP protein and mRNA might be occurred through symplastic unloading from the phloem into the SAM regions of the mulberry seedlings. Corbesier et al. have found that FT fusion proteins expressed in leaf vascular tissue cells of Arabidopsis move to the SAM to trigger Arabidopsis flowering indicating FT expression may act as a mobile flower-inducing signal, such as the proposed ‘florigen’ signal.Citation38) Taken together, we could speculate that MaFT might play an important role in precocious flowering of mulberry through the movement of MaFT protein and mRNA from leaves to SAM to induce the flowering. However, the transient expressed MaFT was insufficient to induce flowering even though we conducted infiltrations in different sites of the same leaf. This result might be caused by the shorter life of transient expression (3–9 days) compared with contrast stable transformation. Thus, subsequent experiments are still needed to validate the above speculation.
In conclusion, a simple and efficient transient transformation assay could be implemented in leaves of mulberry seedlings by syringe infiltration using A. tumefaciens GV3101 at the concentration of OD600 = 0.5. This method might be useful for exploitation of multiple mulberry genomic resources and understanding of mulberry biology. Moreover, we also found that the MaFT might contribute to the precocious flowering of mulberry due to the movement of MaFT from leaves to SAM.
Acknowledgment
We thank Prof Mitsuhara Ichiro (National Institute of Agrobiological Resources, Japan) for providing pBE2133 vectors.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Ionescu-Tîrgovişte C, Popa E, Mirodon Z, Simionescu M, Mincu I. The effect of a plant mixture on the metabolic equilibrium in patients with type-2 diabetes mellitus. Rev Med Interna Neurol Psihiatr Neurochir Dermatovenerol Med Interna. 1989;41:185–192.
- Zhu Y-P. Chinese materia medica: chemistry, pharmacology and applications. Boca Raton: CRC Press; 1998.
- He N, Zhang C, Qi X, Zhao S, Tao Y, Yang G, Lee TH, Wang X, Cai Q, Li D, Lu M, Liao S, Luo G, He R, Tan X, Xu Y, Li T, Zhao A, Jia L, Fu Q, Zeng Q, Gao C, Ma B, Liang J, Shang J, Song P, Wu H, Fan L, Wang Q, Shuai Q, Zhu J, Wei C, Zhu-Salzman K, Jin D, Wang J, Liu T, Yu M, Tang C, Wang Z, Dai F, Chen J, Liu Y, Lin T, Zhang S, Yang H, Paterson AH, Xia Q, Ji D, Xiang Z. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013;4:1–9.
- Ow DW, De Wet JR, Helinski DR, Howell SH, Wood KV, Deluca M. Transient and stable expression of the firefly luciferase gene in plant cells and transgenic plants. Science. 1986;234:856–859.10.1126/science.234.4778.856
- Li J, Krichevsky A, Vaidya M, Tzfira T, Citovsky V. Uncoupling of the functions of the Arabidopsis VIP1 protein in transient and stable plant genetic transformation by Agrobacterium. Proc. Nat. Acad. Sci. 2005;102:5733–5738.10.1073/pnas.0404118102
- Klein I, Sanchez-Alavez M, Tabarean I, Schaefer J, Holmberg KH, Klaus J, Xia F, Marcondes MC, Dubins JS, Morrison B, Zhukov V, Sanchez-Gonzalez A, Mitsukawa K, Hadcock JR, Bartfai T, Conti B. AdipoR1 and 2 are expressed on warm sensitive neurons of the hypothalamic preoptic area and contribute to central hyperthermic effects of adiponectin. Brain Res. 2011;1423:1–9.10.1016/j.brainres.2011.09.019
- Zhang Y, Su J, Duan S, Ao Y, Dai J, Liu J, Wang P, Li Y, Liu B, Feng D, Wang J, Wang H. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes. Plant Methods. 2011;7:1–14.10.1186/1746-4811-7-30
- Rasco-Gaunt S, Riley A, Barcelo P, Lazzeri PA. Analysis of particle bombardment parameters to optimise DNA delivery into wheat tissues. Plant Cell Rep. 1999;19:118–127.10.1007/s002990050721
- Orzaez D, Mirabel S, Wieland WH, Granell A. Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit. Plant Physiol. Behav. 2006;140:3–11.
- Bhaskar PB, Venkateshwaran M, Wu L, Ane JM, Jiang J. Agrobacterium-mediated transient gene expression and silencing: a rapid tool for functional gene assay in potato. PLoS One. 2009;4:e5812.
- Schöb H, Kunz C, Meins Jr. F. Silencing of transgenes introduced into leaves by agroinfiltration: a simple, rapid method for investigating sequence requirements for gene silencing. Mol. Gen. Genet. 1997;256:581–585.
- Marion J, Bach L, Bellec Y, Meyer C, Gissot L, Faure JD. Systematic analysis of protein subcellular localization and interaction using high-throughput transient transformation of Arabidopsis seedlings. Plant J. 2008;56:169–179.10.1111/tpj.2008.56.issue-1
- Zottini M, Barizza E, Costa A, Formentin E, Ruberti C, Carimi F. Agroinfiltration of grapevine leaves for fast transient assays of gene expression and for long-term production of stable transformed cells. Plant Cell Rep. 2008;27:845–853.10.1007/s00299-008-0510-4
- Chen X, Equi R, Baxter H, Berk K, Han J, Agarwal S, Zale J. A high-throughput transient gene expression system for switchgrass (Panicum virgatum L.) seedlings. Biotechnol. Biofuels. 2010;3:1–10.10.1186/1754-6834-3-9
- Nowak K, Luniak N, Meyer S, Schulze J, Mendel R, Hänsch R. Fluorescent proteins in poplar: a useful tool to study promoter function and protein localization. Plant Biol. (Stuttg). 2004;6:65–73.
