
ABSTRACT
Herpes simplex virus, varicella zoster virus, human cytomegalovirus, and rubella virus are the most common causes of virus-induced anterior uveitis. They can present in a variety of entities not only with typical but also overlapping clinical characteristics. These viral infections are commonly associated with ocular infiltration of T cells and B/plasma cells, and expression of cytokines and chemokines typical of a proinflammatory immune response. The infections differ in that the herpes viruses cause an acute lytic infection and inflammation, whereas rubella virus is a chronic low-grade infection with slowly progressing immunopathological responses. The outcome of an intraocular viral infection may largely be guided by the characteristics of the virus, which subsequently dictates the severity and type of the immune response, and the host immune status.
Virus-induced anterior uveitis is a combination of a viral infection and resulting antiviral inflammatory responses in the eye. In general, the virus will infect permissive ocular cells, which may differ for every virus. This will induce an innate immune response including the release of cytokines and chemokines, which will subsequently attract immune cells, such as neutrophils, natural killer (NK) cells, T and B cells, which will further contribute to the inflammation by for instance secreting more inflammatory cytokines and chemokines and/or by exerting cytotoxicity toward virus-infected cells. On the other hand anti-inflammatory cytokines and regulatory T cells may be present, which presumably counteract the inflammation. Depending on the characteristics of the virus, the immune status and most likely also the genetic make-up of the patient the resulting uveitis may vary in severity and outcome. The most prevalent causes of virus-induced anterior uveitis are herpes simplex virus (HSV), varicella zoster virus (VZV), human cytomegalovirus (HCMV), and rubella virus.Citation1 Other viruses are also known to cause or are associated with anterior uveitis, such as chickungunya virus, dengue virus, influenza virus H1N1, parechovirus, and parvovirus B19;Citation2–Citation5 however, immunopathology in relation to these viruses is reported only limitedly. This article will therefore focus on the main viral causes: HSV, VZV, HCMV, and rubella virus.
HERPETIC ANTERIOR UVEITIS
The human neurotropic alphaherpesviruses, HSV type 1 (HSV-1) and particularly VZV, are the most common infectious causes of anterior uveitis in otherwise healthy individuals, accounting for 5–10% of cases in the Western world.Citation6,Citation7 Herpetic anterior uveitis (HAU) usually follows an acute recurrent course, is typically unilateral, and associated with blurred vision, eye pain, photophobia, redness, the presence of keratic precipitates, elevated intraocular pressure (IOP), and sectoral iris atrophy.Citation7–Citation9 Compared to HSV-1-associated HAU, VZV anterior uveitis is more commonly diagnosed in older individuals and more frequently associated with conjunctival redness, corneal edema, posterior synechiae, vitritis, and higher IOP.Citation8–Citation10 The visual prognosis of HAU, when diagnosed promptly and if treated correctly with specific antiviral therapy and topical steroids, is favorable.Citation8,Citation9 Because no specific clinical parameters are known to differentiate HSV-1 and VZV anterior uveitis, diagnosis of HAU is challenging particularly when patients show no typical clinical signs or recent history of herpetic diseases like HSV-1 keratitis and herpes zoster ophthalmicus.Citation6–Citation9 Clinical examination of aqueous humor (AH), showing cellular activity and flare, and laboratory analysis of AH to detect herpesvirus-specific DNA and local antibody production in the diseased eye are advocated to determine the severity and etiology of anterior uveitis, respectively.Citation8,Citation11,Citation12 Because the patient’s immune status and duration of disease affect the outcome of either diagnostic assay, both assays should ideally be performed in parallel.Citation7–Citation9,Citation11 Whereas early at onset of disease viral DNA is readily detectable and local antibody absent, the reverse outcome is more common when AH is sampled at later times after onset of disease. Furthermore, local antibody production is rarely contributive to the differential diagnosis in severely immunocompromised uveitis patients.Citation8,Citation11,Citation12
While the majority of adults are latently infected with HSV-1 and VZV, only a limited number of latently infected individuals will develop HAU. Various factors that determine the incidence and spectrum of ocular herpetic diseases are immune status, age, genetic predisposition, time and route of infection, and potentially the virus strain.Citation6–Citation8,Citation13 HSV-1 and VZV are endemic worldwide infecting about 70% and >90% of individuals, respectively.Citation14 Upon primary infection, commonly during early childhood, both viruses establish a lifelong latent infection in sensory and autonomic neurons from which they reactivate leading to regular asymptomatic virus shedding and incidentally recurrent disease of variable clinical severity ranging from cold sores and shingles to potentially sight- (keratitis and uveitis: both viruses) or even life-threatening neurological diseases (HSV-1 encephalitis).Citation14,Citation15 Intra-ganglionic innate (neuron-interacting satellite glial cells) and adaptive immunity (T cells) are essential to control herpesvirus latency in neurons. Whereas innate immunity is most likely sufficient to prevent VZV to escape from latency, HSV-1-specific CD8 T cells are of additional importance to prevent HSV-1 reactivation.Citation14,Citation16–Citation18 Because most if not all HAU patients have already antibodies to the triggering herpesvirus and latency has not been detected in ocular tissues,Citation11,Citation19 the disease is not initiate by a primary infection but most likely due to reactivation of endogenous virus that entered the eye via trans-axonal transport from the innervating sensory or autonomic ganglia.Citation7,Citation10 The independent distribution of latent HSV-1 and VZV in these ganglia, particularly the dissimilarity in latent virus between paired left and right ganglia of the human head and neck, may explain the clinical observation that HAU usually occurs unilaterally.Citation20,Citation21
