Abstract
A section near the Kilgen Lake (Adana Province) has yielded a fauna of rugose and tabulate corals including Disphyllum cf. rugosum (Wedekind, 1922), D. cf. curtum Hill, 1954, Wapitiphyllum sp., Thamnopora sp., and Alveolites sp. This coral assemblage is consistent with the previous Frasnian age assigned to the limestones of the Gümüşali Formation. The rather well-preserved material provides new data on the structure and microstructure of Disphyllum and allows to describe in Thamnopora unusual calicinal morphology (septal ridges, median teeth, and pseudopercula) as well as new structures linked to the lateral increase (basal low wall, apical cul-de-sac).
Introduction (O.M. & R.G.)
The aim of this study is to complete the inventory of the benthic fauna of the Gümüşali Formation by that of the rugose and tabulate corals and to test the age previously obtained, based on brachiopods (Gourvennec, Citation2006). The fossil material described here was collected in the eastern Taurus, near Kozan, in a succession ranging from the infra-Cambrian to the Tertiary (Özgül et al., Citation1973).
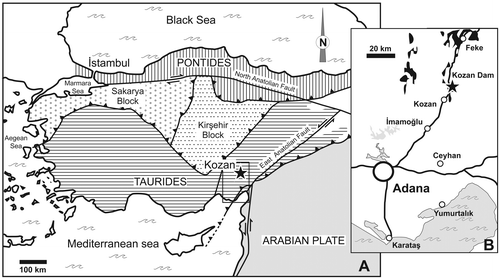
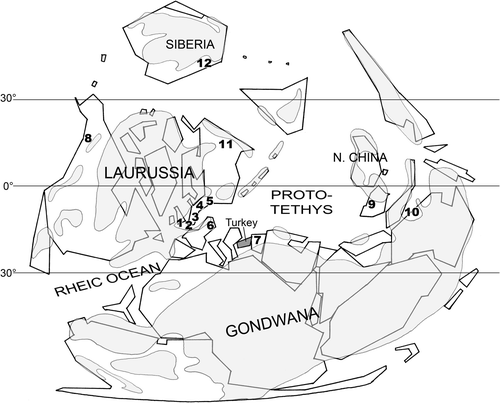
Turkey is a structurally complex assemblage of several microplates which occurred in Mesozoic and Tertiary times (Figure ). Schematically, to the north, the Pontides Chain exhibits Eurasian affinities whereas to the south, the Taurus Chain was part of Gondwana, as well as southeastern Turkey which is situated on the northern edge of the Arabian Plate. In the eastern Taurus, in spite of tectonic disturbances, an overall regressive trend has been recognized in the Devonian succession which conformably follows the Silurian transgressive episode (Wehrmann et al., Citation2010). Brachiopods collected in the Kozan section (Gümüşali Fm.) show Gondwanan affinities and indicate a Late Devonian age, probably the Frasnian (Gourvennec, Citation2006). Further to the north, two continuous Devonian sections (Kocadere and Halevikdere sections, Wehrmann et al., Citation2010) have been described from Lockhovian to Famennian; however in the latter, the brachiopod fauna is lacking in distinctive characteristics to precise its palaeogeographic affinity.
Figure 1. Simplified tectonic map of Turkey showing (A) major terranes and (B) location of the studied section (black star) (after Yalçin & Yılmaz, Citation2010, modified). Devonian outcrops are figured in black.
A conspicuous Palaeozoic succession lies near the dam closing the Kilgen Lake, 12 km north of Kozan, on the road from Adana to Sarız (Figure ). In this area, Lower Palaeozoic strata form a large outcrop which extends along both sides of the lake. The succession ranges in age from Middle Cambrian in the east to Silurian in the west where the strata are faulted against Devonian carbonates. Cambrian carbonates are exposed along the eastern margin of the Kilgen Lake (Figure ) and include the typical red nodular limestone Member of the stratotype in the western Taurus (Dean & Monod Citation1970; Dean, Citation2005). The highest Cambrian rocks are overturned and the base is not exposed. Above, occupying much of the floor of the lake comes the Seydişehir Formation (late Cambrian-Arenig), which has a large extension in the Taurus (Özgül et al., Citation1973) and is typical of the peri-Gondwanaland northern provinces. The youngest strata of the Seydişehir Formation are well exposed near the forestry Fire Station (see map Figure ) and consist of alternating grey shales and sandstones containing nodules with asaphid trilobites, orthid brachiopods, and rare orthocone cephalopods, a comparable assemblage to the type area (Monod, Citation1967). Moreover here, a small graptolitic fauna including Phyllograptus sp. and Corymbograptus v-deflexus (Harris, Citation1924) indicates the middle part of the Upper Arenig (Dean, Monod, Ghienne, Kozlu, & Günay, Citation2003). It is followed by a discontinuous limestone horizon with cystoids and brachiopods (Llanvirn) and silty shales with graptolites, chitinozoans, and acritarchs of Sandbian to Early Katian age (Paris et al., Citation2007) which are equated with part of the Bedinan Formation in eastern Turkey (Dean, Citation1967, Citation1983). Above, an erosional unconformity (Figure ) truncates the underlying strata, from the Caradoc in the north down to the Arenig along the southern shore of the lake. This unconformity is due to the arrival of coarse polymict conglomerates and sandstones of glacial origin (Halevikdere Fm., Hirnantian) (Ghienne, Monod, Kozlu, & Dean, Citation2010; Monod et al., Citation2003). These glacial deposits are followed in continuity by graptolitic black shales of Early-Middle Silurian age (Kozlu, Sachanski, & Göncüoglu, Citation2012). Above, in faulted contact, Upper Devonian strata (Gümüşali Fm.) contain a rich brachiopod and coral fauna, typical of the peri-reefal environment of this formation in the eastern Taurus (Yalçın & Yılmaz, Citation2010).
Figure 2. Geological map of the studied area, along the Kilgen Lake, NE of Kozan. The fossil locality is indicated as a black star.
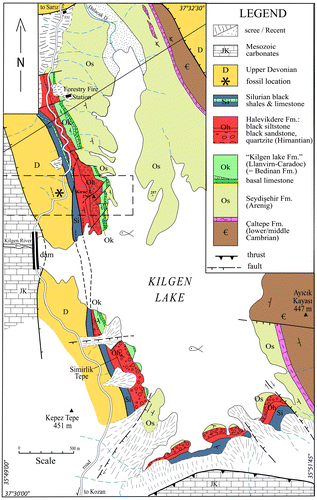
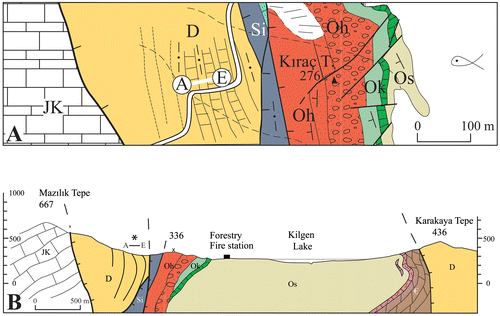
Figure 3. Detailed map of the fossil locality (A) and approximately W–E section (B) of the studied area.
