
Abstract
Purpose: Cancer initialization can be explained as a result of parasitic virus energy consumption leading to randomized genome chemical bonding.
Materials and methods: Analysis of experimental data on cell-mediated immunity (CMI) containing about 12,000 cases of healthy humans, cancer patients and patients with precancerous cervical lesions disclosed that the specific cancer and the non-specific lactate dehydrogenase-elevating (LDH) virus antigen elicit similar responses. The specific antigen is effective only in cancer type of its origin but the non-specific antigen in all examined cancers. CMI results of CIN patients display both healthy and cancer state. The ribonucleic acid (RNA) of the LDH virus parasitizing on energy reduces the ratio of coherent/random oscillations. Decreased effect of coherent cellular electromagnetic field on bonding electrons in biological macromolecules leads to elevating probability of random genome reactions.
Results: Overlapping of wave functions in biological macromolecules depends on energy of the cellular electromagnetic field which supplies energy to bonding electrons for selective chemical bonds. CMI responses of cancer and LDH virus antigens in all examined healthy, precancerous and cancer cases point to energy mechanism in cancer initiation.
Conclusions: Dependence of the rate of biochemical reactions on biological electromagnetic field explains yet unknown mechanism of genome mutation.
Introduction
Oncogene mutation is a process of cancer initiation. Different somatic mutations are analyzed in a number of papers, for instance in Alexandrov et al. (Citation2013), Davies et al. (Citation2002), Dhomen et al. (Citation2009), Dankort et al. (Citation2009), Tsai et al. (Citation2008) and Vredeveld et al. (Citation2012) but understanding the biological processes underlying genome somatic mutations is limited. Thousands of somatic mutations are identified in a single cancer sample but the majority of them are only accompanying events of oncogene mutation (Alexandrov et al. Citation2013) which indicates that genome resistance to mutations is low.
Tomasetti and Vogelstein (Citation2015) proved that the lifetime risk of cancers of many different types is strongly correlated with the total number of divisions of the normal self-renewing cells maintaining homeostasis of the tissue. These results suggest that the total probability of cancer initiation depends on the sum of individual probabilities in particular stem cell divisions. During cancer initiation, the genome memory seems to be open for random access. The mechanism of disturbance of the protection against storing arbitrary information has not been satisfactorily explained yet.
Knox and Funk (Citation2014) claim that biophysical mechanisms in microenvironmental signaling play an important role in gene regulation. It is known that chemical reactions depend on strong enough electric field which can power catalysis and affect their rate (Fried et al. Citation2014, Hildebrandt Citation2014). Therefore, biochemical activity may be affected by near electromagnetic field (EMG) of coherent electric polar vibrations which are present in the cell (Pokorný et al. Citation2013a, Citation2015). If the power of electric oscillations considerably decreases, the EMG field is lowered which may result in deceleration of reactions. Such a situation appears in a cancer cell. Cancer is a pathological state of disturbed oxidative metabolism – mitochondrial dysfunction (Warburg et al. Citation1924, Warburg Citation1956) and inhibited apoptosis. The effects resulting from mitochondrial dysfunction can be explained by Fröhlich’s (Citation1969, Citation1978, Citation1980) hypothesis of coherent electric polar vibrations and generation of EMG field in biological systems and their disturbance in cancer cells. The disturbances of EMG field may also explain the mutational process. Seyfried and Shelton (Citation2010) claimed that nuclear genome integrity is largely dependent on mitochondrial energy homeostasis which may be disturbed by mitochondrial dysfunction. However, the energy level of oscillations in the cell might be lowered by parasitic energy consumption even before development of the mitochondrial dysfunction. In both cases the genome mutation and cancer initiation may result from decrease of EMG generated in the cell.