Due to the lack of animal models that appropriately mimic the pathogenesis of HAU, especially VZV that does not productively infect nonhuman cells, current knowledge on the virus and host factors involved in the disease process is largely based on laboratory studies performed on surplus AH samples of HAU patients.Citation7–Citation10,Citation14 Whereas these studies provided important insight into the cell types and mediators involved during fulminant HAU, inevitably no information was obtained on key parameters that coordinate the initiation and resolution phase of HAU. This information is at least of equal importance to develop novel intervention strategies aimed to block disease progression and prevent recurrences.Citation8,Citation10,Citation14 The association between timing of AH sampling and the differential presence of herpesvirus DNA and local antibody productionCitation11 hint at the prominent role of a direct cytopathic effect (CPE) of the virus on ocular tissue resident cells and an immune-mediated pathology during the early and more progressed clinical phase of HAU, respectively. The correlation between intraocular VZV load and IOP may be due to an obstruction of aqueous outflow due to viral CPE on the trabecular meshwork, whereas the beneficial effect of corticosteroid treatment supports the detrimental role of a local inflammatory response in HAU pathogenesis.Citation8,Citation10,Citation14 Anterior chamber inflammation in HAU can vary from mild-to-severe inflammation ultimately leading to hypopyon formation, which represents a dense leukocyte sediment at the bottom of the anterior chamber. AH of HAU patients is devoid of neutrophils and predominantly contains T cells. Compared to paired blood, AH-derived T cells are predominantly memory CD8 T cells expressing the activation marker Human leukocyte antigen-DR (HLA-DR) implicating their direct role in HAU pathogenesis.Citation22 Detailed phenotypic and functional studies on short-term cultures of AH-derived T cells of HAU patients demonstrated that they specifically recognized the triggering herpesvirus.Citation23 These virus-specific T cells, both CD4 and CD8 T cells, expressed a T0/T1 phenotype and killed virus-infected cells in vitro demonstrating their potential uveitogenic properties.Citation23,Citation24 These data match the cytokine expression profiles in AH of HAU patients, implicating that T cells are the major source of tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ), antiviral cytokines that can also mediate deleterious inflammatory responses.Citation25,Citation26 In addition to inflammatory cytokines, interleukin 10 (IL-10) levels are also elevated in AH from infectious uveitis patients. The significance and cellular source of this inhibitory cytokine in HAU patients is unclear.Citation25,Citation26 T-cell reactivity to the non-causative human alphaherpesvirus is also detected in the affected eyes of herpetic uveitis patients,Citation23 suggesting the role of cross-reactive T cells in the disease. The identification of CD4 T-cell clones, cultured from AH of a VZV uveitis patient, which recognized both VZV- and HSV-1-infected cells, supports this hypothesis.Citation27 Moreover, the respective VZV/HSV-1 cross-reactive CD4 T cell epitope bound with high affinity to multiple HLA-DR alleles and was readily recognized by blood-derived T cells of multiple healthy adults. These type of herpesvirus cross-reactive CD4 T cells may be beneficial to combat HSV-1 and VZV infection in skin but may be detrimental upon infiltration and activation in ocular tissues infected with either herpesvirus.Citation27 In conclusion, HSV-1 and VZV are common causes of anterior uveitis with a good prognosis if treated appropriately to control both intraocular virus replication and virus-specific T cell-mediated immunopathology.
CMV ANTERIOR UVEITIS
HCMV is a double-stranded DNA virus and is a member of the Herpesviridae family.Citation28–Citation30 Productive HCMV ocular infections are more commonly present as a retinitis in immunocompromised patientsCitation31,Citation32 but are increasingly recognized as a cause of anterior uveitis, corneal endotheliitis, and corneal graft failure in immunocompetent individuals.Citation29,Citation33–Citation37 Chronic CMV anterior uveitis may present as a Fuchs heterochromic cyclitis or with acute IOP spikes similar to Posner–Schlossman syndrome (PSS).Citation38,Citation39 During such IOP spikes, studies have shown that CMV DNA can be detected and quantified by real-time polymerase chain reaction analysis, indicating that the reactivation of the CMV infection can be correlated to clinical inflammation and IOP elevation.Citation36 However, these studies also demonstrate the transient nature of this reactivation which is demonstrated by false negative results due to delayed testing.Citation36
HCMV is capable of infecting a variety of cell types including ocular cells, such as retinal cells, keratocytes, glial, and inflammatory cells.Citation28–Citation32,Citation40 After initial infection, the HCMV genome enters the cell nucleus and leads to a variety of viral gene transcriptions from immediate early genes to late genes resulting in production of infectious particles.Citation28,Citation30,Citation41,Citation42 The outcome of HCMV replication leads to lytic HCMV infection and depending on the cell type that is infected can result in lifelong latency. Latent infection is known to establish in monocytes and CD34+ cells of myeloid lineage.Citation28,Citation30,Citation42,Citation43 Both the innate and adaptive immunity play a role in the host defense against HCMV infections.