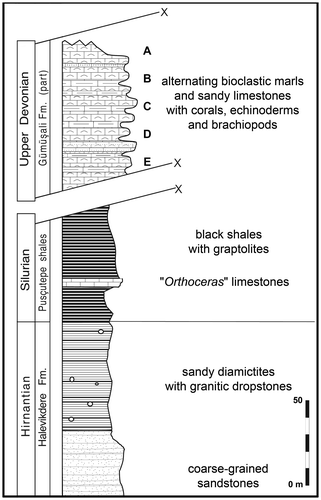
Three levels (Figure ) have yielded corals (Rugosa and Tabulata) and, from the base to the top, the following species are recorded: Disphyllum cf. rugosum (Wedekind, Citation1922) (level PK66 D), Wapitiphyllum sp. and Alveolites sp. (PK66 B), Disphyllum cf. curtum Hill, Citation1954 and Thamnopora sp. (PK66 A). This coral assemblage is consistent with the previous Frasnian age assigned to the limestones of the Gümüşali Formation.
Figure 4. Diagrammatic sketch of the top of the succession in the Kozan section along the road of the Kilgen Lake, with samples location.
Preliminary remarks
The material studied herein was collected by Olivier Monod in 2001; it is housed at the Université de Bretagne Occidentale, Brest (France), Laboratoire de Paléontologie, catalogue number LPB 19360–19390. In addition, the thin sections’ numbers (prefix Bb or L) are given for the illustrated sections.
Original skeleton material used in this study is preserved in calcite and most data were obtained from transverse and longitudinal standard thin sections (ca. 30 μm), acetate peels, and polished ultra-thin sections, the so-called “LFP” (Lafuste, Citation1970). Generally speaking, some (micro-) structures such as the “median dark line” are difficult to see, even not seen in acetate peels; it is the reason why the figure captions always indicate “thin section” or “acetate peel”.
The terms proximal and distal thin sections or acetate peels are here used for transverse sections made in both extremities of the short available fragments of corallite.
In all figures, the septa of Rugosa are identified as follow: black triangle = protoseptum, black circle = major septum, open circle = minor septum and half black circle = minor-like septum; cardinal septum (when identified) always in “north” position. Except for the photos, the figures are camera lucida drawings.
Systematic palaeontology
Rugose corals (M. C.-A. & Y.P.)
In addition to the specimens of Disphyllum and Wapitiphyllum described herein, the Gümüşali Formation has yielded some unidentified fragments of Rugosa mainly belonging to disphyllid representatives. They were collected in levels PK66 D (LPB 19364, LPB 19384 and LPB 19385, probably a trochoid solitary coral) and PK66 A (LPB 19361- 363, LPB 19377).
Subclass Rugosa Milne-Edwards and Haime, Citation1850
Order Stauriida Verrill, Citation1865
Family Disphyllidae Hill, Citation1939
Genus Disphyllum De Fromentel, Citation1861
Type species. By subsequent designation of Lang and Smith (Citation1934, p. 80), Cyathophyllum caespitosum Goldfuss, Citation1826.
Diagnosis. Fasciculate rugose corals. Septa of two orders, occasionally carinate, more or less dilated in the dissepimentarium and thin in the tabularium. Major septa reaching the axis of the corallites or leaving an open space in the centre of the tabularium. Minor septa traversing the entire dissepimentarium. Dissepimentarium composed of several rows of globose dissepiments, often arranged in horizontal layers in its outer part and inclined towards the axis of the corallites in its inner part. Tabulae usually incomplete or compound.
Disphyllum cf. rugosum (Wedekind, Citation1922)
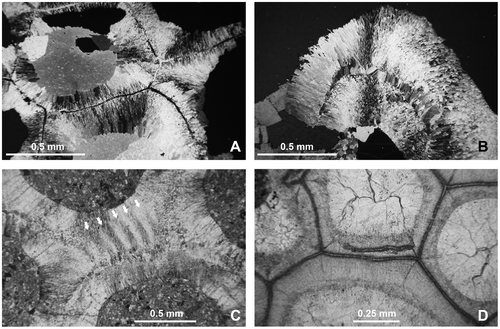
Figures –, A, A–D
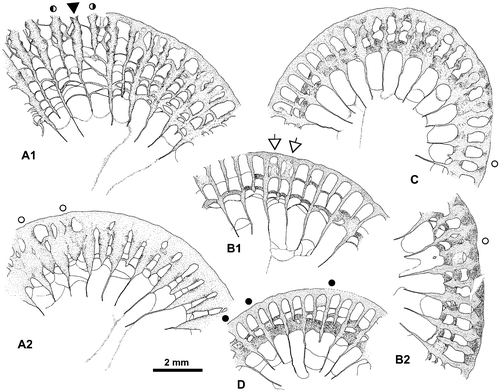
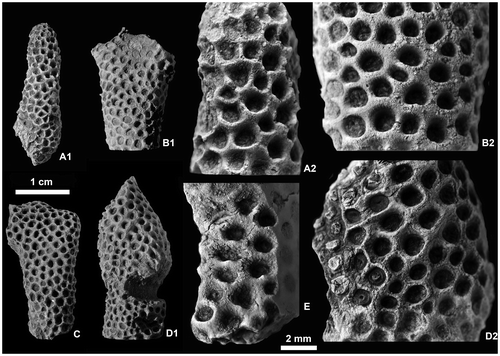
Figure 5. Disphyllum cf. rugosum (Wedekind, Citation1922). Views of corallites in transverse sections. A1, A2. Two opposite sectors of the same section. (A1), major and minor septa moderately thickened and bearing knobbly carinae; note two clearly shorter minor-like septa on both sides of a rather short major septum, this latter could be the cardinal one; (A2), septa strongly dilated and smooth in the dissepimentarium; note two slightly shorter and somewhat stocky minor septa on both sides of a major septum opposite the alleged cardinal, probably the counter septum. LPB 19386, distal thin section L 3154a. B1, B2. Two opposite sectors of the same section, with septa narrow and non-carinate (B1) or more thick with carinae (B2); note an unusual discontinuous minor septa and x-shaped motif (arrows) characteristic of saddle-like dissepiments in tangential section (i.e. concave parallel to the margin of corallum and convex parallel to the septa). LPB 19375, proximal acetate peel. C. Variations of septal morphology depending of the sector; note minor septum entering in the tabularium and the thickening of the inner layer of dissepiments. LPB 19376, distal acetate peel (= Figure D). D. Rather non-carinate narrow septa; some major septa are shorter and display a random setting; thus, the protosepta cannot be identified; note the development of inner wall. LPB 19389, proximal acetate peel (see distal section in Figure C1).
cf. Citation1922 Schlüteria rugosa Wdkd; Wedekind: 5, Figures –.
cf. Citation2005 Disphyllum rugosum (Wedekind, Citation1922); Schröder: 92, pl. 14: Figures –.
cf. Citation2009 Disphyllum rugosum (Wedekind, Citation1922); Coen-Aubert: 70, pl. 3: Figures –.
