
ABSTRACT
We compared the dry matter intake (DMI), body condition score (BCS), body weight (BW), beta-hydroxy-butyrate (BHB), milk yield, and milk composition during the transition period of Holstein and crossbred Holstein x Simmental cows. Thirty cows were used in the study. Each cow entered the study 21 days before their expected calving day (prepartum) and remained in the study until day 21 after calving (postpartum). DMI, and milk yield were recorded daily. Once a week, BW and BCS were recorded and blood was collected to measure the BHB. The SAS software was used for the statistical analysis. Holstein and crossbred Holstein x Simmental cows had similar DMI during the prepartum and postpartum periods with higher intakes after calving. Crossbred Holstein x Simmental cows had a higher BCS than Holstein cows during the prepartum and postpartum periods. BHB does not differ between genetic groups, but between periods – with lower values in the prepartum period. The genetic groups had similar milk yield and composition. Both genetic groups lost BCS after calving, but Holstein cows showed a greater decrease in BCS. The use of a crossbreeding programme with Simmental has no negative effect on the performance during the transition period in Holstein herds.
Introduction
Crossbreeding between dairy breeds is an alternative to pure breeding that has been used to improve phenotype characteristics (related mainly to fitness) or to reduce the problems associated with the relatively high inbreeding rates in purebred herds (Weigel and Barlass Citation2003; Cassell and McAllister Citation2009). In addition to the reducing inbreeding, the genetic variability of the herds should improve and combine utilizing heterosis with complementarity effects (Mendonça et al. Citation2014). Positive heterosis effects can improve feed efficiency by reducing the energy required for maintenance, and deposition of body tissues due to the improved partitioning of consumed energy (Prendiville et al. Citation2009; Olson et al. Citation2010). Heins et al. (Citation2008), for example, showed that crossbred Holstein x Jersey cows had higher dry matter intakes proportional to body weight (BW), better feed efficiency, allocated a bigger portion of energy to body reserves which enabled cows a quick recuperation of their body condition score (BCS) after calving. A favourable BCS of the crossbred Holstein x Jersey cows might have a positive impact on the reproductive performance for example, in comparison to Holstein cows (Heins et al. Citation2008).
The transition period of dairy cows is receiving more attention resulting in more studies investigating the physiological and metabolic changes at the end of the dry period and the beginning of a new lactation. One of the changes is the reduction in the dry matter intake (DMI) at the end of the dry period, which may amount to almost 30% 24 h before calving (Dewhurst et al. Citation2000). The DMI reduction can reach 3.8 kg of dry matter, leading to a reduction of the rumination time – of up to 60 min/day (Schirmann et al. Citation2013). After calving, a gradual increase of DMI is expected (Youssef and El-Ashker Citation2017), although the DMI does not increase as fast as the rise in demand for nutrients for milk production. The animals go through a deficient energy period, well recognized as a negative energy balance (NEB) (Mann et al. Citation2016; Barletta et al. Citation2017; Djoković et al. Citation2017). An NEB may lead to an increase in the concentration of non-esterified fatty acids (NEFA), which indicates body fat mobilization (Mendonça et al. Citation2014).
NEFA can be used as a direct energy source for the tissues or may be completely oxidized in the liver resulting in ketone bodies, especially beta-hydroxy-butyrate (BHB), which can be used as an energy source by the liver or other tissues (Barletta et al. Citation2017; Djoković et al. Citation2017). Situations when the NEFA and BHB levels are higher than the healthy limits for a cow (plasma or serum concentration >0.4 mmol/l or >1.2 mmol/l, respectively), have been associated with lower milk yields, a reduced reproductive performance, higher risks of clinical and metabolic diseases, and finally an increased culling rate of the animals (Leblanc Citation2010; McCarthy et al. Citation2015).
The effects of NEB around the transition period have been frequently studied in high yielding herds, especially Holstein herds. Limited studies exist comparing Holstein and crossbred cows in this specific period. Crossbred cows may differ from Holsteins in DMI, the energy use efficiency, and energy metabolism due to heterosis and complementarity effects (Mendonça et al. Citation2014). Our study tests the hypothesis that crossbred Holstein x Simmental cows reduces DMI during the weeks before calving to a lower degree while showing a higher DMI immediately after calving with a similar milk yield to purebred Holsteins. Therefore, we aimed to compare DMI, BCS, BW, BHB, milk yield, and milk composition during the transition period (3 weeks before calving until 3 weeks after calving) for both breeding groups.
Material and methods
All procedures used in this research were approved by the Santa Catarina State University Ethical Committee, protocol n° 6330030517.