Toll-like receptors (TLRs) are components of the innate immunity and TLR2 has been shown to interact specifically with the HCMV glycoproteins B and gH resulting in activation of the innate immune system.Citation30 In murine CMV infection, TLR4 and 9 were also shown to be involved.Citation44 Once the innate response is activated, inflammatory cytokines and interferons are produced and co-stimulatory molecules are upregulated. Antigen-presenting cells (APCs) and NK cells are also activated to prime the adaptive immunity.Citation28,Citation30,Citation41 NK cells are thought to bridge the innate and adaptive immune system and their role in CMV infection is increasingly being recognized.Citation30
HCMV-derived peptides are processed by APCs and result in a strong adaptive immune response involving both humoral and cellular immunity.Citation30,Citation41 The main cellular mediators against HCMV infections are CD4 and CD8 T cells which recognize peptides presented by MHC Class II and Class I molecules, respectively.Citation41 Once activated, they proliferate and differentiate to effector T cells that secrete cytokines and may destroy the CMV virus-infected cells.Citation30 The humoral immune response is characterized by the formation of specific antibodies to HCMV structural and nonstructural viral proteins, which contribute to restriction of viral dissemination and its clinical manifestations. In immunocompetent individuals, the adaptive immunity is important for control of HCMV latency aimed to prevent reactivation. During CMV latency, CMV-specific T cells persist as memory T cells that continue to expand and become activated to production interferon-gamma (IFN-γ) and a Th1 cytokine signature upon viral reactivation. In individuals with impaired T-cell immunity, such as in retrovirus infection (AIDS) and solid organ or bone marrow transplantation, the risk of HCMV reactivation is increased due to T-cell impairment and with that the clinical manifestation of HCMV retinitis rather than anterior uveitis or endotheliitis.Citation30,Citation41
Intraocular CMV infections have been studied in mice using MCMV.Citation43,Citation45–Citation47 In immunocompetent mice, MCMV intraocular infection presents as an anterior uveitis characterized by inflammatory cells within the iris and ciliary body without accompanying retinitis.Citation43,Citation45,Citation46 In these cases, MCMV DNA was detected by in situ hybridization in the cornea, ciliary body, and iris early after viral infection, whereas only occasional viral DNA was detected in the choroid and transiently in the retina.Citation43,Citation45,Citation46 MCMV retinitis did not develop until immunosuppression was instigated.Citation45 In systemic MCMV infection, the virus was detected in the anterior segment less frequently than with intraocular inoculation or immunosuppression.Citation47 These data suggest that other factors, such as blood retinal barrier breakdown and altered immune responses, are necessary for a productive ocular MCMV infection. In contrast, when immunosuppression was induced in immunocompetent MCMV-infected mice, necrotizing retinitis developed suggesting that the route of MCMV retinal infection was via the ciliary body and choroid.Citation45 The presence of MCMV DNA in the retinal pigment epithelium and retinal outer nuclear layers in early infections supports this theory.Citation45,Citation46 In addition, in immunosuppressed mice, corneal keratocytes appeared to be infected via limbal vessels but this did not cause clinical corneal pathology.Citation47
Macrophages and its precursor monocytes are thought to harbor MCMV in systemic infection and aid spread to other organs, yet these murine studies also revealed that when macrophages were depleted, viral dissemination still occurred demonstrating that it was not macrophage dependent.Citation47 As previously mentioned, T and B cells have also been shown to harbor MCMV and can be a potential source of latency.Citation43 It is interesting to note that MCMV DNA could be detected in ocular fluids and tissues, particularly in the ciliary body and iris, as long as 3–4 months post-infection and suggests that the anterior uvea may represent a potential reservoir for latent MCMV.Citation45,Citation46
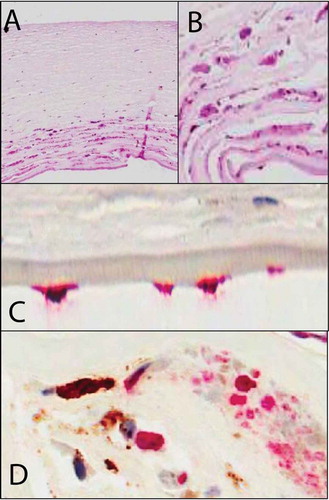
Although there is a paucity of knowledge in the pathogenesis of anterior CMV uveitis in immunocompetent individuals, we are beginning to find similarities to the MCVM model.Citation45,Citation46 HCMV-associated owl’s eye inclusions have previously been demonstrated in the cornea stroma and endothelium in patients with HCMV endotheliitis by in vivo confocal microscopy as well as HCMV DNA by real-time polymerase chain reaction (qPCR) in anterior chamber fluid.Citation33,Citation34 Moreover, a recent study by Chan et al. demonstrated HCMV viral protein expression in the cornea and iris that appears to support the MCVM anterior uvea latency theory.Citation29 In HCMV graft-related infections, CMV-antigen-positive endothelial cells were found by immunohistochemistry (), confirming the presence of an underlying HCMV-induced endotheliitis.Citation29 Furthermore, diffuse stromal involvement characterized by HCMV-infected keratocytes was demonstrated (. The location of the HCMV infection in the cornea seen at the limbusCitation29 seems to reflect the findings in the MCMV immunosuppression model.Citation47 Indeed, the majority of the patients with corneal HCMV endotheliitis or keratitis have had local or systemic immunosuppressive therapy. Similar to HCMV retinal studies, co-localization of HCMV-infected cells with CD68-positive macrophages was not found.Citation29 Finally, CD163 type 2 macrophages showed close association with CMV-infected keratocytes suggesting that rather than harboring the virus, these macrophages may play a role in the immunopathology of CMV graft infection ().