Lectotype. Figures 3–4 in Wedekind (Citation1922) and pl. 1, Figures 4–5 in Bulvanker (Citation1941), chosen by Bulvanker (Citation1941, p. 134) according to Rohart (Citation1988, p. 252). Specimen of the Wedekind collection stored in the Forschungsinstitut Senckenberg at Frankfurt am Main, Germany. Probably Frasnian of Refrath in the Bergisches Land, Germany.
Material. 6 specimens with 7 thin sections (of which 2 “LFP”) and 13 acetate peels. Level PK66 D: LPB 19375, 19376, 19386–19389.
Diagnosis. A species of Disphyllum with 42–52 septa at a diameter of 7–13 mm. Septa carinate and dilated in the dissepimentarium, thin in the tabularium. Major and minor septa rather long. Tabulae incomplete, frequently with flat-topped axial parts.
Description. The material consists of cylindrical corallites with a few lateral offsets. Longitudinal ribs and growth lines are occasionally observed. The outer wall is often well preserved.
The septa are non-carinate or bear some small spinose and knobbly carinae. They are typically dilated in the dissepimentarium and become usually thin beyond their entry into the tabularium (Figure ). A stereoplasmic thickening is locally present between the septa or on an inner layer of dissepiments. This deposit forms a nearly complete inner wall in specimen LPB 19389 (Figure D).
The major septa leave an extensive open space in the center of the corallites. The minor septa traverse the entire dissepimentarium or even enter into the tabularium; they may be slightly shorter.
The dissepimentarium consists of 1–5 rows of globose dissepiments which are in horizontal layers in its outer part and inclined in its inner part; in specimens LPB 19387 and 19389, a few more or less peneckielloid dissepiments are present at the periphery. The tabulae are incomplete and intersecting laterally; their axial parts are occasionally flat-topped.
There are 46–56 septa per corallite. The diameter of the corallites ranges from 9 to 13 mm whereas the width of the tabularium varies between 5.8 and 8.6 mm and even reaches 9.5 mm in LPB 19375.
Microstructure. Only few microstructural data are available for the genus Disphyllum, and generally they only consist of descriptions of trabeculae (Bronwlaw & Jell, Citation2008; Hill Citation1981; Rohart Citation1988). Thus, some of the following data are new. Detailed data about the microstructure of our material are provided both by standard thin sections and mainly by “LFP” thin sections.
The wall consists of two parts (data from “LFP” transverse section, Figures –):
A very thin layer of radially oriented fibroids (size of fibroids ca. 5–8 × 25–30 μm).
An inner thick lining of lamellae (the main part of the wall, .0.25 mm) bridging the space between the peripheral edge of the septa. The asymmetrical outlines of the cupulate lamellae indicate the centripetally direction of skeletal accretion. Size of the lamellae ca. 20–30 even 40 × 4–5 μm.
Figure 6. Disphyllum cf. rugosum (Wedekind, Citation1922). Peripheral part of a transverse section showing the microstructure of the wall (W) and some dissepiments (Ds) (synthesis between observations in crossed nicols and natural light); explanations in text. LPB 19387, “LFP” I.
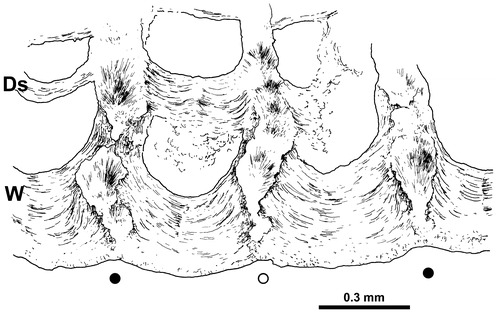
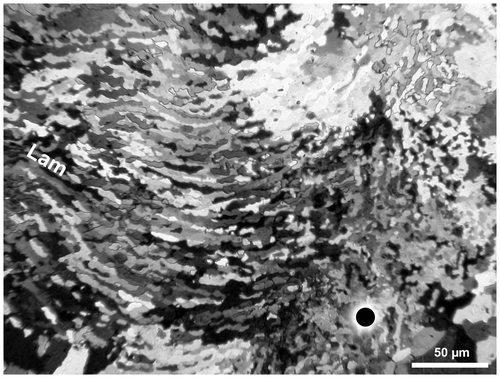
Figure 7. Disphyllum cf. rugosum (Wedekind, Citation1922). Photomicrograph to illustrate the microstructure of the wall, transverse section, crossed nicols; outer layer of fibroids (Fr), lamellae of the inner lining (Lam), minor septum on right, major septum outside of left margin of photo and axis of corallum towards the top of figure. LPB 19387, “LFP” I.

In longitudinal sections (standard thin sections), the lamellae of the wall (at least the outer part of the lining) are inclined upwards and inwards (Figure B).
Figure 8. Disphyllum cf. rugosum (Wedekind, Citation1922). Trabecular microstructure of two septa in longitudinal/tangential section (detailed caption in text). A. LPB 19387, rather thick thin section. B. Note the inclined lamellae in the wall (arrow). LPB 19388, thin section Bb 1408.
This mural structure is generally assigned to archeotheca (or archaeotheca) by the authors dealing with Rugosa. Its features seem to be consistent with the characteristics of the wall of Cyathaxonia cornu Michelin (Citation1847), designated by Weyer (Citation1975) as the type example of this category. However, Stolarski (Citation1995) “proposes to reject the term archeotheca as incorrectly defined”, but this is another story. In addition, according to Roniewicz and Stolarski (Citation1999), the wall of D. cf. rugosum suggests a significant similarity with the “epithecate wall” (Roniewicz & Stolarski Citation1999, Figure 1A). Nevertheless, in the examples studied by these authors (Scleractinia), the inner epithecal layer is built of radially arranged fibers (however, they grow centripetally as the lamellae in Disphyllum) and the fibers of the outer layer “are oriented distally, rarely centripetally” (Stolarski, Citation1996, p. 342).