Animals and management
A commercial dairy farm located in Santa Catarina, South Brazil served as the experimental location for this study. The farm uses a compost bedded pack barn confinement system (CBP). The herd consists of approximately 280 lactating cows with 60% purebred Holstein cows and 40% crossbred Holstein x Simmental cows. A total number of 30 multiparous cows (18 Holstein and 12 crossbred F1 Holstein x Simmental cows) were used in the study. The crossbred cows originated from random crossings between purebred Holstein cows inseminated with semen from five Simmental bulls (DE 000935703404, DE 000811017046, DE 000812666322, DE 000812187178, DE 000939181646) which were imported from Germany and selected for their dairy breeding values (https://www.lfl.bayern.de/itz/rind/bazi/index.php?suche=#). The study was performed from August to November 2017. All cows with three or more parities that calved within the experimental time were included in the study. The cows underwent a dry off period of 60 days before the expected calving day. Each cow entered the study 21 days before the expected calving day (prepartum) and stayed in the research group until day 21 after calving (postpartum). Therefore, the study covered a six-week transition period. Cows entered and left the pre- and postpartum groups according to their expected and actual calving dates. The prepartum group stayed with all other prepartum cows in the farm in an area outside the barn with free access to water and shade. Twice a day, for approximately 2 h (totally 4 h per day), the prepartum cows had access to the feed parlour, where they received a total mixed ration based on maize silage and a commercial concentrate specifically prepared for this pre-lactation transition period. After calving, the (lactating) ‘research’ cows were kept as a separate group in the CBP. The diet offered to postpartum cows in the experimental group was the same as the diet offered to the other postpartum and high yielding cows in the farm. The diet was a total mixed ration (TMR) based on maize silage, ryegrass (fresh and silage), and concentrates. The ingredients and the chemical composition of the pre and postpartum diet are shown in . The diet offered to the cows was calculated to provide 100% of their nutritional requirements (NRC Citation2001). Both groups (prepartum and postpartum) had restricted access to food, food was only provided in the feed parlour. This routine corresponds to the dairy farm regime. During the rest of the day, the cows rested in the CPB (postpartum) or in the dry-cow paddock (prepartum).
Table 1. Ingredients and composition of TMRa (% of dry matter) offered to the dry (prepartum) and lactating (postpartum) Holstein and crossbred Holstein x Simmental dairy cows.
Milking took place three times a day (05:00 h, 13:30 h, and 20:30 h) and was combined with an electronic recording (DeLaval®) of the individual milk yield (MY). Individual milk samples were taken every 7 days in 40-mL bottles containing Bronopol. Each sample consisted of a pooled sample of the three milkings for the day. Samples were sent to the laboratory for milk analysis. The samples were analyzed for milk composition by an infrared method using the DairySpec equipment (Dairy Spec FT, Bentley®, USA).
After each milking, the postpartum cows had access to the feed parlour for approximately 2 h and 30 min – totalling approximately 7.5 h per day. The feed parlour had a self-locking feed front. The cows were already familiar with the system as part of the usual farm routine. The TMR offered was weighed and provided individually for measuring individual feed intake. The TMR was offered semi-ad libitum for each cow allowing a 5–10% residual and was prepared using a horizontal forage mixer. After the cows left the feed parlour, the non-consumed feed was weighed. Once a week, samples of the TMR offered and the residual from each cow (as well as the individual ingredients of the diet), were collected, and then dried in a forced-air oven at 55 °C for 72 h. After this procedure, the samples were milled through a 1-mm screen for chemical analyses. The dry matter content was determined by drying the samples at 105 °C for 24 h. The ash was quantified by combustion in a muffle furnace at 550 °C for 4 h, and the organic material was quantified by the mass difference. The total nitrogen was assayed using the Kjeldahl method (method 984.13; AOAC International, Citation1998). The neutral detergent fibre (NDF) concentration was assessed according to Mertens (Citation2002), except that the samples were weighed in filter bags and treated with a neutral detergent using Ankom A220 equipment (Ankom Technology, Macedon, NY). The concentration of the acid detergent fibre (ADF) was analyzed according to AOAC International (Citation1998). The procedures described for DMI measurements, TMR sampling, and chemical TMR analyses were the same regardless of stage in the transition period.