There exists extensive experimental support. Coherent mechanical vibrations of the membrane of yeast cells dependent on oxidative energy supply were measured by Pelling et al. (Citation2004, Citation2005). These nanoscale vibrations are considered to be a signature of life (Kasas et al. Citation2015). The vibrations can generate EMG field as the majority of biological macromolecules and structures are electrically polar, in particular microtubules (Pokorný et al. Citation1997, Citation2001, Citation2008, Pokorný and Wu Citation1998, Jelínek et al. Citation2009). Mechanisms related to excitation of coherent EMG field in living system was described by Pokorný et al. (Citation2011, Citation2013a, Citation2013b, Citation2014, Citation2015). Further experimental support was provided by Sahu et al. (Citation2013, Citation2014) by measuring microtubule oscillations and absorption in a broad frequency range (0.1–0.3 MHz, 10–30 MHz, 100–200 MHz, 1–20 GHz, infrared band at about 20 THz, and UV band at about 276–334 nm).
Energy of cellular EMG field can be transferred to bonding electrons which exhibit a special behavior in macromolecules. Ventegodt et al. (Citation2006) supposed that the electrons on the orbitals of large biological molecules make one huge ‘cell-orbital’ (‘super-orbital’), which is structured according to the nature of quantum fields. Non-localized electrons on the ‘cell-orbital’ (or macromolecular orbitals) may oscillate in phase with electromagnetic oscillations in the cell. Decrease of energy of the cellular EMG field means decrease of the coherence/randomness ratio in the tissue. If the decrease is sufficiently large, it may result in randomization of chemical reaction and increase of the probability of random effects such as pathological mutations in any cell in the inflicted region. Increase of randomness connected with parasitic energy consumption is assumed to be disclosed by the cell-mediated immunity (CMI) measurement based on cancer and the lactate dehydrogenase-elevating (LDH) virus antigens. Parasitic energy consumption is analysed on the basis of CMI experimental data published by Jandová et al. (Citation1979, Citation2001, Citation2004, Citation2005, Citation2009a, Citation2009b, Citation2015). Elicitation of the immune response depends on the pathological deviations from the normal state. The immune system provides supervision of essential biological functions of living cells; energy and coherence level of electric polar vibrations in cells should be also supervised. The immune response should also include assessment of energy transfer from coherent electric polar vibrations to bonding electrons to exclude random chemical reactions. Analysis of the cellular processes of physical nature involved in randomization and mutability has not been done yet.
Materials and methods
(a) Cell-mediated immunity
CMI is considered to correlate with the adherence of the T lymphocytes from examined patients to solid state surfaces. The leukocyte adherence inhibition (LAI) assay is based on observation of in vitro adherence of T lymphocytes to surfaces of glass or some plastic materials in the presence and in the absence of antigens. CMI response to antigens is related to specific processes in the cells which are under immune surveillance. Not only chemical but also physical agents can affect CMI and change adherence of T lymphocytes. Two types of antigens were used. The specific antigen (AC) is an immunological functional fraction prepared from a malignant tumor of the same type as the malignancy of the patient the blood was taken from. Organs with cancer were taken during surgery. The non-specific antigen (AV) is an immunological functional fraction prepared from the serum of inbred laboratory mouse C3H H2k strain infected with the LDH virus. Both antigens prepared by high-pressure gel chromatography at 340 nm are ribonucleic acids (RNA). A comprehensive description of the preparation method of T lymphocytes, the antigens, and the measurement method is available in the Supplementary data, available online only (Jandová et al. Citation2001, Citation2009a).
LDH virus is a silent virus which does not seemingly harm the host. Its single-stranded RNA does not cause any observable morphological changes but provides a chronic parasitic consumption of energy in the cell. Essential information on LDH virus is based on experimental studies on mice (Brinton Citation1994, Plagemann Citation1996). Infection with LDH virus increases the level of the enzyme lactate dehydrogenase (now classified as NAD 1.1.1.27 Oxidoreductase) in plasma and production of lactate from pyruvate.
The CMI response measured by the described method on patients with cancer of the cervix, endometrium, ovary, breast, lung, melanoblastoma, and otorhinolaryngological cancers was investigated. Histopathological verification of the cancer diagnosis was provided. Conditions for malignant transition in cancer development were studied on precancerous cervical lesions. These lesions were diagnosed by cytological (PAP) smears, from examination by colposcopy, and from colposcopically directed ‘punch’ biopsy material by histology. The grades of development of precancerous lesions LSIL and HSIL (low- and high-grade squamous intraepithelial lesions, respectively) were determined by cytology, and CIN 1, 2, and 3 (cervical intraepithelial neoplasia) grades by histology (Jandová et al. Citation2009a).
