Abstract
Structure and length of transitional zone in the dog's external carotid artery was documented. Six normal adult male dogs were studied. The specimens were processed in the routine way by light and electron microscopy. It was established that the transitional zone with 17-mm length contains parallel and fenestrated elastic fibres. In the transitional zone, the elastic fibres in the extracellular space were greater in number than collagen fibres. However, in the non-transitional zone, the rows of smooth muscle cells were greater in number. There were more collagen fibres than elastic fibres in the extracellular space and the elastic fibres appeared to be fragmented. Therefore, the transitional zone showed a structure intermediate between those of elastic and muscular arteries. The involvement of a special structure of transitional zone in vascular disease such as atherosclerosis is suggested.
Introduction
The two common carotid arteries are the principal arteries supplying the head and neck, each dividing into two branches: the external and the internal carotid artery. The external carotid artery supplies the exterior of the head, the face and the greater part of the neck (Evans Citation1993). Elastic arteries include the aorta, the common carotid artery and the subclavian arteries. It is well known that the external carotid artery is classified predominantly as a muscular type of artery (Janzen et al. Citation2001). Major components of the elastic arteries are elastic fibres that replace with smooth muscle cells in muscular arteries. Elastic fibres are composed of elastin and fibrillin microfibrils (Kielty et al. Citation2002). Elastic fibres are major components of extracellular matrix in dynamic tissues such as blood vessels (Davis Citation1993), skin (Braverman and Fonferko Citation1982) and the lungs (Pierce and Hocott Citation1960). Elastic fibre-rich dynamic tissues are therefore able to deform and store energy under normal physiological loads and to use this energy to drive recoil back to a resting state (Gosline et al. Citation2002). Elastic fibres allow arteries and lungs to expand and contract, thus controlling variations in blood pressure and returning the pulmonary system to a resting state (Robert et al. Citation2008). The structural transition between the elastic origin and the muscular pattern in the tunica media appears to be gradual, not abrupt (Young et al. Citation2007). Histologically, the carotid artery tripod is a region of a structural transition between the elastic and muscular pattern in the tunica media (Janzen et al. Citation2001). The microscopic transitional zone is the segment of the arterial tree in which the elastic wall is replaced by a muscular type (Janzen Citation2004).
The structure of arteries has interested researchers for a long time. Van Baardwijk et al. (Citation1985) showed that the bundles of elastin appear to be continuous from the aorta into its branches. Light microscopic studies (Rees Citation1968) have shown marked differences in the pattern of elastin at aorta–branch junctions on the proximal and distal lips of the junction and have shown that the medial layer of the branches is transitional. The proximal part of the renal artery in rats shows a gradual transition from an elastic to a muscular type as well (Osborn-Pellegrin Citation1978). In fact, a preponderance of atherosclerosis at aortic branching points and bifurcations has been well documented in humans (Schulz and Rothwell Citation2001). In the hypercholesterolemic rabbit, atherosclerotic lesions occur at the distal and lateral sides of the orifice of dorsal inter-costal arteries (Emura et al. Citation1992). Therefore, the present study can increase our knowledge to manage and reduce the risk of disease in this artery.
There is little information regarding the transitional zone structure. The transitional zone may play an important role in the development of arterial disease such as atherosclerosis. In this study, the transitional zone in the dog's external carotid artery was investigated. The dog was chosen for study because there have been no reports concerning the special structure of this area in the normal dog.
Materials and methods
Twelve external carotid arteries were harvested from six clinically healthy adult male dogs (three for histology and three for ultrastructure). The dogs were euthanised with an overdose of anaesthetic (thiopental sodium) and the external carotid arteries exposed.
For histology, the carotid artery tripods were dissected from the neck and the external carotid arteries were freed and transected from their origin from common carotid artery to the final branch of external carotid artery (superficial temporal artery). After removal, the arteries were fixed in 10% formalin. After 24 hours of fixation, each artery was divided into three samples of proximal, middle and distal parts. Each sample was processed by routine paraffin processing. The paraffin-blocked tissue was cut into 6 µm serial sections. Each tenth section was mounted and stained with Orcein for elastic fibres. The microscopic structure was viewed to determine and record the presence of transitional zone structures. The length of the transitional zone was determined by totalling the number of transitional sections and multiplied by the section thickness.