Once a week, BW and BCS were recorded. We performed the BCS evaluation using a scale between 1 (extremely thin) and 5 (very fat), with 0.25 intervals (Ferguson et al. Citation1994). On the same day, all cows were weighed (after milking for the lactating cows), just before feeding. On the same day, blood was sampled (in the morning, prior to the first feeding) from the coccygeal vein of each cow for the measurement of β-hydroxybutyrate (BHB). The concentration of BHB was immediately tested using an electronic handheld device (Precision Xtra metre, Abbott Diabetes Care). We performed all measurements throughout the 6-week observation period.
To obtain the daily rumination data, we used the data collected by the Heatime® (SCR/Allflex) system, an automatic system composed of a neck collar with a tag that records the rumination time (in minutes) of each cow.
Statistical analysis
The energy corrected milk yield (ECM) was obtained by the equation: ECM = (0.327 * MY) + (12.95% * F* MY / 100) + (7.65% P * MY / 100), where, MY = milk yield in l/day, F = fat percentage and P = protein percentage (Tyrrell; Reid, Citation1965). For the variance analysis, we used the MIXED procedure of the SAS (SAS Citation2002) statistical package after testing the data for normality of the residuals by the Kolmogorov–Smirnov test and the homogeneity of the variances using the Levene test. The first variance analysis model (model 1) included the fixed effects genetic group, period (pre/postpartum), and the interaction between them. With model 1 we aimed to show the differences pre and postpartum and the possible interaction of the period with the genetic groups. The variables analyzed were DMI, DMI to BW (%), BCS, BW, BHB, and rumination time. In a second model (model 2), for the analyses of milk yield and quality (ECM, Fat (%), Protein (%), Lactose (%), Fat + Protein yield (kg)), and to further analyze DMI, BHB, BCS, and BW, we included the week relative to calving as a fixed effect instead of the pre/postpartum period. With the second model, we aimed to demonstrate the weekly variation of the variables. By investigating possible interactions of the genetic group and week, we can demonstrate if the weekly variation of the variables is similar between the genetic groups. In all analyses a probability value less than 0.05 was considered statistically significant (P < 0.05).
Results
Holstein and crossbred Holstein x Simmental cows had similar DMI (P = 0.46; ). There was a difference between pre and postpartum DMI (P < 0.01) with a higher value after calving. Both genetic groups reduced the DMI in the week prior to calving (A). After calving, the DMI increased quickly (model 2). We observed an interaction between genetic group and period (P = 0.0074, model 1). The Holstein x Simmental cows had a higher DMI than purebred Holsteins in the third week after calving (A, model 2). Conversely, the DMI proportion to BW (in %) did not differ between Holstein and Holstein x Simmental crossbred cows (P = 0.96; ). There was no interaction between the genetic group and period (P = 0.14; model 1). There was also no difference in rumination time between the genetic groups (P = 0.41).
Figure 1. Weekly means of dry matter intake (A) and beta-hydroxybutyrate (BHB) (B) at three weeks before calving to three weeks after calving for purebred Holstein (H) and crossbred Holstein x Simmental cows (H x S) – results (LSM ± SEM) of variance analysis model 2.
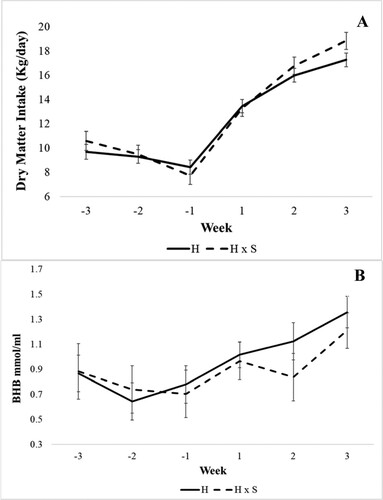
Table 2. Least Squares Means (±SEM) and results of the variance analysis (P-values) comparing genetic group (GG), period (prepartum and postpartum), and their interaction for the variables of dry matter intake (DMI), body weight (BW), body condition score (BCS), rumination time (RT), and beta-hydroxybutyrate (BHB) for purebred Holstein (H) and F1 Holstein x Simmental (H x S) crossbred cows (model 1).