(b) Energy of electrons
DNA is a memory maintaining information for the cell activity. Data storage should be feasible to preserve information gained during evolution. It is reasonable to assume DNA as a memory system capable of permanent localized storage with a limited access controlled by a special cellular mechanism to prevent random events. Mass non-localized genome mutation signifies disturbance of the storage controlling mechanism. A role of electric and electromagnetic fields in biochemical reactions has not been comprehensively understood. Analysis of the effect of the cellular electromagnetic field on biochemical bonds is based on the reverse vibrational Stark effect (see also the Supplementary data).
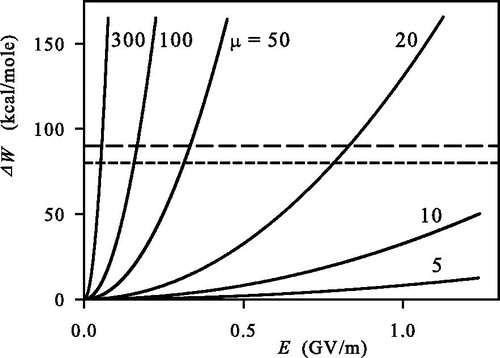
Polar vibrations in biological systems generate cellular electromagnetic field. Coherent polar vibrations excited in cells are an essential feature of biological systems (Pokorný et al. Citation2013a). Electrons on delocalized molecular orbitals in living cells may oscillate in the coherent electric field and can display strong interaction and correlation. Motion of electrons in biological structures and dielectric materials and semiconductors may display similar features. A solid state approximation may be used for assessment of the effect of the electric field on electrons in biological molecules. However, biological structures do not exhibit translational symmetry and specific properties are valid only within a limited region. In such a region of a biological structure, delocalized electrons may resemble electrons in a conduction band of the solid state materials. Effective mass and mobility of electrons depend on scattering and interaction with surrounding atoms and their random vibrations, i.e. on processes connected with energy changes. Intensity of the near electric field at a distance up to several nanometers from the oscillating dipoles may be extremely high, typically in the range around 1 GV/m as illustrated in Figure 2 in the Supplementary data. Increase of the electron energy by the electric field may be written in the form ΔW = p2/(4m*), where m* is the effective mass and the momentum p = m*Eμ, where E is the amplitude of the intensity of the electric field and μ is the mobility of electrons. We assessed the mobility of electrons in the range 5–300 cm2/(Vs); the effective mass is approximately equal to the mass of a free electron. These values were used to calculate the curves in . For comparison, the mobility of electrons in polycrystalline silicon is about 300 cm2/(Vs).
Results
(a) Parasitic energy consumption
An overview of the CMI results is in which displays the effect of AC and AV antigens on T lymphocytes of healthy humans and cancer patients. CMI response to antigens is evaluated from the changes of adherence of T lymphocytes to solid state surfaces (M is a number of non-adherent T lymphocytes in %). The number of investigated cases is given in . T lymphocytes from healthy humans do not produce significant CMI response to antigens (negative results) and only 0.42% (0.67%) of cases for AC (AV) antigens are in the cancer region above the MC boundary (). Both AC and AV antigens elicit a strong CMI response of the majority of T lymphocytes taken from cancer patients (positive results). Only a small amount of cases (0.56% and 0.6% for AC and AV antigen, respectively) remained in the healthy region below MH boundary (). It should be emphasized that the specific antigen AC elicited the CMI response only of T lymphocytes taken from patients with the corresponding tumors, and that the non-specific AV antigen produced the CMI response of T lymphocytes taken from all cancer patients under investigation. More detailed analysis and assessment of statistical significance are available in the Supplementary data (Jandová et al. Citation2001, Citation2009b).