- Zhang C, Gai Y, Zhu Y, Chen X, Jiang X. Construction of a bidirectional promoter and its transient expression in Populus tomentosa. Front. For. China. 2008;3:112–116.10.1007/s11461-008-0018-7
- Araki T. Transition from vegetative to reproductive phase. Curr. Opin. Plant Biol. 2001;4:63–68.10.1016/S1369-5266(00)00137-0
- Pin PA, Nilsson O. The multifaceted roles of FLOWERING LOCUS T in plant development. Plant, Cell Environ. 2012;35:1742–1755.10.1111/pce.2012.35.issue-10
- Lifschitz E, Eviatar T, Rozman A, Shalit A, Goldshmidt A, Amsellem Z, Alvarez JP, Eshed Y. The tomato FT ortholog triggers systemic signals that regulate growth and flowering and substitute for diverse environmental stimuli. Proc. Nat. Acad. Sci. 2006;103:6398–6403.10.1073/pnas.0601620103
- Endo T, Shimada T, Fujii H, Kobayashi Y, Araki T, Omura M. Ectopic expression of an FT homolog from Citrus confers an early flowering phenotype on trifoliate orange (Poncirus trifoliata L. Raf.). Transgenic Res. 2005;14:703–712.10.1007/s11248-005-6632-3
- Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, Yano M. Hd3a, a rice ortholog of the Arabidopsis FT Gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002;43:1096–1105.10.1093/pcp/pcf156
- Bohlenius H, Huang T, Charbonnel-Campaa L, Brunner AM, Jansson S, Strauss SH, Nilsson O. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science. 2006;312:1040–1043.10.1126/science.1126038
- Hecht V, Laurie RE, Vander Schoor JK, Ridge S, Knowles CL, Liew LC. The pea GIGAS gene is a FLOWERING LOCUS T homolog necessary for graft-transmissible specification of flowering but not for responsiveness to photoperiod. Plant Cell. 2011; 23:147–161.10.1105/tpc.110.081042
- Lin MK, Belanger H, Lee YJ, Varkonyi-Gasic E, Taoka KI, Miura E, Xoconostle-Cázares B. FLOWERING LOCUS T protein may act as the long-distance florigenic signal in the cucurbits. Plant cell. 19:1488–1506.
- Koncz C, Schell J. The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimaeric genes carried by a novel type of Agrobacterium binary vector. MGG Mol. Gen. Genet. 1986;204:383–396.10.1007/BF00331014
- Sparkes IA, Runions J, Kearns A, Hawes C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006;1:2019–2025.10.1038/nprot.2006.286
- D’aoust M-A, Lavoie P-O, Belles-Isles J, Bechtold N, Martel M, Vézina L-P, Transient expression of antibodies in plants using syringe agroinfiltration. In: Faye L, Gomord V, editors. Recombinant proteins from plants. Clifton, NJ: Springer; 2009. p. 41–50.
- Jefferson R. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Mol. Biol. Rep. 1987;5:387–405.10.1007/BF02667740
- Manavella PA, Chan RL. Transient transformation of sunflower leaf discs via an Agrobacterium-mediated method: applications for gene expression and silencing studies. Nat. Protoc. 2009;4:1699–1707.10.1038/nprot.2009.178
- Hosein FN, Lennon AM, Umaharan P. Optimization of an Agrobacterium-mediated transient assay for gene expression studies in Anthurium andraeanum. J. Am. Soc. Hortic. Sci. 2012;137:263–272.
- lker B, Li Y, Rosso MG, Logemann E, Somssich IE, Weisshaar B. T-DNA–mediated transfer of Agrobacterium tumefaciens chromosomal DNA into plants. Nat. Biotechnol. 2008;26:1015–1017.
- Schell J, Montagu Van M. The Ti-plasmid of Agrobacterium tumefaciens, a natural vector for the introduction of nif genes in plants? In: Hollaender A, Burris RH, Day PR, Hardy RWF, Helinski DR, Lamborg MR, Owens L, Valentine RC, editors. Genetic engineering for nitrogen fixation. USA: Springer; 1977. p. 159–179.
- Zambryski P, Joos H, Genetello C, Leemans J, Van Montagu M, Schell J. Ti plasmid vector for the introduction of DNA into plant cells without alteration of their normal regeneration capacity. EMBO J. 1983;2:2143–2150.
- Bhaskar PB, Venkateshwaran M, Wu L, Ané J-M, Jiang J. Agrobacterium-mediated transient gene expression and silencing: a rapid tool for functional gene assay in potato. PLoS One. 2009;4:e5812.10.1371/journal.pone.0005812
- Voinnet O, Rivas S, Mestre P, Baulcombe D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003;33:949–956.10.1046/j.1365-313X.2003.01676.x
- Wydro M, Kozubek E, Lehmann P. Optimization of transient Agrobacterium-mediated gene expression system in leaves of Nicotiana benthamiana. Acta Biochim. Pol. 2006;53:289–298.
- Lough TJ, Lucas WJ. INTEGRATIVE PLANT BIOLOGY: role of phloem long-distance macromolecular trafficking. Annu. Rev. Plant Biol. 2006;57:203–232.10.1146/annurev.arplant.56.032604.144145
- Corbesier L, Vincent C, Jang S, Fornara F, Fan Q, Searle, I, Giakountis A, Farrona S, Gissot L, Turnbull C, Coupland G. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science. 2007;316:1030–1033.10.1126/science.1141752