Abstract
The vegetative growth of Ulva lactuca was studied to determine if the growth rate of the alga is driven by infradian rhythmicity. The influence of temperature on the infradian rhythm of growth was also investigated. Discs of Ulva were grown in controlled laboratory conditions at different combinations of temperature (5, 10, 15, 20°С) and irradiance (40 and 60 μmol photons m−2 s−1) under 12 : 12 h light : dark cycles. The growth rates exhibited a rhythmic pattern with one major peak every 2 or 3 days. Growth at 5 or 10°С increased the prevalence of 3-day cycles and maintained U. lactuca in the vegetative growth stage. In contrast, growth at 15 or 20°С provoked a predominance of the 2-day cycle and induced reproduction. The 2- or 3-day cycles were combined in longer cycles having a period close to 6 days. We suppose that the 2-, 3- and 6-day rhythms of physiological processes are related to large-scale Rossby and Kelvin waves, which produce oscillations in the geomagnetic field and seawater temperature with the same periods. The predominance of 2-day or 3-day fluctuations of the geomagnetic field and temperature probably determine the prevalence of reproduction and vegetative growth, respectively, in Ulva.
Introduction
According to modern opinion, there are three types of endogenous rhythms, namely ultradian (with periods of several hours or much less), circadian (with periods between 20 and 28 h), and infradian (with periods longer than 28 h) (Koukkari & Sothern, Citation2006). Various schemes have been proposed for dividing the infradian rhythms further into subgroups, such as periods of about 2 days, half a week (circasemiseptan), 1 week (circaseptan), 10 days, 14 days (circasemilunar), 1 month (circatrigintan), 1 year (circannual), and even longer (Reinberg & Smolensky, Citation1983; Spruyt et al., Citation1987; Yamashita et al., Citation1999; Chumakov et al., Citation2000; Buda et al., Citation2003; Halberg et al., Citation2003; Vignoles et al., Citation2006).
There is evidence for the control of physiological processes in animals by the short-tern infradian clock, for example, infradian rhythms with periods of 2 to 7 days occur in bivalves, rats, ducks, horses and humans (Kennish & Olsson, Citation1975; Temuryants et al., Citation1995; Marques et al., Citation1996; Thébault et al., Citation2006; Díaz-Sandoval et al., Citation2008; Yarmolyuk, Citation2010). There are also some reports of infradian rhythms in higher plants; clear examples are the description of circaseptan rhythms of water uptake in Phaseolus vulgaris seeds (Spruyt et al., Citation1987) and circumnutation intensity in Helianthus annuus (Buda et al., Citation2003). However, information about such infradian rhythms in seaweeds is sparse and the majority of reports concern reproduction. For example, a semilunar rhythmicity of gamete release occurs in Fucus vesiculosus (Andersson et al., Citation1994); a weekly or 5-day reproductive rhythm occurs in Ulva lactuca [=U. pseudocurvata] (Lüning et al., Citation2008) and U. lactuca [=U. fenestrata] (Kalita & Titlyanov, Citation2011), respectively, and a 4- to 5-day interval between formation of gametangia occurs in Derbesia marina (Ziegler-Page & Kingsbury, Citation1968). A semilunar rhythm in melatonin accumulation was also detected recently in Ulva sp., with maximum melatonin content correlating with minimum sea level (Tal et al., Citation2011); this rhythm triggers a priori antioxidant mechanisms enabling the organisms to successfully adapt to stress conditions (desiccation, higher radiation, salinity and temperature) in the intertidal-zone.
Reports of 2-day periodicity in the biological rhythms of living organisms, including algae, are rare. However, some 2-day rhythms have been identified: in microstructural growth of the bivalve Mercenaria mercenaria (Kennish & Olsson, Citation1975), in stria formation by the tropical scallop Comptopallium radula (Thébault et al., Citation2006), and in division and degradation of the zooxanthellae in the scleractinian coral Seriatopora caliendrum, Stylophora pistillata and the colonial hydroid Milllepora intricata (Titlyanov et al., Citation2004, Citation2006). Recently we reported that infradian rhythms (with periods of 2 or 3 and 6 days) of growth rate and cell division occur in U. lactuca [= U. pseudocurvata] and Porphyra umbilicalis from the North Sea (Helgoland Island, Germany, 53º56′N, 14º05′E) (Kalita et al., Citation2007) and we hypothesize that such rhythms will be found in algae from different geographical areas and may be global phenomena. On the Russian Far East coast the genus Ulva is represented by U. lactuca [=U. fenestrata]; little is known about the infradian rhythm of the growth rate of thalli in U. lactuca (Titlyanova, Citation1990; Kepel, Citation1999); and examination of this rhythm was the first objective of our study.
According to the majority of scientists semilunar and weekly rhythms are entrained by moonlight and (or) lunar gravity (Franke, Citation1985; Neumann & Heimbach, Citation1985; Spruyt et al., Citation1987; Neumann, Citation1989; Andersson et al., Citation1994; Lüning et al., Citation2008). However, there is no easy explanation for the origin of infradian modulation with periods of less than 1 week. Although the presence of quasi-2-day waves in the mesosphere and thermosphere () is well-known to geophysicists (Muller, Citation1972; Salby, Citation1981; Palo et al., Citation1999; Limpasuvan et al., Citation2000), as a rule, infradian biological rhythms are related to solar activity with periods close to 3.5 and 7 days (Díaz-Sandoval et al., Citation2008); or with 2- and 3.5-day fluctuations of sea level pressure and wind velocity (Delyukov et al., Citation2001; Thébault et al., Citation2006). Two-day variations of meteorological and oceanographic parameters, including sea surface temperature, sea level pressure, and wind velocity, have been observed in the Pacific Ocean (Kenyon, Citation1996). Kenyon (Citation1996) found that all meteorological properties at sea level across the Pacific along 35°N were ‘based on the correlation between air temperature and the north-wind velocity component, which was consistent with a net pole-ward heat flux all across the ocean’, and this was related to the heating and cooling by the sun. In other words the atmosphere may be partially adjusting its heat balance by horizontal advection within a period of 2 days (Kenyon, Citation1996). This hypothesis has been confirmed by investigations of the 2-day temperature wave in the upper atmosphere (Wu et al., Citation1996; Limpasuvan et al., Citation2000). Bearing this information in mind, a second objective of our study was to investigate primarily the influence of temperature, but also irradiation and photoperiod on the growth rate kinetics of U. lactuca.