The septa exhibit a trabecular microstructure, well exposed in longitudinal/tangential sections (Figure ), consisting of parallel and contiguous trabeculae sloping upwards and inwards from the periphery at varying angles (generally ca. 30°, rarely up to 60°). Dentations on the inner margin of the septa could be exposed (LPB 19389). In some cases, the trabeculae appear to be progressively inclined, projecting close to the horizontal in the proximal part of the septum, and at moderate angle (35°) in its distal part (Figure A). A close feature has been described and well illustrated by Brownlaw and Jell (Citation2008, Figure 25). A single specimen (LPB 19386) possesses trabeculae directed at moderate angle from the periphery (28°–30°), flexing at 8°–10° at the tabularium boundary. When the section is not exactly in the median plane of the septum, the trabeculae seem to be not contiguous (see upper part of Figure B) and, probably, the setting of fibers in successive bundles within the trabeculae is emphazised (Figures B and A). Also, depending of the position of the section with regard to the median plane of the septum, its inner margin could show dentations (Figure C2 upper right) reflecting an underlying trabecular microstructure. Diameter of trabeculae measured on longitudinal sections, mostly between 0.20 and 0.33 mm.
The dissepiments are built of lamellae (Figure ) showing the same size and morphology as those of the wall. In places, within the dissepimentarium, the septa are coated by a thin layer of lamellae.
Figure 9. Disphyllum cf. rugosum (Wedekind, Citation1922). Microstructure of a dissepiment of the thickened inner layer in transverse section, crossed nicols; lamellae (Lam), dissepiment concave towards the axis of corallum (top of fig.). Oblique section of a major septum on the right of figure. LPB 19387, “LFP” II (see also overall section Figure B1).
Discussion. The Middle Frasnian material of Disphyllum rugosum collected in various areas of Belgium has been investigated in detail by Coen-Aubert (Citation2009). The six corallites of Turkey assigned to D. cf. rugosum differ from it by septa less carinate and typically dilated in the dissepimentarium, by some stereoplasmic thickenings and by a wide open space in the centre of the tabularium; additionally, the major septa of the Turkish specimens are often thinning beyond their entry into the tabularium. McLean (Citation2010, p. 32) referred to D. rugosum Middle Frasnian colonies coming from Alberta and the Northwest Territories in Canada which are different from this taxon in having few carinae, a stereozone in some corallites and a somewhat smaller septal number giving a looser texture in transverse section. In fact, McLean (Citation2010) synonymises D. rugosum with D. kostetskae (Soshkina, Citation1949) introduced in the Frasnian of the Urals in Russia. On the contrary, D. rugosum and D. kostetskae are considered as two separate species by Coen-Aubert (Citation2009, p. 71). The Late Frasnian material from the British Columbia, Alberta and the Northwest Territories identified by Mclean (Citation2010, p. 35) as D. catenatum Smith, Citation1945, is very similar to D. rugosum and is distinguished from the Turkish sampling by slightly fewer septa, smaller corallites and longer major septa.
Superficially, there seems to be some affinities between D. cf. rugosum and the Turkish specimen from the Middle Devonian of Feke in eastern Taurus ascribed to D. caespitosum emsti (Wedekind, Citation1922) by Flügel and Flügel (Citation1961, p. 382). However, the latter colony is different in having no carinae, major septa reaching the axis of the corallites and minor septa often extending a long way into the tabularium where they may be contratingent. The lectotype of D. emsti coming from the Early Givetian Oberhonsel Formation of the Sauerland in Germany has been selected by Flügel and Flügel (Citation1961) and is only represented by the longitudinal section figured by Wedekind (Citation1922, Figure 1). So, D. emsti is inadequately known; it is the type species of Schlueteria Wedekind, Citation1922, which is commonly considered as a synonym of Disphyllum as mentioned among others by McLean (Citation2010, p. 20). In fact, the corallite illustrated by Flügel and Flügel (Citation1961) resembles more D. caespitosum (Goldfuss, Citation1826) from which it is separated by the stronger dilation of the septa in the dissepimentarium and the greater length of the minor septa that is striking. The lectotype of D. caespitosum has been collected in the Givetian of the Bergisches Land in Germany and this species occurs also in the Early Givetian of Belgium where it has been revised by Coen-Aubert (Citation2008).
In transverse section, D. cf. rugosum based on the Turkish material is related to Sinodisphyllum kielcense (Rozkowska, Citation1979) reported in the Middle Frasnian of Belgium by Boulvain and Coen-Aubert (Citation2006). However, the latter species is characterized by solitary coralla having more septa for a greater diameter, a wider dissepimentarium and longer major septa as well as by the thickening of the septa which is weaker and restricted to the dissepimentarium.
Geographic and stratigraphic range. Besides the questionable material from Turkey, Disphyllum rugosum is known in the Middle Frasnian from the Bergisches Land in Germany and from various areas of Belgium which have been detailed by Coen-Aubert (Citation2009). In both countries, it seems that the species has been collected in the Palmatolepis punctata conodont Zone (base of the Middle Frasnian).
Disphyllum cf. curtum Hill, Citation1954
Figures –B, E–H
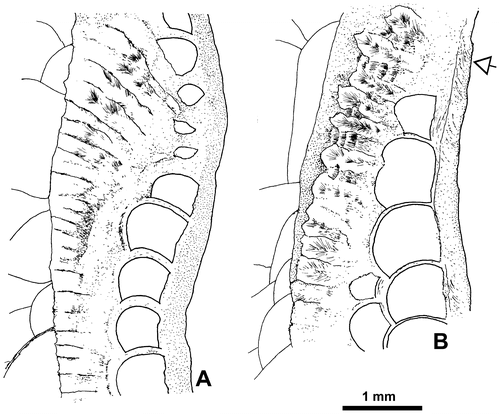
Figure 10. Disphyllum cf. curtum Hill, Citation1954. Views of corallites in transverse and longitudinal sections. A1, A2. Two closely spaced proximal transverse sections with the same orientation. (A1), cardinal sector (inferred from section A2) showing five major septa with large discontinuities in the tabularium, acetate peel ( = Figure F1); (A2), section exhibiting a well-marked bilateral symmetry emphasized by a pair of discontinuous major septa and the alar protosepta longer than the metasepta located on their counter side; cardinal sector on top of figure, counter sector on bottom and corresponding protosepta not identified (thin section Bb 1306, corresponding acetate peel Figure F2). LPB 19381. B1, B2. Respectively proximal and distal transverse sections (acetate peels); (B1) (= Figure H1), section showing a bilateral symmetry, note on both sides of the thickened cardinal septum a pair of shorter minor-like septa, a pair of short major septa, the gradual shortening of the metasepta especially on the counter side of the right alar septum and opposite to the cardinal septum a slightly longer major septum identified as the counter septum; (B2), probably counter sector, note two strongly reduced minor septa and defective herringbone dissepiments (open arrows). LPB 19383. C1, C2. Respectively proximal transverse and longitudinal sections (acetate peels); (C1), note a reduced major septum, other reduced septa occur in the section, but no bilateral symmetry is exhibited; (C2), parent corallite and offset, section showing numerous flat topped incomplete tabulae in the axial zone of the tabularium, and in the tangential sections of the septa, the trabeculae flexing considerably at the tabularium boundary. LPB 19379. D. Longitudinal section (acetate peel), parent corallite and two offsets on its left side, scarce complete tabulae. LPB 19380 (see proximal transverse section Figure G).