FIGURE 1. Immunohistochemistry on corneal tissue removed during penetrating keratoplasy from patients with CMV graft infection. (A,B) Histology (hematoxylin and eosin stain 20×, 40×): Viral cytopathic changes are characterized by cellular enlargement (up to 2 times a normal keratocyte) with eosinophilic intranuclear and intracytoplasmic inclusions. Note the significant lack of inflammation and vascularisation as seen in other herpetic keratitis. (C) CMV immunohistochemistry, 40× (Dako, USA with alkaline phosphatase red chromagen) demonstrating CMV antibody staining in the endothelial cells confirming the presence of CMV endotheliitis. (D) Dual CMV and CD163 macrophage (Dako, USA) immunohistochemistry (40×) with alkaline phosphatase red chromagen, CMV antibody, and brown diaminobenzidine staining for CD163 shows no colocaliztion of CD163 with CMV demonstrating that CMV is not latent in macrophages.
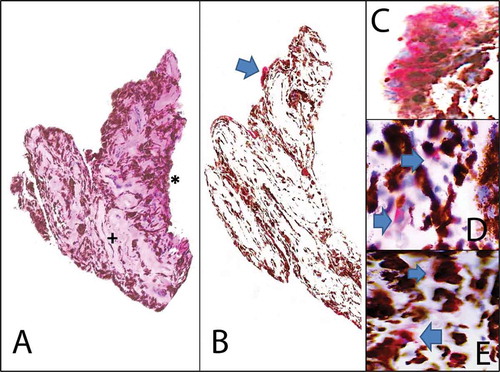
Chronic HCMV uveitis is associated with iris stromal atrophy.Citation36,Citation37 Iris iridectomy specimens from patients with previous qPCR-positive HCMV anterior uveitis show stromal atrophy with a relatively intact posterior iris pigment epithelium (A; Chan et al., unpublished data). Immunohistochemical (IHC) analysis for HCMV antigens showed variable atypical staining in some iris pigment epithelial cells characterized by only cytoplasmic staining (, unlike active HCMV-positive controls where both nuclear and cytoplasmic immunoreactivity were observed (D). HCMV-specific qPCR of these irides is commonly negative. Light microscopy also does not show the typical cytomegaly of CMV CPE. The clinical significance of such staining pattern has yet to be explored. A possible explanation might be cross-reactivity of HCMV proteins with iris proteins. Alternatively, it may be due to staining of viral protein remnants in the cytoplasm. The question whether HCMV DNA can be eradicated post-infection, but not the CMV protein, remains unanswered but is of interest for future studies. Immunohistochemistry for other inflammatory cell markers in these post-CMV irides showed incidental CD3 T cells and CD68 macrophage infiltrates suggesting the presence of a subclinical chronic inflammatory response (D and E).
FIGURE 2. Immunohistochemistry on iris iridectomy specimens from patients with chronic CMV anterior uveitis. (A) Histology (hematoxylin and eosin stain 20×): Iris iridectomy specimen showing stroma fibrosis and atrophy (+). The posterior iris pigmented epithelium remains relatively intact (*). Viral cytopathic changes characterized by cytomegalic cells with eosinophilic nuclear and cytoplasmic inclusions are absent. Rare inflammatory cells may be seen within the stroma. (B) Low power view of CMV immunohistochemistry (20×, alkaline phosphatase red chromagen) highlights the atypical CMV antibody in occasional cells (blue arrow). (C) Higher power view of CMV immunohistochemistry (40×, alkaline phosphatase red chromagen) reveals that the CMV antibody staining is limited to the cytoplasm (blue arrows) only without the nuclear staining seen in the positive controls. (D) CD3, T-cell immunohistochemistry (40×) with alkaline phosphatase red chromagen highlights the presence of rare T cells (blue arrow). (E) CD68, macrophage immunohistochemistry (40×) with alkaline phosphatase red chromagen highlights the presence of rare macrophages (blue arrow).
Little data are available on intraocular cytokines in CMV anterior uveitis. The AH of immunocompetent patients with HCMV anterior segment infections was reported to have significantly higher levels of IL-5, IL-6, IL-8 (CXCL8), monocyte chemotactic protein-1 (MCP-1;CCL-2), macrophage inhibitory protein-1β (MIP-1β; CCL-4), granulocyte colony stimulating factor, and transforming growth factor-β, but low levels of IL-2, IL-12, TNF-α, and INF-γ compared to the control groups.Citation48,Citation49 Interestingly, no significant difference in aqueous cytokines was detected in HCMV qPCR-positive immunocompetent PSS patients when compared to HCMV qPCR-negative PSS patients.Citation49 Future studies are warranted to provide better insights into the inflammatory response or intraocular milieu that may potentiate HCMV anterior segment infection.
In conclusion, HCMV infection induces a profound immune response involving both innate and adaptive immunity. Impairment of both, and in particularly the virus-specific T-cell response, can induce HCMV reactivation. In immunocompetent mice and humans, CMV presents a transient, self-limiting viral infection that predominantly occurs in the anterior uvea presenting as an anterior uveitis or corneal endotheliitis. In latent or chronic CMV infections, viral DNA has been detected in the anterior uvea of murine CMV models, even several months after initial infection suggesting that it may be a potential reservoir of latent CMV. An intraocular immunocompromised state may result in the reactivation of CMV. Case reports have described HCMV reactivation in patients using topical cyclosporine A or with ocular corticosteroid implants.Citation50,Citation51 Both of these drugs are known to suppress T-cell immunity, which is crucial in inhibiting CMV reactivation.Citation52,Citation53 Understanding the immunopathogenesis of CMV ocular infections is vital in the development of future diagnostic and therapeutic strategies.