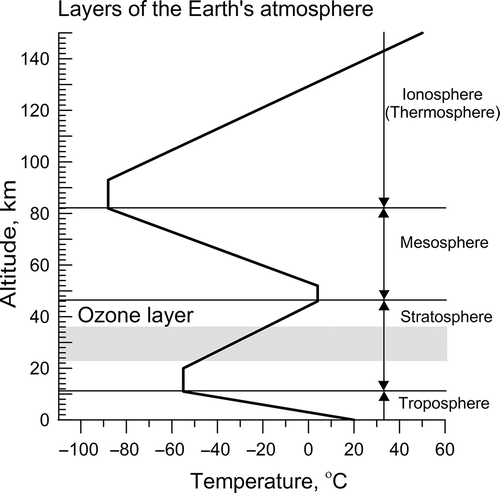
Figs 1. The Earth's atmosphere may be divided into several distinct layers. The troposphere is where all weather takes place; it is the region of rising and falling packets of air. The troposphere is known as the lower atmosphere. Above the troposphere is the stratosphere, where air flow is mostly horizontal. The ozone layer, which absorbs and scatters the solar ultraviolet radiation, is in this layer. Above the stratosphere is the mesosphere and, together, the stratosphere and the mesosphere comprise the middle atmosphere. The ionosphere (or thermosphere) starts just above the mesosphere and extends to 600 km altitude, where many atoms are ionized (have gained or lost electrons so they have a net electrical charge); this layer is known as the upper atmosphere. The regions of mesosphere and lower thermosphere are called the MLT region. The line within the graph shows the temperature changes with altitude. The diagram of website http://http://meteoweb.ru/phen058.php was modified by authors.
Materials and methods
Organism and cultivation conditions
Thalli of Ulva lactuca (Chlorophyta) were collected from upper subtidal habitat in Amursky Bay, in the north-west of the Japan Sea (43º10′N, 132º10′E) at a depth of 3–4 m in March. At the time of collection, the Ulva was growing under a canopy of Zostera marina and Sargassum miyabei, where the light intensity was 15% of the incident surface photosynthetic active radiation (PAR0) or about 60 μmol photons m−2 s−1 in the afternoon of an average bright day. Adult plants of U. lactuca were acclimated to a temperature of 15ºС, photon irradiance of 40 μmol m−2 s−1, and 12 h light per day (start of light phase at 06.00 h) for 2 weeks. The algae were illuminated by cool-white fluorescent lamps (LD-20, 40W). Photon flux density was measured with a LI-190SA cosine quantum sensor (Lambda Instruments Corporation, Lincoln, Nebraska). Thalli of U. lactuca were cultivated in aerated 5000 ml glass beakers in enriched seawater (PES; Provasoli, Citation1968) and the medium was changed weekly.
Design of experiment 1. Effects of temperature and irradiation on the infradian rhythm of growth rate
A total of 48 discs of 20 mm diameter were punched from marginal portions of the blade thalli (excluding thallus portions near the holdfast area), transferred into aerated 1000-ml glass beakers with PES medium, and pre-cultivated for 1 week with different combinations of temperature (5, 10, 15, 20ºС) and irradiance (40 or 60 μmol photons m−2 s−1) in 12 h light per day (). Fresh weight of each disc was measured by torsion balance (WAGA Torsyjna-WT, Poland, 0.0001 g) daily at 10.00 h for 25 days, and then every third day for 3 months after the 25 days. Each disc was blotted with blotting paper to remove excess seawater, weighed, and placed back into a glass beaker with the medium. This procedure was performed three times with each disc, and the average weight of each disc was calculated. Although we were aware of the intrinsic problems with determining growth from fresh weight changes, this was the only possible way to determine growth in our experiments, because we needed to work with live cells. For each disc, information on the following morphological stages was recorded: vegetative thallus, light-green margin, formation of spores or gametes, and release of spores or gametes (Titlyanova, Citation1990).
Table 1. Fresh weight of discs of Ulva lactuca grown at different temperatures, irradiances, and photoperiods. ‘–’ = all cells in the discs were converted into reproductive tissue
Design of experiment 2. Effects of photoperiod on the infradian rhythm of growth rate
Nine adult plants of U. lactuca consisting of three male gametophytes, three female gametophytes and three sporophytes (selection based on size of reproductive cells) were selected. From marginal portions of each thallus nine discs of 15 mm were punched (total 81 discs). These stock discs were immediately transferred into separate aerated 400 ml glass beakers of PES: for each plant six discs were transferred to one beaker and three to another, this was repeated for all plants resulting in two groups of samples (). The first group of nine beakers each containing six discs from each plant were cultivated under a photoperiod of 8 h light : 16 h dark (8 : 16 L : D) and the second group of nine beakers containing three discs from each plant under 12 : 12 L : D. These samples were pre-cultivated under a temperature of 5°C and 40 µmol photons m−2 s−1 for 1 week. No reproduction occurred under these conditions as determined in experiment 1.
Figs 2. Design of experiment 2: effects of photoperiod on the infradian rhythm period of growth rate in Ulva lactuca (for explanation, see text).
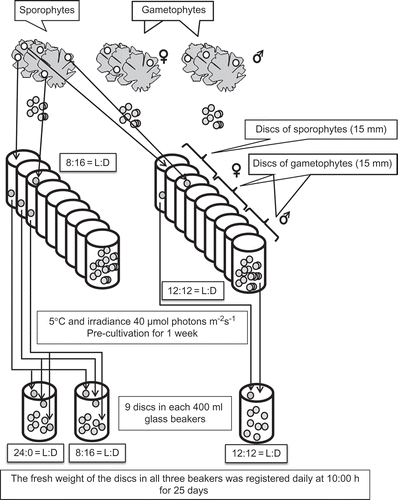
After pre-cultivation one disc was taken from each beaker in the 12 : 12 L : D group and combined into one sample; this beaker was maintained under the same photoperiod conditions for the remainder of the experiment. From the 8 : 16 photoperiod group one disc from each beaker was combined as above and maintained at the same photoperiod, whilst a second combined sample containing one disc from each beaker was cultivated under continuous white light (24 : 0 L : D), from the tenth day of the experiment onwards. This resulted in three aerated 400 ml beakers; each containing nine discs (three discs each of sporophyte, male gametophyte and female gametophyte thalli from nine different plants). In order to follow the growth of a disc from a particular plant, the discs were differentiated in each beaker by marking them with small holes (1 mm in diameter). The fresh weight of the discs in all three beakers was measured daily at 10.00 h (as in experiment 1) for 25 days.
Relative growth rates were calculated as the increase in algal fresh weight assuming exponential growth, μ = [{ln(w 2) – ln(w 1)} ∕ (t 2 − t 1)] × 100%, where w 1 and w 2 are the algal biomass before t 1 and after t 2 days of growth.