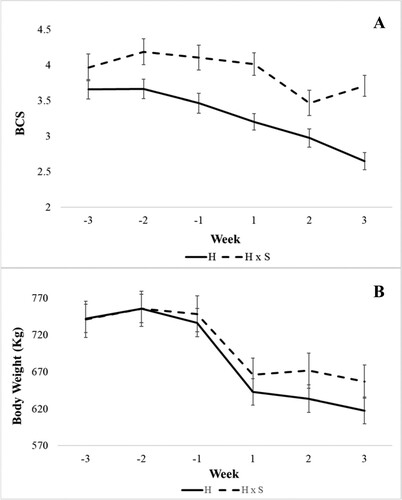
Crossbred Holstein x Simmental cows had a higher BCS than Holstein cows (P < 0.01; ). This difference was observed in both periods (pre and postpartum) (model 1). Furthermore, we found an interaction between genetic group and week (P = 0.0016, model 2). The BCS decreased in both genetic groups after parturition, but the BCS decrease stopped earlier for crossbred Holstein x Simmental cows (A, model 2). Both genetic groups showed a similar BW (P = 0.51; , model 1). Both genetic groups started to lose weight at week 1 prior to calving (B, model 2). There was an interaction between the genetic group and the transition period (P = 0.04). The BW difference between the genetic groups was approximately 30 kg per cow after calving (model 1).
Figure 2. Weekly means of body condition score (BCS) (A) and body weight (B) from three weeks before calving to three weeks after calving for purebred Holstein (H) and crossbred Holstein x Simmental (H x S) cows – results (LSM ± SEM) of variance analysis model 2.
For the variable BHB, we observed a significant difference for the period (P < 0.01) with lower values prepartum for both genetic groups (model 1). After calving, the BHB concentration increased to more than 1 mmol/liter (B, model 2). There was no interaction between the genetic group and transition period (P = 0.23, , model 1), and between the genetic group and week (P = 0.88, , model 2).
Table 3. Genetic Group least squares means (±SEM) and results of the variance analysis (P-values) for Genetic Group (GG), week relative to calving, and their interaction for the variables related to milk yield and milk composition, dry matter intake (DMI), body weight (BW), body condition score (BCS), rumination time (RT), and beta-hydroxybutyrate (BHB) for purebred Holstein and F1 Holstein x Simmental crossbred cows (model 2).
Holstein cows and crossbred Holstein x Simmental cows yielded similar amounts of milk (P = 0.60; ). Both genetic groups produced approximately 21 liters/day from the day after calving. During the following days, the milk production increased quickly until reaching amounts around 34 liters/day in the third week after calving. Genetic groups did not differ in the protein and lactose content within the milk (P = 0.64 and P = 0.83, respectively). Conversely, crossbred Holstein x Simmental cows tended to produce milk with higher percentages of fat content (P = 0.06).
Discussion
We did not find any difference in the BHB concentration, ECM yield, DMI, and BW between Holstein and crossbred Holstein x Simmental cows during the experimental period. This result suggests the energy status of both genetic groups is similar. Hazel et al. (Citation2013) compared Holstein and crossbred Holstein x Montbeliarde cows and found no difference between the genetic groups for DMI during the first 150 days of lactation. In the same study, crossbred cows produced 96% of the total milk compared to Holstein cows, similar to the yield of the cows in our study (94.5%). The observation of equal DMI in our study may relate to the production of similar amounts of milk combined with an identical milk composition (when measured as ECM). Both genetic groups in our study had similar BW. Therefore, it can be expected that cows of both genetic groups had similar maintenance requirements for energy and nutrients. Depending on the body composition and BW of the genetic groups, the nutrient requirements may differ, which might affect the DMI. By comparing Holstein with Jersey cows or Holstein with crossbred Holstein x Jersey cows, Palladino et al. (Citation2010) and Prendiville et al. (Citation2011) demonstrated that the heavier purebred Holstein cows consume more dry matter than the lighter purebred Jerseys or crossbred cows, respectively.
DMI increases as milk yield increases after calving (Weber et al. Citation2013). The cows almost doubled their DMI by 21 days after calving, starting with approximately 9 kg/day after calving and reaching almost 18 kg/day by day 21 postpartum (, A). The time pattern was similar for DMI in relation to BW (%) in both genetic groups. At the week before calving, cows had a DMI to the BW of approximately 1.2%. This value had more than doubled and reached values close to 3% of the BW by the third week of lactation. The increasing relative DMI is not only related to the increased demand for nutrients after calving. It increased more than the absolute DMI because the cows lost weight after calving (, B) as a consequence of the negative energy balance (NEB) (Carvalho et al. Citation2014; Esposito et al. Citation2014). It is interesting to note that at 21 days after calving the cows had almost reached 3% DMI of BW. This value is similar to the values recommended for high yielding dairy cows (NRC Citation2001).
