
ABSTRACT
Little is known about the immune status of different pig breeds and the role of dietary protein intake. The present study, performed on 96 barrows (48 pure-bred Bama mini-pigs (fatty genotype) and 48 Landrace pigs (lean genotype)) randomly assigned to two dietary treatment groups (low- or adequate protein diet), with twenty-four24 individuals per treatment), evaluated the levels of immunoglobulins and cytokines in serum, and intestinal and splenic innate immunity on the two breeds. Data showed significant interactions between breed and diet for adaptive and innate immunity. Throughout the experiment, Landrace pigs showed significantly higher intestinal immune responses than Bama mini-pigs, and an adequate provision of dietary protein improved the expression of specific antibodies in early growth phases. These finding have important implication for improving the immune status of Landrace pigs, which are a major source of protein for humans.
Abbreviations: BW: Body Weight; NRC diet: National Research Council diet; GB diet: Chinese conventional diet; CSFV: Classical Swine Fever Virus; IgG: Immunoglobulin G; IgM: Immunoglobulin M; SIgA: Secretory Immunoglobulin A.
1. Introduction
Pig is a major protein source for human consumption, and health problems in the swine industry cause significant economical loses worldwide (Tang et al., Citation2014; Tang, Li, et al., Citation2015; Tang, Tan, et al., Citation2015). In addition to vaccines and antibiotics (Gonzalez-Perez et al., Citation2016; Zhang et al., Citation2017), nutritional methods, including dietary supplementation with amino acids (Tan et al., Citation2009), probiotics (Lan & Kim, Citation2017), zinc (Uemura, Katsuge, Sasaki, Goto, & Sueyoshi, Citation2017), or herbs (Bai et al., Citation2016), have been used to improve pig immunity.
More than 30 indigenous pig breeds have been established in China. Bama mini-pig (Liu, Li, He, et al., Citation2015; Tang, Tan, et al., Citation2015), a Chinese breed grown in subtropical southern China, displays tolerance to infection and heat stress, thereby constituting a good model for research on pig immunity status. In contrast, Landrace pig, a typical lean genotype, shows fast-growing ability and has been extensively used to produce a large amount of meat (Liu, Kong, Jiang, et al., Citation2015).
Few reports indicate that dietary protein levels play an important role in regulating immune functions (Opapeju, Rademacher, & Nyachoti, Citation2009; Peng et al., Citation2016; Procajlo, Srzweda, Siwicki, Platt-Samoraj, & Mikulska-Skupien, Citation2010; Yengkokpam et al., Citation2016). The present study aimed to determine the effects of dietary nutrient levels on the specific and innate immune status of Bama mini-pigs and Landrace pigs during three phase of their growth.
2. Materials and methods
2.1. Animals and dietary treatment
Ninety-six barrows (48 Bama mini-pigs, average initial body weight (BW) 3·38 ± 0·96 kg, Chinese local breed; and 48 Landrace pigs, average initial BW 7·68 ± 0·89 kg) were fed different diets from five weeks of age to market weight. Four different treatments were applied in a 2 × 2 factorial arrangement () comprising two breeds (Bama mini-pigs vs. Landrace pigs) and two dietary protein levels [National Research Council diet (NRC diet) vs. Chinese conventional diet (GB diet)]. Barrows from each genotype were randomly assigned to one of the two dietary treatments (24 barrows per treatment). The NRC diet was formulated to meet NRC requirements, whereas the GB diet, with lower protein level than the NRC diet, was formulated to meet the recommendations of the Chinese National Feeding Standard for Swine. The animals were individually housed in 0·6 m × 1·2 m pens with hard plastic slatted flooring, and had free access to drinking water and their respective diets. The dietary phase was based on animals’ physiological stage. The experiment was carried out in accordance with the Chinese Guidelines for Animal Welfare and Experimental Protocol, and approved by the Animal Care and Use Committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences.
Table 1. Animals and treatments.