Statistical analysis
Statistical analysis was performed using the software Statistica 6.0 (StatSoft, Tulsa, Oklahoma, USA). Comparisons of growth rates between different samples were made using t-tests for independent samples. Standard deviations were calculated from six or nine replicates (according to experimental design) and indicated by vertical bars in the figures or ± values in the table. In order to determine whether a periodicity in the time series data existed or not, a Fourier spectral analysis was performed. To verify the correctness of the Fourier spectral analysis, Box–Ljung's Q-test of autocorrelation was used. Also, each time series was tested as a white noise series by producing a histogram of the periodogram values and fitting the exponential distribution to the histogram. If the observations in the time series are independent of each other (i.e. there is no periodicity) and they follow the normal distribution, such a time series is referred to as a white noise series. The periodogram values can be interpreted in terms of variance (sums of squares) of the data at the respective frequency or period. The periods (defined as the length of time required for one full cycle) of the time series data were delineated as the two largest periodogram or spectral density values.
The influence of temperature, irradiance and photoperiod on the length of growth cycles was estimated using a factorial ANOVA with P < 0.05 as the level of significance. The percentage of influence of each factor on the length of the growth cycle was evaluated by the Snedecor function (Snedecor, Citation1961).
The approximation of biomass curves by a trend line was carried out in Microsoft Excel. Adequacy of the trend lines was estimated based on the coefficient of determination, R 2, which is an indicator of how well a model fits the data (e.g. an R 2-square close to 1.0 indicates that almost all of the variability with the variables specified in the model has been accounted for).
Results
Effects of temperature, irradiance and photoperiod on disc biomass
The weights of the U. lactuca discs gradually increased in all treatments (). At the end of the experiment the maximum biomass was obtained with discs at 10ºС, and a decline or rise in water temperature by 5ºС caused an average reduction in biomass of 40% and 20% respectively (). The effect of irradiance on biomass was minimal at 10ºС; at this temperature, increasing the light intensity from 40 to 60 μmol photons m−2 s−1 increased the biomass on average by 13.5%. In other temperature conditions, the increase of light intensity from 40 to 60 μmol photons m−2 s−1 produced a 40% or greater increase in disc weight.
Figs 3. Effects of different temperature and irradiance on the fresh weight of Ulva lactuca discs in the laboratory throughout the experimental period with a 12 : 12 h light–dark regime.
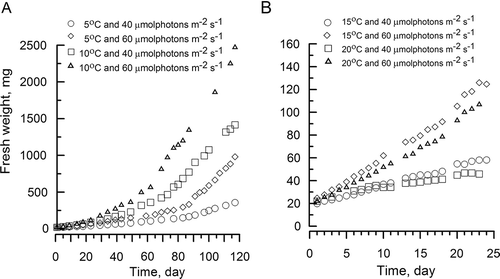
For discs incubated at 5 or 10°C, the relationship between biomass (m, mg) and time (t, day) was fitted to parabolic equations (). The accretion of biomass was maximal at 10ºC and 40 μmol photons m−2 s−1 (m 10/40 = 0.12t 2 − 2.45t + 59.765, R 2 = 0.99) and was minimal at 5ºC and 40 μmol photons m−2 s−1(m 5/40 = 0.03t 2 – 0.86t + 40.38, R 2 = 0.98). The discs remained in the vegetative stage throughout the experiment (117 days) at 5ºC with 40 and 60 μmol photons m−2 s−1, and at 10ºC with 40 μmol photons m−2 s−1. For discs incubated at 15 or 20°C, the relationship between biomass and time was fitted to linear equations (). At these higher temperatures, all cells in the discs were entirely converted into reproductive tissue by day 29. The maximum number of fertile cells per reproduction was obtained at 15ºC and 40 μmol photons m−2 s−1 (m 15/40 = 1.73t + 18.30, R 2 = 0.99).
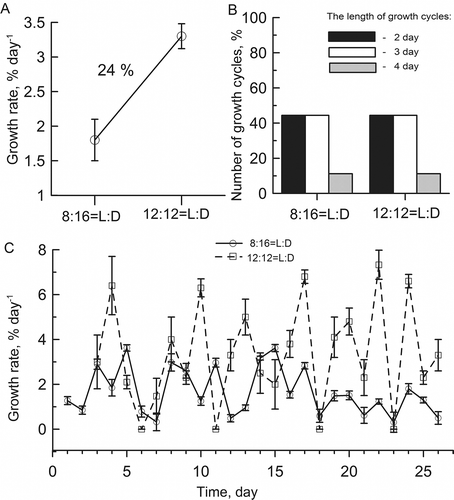
Disc weights increased faster at 12 h light per day than at 8 h light per day (). Under continuous white light (40 μmol photons m−2 s−1) and constant temperature (5ºC), discs increased only 2.9% in weight and died after 15 days.
Growth rate periodicity in light/dark conditions
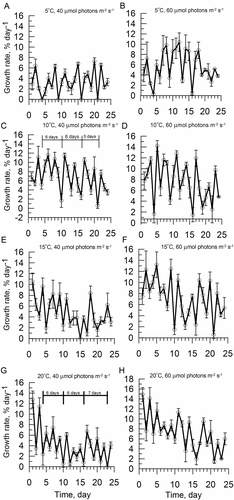
Growth rate, in terms of the rate of biomass increase, exhibited a rhythmic pattern with one major peak every 2 or 3 days (). Growth at 5 and 10ºС caused an increase in the prevalence of 3-day cycles and maintained Ulva lactuca in the vegetative growth stage (–D). Fertile tissue (about 6% of disc area) appeared mainly at 10°C and 60 μmol photons m−2 s−1. Growth cycles of 2 and 3 days occurred with equal frequency at 5°C and 40 μmol photons m−2 s−1 (). Four-day cycles of growth rate were seldom found, occurring more at 60 μmol photons m−2 s−1 (, D). In contrast, exposure at 15 and 20ºС provoked a predominance of 2-day cycles and induced reproduction after 5 days of cultivation (, F). The 2-day cycles accounted for 56–90% of all cycles detected in the growth rate curves at 15 and 20ºС (, C). In these conditions, reproduction prevailed over vegetative growth of discs; hence the biomass of discs declined after each release of gametes or spores.
Figs 4. Infradian rhythms of relative growth rate in Ulva lactuca with a 12 : 12 h light–dark cycle. Vertical bars indicate standard deviation (n = 6).
Figs 5. Effects of temperature on the period of growth rate (A) and the percentage ratio of growth cycles at 40 (B) and 60 (C) μmol photons m−2 s−1 in Ulva lactuca with a 12 : 12 h light–dark regime.
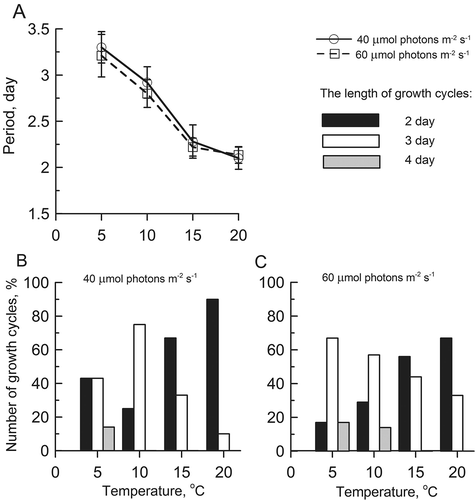
Short 2- or 3-day cycles were combined in larger cycles having a period close to 6 days (). Each of the long cycles (6 days) consisted of either three 2-day cycles or two 3-day cycles (, G). Cycles of 5 and 7 days were also observed, and were formed by combinations of the shorter 2- and 3-day cycles (, G).