Even with similar DMI, crossbred Holstein x Simmental cows had a better BCS during the pre and postpartum periods than had the purebred Holsteins (, A). The more favourable BCS seems to be closely related to the complementarity between the breeds used in the crossbreeding programmes. Simmental is a dual-purpose breed, which has a higher BCS than purebred Holstein cows (Sgorlon et al. Citation2015; Schweizer Citation2020). In our study, Holstein cows lost approximately 0.6 BCS points in the three weeks after calving, while the crossbred Holstein x Simmental cows lost approximately 0.4 BCS points. Młynek et al. (Citation2018) have also shown that Simmental cows decline less in BCS after calving than Holstein cows. We did not find a difference in the BW between breeds (, B). Both genetic groups lost approximately 100 kg of BW during the first three weeks of lactation. These values represent 12% and 15% of the BW for crossbred Holstein x Simmental cows and Holstein cows, respectively. The reduction in BW began in week 1 prior to calving, possibly due to the reduced DMI, which can amount to 30% (Dewhurst et al. Citation2000). A reason for these differences in BW between pre and postpartum may be related to the calving day itself, because at birth cows lose the calf weight and all the fluids originating from the uteri (at a minimum). For our two genetic groups, the calf weight represents on average 45 kg (Knob et al. Citation2016). Furthermore, BW and BCS loss are related to NEB during the first three weeks of lactation, while the energy requirement for milk production is higher than the DMI can provide (Mann et al. Citation2016; Barletta et al. Citation2017; Djoković et al. Citation2017; Youssef and El-Ashker Citation2017).
Our results suggest the genetic group did not affect BHB, because both genetic groups had similar milk yields and DMI ( and ). BHB is closely correlated with NEB, being a marker of oxidation of mobilized fat by the liver (Zachut et al. Citation2020). During NEB (partly characterized by lipomobilization) excess NEFA are converted to acetoacetate which is mainly metabolized to BHB and to a lower extent into acetone to maintain the energy homeostasis for milk production (Barletta et al. Citation2017; Bruckmaier and Gross Citation2017; Djoković et al. Citation2017; Youssef and El-Ashker Citation2017). An important factor regulating the calcium and energy homeostasis during the transition period – as most challenging and critical time in relation to the dairy cow’s health status during the lactation cycle – is serotonin, which acts independent of the parathyroid hormone (Hernández-Castellano et al. Citation2017). A treatment with 5-hydroxytryptophan (the immediate precursor to serotonin synthesis) could actually decrease the BHB concentration at days 6–10 postpartum, what might lead to a more stable energy homeostasis, particularly in Jersey cows (Weaver et al. Citation2017). By comparing Holstein with crossbred Holstein x Jersey cows at two weeks before and eight weeks after calving, Pelizza et al. (Citation2019) did, however, not find any difference between the breeds for BHB. They reported a difference between transition periods with higher BHB values after calving. No difference between breeds was also reported by Sgorlon et al. (Citation2015) when comparing BHB between Holstein and Simmental cows in Italy. In contrast, Lopreiato et al. (Citation2019) found that Simmental cows had lower BHB values, possibly due to the lower milk yield when compared to Holstein cows. Although, genetic groups did not differ, both of them reached BHB values higher than 1.1 mmol/l at the third week after calving, indicating a high use of body reserves to supply the energy requirements because of the high (and increasing) milk yield. Contrary to these observations, elevated BHB concentrations in combination with increased NEFA values are associated with a reduced milk yield (Zachut et al. Citation2020). One reason for the difference between the own results and the statement made in the review by Zachut et al. (Citation2020), might be that the ‘severity of the effects of elevated BHB varies depending on DIM at the onset of hyperketonemia’ (Santschi et al. Citation2016). Because our study stopped after the third week of lactation, we can only speculate that cows with an elevated level of BHB would show a relatively lower (or decreasing) milk yield during the following weeks of lactation in comparison to cows with a normal level of BHB.
The difference between transition periods with higher rumination times after calving is possibly related to a higher DMI after calving (). Cows spent approximately 51 min ruminating for each Kg of DMI before calving. After calving, these values decreased to approximately 32–34 min for each Kg of DMI. This difference may be related to the different diets offered to the cows during the prepartum and postpartum periods. The amount of concentrate offered to the cows increased from around 27% of the prepartum diet to 40% of the DM in the postpartum diet (). The ingredients of the concentrate ration undergo rapid degradation in the rumen and do not favour rumination, because of the lower fibre content (Kargar et al. Citation2010). Besides a lower DMI (especially during the week just before calving), other factors contributing to the lower rumination time (especially on the day before and at the calving day) are the birth mechanisms and body changes involved in calving. Schirmann et al. (Citation2013) observed a reduction of 60 min rumination time and 3.8 kg DMI 24 h before calving. Between 24 and 48 h after calving, the rumination time returns to the same values prior calving.