For comparison with the external carotid artery, the common carotid arteries were taken from three dogs for observation under a light microscope. The specimens were processed in the routine way by light microscopy.
After determination of the external carotid artery transitional zone by light microscope, the external carotid arteries were removed from three more dogs following the same harvesting procedure. After removal of the external carotid artery, the samples for ultrastructural study were taken from the proximal part of the external carotid artery as transitional zone and distal part as muscular artery immediately after euthanasia. The samples were first rinsed with normal saline and fixed in 2.5% glutaraldehyde for 8 hours. The specimens were then rinsed with a cold buffer washing twice (every time for 15 minutes) and post fixed in 1% osmium tetroxide for 2 hours. After that, the specimens were dehydrated in a graded series of alcohol and cleared with the addition of propilenoxide. The specimens were then infiltrated in a mixture of propilenoxide and Epon resin. Gradually, the epon–propilenoxide ratio was increased until pure Epon resin was used. The samples were then embedded in Epon 812 resin. Transverse sections were made and stained with uranyl acetate and the structure of the transitional zone was studied.
Results
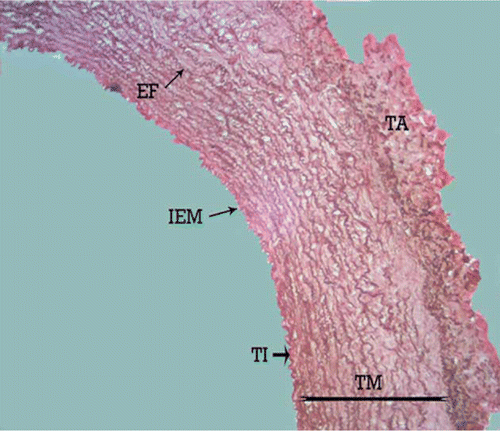
The structure of the external carotid artery of the adult male dog is similar to that of other mammalian arteries with tunica intima, tunica media and tunica adventitia. Light microscopy confirmed the structural change of the tunica media in the length of the canine external carotid artery. The proximal part of the external carotid artery showed the structure between the elastic and muscular artery types. After 17 mm, the external carotid artery showed the typical morphologic features of a muscular artery. The length of the transitional zone in both the left and the right external carotid artery was 17±0.04 mm. The histological study demonstrated that, the media of the common carotid artery is composed of several concentric elastic lamellae separated by layers of smooth muscle cells. It can be observed that the elastic lamellae are essentially continuous sheets. The clear external elastic membrane was not observed ().
Figure 1. Photomicrograph – the tunica media of canine common carotid artery with several continuous and concentric elastic lamellae. TI, tunica intima; TM, tunica media; TA, tunica adventitia; IEM, internal elastic membrane; EF, elastic fibres. Orcein stain, 280×, Bar = 35.7 µm.
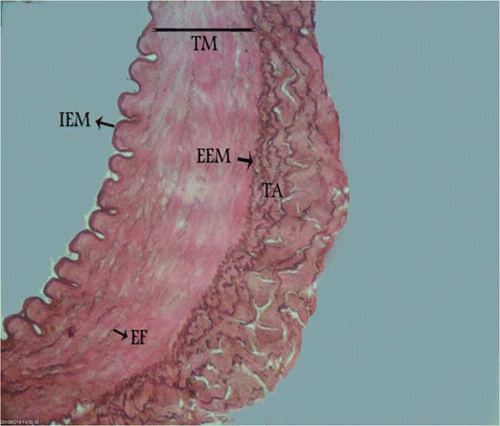
The structure between elastic and muscular artery types was found at the origin of the external carotid artery with parallel and fenestrated elastic fibres between smooth muscle cells in the tunica media. In this section, the internal elastic membrane was present, but the external elastic membrane did not exist between the tunica media and the tunica adventitia (). In the middle part of the external carotid artery, more rows of smooth muscle cells and fewer layers of elastic fibres were seen and the external elastic membrane was not observed (). In the distal part, the typical morphologic features of muscular arteries were found. In this area, smooth muscle cells predominated in the media and rare remnants of elastic fibres between the smooth muscle cells were present. In the distal section an internal elastic membrane separated the tunica intima from the tunica media and there was an external elastic membrane at the junction of the tunica media and tunica adventitia ().