2.2. Vaccination and sample collection
All Landrace pigs and Bama mini-pigs were confirmed negative for classical swine fever virus (CSFV) by real-time PCR (RT–PCR). Pigs of each group were inoculated with the Classical Swine Fever Vaccine (Weike Biotechnology, China) in the muscle of the neck, behind the ear, according to the manufacturer’s recommendation, at the age of 21 days. Following vaccination, pigs were monitored daily for temperature/clinical signs and bled for serology and cytokine analyses during nursery, growing, and finishing phases (hereafter designated phases 1, 2, and 3, respectively). The BW ranges of Landrace pigs in phases 1, 2, and 3 were 7–20, 20–50, and 50–90 kg, respectively, while BW ranges for Bama mini-pigs were 3–15, 15–35, and 35–55 kg, respectively. At the end of each phase, blood samples from eight pigs under each treatment were collected into 10-ml centrifuge tubes containing sodium heparin (14.3 USP units/ml). Plasma was recovered from each sample after centrifugation at 900 × g for 10 min at 4°C, and stored at −80°C until analysis. Spleens were immediately snap-frozen in liquid nitrogen and stored at −80°C for the extraction of total RNA.
2.3. Analysis of serum swine fever antibodies, and immunoglobulins G and M (IgG and IgM, respectively) and cytokines
Specific immune responses were determined by the concentration of swine fever antibodies (Cusabio Biotech Company Limited). Two aliquots (100 μl) of each sample or standard were added to the wells of a microtitre plate previously pre-coated with an appropriate antigen. The negative control serum was used as the negative control. The plate was incubated for 1 h at 37°C, and, after removing the liquid within each well, horseradish peroxidase (HRP)-conjugate antibody (100 µl) was added and the plate incubated for another hour at 37°C. After this period, wells were washed five times with 200 µl of washing buffer, and Trimethylborane (TMB) was added. After incubation for 15 min at 37°C, the reaction was stopped with 50 μl of the terminating solution, and the absorbance of the resulting solution was measured at 450 nm.
Serum concentrations of IgG and IgM were measured using immunoassay kits purchased from Triple J Farms (Bellingham, WA, USA), according to the manufacturer’s instructions. Serum concentrations of interleukin IL-2, IL-10 and tumour necrosis factor (TNF)-a were determined by radio-immunoassays using kits provided by Beijing Chemclin Biotech (Beijing, China).
2.4. Measurement of intestinal innate immunity and cytokine gene expression levels
The abundance of secretory immunoglobulin A (SIgA) was determined using an enzyme-linked immunosorbent assay (ELISA) kit (Abcam. USA.), following the manufacturer’s instructions. The jejunum was collected for RNA extraction and immediately frozen at −80°C until additional processing. Total RNA was extracted using the RNA simple kit (Sangon Inc. Ltd., Shanghai, China), according to the instructions of the manufacturer. Purity and quantity of RNA were measured on a NanoDrop spectrophotometer (Thermo Fisher Scientific, DE, USA), and RT–PCR was performed as described in the following section.
2.5. Analysis of cytokine gene expression in the spleen
Tissue is gently dissociated and whole spleens are prepared to single-cell suspensions of splenocytes. Total RNA was extracted from the splenocytes using the RNA simple kit (Sangon Inc. Ltd) and treated with DNase I (Takara, Dalian, China) at 37°C for 30 min. Purity and quantity of RNA were measured on a NanoDrop spectrophotometer (Thermo Fisher Scientific). Only intact RNA samples with 260:280 nm ratio between 2.0 and 2.2 were used for cDNA synthesis. First-strand cDNA was synthesized from total RNA using the M-MLV First-Strand cDNA synthesis kit (Promega, USA). Quantitative RT–PCR was conducted to detect relative mRNA expression levels using β-actin as the internal control gene and the SYBR® Green Real-time PCR Master Mix kit (ToYoBo Co. Ltd, Japan), according to the manufacturer’s instructions. Reaction mixtures included 7.5 μL PCR master mix, 0.4 μL 10 μM primers (), and 1 μL cDNA, and PCR profile was performed at 95°C for 30 s, followed by 40 cycles at 95°C for 10 s, 55°C for 10 s, and 72°C for 15 s, in the iQ5TM real-time multicolour PCR detector (Bio-Rad, USA). The relative quantification of gene amplification by RT–PCR was performed using the cycle threshold (Ct) method in which expression levels of cytokine genes were determined in relation to those of β-actin.