Cycle length decreased significantly (factorial ANOVA, Р < 0.01) from 3.3–2.8 days at 5 and 10ºС to 2.2 days at 15 and 20ºС () and the shift was not influenced by the photoperiod (; the ratios of 2- and 3-day growth cycles were identical at 12 and 8 h light per day). The length of the growth cycles was modulated by temperature and light intensity, which accounted for 43% and 30% of the variability, respectively, as indicated by a 2-way factorial ANOVA (Supplementary ). No significant changes in the length of the growth cycles were observed with increasing day length (P < 0.05) (Supplementary Table 2). The prolongation of the photoperiod from 8 to 12 h altered the growth rate amplitude, such that the growth accelerated as the light phase lengthened (, C) (Supplementary Table 3).
Figs 6. Effects of photoperiod on the infradian rhythm period of growth rate in Ulva lactuca at 5°C, 40 μmol photons m−2 s−1 (see Fig. 4 for the curve for a light–dark cycle of 12 : 12 h). The average growth rate was 24% higher at 12 h than at 8 h light per day (A). No significant changes in the percentage ratio of cycles were observed with increasing day length (P < 0.05) (B). The prolongation of the photoperiod from 8 to 12 h altered the growth rate amplitude, such that the growth accelerated as the light phase lengthened (C).
Growth rate periodicity in continuous light and temperature
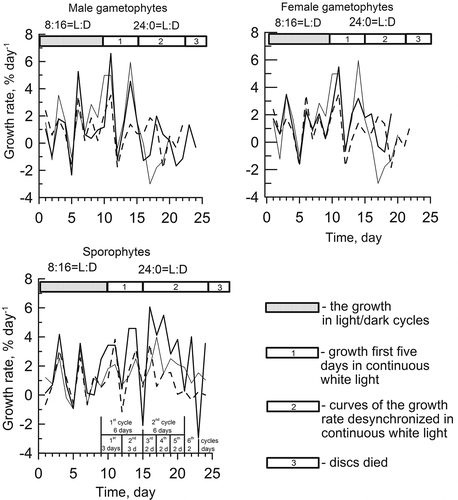
After transfer from light–dark cycles to continuous light, infradian rhythms of growth with periods of 2 or 3 and 6 days persisted through either 5 or 2 cycles, respectively (). Growth trends in all discs were similar to the growth in light–dark during the first 5 days at 40 μmol m−2 s−1 in continuous white light. On the sixth day the growth trends desynchronized, and between the 10th and 14th day of continuous light, the discs died for unknown reasons (). The growth rates of gametophytes and sporophytes were not significantly different (P < 0.05). However, discs of gametophytes died more quickly than those of sporophytes.
Figs 7. Infradian rhythms of growth of different generations of Ulva lactuca in continuous white light at 5°С and 40 μmol m−2 s−1. Each line in the figures represents the curve of growth rate of disc from one plant.
Discussion
Periodic fluctuations in the geomagnetic field: Rossby and Kelvin waves
Our experimental results confirm those of previous studies of U. pseudocurvata and Porphyra umbilicalis (Kalita et al., Citation2007), and zooxanthellae (Titlyanov et al., Citation2004, Citation2006), viz. that the physiological processes (growth, cell division, division and degradation of zooxanthellae) of simply organized algae are driven by biological clocks on 2- or 3- and 6-day scales. A 2-day periodicity has also been observed in stria formation of the tropical scallop Comptopallium radula by Thébault et al. (Citation2006), who suggested that this ‘may be controlled by an endogenous oscillator, synchronized by an environmental cue acting as a Zeitgeber, such as seawater temperature or sea level pressure, both of which exhibit 2-day variations in the Pacific Ocean’. In our opinion, the 2- or 3- and 6-day rhythms of physiological processes in algae could possibly be entrained by 2- or 3- and 6-day fluctuations of the Earth's geomagnetic field. This conclusion is based on the following observations from geophysicists and oceanographers.
Firstly, the large-scale circulation of the atmosphere and ocean is dominated by wave motions caused by the Earth's rotation. These are known as Rossby waves (oceanic and atmospheric). The atmospheric Rossby waves usually have periodicities of 2, 6, 10 and 16 days (Talaat et al., Citation2001; Manson et al., Citation2005; Pancheva et al., Citation2006). Only the 2- and 6-day waves propagate vertically from the stratosphere to the mesosphere/lower thermosphere (MLT) region and reach the ionosphere () (Pancheva et al., Citation2006, Citation2008). In the MLT region, the 6-day Rossby wave reaches maxima just before and just after the equinoxes (Talaat et al., Citation2001; Pancheva et al., Citation2008) and minimizes at solstices (Talaat et al., Citation2001). The wave amplitudes at the equinoxes are smaller than the peaks before and after, but are still larger than the wave amplitudes at solstices (Liu et al., Citation2004). Most scientists suggest that the 2-day Rossby wave is a summer phenomenon in the middle atmosphere, occurring mainly at low- and mid-latitudes, and reaching its maximum amplitude during July and August in the northern hemisphere. A subsidiary maximum of about 70% of the summer peak appears during April–May (Thayaparan et al., Citation1997; Riggin et al., Citation2004). Moreover, the 2-day wave activity seen in the ionospheric currents is forced by the simultaneous 2-day wave activity in the MLT region (Pancheva et al., Citation2006), and the largest enhancements of the oscillation occur during early June, July and early August (Altadill et al., Citation1998).
Secondly, along the equator and along lateral vertical boundaries, such as coastlines or density gradients, a different class of waves known as Kelvin waves (oceanic and atmospheric) is found. The atmospheric Kelvin waves are usually classified as follows: ‘slow’ waves with periods of 10–20 days, ‘fast’ waves with periods of 6–10 days, and ‘ultra-fast’ waves with periods of 3–4 days (Pancheva et al., Citation2008). Like Rossby waves, Kelvin waves originate in the troposphere and propagate through the stratosphere to the MLT region. However, Mayr et al. (Citation2004) proposed that a significant portion of the waves observed in the mesosphere may be excited in situ. The amplitudes and prominent periods of atmospheric Kelvin waves depend on the altitude and season. It is known that the dominant Kelvin waves change from long periods (5–10 days) and short wavelengths (9–13 km) in the stratosphere, to shorter periods (2–3 days) and longer wavelengths (35–45 km) in the MLT region. Ultra-fast Kelvin waves exist intermittently during all months of the year in the MLT region (Forbes et al., Citation2009). What is important is that 3- to 4-day waves have larger amplitudes during winter to spring (Hitchman & Leovy, Citation1988; Yoshida et al., Citation1999). Six-day Kelvin waves amplify during two intervals: in August and in the second half of September to early November (Pancheva et al., Citation2008).