Figure 1. Mean values and standard deviations of the relative number (M) of non-adherent T lymphocytes prepared from blood of (a) healthy humans (H) and cancer patients with different cancers (C) without and with specific (AC) and non-specific (AV) antigens (Jandová et al. Citation2001, Citation2009b), (b) healthy women (H) and patients with precancerous cervical lesions (P) in M < MH, and M > MC region, and cervical cancer (C), with specific (AC) and non-specific (AV) antigens (Jandová et al. Citation2009a). The number of cases is given in . MH and MC determine the boundaries of a noise region between adherence of T lymphocytes from healthy humans and cancer patients.
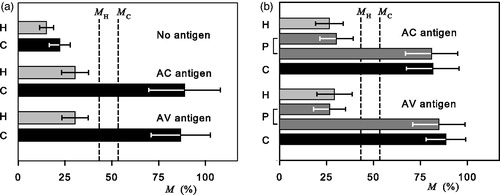
Table 1. Number of cases of non-adherent T lymphocytes (M) prepared from blood of (a) healthy humans (H) and cancer patients (C) with different cancers without antigen and with specific (AC) and non-specific (AV) antigens (Jandová et al. Citation2001; Citation2009b), (b) healthy women and patients with precancerous lesions and cancer of the cervix with specific (AC, in this case the cervical cancer antigen) and non-specific (AV) antigen (Jandová et al. Citation2009a).
and show the cellular immune response of healthy women, patients with precancerous lesions and carcinoma of the cervix to AC and AV antigens (Jandová et al. Citation2009a). The average immune response of T lymphocyte of all precancerous patients is lower than that of the cancer patients. AC and AV antigens, respectively, disclosed that 27.7% and 34.8% of precancerous cases display a healthy state and 67.0% and 59.8% performed transition to cancer. Parts of the CIN subgroups, whose CMI results correspond to healthy humans and cancer patients, were analyzed. Except for one subgroup with the statistical significance 0.05, there are no statistically significant differences between the M values of parts of CIN 1, 2, and 3 subgroups for M < MH or M > MC and healthy women or cervical cancer patients, respectively, for both antigens (Jandová et al. Citation2009a). The results indicate that the transition to cancer malignant properties is connected with the LDH virus infection or a pathological process with a similar CMI response.
The experimental data suggest that information revealed by CMI response to antigens is stored in the memory of T lymphocytes as a marker connected with a factor which is under supervision by the immune system. RNA of the LDH virus is not harmless by itself but parasites on the oxidative metabolism of the cell. Due to interconnection of physical and biochemical processes, disturbances of energy supply and processing must have far-reaching consequences.
About one third of patients with cervical precancerous lesions correspond to healthy women and two thirds to cancer patients. The observed morphological disturbances of precancerous lesions are processes correlating with development of precancerous state to cancer. CMI responses of T lymphocytes taken from cancer patient to AC and AV antigens are similar: the majority of T lymphocytes do not adhere.
(b) Excitation of electrons by cellular electromagnetic field
Energy ΔW of an electron in the electric field is given in . The long-dashed line and the short-dashed line denote the binding energy of the covalent (90 kcal/mole) and the ionic (80 kcal/mole) bonds, respectively. Energy of the ionic bonds in water decreases to 3 kcal/mole (Alberts et al. Citation1994). Even for the intensity of the electric field smaller than 1 GV/m the energy of electron in the electric field generated in a living cell may be much higher than the binding energy. Velocity saturation caused by scattering on oscillating systems (similar to optical phonon scattering) was not considered as the vibrations are coherent and random parts are very weak. In biological systems all charged parts oscillate in phase with coherent polar vibrations. Due to correlation between oscillation parts probability of their bonding might be high.
Figure 2. Increase of electron energy ΔW as a function of the intensity of the electric field E. Mobility of the electrons is equal to or smaller than 300 cm2/(Vs) and effective mass equal to the mass of a free electron. The ΔW energy may be much higher than the energy of the covalent and ionic bonds (the lines with long and short dashes, respectively). Electric field and coherent electromagnetic field can increase probability of bonding and accelerate chemical reactions.
Theoretical analysis of the effect of coherent vibrations on chemical bonds may be performed using relations of the binding energy dependent on the overlap integral S, the Coulomb integral Q, and the exchange integral J (Streitwieser Citation1962). Correlation of the electrons caused by generated electric field is represented by an operator of correlation denoted by R.