2.6. Statistical analysis
Data were analyzed using a mixed-effects model in SAS version 8.2 (SAS Institute Inc., Cary, NC, USA). Diet, breed, and the interactions between these factors were included in the statistical model. Effects were considered statistically significant at P < 0.05. Probability values between 0.05 and 0.10 were considered trends.
3. Results
3.1. Immunoglobulins and cytokines in serum
In phases 1 and 2, Landrace pigs fed the NRC diet showed higher level of specific antibodies for swine fever than Landrace pigs fed the GB diet, while there was no different in phase 3 Landrace pigs. In addition, interaction between Bama mini-pigs and Landrace pigs was not significant, although mean values were greater in Bama mini-pigs ((a)). The concentration of IgG showed significant breed interaction (P < 0.05) in phase 2 but not in phases 1 and 3 ((b)). Similar levels of IgM were found among the three different phases (P > 0.05) ((c)).
Figure 1. Effects of dietary protein intake and breed on the levels of specific classical swine fever virus (CSFV) antibodies and immunoglobulins G and M in serum, in growth phases 1, 2, and 3. (a) concentration of CSFV antibodies; (b) concentration of IgG; (c) concentrations of IgM. 
 Landrace pig/National Research Council (NRC) diet group;
Landrace pig/National Research Council (NRC) diet group;  Bama mini-pig/GB diet group;
Bama mini-pig/GB diet group;  Bama mini-pig/NRC diet group. Values are means (n = 8), and standard errors are represented by vertical bars. Effects were considered statistically significant if P < 0.05. S × D, strain × diet interaction.
Bama mini-pig/NRC diet group. Values are means (n = 8), and standard errors are represented by vertical bars. Effects were considered statistically significant if P < 0.05. S × D, strain × diet interaction.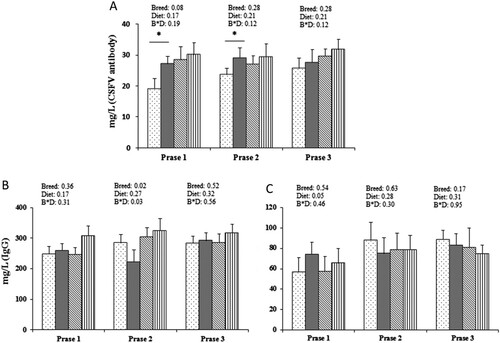
Dietary protein levels showed no significant breed × diet effect (P > 0.05) on serum concentrations of IL-2 and TNF-α in the three phases, except for the slight fluctuations of IL-2 in phase 3 and TNF-α in phase 1 (). The level of IL-10 was significantly affected by breed and diet, and Landrace pigs fed the NRC diet generally produced higher IL-10 than those fed the GB diet. On the contrary, Bama mini-pigs fed the GB diet activated more IL-10 secretion in serum than Bama mini-pigs fed the NRC diet.
Table 2. Effects of dietary protein level and breed on serum concentrations of cytokines in the serum and relative mRNA level of intestinal and spleen.