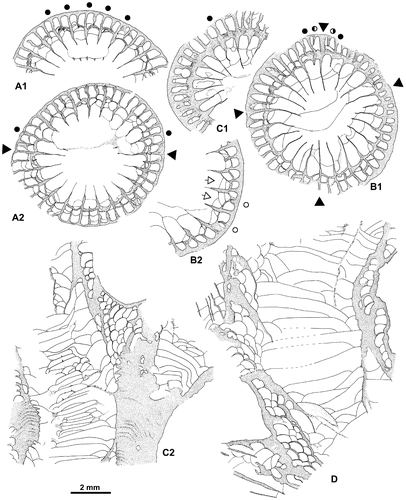
Figure 11. Septal microstructure, trabeculae in longitudinal section. A. Disphyllum cf. rugosum (Wedekind, Citation1922). LPB 19388, thin section Bb 1408 (see also Figure B). B. Disphyllum cf. curtum Hill, Citation1954. LPB 19379, thin section Bb 1409 (see also Figure C2 where the flexing of trabeculae is best exposed; acetate peel corresponding to the thin section).

cf. Citation1954 Disphyllum curtum sp.nov.; Hill: 22, pl. 2: Figure 8.
cf. Citation1970 Disphyllum curtum Hill, Citation1954; Hill and Jell: 42, pl. 7: Figure 11, pl. 8: Figure 3.
Holotype. Pl. 2, Figure 8 in Hill (Citation1954). Specimen UWA 33518 stored in the Geology Museum from the University of Western Australia at Perth. Section south of Mount Wilson, West Kimberleys, Canning Basin, Western Australia. Lower part of the Pillara Limestone, Frasnian.
Material. 7 specimens with 5 thin sections and 16 acetate peels. Level PK66 A: LPB 19360, 19378 to 19383
Diagnosis. A species of Disphyllum with 42–46 septa at a diameter of 6–11 mm. Septa non-carinate and slightly dilated in the dissepimentarium. Major septa withdrawn from the axis of the corallites. Tabulae incomplete.
Description. The material consists of cylindrical corallites which may be curved or even conical, with some lateral offsets. Longitudinal ribs are frequently observed. The outer wall is more or less preserved.
The septa are non-carinate or bear locally a few small spinose and knobbly carinae. They are faintly dilated in the dissepimentarium and become thin in the tabularium or slightly beyond their entry into it (Figure ). Sometimes, there is a weak deposit of stereoplasma on an inner layer of dissepiments. An inner wall is present in the young stage of LPB 19379, close to the tip of the corallite (Figure C1).
The major septa leave a more or less extensive open space in the center of the tabularium or nearly reach the axis of the corallites; poor pseudofossulae are often observed. The minor septa traverse all, nearly all or half the dissepimentarium or even enter into the tabularium. They are occasionally contratingent, shorter, reduced to spines and rarely lacking; a few herringbone dissepiments are present in the inner dissepimentarium of several corallites.
The dissepimentarium consists of 1–5 rows of globose dissepiments which are in horizontal layers in its outer part and inclined in its inner part; in specimen LPB 19383, some peripheral dissepiments are larger and more or less peneckielloid. The tabulae are generally incomplete and intersecting laterally; their axial parts are rather often flat-topped and sometimes horizontal, concave or convex.
There are 42–50 septa per corallite. The diameter of the corallites ranges from 6.5 to 11.5 mm whereas the width of the tabularium varies between 3.5 and 7.6 mm.
Microstructure. The microstructure is roughly identical to that of D. cf. rugosum; however, “LFP” are not available and thus minute details are lacking.
The wall structure probably belongs to the archeotheca sensu Weyer (Citation1975). As shown by specimen LPB 19381 (Figure A2, cardinal area), the septa are clearly not contiguous and their peripheral ends, sharply demarcated from the wall, appear wedged into this structure. In longitudinal section, the lamellae of the wall appear projected inwards and upwards as in D. cf. rugosum. The same feature can be observed in the wall of Cyathaxonia cornu (Plusquellec unpublished). In the longitudinal thin section of specimen LPB 19379 (area not illustrated in Figure B), two short segments of dark line within the wall mark the boundary between parent corallite and offset. This structure has already been mentioned by Coen-Aubert (Citation2008) in D. caespitosum (Goldfuss, Citation1826).
The septa are built of contiguous trabeculae sloping upwards and inwards (25°–45°); their inner margin show dentations (Figure H2 left). In the specimen LPB 19379, the trabeculae are moderately declined at the periphery, but are flexed in the middle dissepimentarium to become near horizontal or slightly below the horizontal in the tabularium boundary (Figure C2 and Figure B); this feature, unusual in our material, is described in numerous specimens belonging to eight species of Disphyllum and Disphyllum? from the Pillara limestone by Brownlaw and Jell (Citation2008, see especially Figure 29C and D).
Discussion. The seven Turkish corallites assigned to Disphyllum cf. curtum are distinguished from D. cf. rugosum by slightly smaller septal number and diameters of the corallites and tabularia, by septa less carinate and thickened in the dissepimentarium, by minor septa not traversing systematically the entire dissepimentarium and by major septa which occasionally nearly reach the axis of the corallites. There are only a few Frasnian colonies of Western Australia that have been described as D. curtum by Hill (Citation1954) and Hill and Jell (Citation1970). This sampling shows major septa withdrawn from the center of the tabularium. Contrary to the opinion of Brownlaw and Jell (Citation2008, p. 32), D. curtum is not considered as a synonym of D. virgatum (Hinde, Citation1890) whose lectotype from the Early Frasnian of the Canning Basin in Western Australia has been illustrated among others by Coen-Aubert (Citation1989). Indeed, this species investigated by this author at the top of the Givetian from Belgium is characterized by larger corallites, by longer major septa and by septa strongly dilated in the dissepimentarium with stereoplasmic thickenings.