Both genetic groups produced similar amounts of milk during the first three weeks of lactation (). Similar or a little lower milk yields for crossbred cows have been reported in other studies that covered the entire lactation (Brähmig Citation2011; Knob et al. Citation2018; Nolte Citation2019). Generally, the milk yield increased quickly during the first week of lactation, starting at about 21 litres on the day of calving to close 30 liters/day after seven days of lactation. By lactation day 21, both genetic groups had a milk yield close to 40 liters/day; they had doubled their milk yield during this time. Additionally, the fat and protein content did not vary between the genetic groups. Both variables decreased from the first until the third week after calving. This is possibly due to the dilution effect as the amount of milk produced doubled during the same period.
One of the limitations of our study is the relatively small number of animals, especially regarding the crossbred Holstein x Simmental cows. This led to a relatively low power (1- beta error) of 0.67 for the statistical analysis. We think, however, that the results of our study are relevant and provide new insights into the transition period of crossbred Holstein x Simmental cows compared to Holstein cows.
Conclusion
After three weeks of lactation, DMI, milk yield, and all the other parameters keep changing. This study, however, demonstrates how the two different genetic groups (Holstein and crossbred Holstein x Simmental) handle the challenging transition period. Both genetic groups had a similar energy requirement prior and post calving. We have successfully demonstrated that the crossbreeding programme with Simmental semen, results in no negative effect on performance during the transition period in Holstein herds. The crossbred cows had the same dry matter intake and milk yield, and presented better body condition scores than purebred Holsteins. Crossbreeding between Holstein and Simmental might have a positive effect on the reproductive performance and lactation curve (persistency).
Declaration of interest
The authors declare that they have no conflict of interest.
Acknowledgements
We would like to thank the dairy farmer that kindly collaborated in this research. We also would like to thank to Ordemilk and Allfex® that kindly donate the cervical collars monitoring system for the realization of this research. This study was financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- AOAC International. 1998. Official Methods of Analysis. CD-ROM.AOAC International. Rockville: MD.
- Barletta RV, Maturana Filho M, Carvalho PD, Del Valle TA, Netto AS, Rennó FP, Mingoti RD, Gandra JR, Mourão GB, Fricke PM, et al. 2017. Association of changes among body condition score during the transition period with NEFA and BHBA concentrations, milk production, fertility, and health of Holstein cows. Theriogenology. 104:30–36. doi:10.1016/j.theriogenology.2017.07.030.
- Brähmig J. 2011. Einfluss der Wechselkreuzung von Deutschen Holsteins und Deutschem Fleckvieh auf Milchleistung und Milchqualität in einem automatischen Melksystem. Munich, Germany: Veterinary Medicine Faculty, Ludwig Maximilians University, 162 pp.
- Bruckmaier RM, Gross JJ. 2017. Lactational challenges in transition dairy cows. Anim Prod Sci. 57:1471–1481.
- Carvalho PD, Souza AH, Amundson MC, Hackbart KS, Fuenzalida MJ, Herlihy MM, Ayres H, Dresch AR, Vieira LM, Guenther JN, et al. 2014. Relationships between fertility and postpartum changes in body condition and body weight in lactating dairy cows. J Dairy Sci. 97(6):3666–3683. doi:10.3168/jds.2013-7809.
- Cassell BG, McAllister J. 2009. Dairy crossbreeding research : results from current projects. Virginia Cooperative Extension. 1–6. http://pubs.ext.vt.edu/404/404-094/404-094.html.
- Dewhurst RJ, Moorby JM, Dhanoa MS, Evans RT, Fisher WJ. 2000. Effects of altering energy and protein supply to dairy cows during the dry period. 1. Intake, body condition, and milk production. J Dairy Sci 83(8):1782–1794. doi:10.3168/jds.S0022-0302(00)75049-1.
- Djoković R, Kurćubić V, Ilić Z, Cincović M, Lalović M, Jašović B, Bojkovski J. 2017. Correlation between blood biochemical metabolites milk yield, dry matter intake and energy balance in dairy cows during early and mid lactation. Adv Diabetes Metab. 5(2):26–30. doi:10.13189/adm.2017.050202.
- Esposito G, Irons PC, Webb EC, Chapwanya A. 2014. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim Reprod Sci. 144(3–4):60–97. doi:10.1016/j.anireprosci.2013.11.007.
- Ferguson JD, Galligan DT, Thomsen N. 1994. Principal descriptors of body condition score in Holstein cows. J Dairy Sci. 77:2695–2703. doi:10.3168/jds.S0022-0302(94)77212-X.