RUBELLA VIRUS-INDUCED UVEITIS (FUCHS UVEITIS)
Rubella virus is a positive-stranded RNA virus and the only member of the Rubivirus genus within the Togavirus family. It is the causative agent of rubella and congential rubella syndrome.Citation54 Only in 2004, rubella virus was identified as an etiological agent of Fuchs uveitis syndrome (FUS).Citation55 Hence, few reports are available on the immunology and pathology in Fuchs uveitis (FU) patients with a confirmed intraocular rubella virus infection. Therefore, this part will focus mostly on the immunopathology of FU in general. Do note, however, that HCMV and more recently chickungunya virus have also been reported to be associated with FUS-like clinical entities and that in most reports on FUS, it is not clear which virus is involved in the population described.Citation2,Citation37,Citation56
Striking immunopathological features of FUS are iris atrophy/heterochromia and the stellate keratic precipitates and vitritis.Citation9,Citation57 Electronmicroscopical (EM) and IHC studies on iris biopsies of FUS patients revealed the abundant presence of plasma cells and other lymphocytes, abnormal or degraded melanocytes, the absence of stromal melanocytes, stromal atrophy, and depigmentation of the posterior pigment epithelium consistent with the described atrophy and/or heterochromia.Citation58–Citation62 Indeed, in longitudinal studies, gradual degeneration of the stroma and posterior pigmented epithelium resulting in the clear blue iris, typical of the diseased eye in bright eyed FUS patients, has been described as well.Citation63,Citation64 IHC analysis on iris and cornea biopsies also showed mononuclear cell and particularly plasma cell infiltration in Koeppe nodules and keratic precipitates, respectively.Citation58,Citation62 Although it is not known whether the patients investigated in these studies had rubella virus-associated FUS, the presence of plasma cells would be in agreement with the high prevalence of intraocular antibody production against rubella virus.Citation55,Citation65–Citation68 Unfortunately, the magnification of the EM pictures was too low to observe rubella virus particles should these have been present.
In addition to plasma cells, T cells were found in the anterior chamber fluid of FUS patients.Citation60,Citation69,Citation70 Muhaya et al. described the presence of both CD4 and CD8 T cells in the AH of FUS patients. Both T-cell subsets were present at higher frequencies than in paired peripheral blood. Moreover, there seemed to be a shift toward CD8 T cells, which was also observed by Laballette et al.Citation69,Citation70 Unfortunately, the distinction between cytotoxic T cells and regulatory CD8 T cells was not made. Both IFN-γ and IL-10 expression were found in the AH of the FUS patients, suggesting that both CD8 T-cell subsets might be present. Laballette et al. investigated the clonal composition of intraocular CD8 T cells of two FUS patients by analyzing the expression of T-cell receptor variable beta (TCRVβ) genes.Citation69 The authors found a limited number of TCRVβ clonotypes in both patients suggestive of an antigen-driven process, though it was not determined whether this intraocular T-cell response was directed to an infectious agent, such as rubella virus, or self-antigens or both.
Whereas no clear genetic associations with specific HLA alleles have been described for FUS, Spriewald et al. reported an association between polymorphisms in the cytotoxic T-lymphocyte antigen 4 (CTLA4) gene.Citation57,Citation71 However, these data have to be corroborated in a larger patient cohort.
In intraocular fluid of FUS patients, expression of several, mostly inflammatory, cytokines, and chemokines has been reported, including IL-6, IL-8, IL-12, IL-15, IP-10, MCP-1, TNF-α, INF-γ, and MIP-1α. Interestingly, with the exception of IL-12, expression of most inflammatory immune mediators was decreased compared to that in HSV-1 and VZV-induced posterior (acute retina necrosis) and anterior uveitis, reflecting the low-grade inflammation in FUS eyes.Citation25,Citation72 Similarly, de Visser et al. reported a surprising similarity between chemo-/cytokine expression levels in AH of FUS and ocular toxoplasmosis patients, apparently representative of a milder inflammatory pathogenesis.Citation72 Anti-inflammatory cytokines, such as TGF-β and IL-10, have been detected in FUS AH as well, though IL-10 expression in AH was generally low and not significantly different compared to non-inflammatory controls.Citation13,Citation25,Citation70,Citation72 Suzuki et al., however, found high IL-10 expression levels in the vitreous of two FUS patients.Citation13 Since there are no other reports on cytokine expression in vitreous of FUS patients, the implication of these discrepant findings is not clear yet.
In conclusion, the immunopathology of FUS is characterized by a low-grade intraocular antigen-driven inflammation with T-cell and plasma cell infiltration and mild inflammatory immune mediator expression, with currently no apparent genetic associations. In case of rubella virus, one could envisage a pathogenesis where a slow viral infection results in gradual degradation of the anterior layer, stroma, and posterior pigmented epithelium of the iris. Furthermore, in reaction to the viral infection, plasma cells, T cells, and monocytes/macrophages infiltrate in the eye, which in combination with cytokine and chemokine expression also contribute to tissue damage to the iris and to immune depositions on the cornea and iris.