D. cf. curtum is also very similar to D. grabaui Tsien, Citation1970, from the Early Frasnian of Belgium. However, the latter taxon studied in detail by Coen-Aubert (Citation2009) is separated from the former by minor septa of more variable length and by somewhat larger corallites and tabularia. Finally, there are some affinities between D. cf. curtum and the Middle Frasnian corallites of Central Iran referred by Rohart (Citation2000, p. 68) to Disphyllum sp. 1. But, the minor septa of this material are extending across the entire dissepimentarium whereas other Frasnian specimens of Iran ascribed by Rohart (Citation1999, p. 51) to Disphyllum sp. 1 are more different by the occurrence of carinae.
Geographic and stratigraphic occurrence. Besides the questionable material from Turkey, Disphyllum curtum is known in the Frasnian of the Canning Basin in Western Australia.
Genus Wapitiphyllum McLean and Pedder, Citation1984
= Donia Soshkina, Citation1951
= Disphyllia sensu Hill, Citation1981, non He, Citation1978
= Cystihexagonaria Rohart, Citation1988
Type species. By original designation, Wapitiphyllum vallatum McLean and Pedder, Citation1984.
Diagnosis. Massive cerioid rugose corals. Septa of two orders, more or less dilated and carinate in the dissepimentarium, thin in the tabularium. Major septa reaching only occasionally the center of the tabularium and the outer wall of the corallites from which they are separated by presepiments. Minor septa variably developed, commonly reduced to absent. Dissepimentarium composed of several rows of globose dissepiments often arranged in horizontal layers in its outer part and then inclined towards the axis of the corallites. Tabulae complete or incomplete, sometimes compound.
Discussion. The genus Wapitiphyllum has been introduced by McLean and Pedder (Citation1984) as a replacement name for Donia Soshkina, Citation1951; but it is based on three Canadian species: the genotype W. vallatum McLean and Pedder (Citation1984), W. facetum McLean and Pedder (Citation1984) and W. exiguum McLean and Pedder (Citation1984) which are only represented by one to three colonies. So, the variability of this Late Frasnian material from the Northwest Territories and the British Columbia is not very well known. Moreover, McLean and Pedder (Citation1984, p. 22) regarded Wapitiphyllum as a cerioid equivalent of the solitary genus Mictophyllum Lang and Smith, Citation1939, whose type species is M. nobile Lang and Smith, Citation1939, from the Late Frasnian of the Northwest Territories. However, this taxon revised by McLean (Citation2007, p. 39) is characterized by small inclined dissepiments and by very slender septa which are nearly continuous from the outer wall to the axis of the corallum. These features do not occur in the three species of Wapitiphyllum described by McLean and Pedder (Citation1984). Indeed, they show dissepiments mostly arranged in horizontal layers and septa which are discontinuous at the periphery and more or less dilated in the inner part of the dissepimentarium. These characteristics are also present in Cystihexagonaria defecta Rohart, Citation1988, type species of the genus Cystihexagonaria Rohart, Citation1988.
Wapitiphyllum sp.
Figures –, I
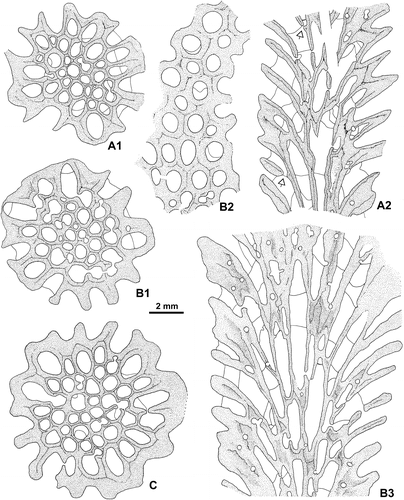
Figure 12. Wapitiphyllum sp. A-D. Transverse sections showing mainly complete major and minor septa (A, large corallite), minor septum reduced to a septal spine/ridge (C, open arrow) or divided into fragments (B). Note in B (lower right) unusual discontinuities in the wall. A, C–D acetate peel; B thin section, see also Figure I1; E. Longitudinal section (acetate peel). LPB 19390.
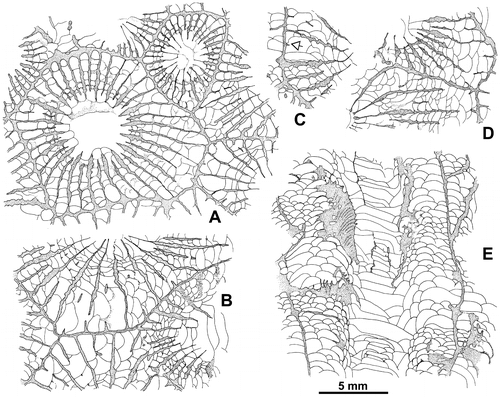
Figure 13. Wapitiphyllum sp. Part of a transverse section in a corallite. A. Outline of some knobbly carinate and discontinuous major and minor septa. B. Enlarged view of two septa showing trabeculae in transverse section. LPB 19390, thin section.