3.2. Intestinal innate immunity and cytokines
As shown in , jejunum SIgA was higher in Landrace pigs than in Bama-mini-pigs, showing breed interaction (P < 0.05) but not diet interaction (P > 0.05). Breed differences in SIgA were consistent with cytokines expression levels. In phase 1, there was no significant breed or diet interaction (P > 0.05) for IL-2, IL-4, IL-10, IL-1β, TNF-α, and TGF-β1, although Landrace pigs had significantly lower IL-6 and significantly higher IFN-γ than Bama mini-pigs. In phase 2, breed × diet interaction was significant (P > 0.05) for IFN-γ and TGF-β1, and breed significantly affected (P < 0.05) the gene expression of IL-4 and IL-10 (). In phase 3, there were significant breed × diet interactions for IFN-γ and TNF-α, and Landrace pigs had significantly higher IL-6, IL-10, IL-1β, and TGF-β1 expression levels than Bama mini-pigs. Diet, on the other hand, did not affect gene expression (P > 0.05) ().
Figure 2. Effects of dietary protein intake and breed on the intestinal secretion of secretory immunoglobulin A (SIgA) in growth phases 1, 2, and 3. (a) concentration of SIgA in jejunum; (b) concentration of SIgA in the lumen of jejunum.
Landrace pig/ National Research Council (NRC) diet group; Bama mini-pig/GB diet group; Bama mini-pig/NRC diet group; Values are means (n = 8), and standard errors represented by vertical bars. Effects were considered statistically significant if P < 0.05. S × D, strain × diet interaction.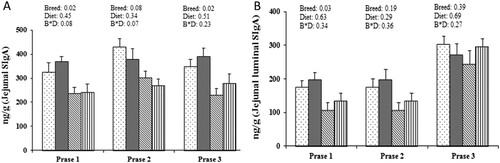
3.3. Cytokines in spleen
As show in , there was no significant breed or diet interaction for IL-4, IL-1β, IFN-γ, and TNF-α in the three different phases. In phase 1, breed interaction was significant (P < 0.05) for the splenic expression of IL-2, IL-6, IL-10, and TGF-β1 as Landrace pigs fed GB or NRC diets had higher expression level than Bama mini-pigs. However, no significant diet interactions (P > 0.05) were observed for the expression of these cytokines. In phases 2 and 3, only IL-6 and IL-10 showed significant breed interactions (P < 0.05). Significant breed × diet interactions were only shown for IL-10 expression during phase 2 (P < 0.05).
4. Discussion
All pigs showed healthy growth throughout the experimental period, and their growth performance, meat quality, plasma metabolites, and lipid metabolism have been described previously (Liu, Kong, Jiang, et al., Citation2015; Liu, Li, He, et al., Citation2015; Liu, Li, Kong, et al., Citation2015). Because the immune status of pigs depends on the interactive effects of genotype, vaccination, antibiotics’ supply, and nutritional condition, the present study focused on evaluating the effects of breed and dietary protein level on adaptive and innate immunity. Diet significantly affected the specific immunity of Landrace pigs, with increasing levels of dietary protein increasing the titer of CFSV antibodies, although both IgG and IgM levels did not significantly differed between diets. Cytokines (Tan et al., Citation2009; Thorum et al., Citation2013), such as IL-2, IL-10, and TNF-α, are crucial protein mediators in humoral immunity, and the mRNA levels of cytokines in serum were affected by both diet and breed, particularly for IL-10, which enhances B cell survival and antibody production. These results indicated that cytokines in serum were significantly affected by breed or diet, and high levels of dietary protein increased the level of specific antibodies on early growth stages.
The intestinal immune system is an important line of defense and can protect the animal from pathogenic microorganisms and toxins (Tang, Li, et al., Citation2015). Levels of SIgA (Min, Yang, Xu, & Gao, Citation2016; Tian et al., Citation2014), as a key mark for intestinal immunity, showed significantly breed interaction with Landrace pigs showing higher SIgA levels than Bama mini-pigs, and the high-protein diet increased SIgA levels in Landrace pigs in phase 1 but not in phases 2 and 3. Consistent with SIgA levels, the expression of partial cytokines in Bama mini-pigs were upregulated by approximately 200%–300%, in relation to Landrace pigs. Thus, Landrace pigs seem to have stronger intestinal immune responses than Bama mini-pigs, and high protein intake seems to improve gene regulation in the intestine of Landrace pigs during phase 1.