Oceanic Rossby and Kelvin waves certainly contribute to the intensification of oceanic circulation, but they are not interesting for us because these waves have a large wavelength (of the order of 1000 km), slow propagation (of the order of 1–10 km per day), and a period of more than 70 days (Cipollini et al., Citation1997; Hendon et al., Citation1998; Quartly et al., Citation2003; Wu et al., Citation2008).
Thirdly, it is known that if a global-scale wave with large amplitude and fairly long wavelength propagates from the MLT region of Earth's atmosphere into the ionosphere, it should cause an electric-current system that follows the period of the global-scale wave (Altadill et al., Citation1998; Forbes, Citation2000; Pancheva et al., Citation2006, Citation2008). This wave-like current system produces perturbations in the geomagnetic field that are easily measured at ground level. It is very likely that such geomagnetic perturbations arise from 6- and 3-day Kelvin waves (Forbes, Citation2000; Pancheva et al., Citation2008) and from 6- and 2-day Rossby waves (Altadill et al., Citation1998; Pancheva et al., Citation2006, Citation2008).
Finally, atmospheric Kelvin and Rossby waves cause oscillations in pressure, temperature, and winds or currents (Wheeler, Citation2002). The magnitude of such oscillations is large enough to influence large-scale weather patterns (Wheeler, Citation2002). For instance, in 1996 Kenyon described 2-day variation of meteorological properties (sea surface temperature, air temperature, sea level pressure, wind velocity, total cloud amount and visibility) in the Pacific Ocean (along 35°N from California to Japan) and suggested that Rossby waves can produce these oscillations (Kenyon, Citation1996).
Thus, on the one hand, large-scale 2- and 6-day Rossby and 3- and 6-day Kelvin waves generate oscillations in the Earth's geomagnetic field with analogous periods, and the dominant period of these oscillations depends on the season. These geomagnetic oscillations may be used directly by algae and may be the main Zeitgeber signal of the infradian clock (similar to light–dark cycles in circadian and circannual rhythms), synchronizing physiological processes. On the other hand, Rossby and Kelvin waves produce oscillations of meteorological properties with dominant seasonal periods that include temperature fluctuations. The temperature oscillations seem to serve as the secondary Zeitgeber signal of the infradian clock, determining the ontogenetic stage (which is true for circadian and circannual rhythms).
It is now established that electromagnetic fields, even if very weak, can have an influence on basic biological functions, such as circadian rhythmicity of Drosophila’s locomotor activity (Yoshii et al., Citation2009), orientation and navigation in birds (Wiltschko & Wiltschko, Citation2005; Rodgers & Horea, Citation2009; Wu & Dickman Citation2012) and larval development of the Japanese newt, Cynops pyrrhogaster (Asashima et al., Citation1991). It is also known that electromagnetic shielding causes sharp phase shifts and amplitude changes of infradian rhythmicity of the pain sensitivity in land snails Helix albescens (Kostyuk & Temuryants, Citation2009) and of mobility in the planarian Dugesia tegrina (Yarmolyuk, Citation2010). Low and ultra-high frequency electromagnetic fields also alter infradian rhythmicity of physiological processes; these electromagnetic oscillations play a role as an external synchronizer at the different types of desynchronosis (desynchronosis is the phase shift of any rhythm in the organism with respect to its other rhythms). For example, an ultra-high frequency electromagnetic field corrects and synchronizes the infradian rhythmicity of neutrophil functional activity in intact rats (Temuryants et al., Citation2007). Thus, the geomagnetic field can be registered and, perhaps, utilized by living organisms for synchronizing infradian rhythmicity of physiological processes (Zhadin, Citation2001; Galland & Pazur, Citation2005; Temuryants et al., Citation2007).
A phenomenon particularly relevant to this study is magnetoreception in plants, reviewed in detail by Galland & Pazur (Citation2005); the authors described two mechanisms for magnetoreception: (1) modulation of chemical reaction rates (radical-pair mechanism) and (2) modulation of transport rates and binding by the ion cyclotron resonance mechanism. However, the influence of electromagnetic fields on the temporal organization of living organisms and, in particular, on the infradian rhythms of macroalgae remains virtually unexplored.
Application to Ulva infradian rhythmicity
As mentioned earlier, 2-day Rossby waves dominate at mid-latitudes of the northern hemisphere in summer (Altadill et al., Citation1998) and produce oscillations in the geomagnetic field and water temperature (Kenyon, Citation1996; Pancheva et al., Citation2006, Citation2008). Possibly such oscillations are superimposed on the seawater temperature at 15°C and above and generate the 2-day rhythms of physiological processes in Ulva. The appearance of 2-day rhythms may be an indicator, for Ulva, of favourable conditions for reproduction. At other times of the year, the predominance of 3- to 4-day Kelvin waves may coincide with seawater temperatures ranging from −1.8 to +12°C and cause a 3-day rhythm of physiological processes in Ulva. A 3-day rhythm may thus be a marker of unfavourable conditions, causing vegetative growth to dominate over reproduction in Ulva. Thus, we suppose that the dominance of 2-day versus 3-day fluctuations of the geomagnetic field and temperature may determine the balance of reproduction versus vegetative growth in Ulva and possibly other simply organized algae.
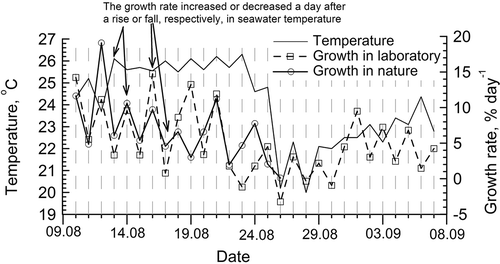
Our hypothesis can explain certain observations on the growth and reproduction of algae. For example, higher minimum light (10 µmol photons m−2 s−1) has been found to be required for reproduction versus vegetative growth (5 µmol photons m−2 s−1) of Ulva pertusa (Han et al., Citation2003). The fertility of Phyllariopsis purpurascens (Phaeophyta) is controlled by temperature such that the maximum sorus surface area coincides with the peak temperature (Flores-Moya et al., Citation1993). Precisely such conditions conducive to reproduction occur in summer months, when the 2-day temperature fluctuation exists. If ambient conditions are unfavourable (when 3- to 4-day temperature oscillations dominate), the period of vegetative growth is increased and reproduction cycles are avoided. Switching from shorter to longer reproductive cycles at the transition from the warmer to the colder season has been shown in detail in Ulva pseudocurvata (Lüning et al., Citation2008), U. lactuca (Kalita & Titlyanov, Citation2011), and Derbesia marina (Ziegler-Page & Kingsbury, Citation1968; Ziegler-Page & Sweeney, Citation1968). In our pilot study, the seawater temperature fluctuated with a 2-day period and amplitude of 0.1–4°C in August to early September 2001 (). The growth rate of Ulva in nature and in the laboratory increased or decreased a day after a rise or fall, respectively, in seawater temperature; therefore, it is possible that temperature synchronizes the infradian oscillations of physiological processes in algae. We cannot be certain of this, but in several groups of organisms, including plants, fungi and vertebrates, very small temperature differences of 0.7–2.0°C can be sufficient to entrain circadian oscillations (Rensing & Ruoff, Citation2002; Ruoff & Rensing, Citation2004). Moreover, according to Dainton (Citation1954), a change of 0.1°C is able to synchronize the circadian rhythm in a slug, Agrolimax reticulatus. On the other hand, if environmental factors oscillate at very low amplitudes and modulate the rhythmic behaviour of biological systems, it will be very important to ensure that experimental conditions are very precisely controlled.