SUMMARY
HSV, VZV, CMV, and rubella virus are the major etiologies of virus-induced anterior uveitis and together cause a variety of anterior uveitis manifestations which may not only have specific characteristics but may also overlap, despite the fact that each virus has its own distinctive pathogenic properties.Citation9,Citation37 Whereas active HSV/VZV and CMV AU are characterized by acute lytic infections, rubella virus-related AU is slowly progressing and does not seem to cause immediate virus-related tissue damage. HSV and VZV are self-limiting infections that establish a lifelong latent infection in neurons of ganglia that are kept under control by both innate and adaptive immune responses. If immune control fails, the endogenous latent virus reactivates and may cause a variety of recrudescent ocular diseases including AU. CMV infection is also self-limiting; however, this virus is not neurotropic but seems to become latent in ocular-resident cells, although a low-grade persistent infection cannot be ruled out. If left untreated, HCMV can cause frequent intermittent recurrences. Rubella virus infection on the other hand does not seem to cease and may represent a long-lasting persistent low-grade infection ultimately culminating in the typical ophthalmoclinical manifestations of FUS probably as a result of continuous immunological activity. From an immunological perspective, ocular infection with the aforementioned viruses results in the influx of T and B cells rather than neutrophils, and a proinflammatory immune mediator profile, though the anti-inflammatory cytokine IL-10 is also expressed, possibly to counteract the inflammation. The combined knowledge suggests that the outcome of an intraocular viral infection is largely guided by the characteristics of the virus, which subsequently dictates the severity and type of the immune response. The immunological conditions for intraocular viral control and reactivation or for prolonged inflammation remain subject of investigation.
DECLARATION OF INTEREST
The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Groen-Hakan F, Babu K, Tugal-Tutkun I, et al. Challenges of diagnosing viral anterior uveitis. Ocul Immunol Inflam. 2017;25:715–725.
- Babu K, Murthy GJ. Chikungunya virus iridocyclitis in Fuchs’ heterochromic iridocyclitis. Indian J Ophthalmol. 2012;60:73–74.
- Jdf DG-M, De Visser L, Zuurveen S, et al. Identification of new pathogens in the intraocular fluid of patients with uveitis. Am J Ophthalmol. 2010;150:628–636.
- Jdf DG-M, Dekkers J, De Visser L, Rothova A, Van Loon AM, De Boer JH. Antibody production against b19 virus in ocular fluid of jia-associated uveitis patients. Ophthalmology. 2015;122:1270–1272
- Khairallah M, Kahloun R. Ocular manifestations of emerging infectious diseases. Curr Opin Ophthalmol. 2013;24:574–580.
- Jakob E, Reuland MS, Mackensen F, et al. Uveitis subtypes in a german interdisciplinary uveitis center–analysis of 1916 patients. J Rheumatol. 2009;36:127–136.
- Jap A, Chee SP. Viral anterior uveitis. Curr Opin Ophthalmol. 2011;22:483–488.
- Pleyer U, Chee SP. Current aspects on the management of viral uveitis in immunocompetent individuals. Clin Ophthalmol. 2015;9:1017–1028.
- Wensing B, Relvas LM, Caspers LE, et al. Comparison of rubella virus- and herpes virus-associated anterior uveitis: clinical manifestations and visual prognosis. Ophthalmology. 2011;118:1905–1910.
- Takase H, Kubono R, Terada Y, et al. Comparison of the ocular characteristics of anterior uveitis caused by herpes simplex virus, varicella-zoster virus, and cytomegalovirus. Jpn J Ophthalmol. 2014;58:473–482.
- Jdf DG-M, Rothova A, Van Loon AM, et al. Polymerase chain reaction and Goldmann-Witmer coefficient analysis are complimentary for the diagnosis of infectious uveitis. Am J Ophthalmol. 2006;141:313–318.
- Sugita S, Shimizu N, Watanabe K, et al. Use of multiplex PCR and real-time PCR to detect human herpes virus genome in ocular fluids of patients with uveitis. Br J Ophthalmol. 2008;92:928–932.
- Suzuki K, Suzuki Y, Matsumoto M, Expression NM. Profile of intravitreous cytokines, chemokines and growth factors in patients with fuchs heterochromic iridocyclitis. Case Rep Ophthalmol. 2010;1:5–13.
- Kinchington PR, Leger AJ, Guedon JM, Hendricks RL. Herpes simplex virus and varicella zoster virus, the house guests who never leave. Herpesviridae. 2012;3:5.
- Ludlow M, Kortekaas J, Herden C, et al. Neurotropic virus infections as the cause of immediate and delayed neuropathology. Acta Neuropathol. 2016;131:159–184.
- Ouwendijk WJ, Laing KJ, Verjans GM, Koelle DM. T-cell immunity to human alphaherpesviruses. Curr Opin Virol. 2013;3:452–460.
- St Leger AJ, Hendricks RL. CD8+ T cells patrol HSV-1-infected trigeminal ganglia and prevent viral reactivation. J Neurovirol. 2011;17:528–534.
- Verjans GM, Hintzen RQ, Van Dun JM, et al. Selective retention of herpes simplex virus-specific T cells in latently infected human trigeminal ganglia. Proc Natl Acad Sci U S A. 2007;104:3496–3501.
- Remeijer L, Duan R, Van Dun JM, Wefers Bettink MA, Osterhaus AD, Verjans GM. Prevalence and clinical consequences of herpes simplex virus type 1 DNA in human cornea tissues. J Infect Dis. 2009;200:11–19.
- Lee S, Ives AM, Bertke AS. Herpes simplex virus 1 reactivates from autonomic ciliary ganglia independently from sensory trigeminal ganglia to cause recurrent ocular disease. J Virol. 2015;89:8383–8391.
- Richter ER, Dias JK, Gilbert JE 2nd, Atherton SS. Distribution of herpes simplex virus type 1 and varicella zoster virus in ganglia of the human head and neck. J Infect Dis. 2009;200:1901–1906.