Figure 2. Photomicrograph – canine external carotid artery transitional zone with parallel and fenestrated elastic fibres in the media. TI, tunica intima; TM, tunica media; TA, tunica adventitia; IEM, internal elastic membrane; EF, elastic fibres. Orcein stain, 280×, Bar = 35.7 µm.
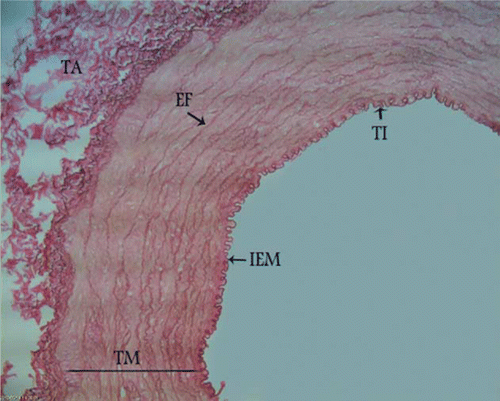
Figure 3. Photomicrograph – transitional zone in the middle part of canine external carotid artery with more rows of smooth muscle cells and fewer layers of elastic fibres. TI, tunica intima; TM, tunica media; TA, tunica adventitia; IEM, internal elastic membrane; EF, elastic fibres. Orcein stain, 280×, Bar = 35.7 µm.
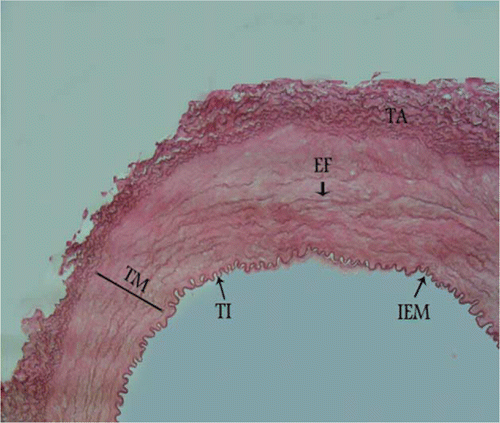
Figure 4. Photomicrograph – non-transitional zone of canine external carotid artery with fragmented elastic fibres in the media. TM, tunica media; TA, tunica adventitia; IEM, internal elastic membrane; EEM, external elastic membrane; EF, elastic fibres. Orcein stain, 700×, Bar = 25.5 µm.
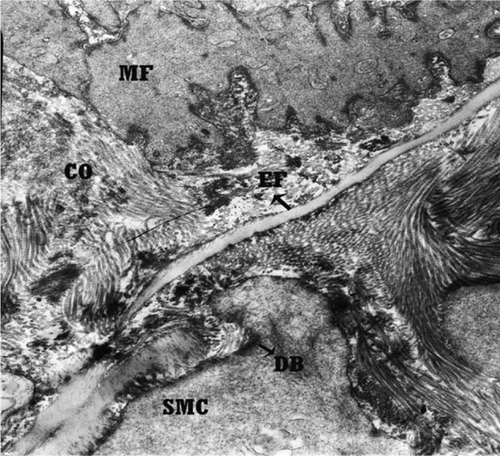
The ultrastructural studies confirmed these differences in the tunica media. AU: the tunica media at the origin of the external carotid artery from the common carotid artery showed more elastic fibres in comparison with the collagen fibres in extracellular space. In this section, the elastic fibres were in a continuous arrangement (), while in the distal part of the external carotid artery section, elastic fibres became rare and fragmented. In this area, collagen fibres were more numerous than elastic fibres in the extracellular spaces between smooth muscle cells (). Myofilaments were found in the cytoplasm of most of the smooth muscle cells of the arterial wall in the origin and in the distal part of the external carotid artery section. Smooth muscle cells contained conspicuous dens bodies which underlied the cell surface and were found between myofilaments ( and ).
Figure 5. Transmission electron micrograph of transitional zone of canine external carotid artery with more elastic fibres in comparison with the collagen fibres in extracellular space. EF, elastic fibres; COF, collagen fibres; SMC, smooth muscle cell; MF, myofilament; DB, dens bodies. Orcein stain, 8900×, Bar = 1.1 µm.