Figs 8. Temperature fluctuations of seawater in the upper sublittoral zone of Amurski Bay (Japan Sea) in nature in August–September 2001 and the growth rate of Ulva lactuca in nature and the laboratory. The growth rate increases on the day after the seawater temperature rises and vice versa, both in nature and in the laboratory. A great drop in temperature on 25–26 August was caused by a typhoon.
To summarize: we have found short infradian rhythms of growth in Ulva, described the relationship between these rhythms and temperature, and identified the probable Zeitgeber for these rhythms in nature. Further investigation will reveal whether geomagnetic field fluctuations might synchronize the infradian rhythms in algae. The most interesting issue will be to determine the origin of infradian rhythms: will these rhythms persist in a constant electromagnetic field?
Acknowledgements
We are grateful for the helpful comments of Professor Mario Giordano and two referees. This research was partially supported by the Russian Foundation of Basic Research 12-04-01424.
References
- Andersson , S. , Kautsky , L. and Kalvas , A. 1994 . Circadian and lunar gamete release in Fucus vesiculosus in the atidal Baltic Sea . Marine Ecology Progress Series , 110 : 195 – 201 .
- Altadill , D. , Solea , J.G. and Apostolov , E.M. 1998 . First observation of quasi-2-day oscillations in ionospheric plasma frequency at fixed heights . Annales de Geophysicae , 16 : 609 – 617 .
- Asashima , M. , Shimada , K. and Pfeiffer , C.J. 1991 . Magnetic shielding induces early developmental abnormalities in the newt, Cynops pyrrhogaster . Bioelectromagnetics , 12 : 215 – 224 .
- Buda , A. , Zawadzki , T. , Krupa , M. , Stolarz , M. and Okulski , W. 2003 . Daily and infradian rhythms of circumnutation intensity in Helianthus annuus . Physiologia Plantarum , 119 : 582 – 589 .
- Chumakov , V.I. , Soldatov , A.A. and Dymochkin , V.N. 2000 . Does the 48-hour biological rhythm exist? . Human Physiology , 26 : 123 – 128 .
- Cipollini , P. , Cromwell , D. , Jones , M.S. , Quartly , G.D. and Challenor , P.G. 1997 . Concurrent altimeter and infrared observations of Rossby wave propagation near 34 in the Northeast Atlantic . Geophysical Research Letters , 24 : 889 – 892 .
- Dainton , B.H. 1954 . The activity of slugs: I. The induction of activity by changing temperatures . Journal of Experimental Biology , 31 : 87 – 110 .
- Delyukov , A. , Gorgo , Yu. , Cornélissen , G. , Otsuka , K. and Halberg , F. 2001 . Natural environmental associations in a 50-day human electrocardiogram . International Journal of Biometeorology , 45 : 90 – 99 .
- Díaz-Sandoval , R. , Sánchez-de la Peñab , S. and Chávez-Negretec , A. 2008 . Seven and 3.5-day rhythms in the incidence of myocardiopathies in Mexico . Archives of Medical Research , 39 : 134 – 138 .
- Flores-Moya , A. , Fernandez , J.A. and Niell , F.X. 1993 . Reproductive phenology, growth and primary production of Phyllariopsis purpurascens (Phyllariaceae, Phaeophyta) from the straits of Gibraltar . European Journal of Phycology , 28 : 223 – 230 .
- Forbes , J.M. 2000 . Wave coupling between the lower and upper atmosphere: case study of an ultra-fast Kelvin Wave . Journal of Atmospheric and Solar-Terrestrial Physics , 62 : 1603 – 1621 .
- Forbes , J.M. , Zhang , X. , Palo , S.E. , Russell , J. , Mertens , C.J. and Mlynczak , M. 2009 . Kelvin waves in stratosphere, mesosphere and lower thermosphere temperatures as observed by TIMED/SABER during 2002–2006 . Earth Planets Space , 61 : 447 – 453 .
- Franke , H.D. 1985 . On a clocklike mechanism timing lunar-rhythmic reproduction in Typosyllis prolifera (Polychaeta) . Journal of Comparative Physiology A , 156 : 553 – 561 .
- Galland , H. and Pazur , F. 2005 . Magnetoreception in plants . Journal of Plant Research , 118 : 371 – 389 .
- Halberg , F. , Cornelissen , G. , Wang , Z. , Wan , C. , Ulmer , W. , Katinas , G. , Singh , R. , Singh , R.K. , Gupta , B.D. , Singh , R.B. , Kumar , A. , Kanabrocki , E. , Sothern , R.B. , Rao , G. , Bhatt , M.L.B. , Srivastava , M. , Rai , G. , Singh , S. , Pati , A.K. , Nath , P. , Halberg , F. , Halberg , J. , Schwartzkopf , O. , Bakken , E. and Shastri , S.V. 2003 . Chronomics: circadian and circaseptan timing of radiotherapy, drugs, calories, perhaps nutriceuticals and beyond . Journal of Experimental Therapeutics and Oncology , 3 : 223 – 260 .
- Han , T. , Han , Y. , Kim , K. , Kim , J. , Shin , H. , Kain , J.M. , Callow , J.A. and Callow , M.E. 2003 . Influences of light and UV-B on growth and sporulation of the green alga Ulva pertusa Kjellman . Marine Ecology Progress Series , 290 : 115 – 131 .
- Hendon , H.H. , Liebmann , B. and Glick , J.D. 1998 . Oceanic Kelvin waves and the Madden–Julian oscillation . Journal of the Atmospheric Sciences , 55 : 88 – 101 .
- Hitchman , M.H. and Leovy , C.B. 1988 . Estimation of the Kelvin wave contribution to the semiannual oscillation . Journal of the Atmospheric Sciences , 45 : 1462 – 1475 .