- Hazel aR, Heins BJ, Seykora aJ, Hansen LB. 2013. Montbéliarde-sired crossbreds compared with pure Holsteins for dry matter intake, production, and body traits during the first 150 days of first lactation. J Dairy Sci. 96(3):1915–1923. doi:10.3168/jds.2012-5667.
- Heins BJ, Hansen LB, Seykora aJ, Johnson DG, Linn JG, Romano JE, Hazel aR. 2008. Crossbreds of Jersey x Holstein compared with pure Holsteins for production, fertility, and body and udder measurements during first lactation. J Dairy Sci. 91(3):1270–1278. doi:10.3168/jds.2007-0564.
- Hernández-Castellano LE, Hernandez LL, Sauerwein H, Bruckmaier RM. 2017. Endocrine and metabolic changes in transition dairy cows are affected by prepartum infusions of a serotonin precursor. J Dairy Sci. 100:5050–5057. doi:10.3168/jds.2016-12441.
- Kargar S, Khorvash M, Ghorbani GR, Alikhani M, Yang WZ. 2010. Short communication: effects of dietary fat supplements and forage:concentrate ratio on feed intake, feeding, and chewing behavior of Holstein dairy cows. J Dairy Sci 93(9):4297–4301. doi:10.3168/jds.2010-3168.
- Knob DA, Alessio DRM, Neto AT, Mozzaquatro FD, Thaler Neto A, Mozzaquatro FD. 2018. Growth, productive performance, and udder health of crossbred Holstein x Simmental cows and purebred Holstein cows. Semin Ciências Agrárias. 39:2597–2606. doi:10.5433/1679-0359.2018v39n6p2597.
- Knob DA, Alessio DRM, Thaler Neto A, Mozzaquatro FD. 2016. Reproductive performance and survival of Holstein and Holstein x Simmental crossbred cows. Trop Anim Health Prod. 48(7):1409–1413. doi:10.1007/s11250-016-1103-9.
- Leblanc S. 2010. Monitoring metabolic health of dairy cattle in the transition period introduction—metabolic challenges in peripartum dairy cows and their associations with reproduction. J Reprod Dev Reprod Dev. 56(56):S29–S35.
- Lopreiato V, Minuti A, Trimboli F, Britti D, Morittu VM, Cappelli FP, Loor JJ, Trevisi E. 2019. Immunometabolic status and productive performance differences between periparturient Simmental and Holstein dairy cows in response to pegbovigrastim. J Dairy Sci. 102:9312–9327. doi:10.3168/jds.2019-16323.
- Mann S, Nydam DV, Lock AL, Overton TR, McArt JAA. 2016. Short communication: Association of milk fatty acids with early lactation hyperketonemia and elevated concentration of nonesterified fatty acids. J Dairy Sci 99(7):5851–5857. doi:10.3168/jds.2016-10920.
- McCarthy MM, Mann S, Nydam DV, Overton TR, McArt JAA. 2015. Short communication: concentrations of nonesterified fatty acids and β-hydroxybutyrate in dairy cows are not well correlated during the transition period. J Dairy Sci 98(9):6284–6290. doi:10.3168/jds.2015-9446.
- Mendonça LGD, Abade CC, da Silva EM, Litherland NB, Hansen LB, Hansen WP, Chebel RC. 2014. Comparison of peripartum metabolic status and postpartum health of Holstein and Montbéliarde-sired crossbred dairy cows. J Dairy Sci. 97(2):805–818. doi:10.3168/jds.2013-7159.
- Mertens DR. 2002. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: Collaborative study. J AOAC Int. 85(6):1217–1240.
- Młynek K, Głowińska B, Salomończyk E, Tkaczuk J, Styś W. 2018. The effect of daily milk production on the milk composition and energy management indicators in Holstein–Friesian and Simmental cows. Turkish J Vet Anim Sci. 42(4):223–229. doi:10.3906/vet-1711-31.
- Nolte O. 2019. Effekte der Rückkreuzung auf Fleckvieh aus einer Kreuzungsherde mit Deutschen Holstein-Genanteilen auf Milchleistung und Milchqualität. Munich, Germany: Veterinary Medicine Faculty, Ludwig Maximilians University.
- NRC. 2001. Nutrient requirement Tables, 7th ed. Washington, DC: Natl. Acad. Press.
- Olson KM, Cassell BG, Hanigan MD. 2010. Energy balance in first-lactation Holstein, Jersey, and reciprocal F 1 crossbred cows in a planned crossbreeding experiment. J Dairy Sci 93(9):4374–4385. doi:10.3168/jds.2010-3195.