The spleen plays important roles in the immune system and in antibodies’ production, which might indicate the immune status of an animal. The present results indicated significant effects of breed, but not of diet, on cytokines’ expression in the spleen.
5. Conclusion
In summary, the results of the present study suggest an interaction effect between breed and diet. Landrace pigs show more diet benefits on intestinal immune responses, and increasing dietary protein intake in animals belonging to this breed might improve their immune status to some extent, while the Bama mini-pigs are suitable to lower protein level for their immune status.
Acknowledgements
This research was supported by a grant from State Key Laboratory foundation of Animal Nutrition (2004DA125184F1701), National Natural Science Foundation of China (NSFC, 31402091) and Natural Science Foundation of Hunan Province (2016JJ4087).
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Bai, X., Yan, X., Xie, L., Hu, X., Lin, X., Wu, C., … See, M. T. (2016). Effects of pre-slaughter stressor and feeding preventative Chinese medicinal herbs on glycolysis and oxidative stability in pigs. Animal Science Journal = Nihon Chikusan Gakkaiho, 87, 1028–1033.
- Gonzalez-Perez, G., Hicks, A. L., Tekieli, T. M., Radens, C. M., Williams, B. L., & Lamouse-Smith, E. S. (2016). Maternal antibiotic treatment impacts development of the neonatal intestinal microbiome and antiviral immunity. The Journal of Immunology, 196, 3768–3779. doi: https://doi.org/10.4049/jimmunol.1502322
- Lan, R. X., & Kim, I. H. (2017). Effects of dietary supplementation with a probiotic (Enterococcus faecium DSM 7134) on growth performance, nutrient digestibility, and gut health status in weaning pigs. Journal of the Science of Food and Agriculture, 98(2). doi: https://doi.org/10.1002/jsfa.8374.10.1002/jsfa.8374
- Liu, Y., Kong, X., Jiang, G., Tan, B., Deng, J., Yang, X., … Yin, Y. (2015). Effects of dietary protein/energy ratio on growth performance, carcass trait, meat quality, and plasma metabolites in pigs of different genotypes. Journal of Animal Science and Biotechnology, 6, 8. doi: https://doi.org/10.1186/s40104-015-0001-8
- Liu, Y., Li, F., He, L., Tan, B., Deng, J., Kong, X., … Wu, G. (2015). Dietary protein intake affects expression of genes for lipid metabolism in porcine skeletal muscle in a genotype-dependent manner. British Journal of Nutrition, 113, 1069–1077. doi: https://doi.org/10.1017/S0007114514004310
- Liu, Y., Li, F., Kong, X., Tan, B., Li, Y., Duan, Y., … Yin, Y. (2015). Signaling pathways related to protein synthesis and amino acid concentration in pig skeletal muscles depend on the dietary protein level, genotype and developmental stages. PloS One, 10, e0138277. doi: https://doi.org/10.1371/journal.pone.0138277
- Min, Y. N., Yang, H. L., Xu, Y. X., & Gao, Y. P. (2016). Effects of dietary supplementation of synbiotics on growth performance, intestinal morphology, sIgA content and antioxidant capacities of broilers. Journal of Animal Physiology and Animal Nutrition, 100, 1073–1080. doi: https://doi.org/10.1111/jpn.12479
- Opapeju, F. O., Rademacher, M., & Nyachoti, C. M. (2009). Effect of dietary crude protein level on jejunal brush border enzyme activities in weaned pigs. Archives of Animal Nutrition, 63, 455–466. doi: https://doi.org/10.1080/17450390903299133
- Peng, X., Hu, L., Liu, Y., Yan, C., Fang, Z. F., Lin, Y., … Che, L. Q. (2016). Effects of low-protein diets supplemented with indispensable amino acids on growth performance, intestinal morphology and immunological parameters in 13 to 35 kg pigs. Animal: An International Journal of Animal Bioscience, 10, 1812–1820. doi: https://doi.org/10.1017/S1751731116000999
- Procajlo, Z., Srzweda, W., Siwicki, A. K., Platt-Samoraj, A., & Mikulska-Skupien, E. (2010). Indices of non-specific and specific humoral immunity in pigs immunomodulated with the Bioimmuno preparation and/or immunised with the respisure one vaccine against mycoplasmal pneumonia of swine. Polish Journal of Veterinary Sciences, 13, 447–455.