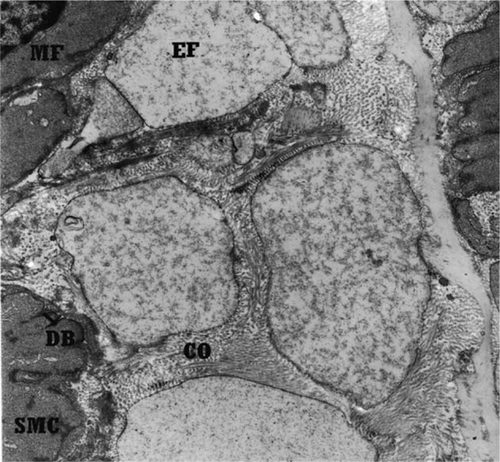
Figure 6. Transmission electron micrograph of non-transitional zone of right canine external carotid artery with rare and fragmented elastic fibres and more collagen fibres in extracellular space. EF, elastic fibres; COF, collagen fibres; SMC, smooth muscle cell; MF, myofilament; DB, dens bodies. Orcein stain, 11500×, Bar = 0.86 µm.
Discussion
Several studies have been performed on arteries (Ball et al. Citation1963; Aleckseevskikh Citation1968; Pinto et al. Citation1998), but little information is available about the transitional zone in arteries (Roach Citation1987; Gabella Citation1995). In these histological studies, we were able to show a gradual transition of about 17 mm in the length of the structure of the tunica media in the dog external carotid artery. In the White Carneau pigeon, a transitional zone has been reported in the coeliac artery which was about 1 cm (Lauper et al. Citation1975). In humans, the typical transitional zone was reported in the renal artery with maximally 10-mm long and in the internal and external carotid artery with between 5- and 15-mm long (Janzen et al. Citation2000, Citation2001). The tunica media in the origin of the external carotid artery showed a structure intermediate between the common carotid artery and the distal external carotid artery and was therefore named the transitional zone, and in this zone the rows of elastic fibres were more numerous and the external elastic membrane was absent. This may be a result of the similarity of the external elastic membrane to the elastic fibres of the tunica media in arteries such as the aorta.
It has been reported that elastin is an important elastic component that provides extensible tissues with resilience (Faury Citation2001). In the walls of the artery, specially in large arteries elastic rebound helps to maintain arterial pressure (Ushiki Citation2002).
In the non-transitional zone tunica media, the distal part of external carotid artery, the rows of smooth muscle cells increased in number and rare remnants of elastic fibres were found between the smooth muscle cells. In this zone, collagen fibres in extracellular space increased in number and the external elastic membrane and a typical muscular morphology were observed clearly. In two zones, there were dens body in smooth muscle cells. This observation is similar to the findings in other studies. Osborn-Pellegrin (Citation1978) reported that the elastic lamellae in the media of the renal artery becomes thinner as the size of the vessel decreases and they disappear completely in the distal renal artery. Different structures occur at other vessel sites. For instance, at the origin of the renal artery and the common carotid artery in humans a segment has been observed in which the elastic type wall architecture is replaced by one of the muscular type (Janzen Citation2004). Studies of the vertebral artery in the giraffe (Kimani Citation1986) have shown that the caudal segment of the vertebral artery has a largely elastic structure, while the cranial segment has a muscular structure. The transition of the arterial wall normally occurs between the seventh and the fifth cervical vertebral levels. It involves diminution of the elastic tissue in the luminal portion of the tunica media and a simultaneous increase of the smooth muscle cells content. Examination of the musculophrenic and superior epigastric arteries (Van Son et al. Citation1993) has shown that the media of the first 1–2 cm of these arteries is elastomuscular or muscular with rare elastic lamellae, whereas more distally the media is purely muscular. We know that during systole the blood enters the large elastic arteries with considerable force and that these arteries distend. They are able to do so because of the large amount of elastic tissue in their walls. During diastole, the arteries return to their original size because of the elastic recoil of the walls. The flow of blood to the organ is controlled by the contraction or relaxation of the smooth muscle cells of the tunica media (Junqueira et al. Citation2005). Research has shown (Roach Citation1983) that the amount of elastin along the thoracic aorta is decreased compared with the abdominal aorta and that this is linearly related to the pulse pressure. Therefore, the existence of more elastic fibres in the transitional zone compared with the non-transitional zone and the replacement of elastic fibres by smooth muscle cells in the non-transitional zone can be viewed as physiological. The larger number of elastic fibres in the transitional zone is due to the proximity of this part of the external carotid artery to the common carotid artery, and it helps to maintain the blood pressure. However, in the distal external carotid artery, which contains smaller amounts of elastic fibres, smooth muscle cells help to maintain the blood pressure and regulate the amount of blood flowing.