- Kalita , T.L. and Titlyanov , E.A. 2011 . The effect of temperature on infradian rhythms of reproduction in Ulva fenestrata Postels et Ruprecht, 1840 (Chlorophyta: Ulvales) . Russian Journal of Marine Biology , 37 : 52 – 61 .
- Kalita , T.L. , Titlyanova , T.V. and Titlyanov , E.A. 2007 . New rhythmic changes in mitosis and growth in low differentiated green and red marine macroalgae . Russian Journal of Marine Biology , 33 : 245 – 252 .
- Kennish , M.J. and Olsson , R.K. 1975 . Effects of thermal discharges on the microstructural growth of Mercenaria mercenaria . Environmental Geology , 1 : 41 – 64 .
- Kenyon , K.E. 1996 . Bi-daily variation of meteorological properties at sea level across the Pacific along 350N . Atmospheric Research , 43 : 31 – 46 .
- Kepel , A.A. 1999 . Seasonal dynamics of rocky intertidal macrophytobenthos in the mouth of a channel of lake Ptichye (southwestern Peter the Great Bay, Sea of Japan) . Russian Journal of Marine Biology , 25 : 355 – 359 .
- Kostyuk , A.S. and Temuryants , N.A. 2009 . Electromagnetic shielding changes infradian rhythmicity of pain sensitivity of land snails Helix albescens . Scientific Notes of Taurida V.Vernadsky National University – Series: Biology, Chemistry , 22 : 87 – 94 .
- Koukkari , W.L. and Sothern , R.B. 2006 . Introducing biological rhythms: a primer on the temporal organization of life, with implications for health, society, reproduction, and the natural environment , 1st Springer, New York.
- Limpasuvan , V. , Leovy , C.B. and Orsolini , Y.J. 2000 . Observed temperature two-day wave and its relatives near the stratopause . Journal of the Atmospheric Sciences , 57 : 1689 – 1701 .
- Liu , H.-L. , Talaat , E.R. , Roble , R.G. , Lieberman , R.S. , Riggin , D.M. and Yee , J.-H. 2004 . The 6.5-day wave and its seasonal variability in the middle and upper atmosphere . Journal of Geophysical Research , 109 : D21112 doi: 10.1029/2004JD004795.
- Lüning , K. , Kadel , P. and Pang , S.H. 2008 . Control of reproduction rhythmicity by environmental and endogenous signals in Ulva pseudocurvata (Chlorophyta) . Journal of Phycology , 44 : 866 – 873 .
- Manson , A.H. , Meek , C.E. , Chshyolkova , T. , Avery , S.K. , Thorsen , D. , MacDougall , J.W. , Hocking , W. , Murayama , Y. and Igarashi , K. 2005 . Wave activity (planetary, tidal) throughout the middle atmosphere (20–100km) over the CUJO network: satellite (TOMS) and medium frequency (MF) radar observations . Annales Geophysicae , 23 : 305 – 323 .
- Marques , N. , Araujo , J.F. , Righini , A.S.F. , Fleury , J.J. , Costa-Neto , J.B.F. and Caldas , M.C.S. 1996 . Circaseptan rhythms of semen characteristics of a brazilian breed (“Mangalarga”) stallion . Biological Rhythm Research , 27 : 343 – 350 .
- Mayr , H.G. , Mengel , J.G. , Talaat , E.R. , Porter , H.S. and Chan , K.L. 2004 . Modeling study of mesospheric planetary waves: genesis and characteristics . Annales Geophysicae , 22 : 1885 – 1902 .
- Muller , H.G. 1972 . Long period meteor wind oscillations . Philosophical Transactions of the Royal Society London A , 271 : 585 – 598 .
- Neumann , D. 1989 . Circadian components of semilunar and lunar timing mechanisms . Journal of Biological Rhythms , 4 : 173 – 182 .
- Neumann , D. and Heimbach , F. 1985 . Circadian range of entrainment in the semilunar eclosion rhythm of the marine insect Clunio marinus . Journal of Insect Physiology , 31 : 549 – 557 .
- Pancheva , D.V. , Mukhtarov , P.J. , Shepherd , M.G. , Mitchell , N.J. , Fritts , D.C. , Riggin , D.M. , Franke , S.J. , Batista , P.P. , Clemesha , B.R. and Kikuchi , T. 2006 . Two-day wave coupling of the low-latitude atmosphere-ionosphere system . Journal of Geophysical Research , 111 : A07313 doi: 10.1029/2005JA011562.
- Pancheva , D.V. , Mukhtarov , P.J. , Mitchell , N.J. , Fritts , D.C. , Riggin , D.M. , Takahashi , H. , Batista , P.P. , Clemesha , B.R. , Gurubaran , S. and Ramkumar , G. 2008 . Planetary wave coupling (5–6-day waves) in the low-latitude atmosphere ionosphere system . Journal of Atmospheric and Solar-Terrestrial Physics , 70 : 101 – 122 .
- Palo , S.E. , Roble , R.G. and Hagan , M.E. 1999 . Middle atmosphere effects of the quasi-two-day wave determined from a General Circulation Model . Earth Planets Space , 51 : 629 – 647 .
- Provasoli , L. 1968 . “ Media and prospects for the cultivation of marine algae ” . In Cultures and collections of algae , Edited by: Watanabe , A. and Hattori , A. 63 – 75 . Hakone : U.S.–Japan Conference, Japanese Society of Plant Physiology .
- Quartly , G.D. , Cipollini , P. , Cromwell , D. and Challenor , P.G. 2003 . Rossby waves: synergy in action . Philosophical Transactions of the Royal Society of London, series A , 361 : 57 – 63 .
- Reinberg , A. and Smolensky , M.H. 1983 . Biological rhythms and medicine: cellular, metabolic, physiopathologic, and pharmacologic aspects , New York. : Springer Verlag .
- Rensing , L. and Ruoff , P. 2002 . Temperature effect on entrainment, phase shifting, and amplitude of circadian clocks and its molecular bases . International Journal of Chronobiology , 19 : 807 – 864 .
- Riggin , D.M. , Lieberman , R.S. , Vincent , R.A. , Manson , A.H. , Meek , C.E. , Nakamura , T. , Tsuda , T. and Portnyagin , Y.I. 2004 . The 2-day wave during the boreal summer of 1994 . Journal of Geophysical Research , 109 : D08110 doi: 10.1029/2003JD004493.
- Rodgers , C.T. and Horea , P.J. 2009 . Chemical magnetoreception in birds: the radical pair mechanism . Proceedings of the National Academy of Sciences of the United States of America , 106 : 353 – 360 .
- Ruoff , P. and Rensing , L. 2004 . Temperature effects on circadian clocks . Journal of Thermal Biology , 29 : 445 – 456 .
- Salby , M.L. 1981 . The 2-day wave in the middle atmosphere: observations and theory . Journal of Geophysical Research , 86 : 9654 – 9660 .