The integrals S, Q, and J can be written in the form
where QH and JH are the classical Hamiltonian components, and JR, QR the correlation components dependent on excited coherent oscillating electric field, Ψa and Ψb are wave functions for molecular orbitals a and b, and (1) and (2) refer to two electrons with opposite spins on orbitals of different molecules. Energy of the bond is given by the relation
where ER is the correlation term corresponding to coherent interaction between electrons on orbital a and b. On condition that ER ≫ EH coherent bonding is preferred; if ER ≈ EH probability of a random chemical bond increases, and for ER ≪ EH the randomness is prevailing. The main effect of the near electromagnetic field is very likely based on correlation of both interacting electrons.
Discussion
The probability of cancer initiation seems to depend on natural aggressiveness of carcinogens and liability of genomic system to undergo the oncogene mutations. Viral infections are of special significance. The effects of the high and the low risk Human Papilloma Viruses (HPV) are well-known. The involvement of LDH virus activity in cancer origin has been confirmed by measurements of CMI response of T lymphocytes from cancer patients to the AV antigen. Similar CMI responses have been found in immunological examination in women who had suffered from recurrent spontaneous abortions in early pregnancy without having apparent gynaecological reasons, and also women who had given birth to stillborn babies, having severe organ malformations revealed on dissection (Mejsnarová et al. Citation1981, Jandová et al. Citation2015). Teratogenicity of the Rubella virus may be caused by an analogous mechanism. The experimental results suggest that cancer initiation follows LDH virus infection or an infection agent eliciting similar CMI response. As LDH virus is a parasite on the energy system of infected cells, the process monitored by the non-specific antigen might detect oxidative energy production, i.e. mitochondrial activity. Decreased mitochondrial activity results in decreased level of the state far from the thermodynamic equilibrium and coherence, the essential conditions for life (Pokorný et al. Citation2013a).
The CMI response of T lymphocytes from cancer patients to the non-specific LDH virus and specific cancer antigens very likely detects a decrease of coherence, but not the causes of its decrease. Both types of antigens used are similar immunological functional fractions containing RNA prepared at 340 nm. Specificity of cancer antigen may depend on addition of a particular cancer type part to the identical RNA component. CMI of T lymphocytes of patients with cervical precancerous lesions disclosed an essential feature of the cancer transformation process. CIN precancerous lesions are classified on the basis of morphological disturbances such as macronucleosis, anisonucleosis, coarse chromatin, wrinklings of the nucleus membrane, nucleolus changes (anisokaryosis) etc. Morphological disturbances in the precancerous lesions manifest internal disorganization and very likely represent a framework of processes, which also depend on the level of coherence. CMI response may be elicited when the level of coherence is below a limit value. Some mutations of the genome can be performed at the coherence level above the limit and CMI response formed later after further lowering the coherence (after development of mitochondrial dysfunction).
Near coherent electromagnetic field generated in living cells increases energy of electrons on molecular orbitals. In the coherent electromagnetic field, motion of electrons is synchronized and interactions are correlated. If the additional energy is much higher than the energy of the bonds then the probability of bonding is high. Increase of randomness, decrease of power of the cellular electromagnetic field, and consequent disturbances of water ordering result in decrease of the probability of bonding which may, in turn, increase the tendency to genome mutation. Therefore, it seems reasonable to assume that genome somatic mutation is connected with disturbances of the oxidative metabolism.
Analysis of the effect of LDH virus infection on genome mutation is based on experimental data of CMI response in healthy humans, precancerous, and cancer patients. Infection with LDH virus causes disturbances of physical metabolic processes by parasitic energy consumption, which creates defects of chemical bonds resulting in genome mutation and initiation of pathological states. Therefore, the process of genome mutation includes strongly interdisciplinary links: infection of a biological system, biophysical mechanism decreasing energy production, physical mechanism of decreased coherence of EMG field, defects of chemical bonds, and final biological link of various diseases. Chemical bonds in biological system are considered to depend on coherent EMG fields whose disturbances result in defective bonding and reactions. Biochemical and physical links are thus strongly coupled and cannot be separated.