Osborn-Pellegrin (Citation1978) in a study of rat arteries reported that dens bodies are more numerous in the renal artery than in the aorta. Dens bodies have been observed previously in the smooth muscle cells of the cat and the monkey in varying detail (Pease and Molinari Citation1960). Dens bodies in the cytoplasm and the cell membrane serve as anchor sites for the myofilaments that are well developed in muscular arteries (Ross Citation1971).
The clinical implications of the transitional zone in the external carotid artery are not yet fully understood but the existence of the transitional zone and its length can be important in vascular diseases. Degenerative diseases of the arterial wall, such as atherosclerosis, often occur at arterial bifurcations and branches (Schulz and Rothwell Citation2001). These malfunctions are probably associated with change in the elasticity of the arterial tissue involved and, perhaps, with factors such as local flow disturbance at bifurcation (Roach Citation1977). Altered fluid dynamics at branching points has been recognised as one of the crucial promoting factors for the development of atherosclerosis (Karino et al. Citation1987). The histopathological findings in rat with an atherogenic diet consisted of marked rarefaction, fat deposition, atheroma in the subendothelial tissue and tunica intima and decrease in elastic fibres. The remaining elastic fibres had such abnormalities as a tortuous course, bends and fissures (Katsunuma et al. Citation1983). Therefore, changing histologic architecture within the transitional zone, and its length, may play an important role in the development of vascular disease such as atherosclerosis.
Conclusion
Transitional zone – an arterial segment with transition from elastic to muscular type – was found at the origin of the external carotid artery of the dog. The length of the transitional zone was 17 mm. Some differences between the transitional and the non-transitional zone of the external carotid artery were found and the involvement of a special structure in vascular disease such as atherosclerosis is suggested.
Acknowledgements
The authors would like to acknowledge the research council of the Islamic Azad University, Kazeroun Branch, Kazeroun, Iran, for their financial support.
References
- Aleckseevskikh , U . 1968 . Morphological changes of renal vessels in condition of distribution . Archives of Pathology and Laboratory Medicine , 300 : 56 – 62 .
- Ball , RA , Sautter , JH and Katter , MS . 1963 . Morphological characteristics of the anterior mesenteric artery of fowl . Anatomical Record , 146 : 251 – 255 .
- Braverman , IM and Fonferko , E . 1982 . Studies in cutaneous aging: I. The elastic fiber network . Journal of Investigative Dermatology , 78 : 434 – 443 .
- Davis , EC . 1993 . Stability of elastin in the developing mouse aorta: a quantitative radioautographic study . Histochemistry , 100 : 17 – 26 .
- Emura , S , Masucko , S and Sunaga , T . 1992 . Fine structures around the orifice of the intercostal artery of the rabbit thoracic aorta . Angiology , 43 : 211 – 218 .
- Evans , H . 1993 . Miller's anatomy of the dog , 3rd ed , Philadelphia, PA : Saunders .
- Faury , G . 2001 . Role of elastin in the development of the vascular function. Knock-out study of the elastin gene in mice . Journal De La Societe De Biologie , 195 : 151 – 156 .
- Gabella , G . 1995 . Complex structure of the common carotid artery of sheep . Anatomical Record , 243 : 376 – 383 .
- Gosline , J , Lillie , M , Carrington , E , Guerette , P , Ortlepp , C and Savage , K . 2002 . Elastic proteins: biological roles and mechanical properties . Philosophical Transactions of the Royal Society , 357 : 121 – 132 .
- Janzen , J . 2004 . The microscopic transitional zone between elastic and muscular arteries . Archives des maladies du coeur et des vaisseaux , 97 : 909 – 914 .
- Janzen , J , Lanzer , P , Rothenberger-Janzen , K and Vuong , PN . 2000 . The transitional zone in the tunica media of renal arteries has a maximal length of 10 millimeters . Archives des maladies du coeur et des vaisseaux , 29 : 168 – 172 .