- Snedecor , G.W. 1961 . Statistical methods applied to experiments in agriculture and biology , Moscow. [In Russian] : Science .
- Spruyt , E. , Verbelen , J.-P. and Greef , D. 1987 . Expression of circaseptan and circannual rhythmicity in the imbibition of dry stored bean seeds . Plant Physiology , 84 : 707 – 710 .
- Tal , O. , Haim , A. , Harel , O. and Gerchman , Y. 2011 . Melatonin as an antioxidant and its semi-lunar rhythm in green macroalga Ulva sp . Journal of Experimental Botany , 62 : 1903 – 1910 .
- Talaat , E.R. , Yee , J.-H. and Zhu , X. 2001 . Observations of the 6.5-day wave in the mesosphere and lower thermosphere . Journal of Geophysical Research , 106 ( 20 ) : 715 – 723 . 20
- Temuryants , N.A. , Chuyan , E.N. and Shekhotkin , A.V. 1995 . Infradian rhythmics of the functional state of neutrophils and lymphocytes of rats with different constitutional features . Biophysics , 40 : 1121 – 1125 .
- Temuryants , N.A. , Martynyuk , V.S. and Chuan , E.N. 2007 . “ Influence of electromagnetic fields of extremely different frequency diapason on infradian rhythms of physiological processes ” . In Biophotonics and coherent systems in biology Edited by: Beloussov , L.V. , Voeikov , V.L. and Martynyuk , V.S. 191 – 201 . Springer, New York
- Thayaparan , T. , Nocking , W.K. , MacDoudall , J. , Manson , A.H. and Meek , C.E. 1997 . Simultaneous observations of the 2-day wave at London (43°N, 81°W) and Saskatoon (52°N, 107°W) near 91 km altitude during the two years of 1993 and 1994 . Annales Geophysicae , 15 : 1324 – 1339 .
- Thébault , J. , Chauvaud , L. and Clavier , J. 2006 . Evidence of a 2-day periodicity of striae formation in the tropical scallop Comptopallium radula using calcein marking . Marine Biology , 149 : 257 – 267 .
- Titlyanov , E.A. , Titlyanova , T.V. , Kalita , T.L. and Yakovleva , I.M. 2004 . Rhythmicity in division and degradation of zooxanthellae in the hermatypic coral Stylophora pistillata . Symbiosis , 36 : 211 – 224 .
- Titlyanov , E.A. , Titlyanova , T.V. , Yakovleva , I.M. and Kalita , T.L. 2006 . Rhythmical changes in the division and degradation of symbiotic algae in hermatypic corals . Russian Journal of Marine Biology , 32 : 17 – 25 .
- Titlyanova , T.V. 1990 . “ Anatomical and morphological description and phenological observations of life cycles in Enteromorpha linza and Ulva fenestrata in Peter the Great Bay ” . In Ontogenetic aspects of photosynthesis in marine macroalgae , Edited by: Titlyanov , E.A. 7 – 22 . Vladivostok. [In Russian] : FEB RAS .
- Vignoles , P. , Alarion , N. , Bellrt , V. , Dreyfuss , G. and Rondelaud , D. 2006 . A 6- to 8-day periodicity in cercarial shedding occurred in some Galba truncatula experimentally infected with Fasciola hepatica . Parasitology Research , 98 : 385 – 388 .
- Wheeler , M.C. 2002 . “ Equatorial waves ” . In Tropical meteorology , Edited by: Shankar , M. Elsevier Science . doi: 10.1006/rwas.2002.0414
- Wiltschko , W. and Wiltschko , R. 2005 . Magnetic orientation and magnetoreception in birds and other animals . Journal of Comparative Physiology, A: Sensory, Neural, and Behavioral Physiology , 191 : 675 – 693 .
- Wu , D.L. , Fishbein , E.F. , Read , W.G. and Waters , J.W. 1996 . Excitation and evolution of the quasi-2-day wave observed in UARS/MLS temperature measurements . Journal of the Atmospheric Sciences , 53 : 728 – 738 .
- Wu , L.-Q. and Dickman , J.D. 2012 . Neural correlates of a magnetic sense . Science , 336 : 1054 – 1057 .
- Wu , X. , Xie , Q. , He , Z. and Wang , D. 2008 . Free and forced Rossby waves in the Western South China Sea inferred from Jason-1 satellite altimetry data . Sensors , 8 : 3633 – 3642 .
- Yamashita , T. , Murakawa , Y. , Hayami , N. , Inoue , M. , Fukui , E. , Kasaoka , Y. and Omata , M. 1999 . Infradian rhythm of paroxysmal atrial fibrillation. A case report . Japanese Heart Journal , 40 : 227 – 232 .
- Yarmolyuk , N.S. 2010 . Electromagnetic shielding changes infradian rhythmicity speeds of planarian Dugesia tigrina . Scientific Notes of Taurida V. Vernadsky National University – Series: Biology, Chemistry , 23 : 200 – 208 .
- Yoshida , S. , Tsuda , T. , Shimizu , A. and Nakamura , T. 1999 . Seasonal variations of 3.0∼3.8-day ultra-fast Kelvin waves observed with a meteor wind radar and radiosonde in Indonesia . Earth Planets Space , 51 : 675 – 684 .
- Yoshii , T. , Ahmad , M. and Helfrich-Förster , C. 2009 . Cryptochrome mediates light-dependent magnetosensitivity of Drosophila’s circadian clock . Public Library of Science: Biology , 7 : 0813 – 0819 .
- Zhadin , M.N. 2001 . Review of Russian literature on biological action of DC and low-frequency AC magnetic fields . Bioelectromagnetics , 2 : 27 – 45 .
- Ziegler-Page , J. and Kingsbury , J.M. 1968 . Culture studies on the marine green alga Halicystis parvula – Derbesia tenuissima. II. Synchrony and periodicity in gamete formation and release . American Journal of Botany , 55 : 1 – 11 .
- Ziegler-Page , J. and Sweeney , B.M. 1968 . Culture studies on the marine green alga Halicystis parvula–Derbesia tenuissima. III. Control of gamete formation by an endogenous rhythm . Journal of Phycology , 4 : 253 – 260 .
Supplementary information
The following supplementary material is available for this article, accessible via the Supplementary Content tab on the article's online page at http://dx.doi.org/10.1080/09670262.2013.796528
Supplementary . Factorial ANOVA results comparing length of growth cycle in Ulva lactuca exposed to four temperatures (5, 10, 15 or 20°C) by two light levels (40 or 60 μmol photons m−2 s−1) over period of 117 days.
Supplementary Table 2. Summary of main effects ANOVA results showing the significance effect of temperature and irradiance on length of growth cycle in Ulva lactuca.
Supplementary Table 3. Result of a one-way ANOVA shows a significant increase of the growth rate in Ulva lactuca at lengthening of daylight from 8 to 12 h.