- Palladino RA, Buckley F, Prendiville R, Murphy JJ, Callan J, Kenny DA. 2010. A comparison between Holstein-Friesian and Jersey dairy cows and their F1 hybrid on milk fatty acid composition under grazing conditions. J Dairy Sci. 93(5):2176–2184. doi:10.3168/jds.2009-2453.
- Pelizza A, Hauser A, Mendes TC, Mattiello CA, Knob DA, Cardozo LL, Parizotto Filho R, Gomes IPO, Alessio DRM, Camera M, Thaler Neto A. 2019. Perfil metabólico de vacas Holandês e mestiças Holandês x Jersey no periparto. Arq Bras Med Veterinária e Zootec. 71(3):741–751. doi:10.1590/1678-4162-10098.
- Prendiville R, Pierce KM, Buckley F. 2009. An evaluation of production efficiencies among lactating Holstein-Friesian, Jersey, and Jersey x Holstein-Friesian cows at pasture. J Dairy Sci. 92(12):6176–6185. doi:10.3168/jds.2009-2292.
- Prendiville R, Pierce KM, Delaby L, Buckley F. 2011. Animal performance and production efficiencies of Holstein-Friesian, Jersey and Jersey x Holstein-Friesian cows throughout lactation. Livest Sci. 138(1–3):25–33. doi:10.1016/j.livsci.2010.11.023.
- Santschi DE, Lacroix R, Durocher J, Duplessis M, Moore RK, Lefebvre DM. 2016. Prevalence of elevated milk beta-hydroxybutyrate concentrations in Holstein cows measured by Fourier-transform infrared analysis in Dairy Herd Improvement milk samples and association with milk yield and components. J Dairy Sci. 99(11):9263–9270. doi:10.3168/jds.2016-11128.
- SAS. 2002. Statistical analysis system. Cary, North Carolina, USA: SAS Institute Inc.
- Schirmann K, Chapinal N, Weary DM, Vickers L, von Keyserlingk MAG. 2013. Short communication: rumination and feeding behavior before and after calving in dairy cows. J Dairy Sci 96(11):7088–7092. doi:10.3168/jds.2013-7023.
- Schweizer H. 2020. Vergleich von deutschen und neuseeländischen Milchproduktionssystemen mit dem Schwerpunkt Tiergesundheit [Dissertation Thesis]. Munich, Germany: Tierärztliche Fakultät, Lehr- und Versuchsgut Oberschleißheim, Ludwig Maximilians Universität München.
- Sgorlon S, Fanzago M, Sandri M, Gaspardo B, Stefanon B. 2015. Association of index of welfare and metabolism with the genetic merit of Holstein and simmental cows after the peak of lactation. Ital J Anim Sci. 14(3):368–373. doi:10.4081/ijas.2015.3841.
- Tyrrell H F, Reid J T. 1965. Prediction of the energy value of cow's milk. J. Dairy Sci. 48:1215–1223. doi: 10.3168/jds.S0022-0302(65)88430-2.
- Weaver SR, Prichard AS, Maerz NL, Prichard AP, Endres EL, Hernandez-Castellano LE, Akins MS, Bruckmaier RM, Hernandez LL. 2017. Elevating serotonin pre-partum alters the Holstein dairy cow hepatic adaptation to lactation. PLoS ONE. 12(9):e0184939. doi:10.1371/journal.pone.0184939.
- Weber C, Hametner C, Tuchscherer A, Losand B, Kanitz E, Otten W, Singh SP, Bruckmaier RM, Becker F, Kanitz W, Hammon HM. 2013. Variation in fat mobilization during early lactation differently affects feed intake, body condition, and lipid and glucose metabolism in high-yielding dairy cows. J Dairy Sci. 96(1):165–180. doi:10.3168/jds.2012-5574.
- Weigel Ka, Barlass Ka. 2003. Results of a producer survey regarding crossbreeding on US dairy farms. J Dairy Sci. 86(12):4148–4154. doi:10.3168/jds.S0022-0302(03)74029-6.
- Youssef M, El-Ashker M. 2017. Significance of insulin resistance and oxidative stress in dairy cattle with subclinical ketosis during the transition period. Trop Anim Health Prod. 49(2):239–244. doi:10.1007/s11250-016-1211-6.
- Zachut M, Šperanda M, De Almeida AM, Gabai G, Mobasheri A, Hernández-Castellano LE. 2020. Biomarkers of fitness and welfare in dairy cattle: healthy productivity. J Dairy Res. 87(1):4–13. doi:10.1017/S0022029920000084.