- Janzen , J , Lanzer , P , Rothenberger-Janzen , K and Vuong , PN . 2001 . Variable extension of the transitional zone in the medial structure of carotid artery tripod . Archives des maladies du coeur et des vaisseaux , 30 : 101 – 106 .
- Junqueira , LC , Carneiro , J and Kelly , RO . 2005 . Basic histology (text and atlas) , 11th ed , New York, NY : McGraw-Hill Medical Pub .
- Karino , T , Goldsmith , H , Motomiya , M , Mabuchi , S and Sohara , Y . 1987 . Flow patterns in vessels of simple and complex geometries . Annals of the New York Academy of Sciences , 516 : 422 – 441 .
- Katsunuma , H , Shimizu , K , Ebihara , T , Iwamoto , T and Kiyokawa , M . 1983 . Anti-atherosclerotic action of elastase-with special reference to its effect on elastic on elastic fibers . Age and Aging , 12 : 183 – 194 .
- Kielty , CM , Sherratt , MJ and Shuttleworth , CA . 2002 . Elastic fibres . Journal of Cell Science , 115 : 2817 – 2828 .
- Kimani , JK . 1986 . Structural organization of the vertebral artery in the giraffe (Giraffa camelopardalis) . Anatomical Record , 217 : 256 – 262 .
- Lauper , NT , Unni , KK , Kottke , BA and Titus , JL . 1975 . Anatomy and histology of aorta of White Carneau pigeon . Laboratory Investigation , 32 : 536 – 551 .
- Osborn-Pellegrin , M . 1978 . Some ultrastructural of the renal artery and abdominal aorta in the rat . Journal of Anatomy , 25 ( pt 3 ) : 641 – 651 .
- Pease , DC and Molinari , S . 1960 . Electron microscopy of muscular arteries; pial vessels of the cat and monkey . Journal of Ultrastructure Research , 3 : 447 – 468 .
- Pierce , JA and Hocott , JB . 1960 . Studies on the collagen and elastin content of the human lung . The Journal of Clinical Investigation , 39 : 8 – 14 .
- Pinto , YM , Pinto , SJ , Paul , M and Merker , HJ . 1998 . The electron microscopic morphology of the common carotid artery in rat . Annals of Anatomy , 180 : 223 – 235 .
- Rees , PM . 1968 . Electron microscopical observations on the architecture of the carotid arterial walls. With special reference to the sinus portion . Journal of Anatomy , 103 ( pt 1 ) : 33 – 47 .
- Roach , MR . 1977 . Biophysical analyses of blood vessel walls and blood flow . Annual Review of Physiology , 39 : 51 – 71 .
- Roach , MR . 1983 . The pattern of elastin in the aorta and large arteries of mammals . Ciba Foundation Symposium , 100 : 37 – 55 .
- Roach , MR . 1987 . The structure and elastic properties of arterial junction . Connective Tissue Research , 15 : 77 – 84 .
- Robert , L , Robert , AM and Fulop , T . 2008 . Rapid increase in human life expectancy: will it soon be limited by the aging of elastin? . Biogerontology , 9 : 119 – 133 .
- Ross , R . 1971 . The smooth muscle cell . The Journal of Cell Biology , 50 : 172 – 186 .
- Schulz , UG and Rothwell , PM . 2001 . Sex differences in carotid bifurcation anatomy and the distribution of an atherosclerotic plaque . Stroke , 32 : 1525 – 1531 .
- Ushiki , T . 2002 . Collagen fibers, reticular fibers and elastic fibers. A comprehensive understanding from a morphological viewpoint . Archives of Histology and Cytology , 65 : 109 – 126 .
- Van Baardwijk , C , Barwick , SE and Roach , MR . 1985 . Organization of medial elastin at aortic junction in sheep and lambs . Canadian Journal of Physiology and Pharmacology , 63 : 855 – 862 .
- Van Son , JA , Smedts , FT , Wilde , PC , Pijls , NH , Wong-Alcala , L , Kubat , K , Tavilla , G and Lacquet , LK . 1993 . Histological study of the internal mammary artery with emphasis on its suitability as a coronary artery bypass graft . The Annals of Thoracic Surgery , 55 : 106 – 113 .
- Young , B , Low , JS , Stevens , A and Heath , JW . 2007 . Functional histology a text and color atlas , 5th ed , Philadelphia, PA : Churchill Livingstone .