Conclusion
A novel hypothetical insight into initiation of mass somatic mutations with an essential biophysical mechanism is presented. Mutations of oncogenes seem to be a random event which enables storage of arbitrary information during genome manipulation. The storage is performed by chemical reactions very likely not controlled by the cell. An extensive ensemble of experimental data on the cell-mediated immunity of healthy humans, precancerous, and cancer patients was analyzed to elucidate the process of increased genome mutability in cancer cells and the role of the LDH virus. Cell-mediated immunity responses of T lymphocytes taken from healthy humans and cancer patients to the specific cancer and the non-specific LDH virus antigen are similar which suggests a causal factor. RNA of the LDH virus parasites on the oxidative energy system of the cell. Parasitic energy consumption should reduce energy level and coherence of the electromagnetic oscillations. Therefore, the results support a hypothesis that CMI response corresponds to reduced coherence and disturbed excited coherent state far from the thermodynamic equilibrium. The CMI response of T lymphocytes from patients with cervical precancerous lesions to both antigens corresponds to healthy humans or cervical cancer patients which indicates the period of oncogene mutation and suggests that the cell-mediated immune response is elicited in connection with genome mutation.
Massive non-local genome somatic mutations in cancer cells seem to correspond to a substantial increase of randomness of the electromagnetic activity which might elicit the CMI response. A genome somatic mutation can be understood as an interdisciplinary process which displays a failure of cellular control over chemical reactions caused by disturbance of the biophysical mechanisms. Defects of oxidative metabolism result in low level of excitation, diminished coherence/random ratio, increased randomness, and uncontrolled genome mutations. Coherence of biological electromagnetic field enables control of chemical reactions in living cells.
Supplemental content
Download PDF (452.8 KB)Acknowledgements
This work was supported by the Czech Science Foundation, project 16-12757S. The authors pay tribute to their late colleagues Josef Koštíř, Karel Heyberger, Tomáš Hraba, Jiří Čoupek, Josef Sanitrák, Blanka Mejsnarová and Otto Wichterle for their great contributions to the development of materials, facilities for antigen purification, and measurement.
Disclosure statement
The authors report no conflicts of interest. The authors alone are response for the content and writing of the paper.
References
- Alberts B, Bray D, Lewis D, Raff M, Roberts K, Watson JD. 1994. Molecular biology of the cell; 3rd ed. New York & London: Garland Publishing, Inc.
- Alexandrov LB, Nik-Zainal S, Wedge DC, Aparicio SAJR, Behjati S, Biankin AV, Bignell GR, Bolli N, Borg A, Børresen-Dale A-L, et al. 2013. Signature of mutational processes in human cancers. Nature. 500:415–421.
- Brinton MA. 1994. Lactate dehydrogenase-elevating, equine arteritis and lelystad viruses. In: Webster RG, Granoff A, editors. Encyclopedia of virology 2. London/San Diego, CA: Academic Press Ltd. pp. 763–771.
- Dankort D, Curley DP, Cartlidge RA, Nelson B, Karnezis AN, Damsky WE Jr, You MJ, DePinho RA, McMahon M, Bosenberg M. 2009. BrafV600E cooperates with Pten loss to induce metastatic melanoma. Nat Genet. 41:544–552.
- Davies H, Bignell GR, Cox Ch, Stephens P, Edkins S, Clegg S, Teague J, Woffendin H, Garnett MJ, Bottomley W, et al. 2002. Mutations of the BRAF gene in human cancer. Nature. 417:949–954.
- Dhomen N, Reis-Filho JS, da Rocha Dias S, Hayward R, Savage K, Delmas V, Larue L, Pritchard C, Marais R. 2009. Oncogenic Braf induces melanocyte senescence and melanoma in mice. Cancer Cell. 15:294–303.
- Fried SD, Bagchi SD, Boxer SG. 2014. Extreme electric fields power catalysis in the active site of ketosteroid isomerase. Science. 346:1510–1514.