- Tan, B., Li, X. G., Kong, X., Huang, R., Ruan, Z., Yao, K., … Wu, G. (2009). Dietary L-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids, 37, 323–331. doi: https://doi.org/10.1007/s00726-008-0155-1
- Tang, Y., Li, J., Li, F., Hu, C. A., Liao, P., Tan, K., … Yin, Y. (2015). Autophagy protects intestinal epithelial cells against deoxynivalenol toxicity by alleviating oxidative stress via IKK signaling pathway. Free Radical Biology and Medicine, 89, 944–951. doi: https://doi.org/10.1016/j.freeradbiomed.2015.09.012
- Tang, Y., Li, F., Tan, B., Liu, G., Kong, X., Hardwidge, P. R., & Yin, Y. (2014). Enterotoxigenic Escherichia coli infection induces intestinal epithelial cell autophagy. Veterinary Microbiology, 171, 160–164. doi: https://doi.org/10.1016/j.vetmic.2014.03.025
- Tang, Y., Tan, B., Xiong, X., Li, F., Ren, W., Kong, X., … Yin, Y. (2015). Methionine deficiency reduces autophagy and accelerates death in intestinal epithelial cells infected with enterotoxigenic Escherichia coli. Amino Acids, 47, 2199–2204. doi: https://doi.org/10.1007/s00726-014-1781-4
- Thorum, S. C., Comstock, S. S., Hester, S. N., Shunk, J. M., Monaco, M. H., & Donovan, S. M. (2013). Impact of diet on development of bronchial-associated immunity in the neonatal piglet. Veterinary Immunology and Immunopathology, 151, 63–72. doi: https://doi.org/10.1016/j.vetimm.2012.10.006
- Tian, E. J., Zhou, B. H., Wang, X. Y., Zhao, J., Deng, W., & Wang, H. W. (2014). Effect of diclazuril on intestinal morphology and SIgA expression in chicken infected with Eimeria tenella. Parasitology Research, 113, 4057–4064. doi: https://doi.org/10.1007/s00436-014-4074-7
- Uemura, R., Katsuge, T., Sasaki, Y., Goto, S., & Sueyoshi, M. (2017). Effects of zinc supplementation on Shiga toxin 2e-producing Escherichia coli in vitro. Journal of Veterinary Medical Science, 79, 1637–1643. doi: https://doi.org/10.1292/jvms.16-0471
- Yengkokpam, S., Debnath, D., Sahu, N. P., Pal, A. K., Jain, K. K., & Baruah, K. (2016). Dietary protein enhances non-specific immunity, anti-oxidative capability and resistance to Aeromonas hydrophila in Labeo rohita fingerlings pre-exposed to short feed deprivation stress. Fish & Shellfish Immunology, 59, 439–446. doi: https://doi.org/10.1016/j.fsi.2016.10.052
- Zhang, C., Yu, M., Yang, Y., Mu, C., Su, Y., & Zhu, W. (2017). Differential effect of early antibiotic intervention on bacterial fermentation patterns and mucosal gene expression in the colon of pigs under diets with different protein levels. Applied Microbiology and Biotechnology, 101, 2493–2505. doi: https://doi.org/10.1007/s00253-016-7985-7