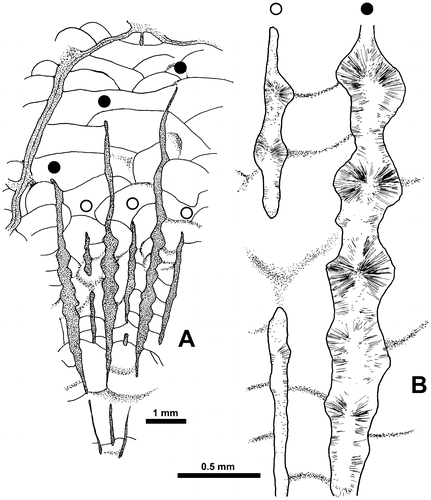
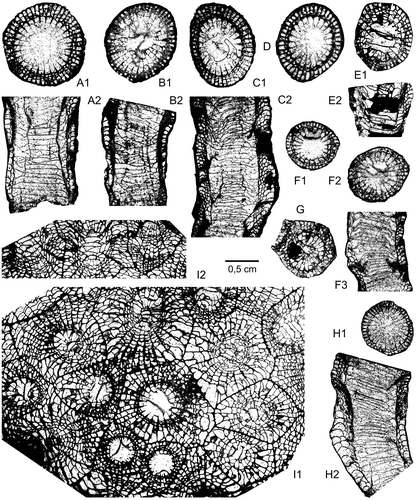
Figure 14. Frasnian rugose corals of the Kozan section. A–D. Disphyllum cf. rugosum (Wedekind, Citation1922); level PK66 D. A1–A2. LPB 19388, transverse (distal) and longitudinal sections, acetate peels. B1–B2. LPB 19387, transverse (distal) and longitudinal thin sections. C1–C2. LPB 19389, transverse (distal) and longitudinal sections, acetate peels. D. LPB 19376, transverse (distal) section, acetate peel. E–H. Disphyllum cf. curtum Hill, Citation1954; level PK66 A. E1–E2. LPB 19378, transverse and longitudinal thin sections. F1–F3. LPB 19381, transverse proximal (F1), distal (F2) and longitudinal sections, acetate peels. G. LPB 19380, transverse section, acetate peel. H1–H2. LPB 19383, transverse (proximal) and longitudinal sections, acetate peels. I1–I2. Wapitiphyllum sp.; level PK66 B. LPB 19390, transverse and oblique thin sections.
Material. 1 colony with 2 thin section and 2 acetate peels. Level PK66 B: LPB 19390.
Description. The only colony available has a height of 3.5 cm and an area of 5.5 × 5 cm. It is developed on an alveolitid. The calices are excavated and bordered by a flat platform. The walls between adjacent corallites are straight with a dark median line.
The septa are non-carinate or bear a few small spinose and knobbly carinae. They are dilated in the dissepimentarium and become thin in the tabularium or slightly beyond their entry into it (Figure ). In some corallites, the septa are typically spindle-shaped. At the periphery, they are occasionally slender or even discontinuous and divided into segments with some presepiments (Figure A). A few small lateral dissepiments are developed along the sides of the septa.
The major septa leave a more or less extensive open space in the center of the tabularium where they may be discontinuous; sometimes also, they are reaching the axis of the corallites. The minor septa are highly variable in length. Indeed, they are short, lacking, divided into fragments or more or less long, that is to say traversing all, nearly all or half the dissepimentarium. Some herringbone dissepiments are present.
The dissepimentarium consists of 3–6 rows of large globose dissepiments which are arranged in horizontal layers and are only inclined at the border of the tabularium. The tabulae are incomplete and intersecting laterally; their axial parts are rather often flat-topped.
There are 36–38 septa per corallite. The diameter of the corallites ranges from 8 to 15 mm whereas the width of the tabularium varies between 3.3 and 5.5 mm.
Microstructure. The microstructural data are provided by the study of standard thin sections and acetate peel (“LFP” not available).
The wall is thin and belongs to a three-layered type; the median layer appears as a continuous dark line in transverse section flanked on both sides by a stereoplasm which seems to be fibrous.
The trabeculate construction of the septa is clearly exposed in transverse section as well as in longitudinal/tangential one. In transverse section, especially in the dilated part of the major septa, an obvious relationship between lateral expansion of trabeculae and knobbly carinae is expressed (Figure ). In longitudinal section, the trabeculae appear contiguous and progressively inclined towards the tabularium in the inner dissepimentarium (Figure E); in the outer dissepimentarium, the trabeculae are only seen in oblique sections, but it seems that they are subparallel to the wall. Diameter of larger trabeculae ca. is 0.20-0.28 x 0.40 mm.
This microstructure is consistent with that described by Sorauf (Citation1967) in some species assigned to Hexagonaria Gürich, Citation1896; it is also visible on the figures or briefly mentioned in Wapitiphyllum by McLean and Pedder (Citation1984).
Discussion. In transverse section, the Turkish specimen shows much variation concerning the septal thickening and the length of the minor septa. There are some similarities between Wapitiphyllum sp. and W. defectum coming from the Middle Frasnian of the Boulonnais in France. However, the latter taxon is separated from the former by thinner septa which are more discontinuous at the periphery and by minor septa less developed. By its septa showing some fusiform dilation and not systematically interrupted by presepiments near the outer wall, the Turkish colony described herein is a transitional form between the genera Wapitiphyllum and Hexagonaria. It is assigned to Wapitiphyllum due to some peripheral discontinuity of the septa and the variable length of the minor septa. These two features are not characteristic of the diverse Frasnian species of the genus Hexagonaria investigated among others by Rohart (Citation1988), Coen-Aubert (Citation1994), Sorauf (Citation1998) and McLean (Citation2010).
Geographic and stratigraphic range. The form has only been found in the Frasnian of eastern Taurus in Turkey.
Tabulate corals (Y.P., F.T. & R.G.)
Subclass Tabulata Milne-Edwards and Haime, Citation1850
Order Favositida Wedekind, Citation1937
Family Pachyporidae Gerth, Citation1921
Genus Thamnopora Steininger, 1921
Type species. Thamnopora cervicornis (de Blainville, Citation1830)
Diagnosis. See emended diagnosis in Tourneur (Citation1986).
Thamnopora sp.
Figures –
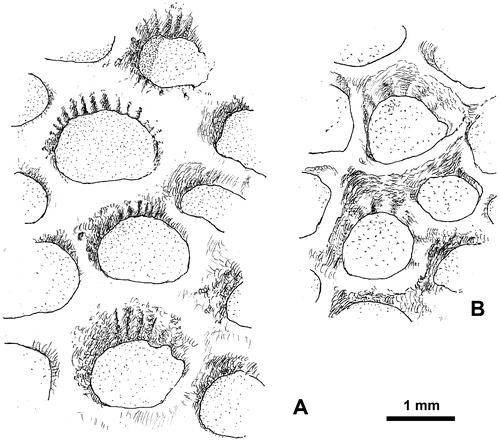
Figure 15. Thamnopora sp. View of exterior of branches and detailed morphology of the calices. A1–A2. LPB 19369. B. LPB 19373. C. LPB 19370. D. LPB 19371. E. LPB 19374. Enlargement of calices showing rounded ventral lips (A2), septal ridges (open arrow) and median tooth (B2, E), pseudopercula and septal ridges (D).
Material. The material consists of 9 short branch fragments, 4 plaster cast replica, 14 thin sections, 14 acetate peels, level PK66 A, catalogue number LPB 19366–19374.
Description. The branches are cylindrical or dichotomously coarsely branching (Figures B, C). The corallites opening roughly perpendicular or, more often, not very obliquely to the surface show rounded calices. In some specimens (or in some areas), the ventral outline of the corallites is locally curved; a kind of lip develops wearing in some cases a smooth median teeth (Figures B2, E).