- Wang XC, Norose K, Yano A, Ohta K, Segawa K. Two-color flow cytometric analysis of activated T lymphocytes in aqueous humor of patients with endogenous vs. exogenous uveitis. Curr Eye Res. 1995;14:425–433.
- Verjans GM, Feron EJ, Dings ME, et al. T cells specific for the triggering virus infiltrate the eye in patients with herpes simplex virus-mediated acute retinal necrosis. J Infect Dis. 1998;178:27–34.
- Milikan JC, Kinchington PR, Baarsma GS, Kuijpers RW, Osterhaus AD, Verjans GM. Identification of viral antigens recognized by ocular infiltrating T cells from patients with varicella zoster virus-induced uveitis. Invest Ophthalmol Vis Sci. 2007;48:3689–3697.
- Curnow SJ, Falciani F, Durrani OM, et al. Multiplex bead immunoassay analysis of aqueous humor reveals distinct cytokine profiles in uveitis. Invest Ophthalmol Vis Sci. 2005;46:4251–4259.
- Takase H, Futagami Y, Yoshida T, et al. Cytokine profile in aqueous humor and sera of patients with infectious or noninfectious uveitis. Invest Ophthalmol Vis Sci. 2006;47:1557–1561.
- Ouwendijk WJ, Geluk A, Smits SL, Getu S, Osterhaus AD, Verjans GM. Functional characterization of ocular-derived human alphaherpesvirus cross-reactive CD4 T cells. J Immunol. 2014;192:3730–3739.
- Varani S, Landini MP. Cytomegalovirus-induced immunopathology and its clinical consequences. Herpesviridae. 2011;2:6.
- Chan AS, Mehta JS, Al Jajeh I, Iqbal J, Anshu A, Tan DT. Histological features of Cytomegalovirus-related corneal graft infections, its associated features and clinical significance. Br J Ophthalmol. 2016;100:601–606.
- La Rosa C, Diamond DJ. The immune response to human CMV. Future Virol. 2012;7:279–293.
- Rao NA, Zhang J, Ishimoto S. Role of retinal vascular endothelial cells in development of CMV retinitis. Trans Am Ophthalmol Soc. 1998;96:111–123. discussion 124-116
- Read RW, Zhang JA, Ishimoto SI, Rao NA. Evaluation of the role of human retinal vascular endothelial cells in the pathogenesis of CMV retinitis. Ocul Immunol Inflamm. 1999;7:139–146.
- Kobayashi A, Yokogawa H, Higashide T, Nitta K, Sugiyama K. Clinical significance of owl eye morphologic features by in vivo laser confocal microscopy in patients with cytomegalovirus corneal endotheliitis. Am J Ophthalmol. 2012;153:445–453.
- Shiraishi A, Hara Y, Takahashi M, et al. Demonstration of. “owl’s eye” morphology by confocal microscopy in a patient with presumed cytomegalovirus corneal endotheliitis. Am J Ophthalmol. 2007;143:715–717.
- Chee SP, Bacsal K, Jap A, Se-Thoe SY, Cheng CL, Tan BH. Corneal endotheliitis associated with evidence of cytomegalovirus infection. Ophthalmology. 2007;114:798–803.
- Chee SP, Bacsal K, Jap A, Se-Thoe SY, Cheng CL, Tan BH. Clinical features of cytomegalovirus anterior uveitis in immunocompetent patients. Am J Ophthalmol. 2008;145:834–840.
- Chee SP, Jap A. Presumed fuchs heterochromic iridocyclitis and Posner-Schlossman syndrome: comparison of cytomegalovirus-positive and negative eyes. Am J Ophthalmol. 2008;146:883–889.
- Choi JA, Kim J-E, Noh S-J, Kim EK, Park CK, Paik S-Y. Enhanced cytomegalovirus infection in human trabecular meshwork cells and its implication in glaucoma pathogenesis. Sci Rep. 2017;7:43349.
- Park SW, Yu HG. Association of cytomegalovirus with idiopathic chronic anterior uveitis with ocular hypertension in Korean patients. Ocul Immunol Inflamm. 2013;21:192–196.
- Burd EM, Pulido JS, Puro DG, O’Brien WJ. Replication of human cytomegalovirus in human retinal glial cells. Invest Ophthalmol Vis Sci. 1996;37:1957–1966.
- Carmichael A. Cytomegalovirus and the eye. Eye (Lond). 2012;26:237–240.
- Taylor-Wiedeman J, Sissons JG, Borysiewicz LK, Sinclair JH. Monocytes are a major site of persistence of human cytomegalovirus in peripheral blood mononuclear cells. J Gen Virol. 1991;72;(Pt 9):2059–2064.
- Hayashi K, Kurihara I, Uchida Y. Studies of ocular murine cytomegalovirus infection. Invest Ophthalmol Vis Sci. 1985;26:486–493.
- Xagorari A, Chlichlia K. Toll-like receptors and viruses: induction of innate antiviral immune responses. Open Microbiol J. 2008;2:49–59.
- Bale JF Jr., O’Neil ME, Folberg R. Murine cytomegalovirus ocular infection in immunocompetent and cyclophosphamide-treated mice. Potentiation of ocular infection by cyclophosphamide. Invest Ophthalmol Vis Sci. 1991;32:1749–1756.
- Bale JF Jr., O’Neil ME, Lyon B, Perlman S. The pathogenesis of murine cytomegalovirus ocular infection. Anterior chamber inoculation. Invest Ophthalmol Vis Sci. 1990;31:1575–1581.