- Fröhlich H. 1969. Quantum mechanical concepts in biology. In: Marois M, editor. Theoretical physics and biology. Proceedings of the 1st Int. Conference on Theoretical Physics and Biology; 26–30 June 1967; Versailles. Amsterdam, London: North Holland; pp. 13–22.
- Fröhlich H. 1978. Coherent electric vibrations in biological systems and cancer problem. IEEE T Microw Theory. 26:613–617.
- Fröhlich H. 1980. The biological effects of microwaves and related questions. In: Marton L, Marton C, editors. Advances in electronics and electron physics, vol. 53. New York: Academic Press; pp. 85–152.
- Hildebrandt P. 2014. More than fine tuning. Science. 346:1456–1457.
- Jandová A, Motyčka K, Čoupek J, Sanitrák J, Heyberger K, Laurová L, Mejsnarová B, Novotná J, Pezlarová J, Macholda F, et al. 1979. [“Diseases” of cellular energy system]. Sb Lek. 81:321–327 [in Czech].
- Jandová A, Hurych J, Pokorný J, Čoček A, Trojan S, Nedbalová M, Dohnalová A. 2001. effects of sinusoidal magnetic field on adherence inhibition of leukocytes. Electro- Magnetobiol. 20:397–413.
- Jandová A, Pokorný J, Čoček A, Trojan S, Nedbalová M, Dohnalová A. 2004. Effects of sinusoidal magnetic field 0.5 mT on leukocyte adherence inhibition. Electromagn Biol Med. 23:81–96.
- Jandová A, Mhamdi L, Nedbalová M, Čoček A, Trojan S, Dohnalová A, Pokorný J. 2005. Effects of magnetic field 0.1 and 0.05 mT on leukocyte adherence inhibition. Electromagn Biol Med. 24:283–292.
- Jandová A, Pokorný J, Kobilková J, Janoušek M, Mašata J. Trojan S, Nedbalová M, Dohnalová A, Beková A, Slavík V, et al. 2009a. Cell-mediated immunity in cervical cancer evolution. Electromagn Biol Med. 28:1–14.
- Jandová A, Pokorný J, Kobilková J, Trojan S, Nedbalová M, Dohnalová A, Čoček A, Mašata J, Holaj R, Tvrzická E, et al. 2009b. Mitochondrial dysfunction. Neural Netw World. 19:379–391.
- Jandová A, Pokorný J, Pokorný J, Kobilková J, Nedbalová M, Čoček A, Jelínek F, Vrba J, Vrba J Jr, Dohnalová A, et al. 2015. Diseases caused by defects of energy level and loss of coherence in living cells. Electromagn Biol Med. 34:151–155.
- Jelínek F, Cifra M, Pokorný J, Vaniš J, Šimša J, Hašek J, Frýdlová I. 2009. Measurement of electrical oscillations and mechanical vibrations of yeast cells membrane around 1 kHz. Electromagn Biol Med. 28:223–232.
- Kasas S, Ruggeri FS, Benadiba C, Maillard C, Stupar P, Tournu H, Dietler G, Longo G. 2015. Detecting nanoscale vibrations as signature of life. Proc Natl Acad Sci USA. 112:378–381.
- Knox SS, Funk RHW. 2014. Oncology and biophysics: a need for integration, J Clin Exp Oncol S1–S001.
- Mejsnarová B, Jandová A, Kupec M, Motyčka K, Heyberger K, Čoupek J, Kobilková J. 1981. [Increased frequency of associations of acrocentric chromosomes brought about by the LDH virus in fertile women]. Sb Lek. 83:332–335 [in Czech].
- Pelling AE, Sehati S, Gralla EB, Valentine JS, Gimzewski JK. 2004. Local nano-mechanical motion of the cell wall of Saccharomyces cerevisiae. Science. 305:1147–1150.
- Pelling AE, Sehati S, Gralla EB, Gimzewski JK. 2005. Time dependence of the frequency and amplitude of the local nanomechanical motion of yeast. Nanomed-Nanotechnol. 1:178–183.
- Plagemann PGW. 1996. Lactate dehydrogenase-elevating virus and related viruses. In: Fields BN, Knipe DM, Howley PM, editors. Virology; 3rd ed. New York: Raven Press; pp. 1105–1120.
- Pokorný J, Jelínek F, Trkal V, Lamprecht I, Hölzel R. 1997. Vibrations in microtubules. J Biol Phys. 23:171–179.
- Pokorný J, Wu T-M. 1998. Biological aspects of coherence and biological order. Heidelberg: Springer.
- Pokorný J, Hašek J, Jelínek F, Šaroch J, Palán B. 2001. Electromagnetic activity of yeast cells in the M phase. Electro- Magnetobiol. 20:371–396.
- Pokorný J, Hašek J, Vaniš J, Jelínek F. 2008. Biophysical aspects of cancer – electromagnetic mechanism. Indian J Exp Biol. 46:310–321.
- Pokorný J, Vedruccio C, Cifra M, Kučera O. 2011. Cancer physics: diagnostics based on damped cellular elastoelectrical vibrations in microtubules. Eur Biophys J. 40:747–759.
- Pokorný J, Pokorný J, Kobilková J. 2013a. Postulates on electromagnetic activity in biological systems and cancer. Integrat Biol. 5:1439–1446.
- Pokorný J, Foletti A, Kobilková J, Jandová A, Vrba J, Vrba J Jr, Nedbalová M, Čoček A, Danani A, Tuszyński JA. 2013b. Biophysical insights into cancer transformation and treatment. Sci World J. 2013:195028.11p.
- Pokorný J, Pokorný J, Kobilková J, Jandová A, Vrba J, Vrba J Jr. 2014. Cancer – pathological breakdown of coherent energy states. Biophys Rev Lett. 9:115–133.
- Pokorný J, Pokorný J, Foletti A, Kobilková J, Vrba J, Vrba J Jr. 2015. Mitochondrial dysfunction and disturbed coherence: gate to cancer. Pharmaceuticals. 8:675–695.
- Sahu S, Ghosh S, Ghosh B, Aswani K, Hirata K, Fujita D, Bandyopadhyay A. 2013. Atomic water channel controlling remarkable properties of a single brain microtubule: correlating single protein to its supramolecular assembly. Biosens Bioelectron. 47:141–148.
- Sahu S, Ghosh S, Fujita D, Bandyopadhyay A. 2014. Live visualizations of single isolated tubulin protein self-assembly via tunneling current: effect of electromagnetic pumping during spontaneous growth of microtubule. Sci Reports. 4:7303.
- Seyfried TN, Shelton LM. 2010. Cancer as a metabolic disease. Nutr Metab. 7:7.
- Streitwieser A. 1962. Molecular orbital theory. New York/London: Wiley.
- Tomasetti C, Vogelstein B. 2015. Variation in cancer risk among tissues can be explained by the number of stem cell divisions. Science. 347:78–81.
- Tsai J, Lee JT, Wang W, Zhang J, Cho H, Mamo S, Bremer R, Gillette S, Kong J, Haass NK, et al. 2008. Discovery of a selective inhibitor of oncogenic B-Raf kinase with potent antimelanoma activity. Proc Natl Acad Sci USA. 105:3041–3046.
- Ventegodt S, Hermansen TD, Flensborg-Madsen T, Nielsen ML, Merrick J. 2006. Human development VIII: a theory of “deep” quantum chemistry and cell consciousness: quantum chemistry controls genes and biochemistry to give cells and higher organisms consciousness and complex behavior. Sci World J. 6:1441–1453.
- Vredeveld LCW, Possik PA, Smit MA, Meissl K, Michaloglou Ch, Horlings HM, Ajouaou A, Kortman PC, Dankort D, McMahon M, et al. 2012. Abrogation of BRAFV600E-induced senescence by PI3K pathway activation contributes to melanomagenesis. Gene Dev. 26:1055–1069.
- Warburg O, Posener K, Negelein E. 1924. Über den Stoffwechsel der Carcinomzelle. [Metabolism of a cancer cell]. Biochem Z. 152:309–344 [in German].
- Warburg O. 1956. On the origin of cancer cells. Science. 123:309–314.