- Hayashi K, Suwa Y, Shimomura Y, Ohashi Y. Pathogenesis of ocular cytomegalovirus infection in the immunocompromised host. J Med Virol. 1995;47:364–369.
- Nahdi I, Abdelwahed RB, Boukoum H, et al. Herpesvirus detection and cytokine levels (IL-10, IL-6, and IFN-gamma) in ocular fluid from Tunisian immunocompetent patients with uveitis. J Med Virol. 2013;85:2079–2086.
- Li J, Ang M, Cheung CM, et al. Aqueous cytokine changes associated with Posner-Schlossman syndrome with and without human cytomegalovirus. PLoS One. 2012;7:e44453.
- Siak J, Chee SP. Cytomegalovirus Anterior Uveitis Following Topical Cyclosporine A. Ocul Immunol Inflamm. 2017;1–4.
- Sims JL, Chee SP. Cytomegalovirus endotheliitis following fluocinolone acetonide (Retisert) implant. Eye (Lond). 2010;24:197–198.
- Ashwell JD, Lu FW, Vacchio MS. Glucocorticoids in T cell development and function*. Annu Rev Immunol. 2000;18:309–345.
- De Smet MD, Nussenblatt RB. Clinical use of cyclosporine in ocular disease. Int Ophthalmol Clin. 1993;33:31–45.
- Weir E, Sider D. A refresher on rubella. Canadian Med Ass J. 2005;172:1680–1681.
- Quentin CD, Reiber H. Fuchs heterochromic cyclitis: rubella virus antibodies and genome in aqueous humor. Am J Ophthalmol. 2004;138:46–54.
- Mahendradas P, Shetty R, Malathi J, Madhavan HN. Chikungunya virus iridocyclitis in Fuchs’ heterochromic iridocyclitis. Indian J Ophthalmol. 2010;58:545–547.
- Jones NP. Fuchs’ heterochromic uveitis: an update. Surv Ophthalmol. 1993;37:253–272.
- Dithmar S, Tetz MR, Volcker HE. [Fuchs’ heterochromic cyclitis. Clinico-histopathologic findings of nodular iritis]. Klin Monbl Augenheilkd. 1996;209:158–162.
- Melamed S, Lahav M, Sandbank U, Yassur Y, Ben-Sira I. Fuch’s heterochromic iridocyclitis: an electron microscopic study of the iris. Invest Ophthalmol Vis Sci. 1978;17:1193–1199.
- Murray PI, Mooy CM, Visser-De Jong E, et al. Immunohistochemical analysis of iris biopsy specimens from patients with Fuchs’ heterochromic cyclitis. Am J Ophthalmol. 1990;109:394–399.
- Wobmann P. Fuchs’s heterochromic cyclitis. Electron-microscopic study of nine iris biopsies (author’s transl). Albrecht Von Graefes Arch Klin Exp Ophthalmol. 1976;199:167–178.
- La Hey E, Mooy CM, Baarsma GS, De Vries J, De Jong PT, Kijlstra A. Immune deposits in iris biopsy specimens from patients with Fuchs’ heterochromic iridocyclitis. Am J Ophthalmol. 1992;113:75–80.
- Ueber FE. Komplikationen der Heterochromie. Z Augenheilk. 1906;15:191–212.
- Norrsell K, Sjodell L. Fuchs’ heterochromic uveitis: a longitudinal clinical study. Acta Ophthalmol. 2008;86:58–64.
- Cimino L, Aldigeri R, Parmeggiani M, et al. Searching for viral antibodies and genome in intraocular fluids of patients with Fuchs uveitis and non-infectious uveitis. Graefes Arch Clin Exp Ophthalmol. 2013;251:1607–1612.
- Jdf DG-M, De Visser L, Rothova A, Schuller M, Van Loon AM, Weersink AJ. Rubella virus is associated with fuchs heterochromic iridocyclitis. Am J Ophthalmol. 2006;141:212–214.
- Ruokonen PC, Metzner S, Ucer A, Torun N, Hofmann J, Pleyer U. Intraocular antibody synthesis against rubella virus and other microorganisms in Fuchs’ heterochromic cyclitis. Graefes Arch Clin Exp Ophthalmol. 2010;248:565–571.
- Suzuki J, Goto H, Komase K, et al. Rubella virus as a possible etiological agent of Fuchs heterochromic iridocyclitis. Graefes Arch Clin Exp Ophthalmol. 2010;248:1487–1491.
- Labalette P, Caillau D, Grutzmacher C, Dessaint JP, Labalette M. Highly focused clonal composition of CD8(+) CD28(neg) T cells in aqueous humor of fuchs heterochromic cyclitis. Exp Eye Res. 2002;75:317–325.
- Muhaya M, Calder V, Towler HM, Shaer B, McLauchlan M, Lightman S. Characterization of T cells and cytokines in the aqueous humour (AH) in patients with Fuchs’ heterochromic cyclitis (FHC) and idiopathic anterior uveitis (IAU). Clin Exp Immunol. 1998;111:123–128.
- Spriewald BM, Lefter C, Huber I, Lauer B, Wenkel H. A suggestive association of fuchs heterochromic cyclitis with cytotoxic T cell antigen 4 gene polymorphism. Ophthalmic Res. 2007;39:116–120.
- De Visser L, De Boer JH, Rijkers GT, et al. Cytokines and chemokines involved in acute retinal necrosis. Invest Ophthalmol Vis Sci. 2017;58:2139–2151.