The calicinal wall seems to be smooth at first sight. Nevertheless, in some cases, it bears very inconspicuous septal ridges preferentially developed on the upper “lip” of the calice. The ridges are recorded in five of the nine specimens (LPB 19369, 19370, 19371, 19373 and 19374; calices not exposed in LPB 19368 and 19372), but they are visible in a limited number of corallites, sometimes only in one of them. The ridges are narrow and their number reaches up to 18 on the half diameter (Figures B2, 16A) or less usually rather broad (ca. 7 on the half corallite diameter, Figure B). Exceptionally, the two kinds of ridges are recorded on the same branch.
Figure 16. Thamnopora sp. Calices showing narrow (A) and broad (B) septal ridges (explanations in text). A. LPB 19373 (see also Figure B2). B. LPB 19369.
In one specimen (LPB 19370), near the apex of the branch, the intercalicinal area shows a row of more or less transverse granules (5–6 along a section wall), probably emphasized by weathering.
Pseudopercula are recorded in two specimens; they are rare in LPB 19370, rather numerous and well preserved in LPB 19371 (Figure D2). There, the pseudopercula show concentric coarse wrinkles or more smooth structures whose center is occupied by a small elongated depression. Pseudopercula affect only a small portion of the corallum located in the middle part of the branch (Figure D2) and, according to Nothdurft and Webb (Citation2009, p. 143), these structures are interpreted “as a response to stress that may help protect the colony from invasion of unhealthy tissues by parasites or disease”. The presence of pseudopercula has already been mentioned in the genus Thamnopora by Hubmann and Messner (Citation2011) in the Eifelian of “Graz Palaeozoic”.
The branch diameters do not exceed 12 mm except in the area of branching; 8.5–9.5 mm is most common. Owing to the fragmentary specimens, the length is unknown (more than 30 mm). In the same way, it is difficult to assume the growth form of the colony, probably bushy like in the classical species (Th. reticulata de Blainville, Citation1830) of western Europe, easily broken into short fragments.
In transverse sections, the axial zone of the branch is surrounded by a crown of obliquely cut corallites (this feature corresponding to their gently curved orientation clearly visible in longitudinal sections) opening at the surface of the branch in somewhat oblique calices. The distal thickening of the walls is rather limited as shown by the tangential sections (Figure A2).
Figure 17. Thamnopora sp. Sections showing the structure of the corallum. A1–A2. Respectively transverse and longitudinal sections, note on A2 (arrows) the location of detailed Figures D and (top) and 20B2 (bottom). LPB 19366, thin sections Bb 1132 and Bb 1133. B1–B3. Transverse, tangential and longitudinal sections. LPB 19373, acetate peels. C. Transverse section. LPB 19374, acetate peel.
The axial zone occupies usually slightly more than half of the branch diameter (Figure A–B); nevertheless, in some cases, this area is more developed (Figure ). In the axial zone, the corallite lumina are subpolygonal to moderately rounded. In the center of the transverse sections, the corallites are rather unequal, with many young smaller ones (Figure B1, C), consequence of active budding in this part of the branch. The tangential section shows more equal corallites (Figure B2) (and this is consistent with the rather uniform dimensions of the calices on the surface of the branches), with a conspicuous widening of the diameter of the corallite and at the same time, of the diameter of the lumen (due to the rather limited thickening of the walls).
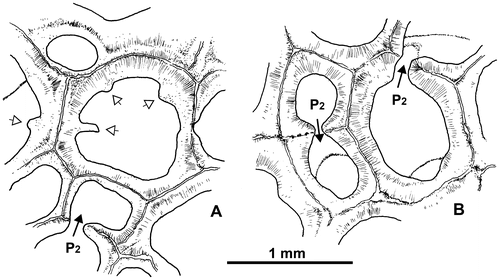
Mural pores belong to the wall pore variety (Powell & Scrutton, Citation1978,= P2 sensu Plusquellec, 1976); they are generally situated on the center of the faces, scarce on their edge (Figure B).The angle pores (=P1) are missing as usual in Thamnopora. Some pore plates are recorded.
Figure 18. Thamnopora sp. Transverse sections showing (A) unusual corallites bearing squamulae (or spines?) and (B) two kinds of pores P2 : midface wall pore (left) and side face wall pore (upper right). Respectively LPB 19367, thin section Bb 1130 and LPB 19366, thin section Bb 1131.
The presence of squamulae (or septal spines?) in the axial zone is very unusual and only recorded in one specimen (Figure ).
The tabulae are rather scarce and irregularly distributed, spaced from 0.3-0.4 mm up to 2.0 mm, even in rare cases 2.9 mm. They are flat, slightly concave or convex, rarely incomplete.
Microstructure. The microstructural data of the corallite wall are provided both by standard thin sections and by the so-called “LFP”. The wall consists of a narrow median plate flanked on each side by thick fibrous stereoplasm. In 30 μm thin sections, the median plate appears as a light-colored structure lined on both sides by a narrow dark line (Figure ) which in fact belongs to the very proximal part of the stereoplasm; this latter is organized in two layers. In some cases, in the peripheral zone observed in longitudinal section, the stereoplasm shows low-angled growth lamination.
Figure 19. Thamnopora sp. Microstructure of the wall. A. Transverse section, periphery of the axial zone showing the median plate flanked by the two layers of stereoplasm. LPB 19373, “LFP” thin section, crossed nicols. B. Slightly oblique section, peripheral zone, showing the “plaquettes” of the median plate. LPB 19373, “LFP” thin section, crossed nicols. C. Tangential section to the branch showing the trabecular structure of the inner layer of the stereoplasm of the “upper “corallite. LPB 19374, thin section Bb 1122. D. Transverse section, axial zone showing the light-colored “dark line” (= median plate) and its duplication on one of its sides. LPB 19374, thin section Bb 1123.
The “LFP” allow more accurate observation detailed as follows. In the peripheral zone of the corallum, the median plate is built of flattened and radially arranged units, the so-called “plaquettes” of Tourneur (Citation1986) (Figure B). In the axial zone, this setting is less obvious and the “plaquettes” could be longer than wide (such as 20 × 12, 17 × 12 μm or more or less square 11 × 11, 10 × 11 μm). The width of the median plate is of ca. 10–12 μm. In the margin of the corallum, a water jet bundle of fibers replaces the “plaquettes”. This evolution of the median plate has already been described in tabulate corals, for example, in Praemichelinia (Lafuste & Plusquellec, Citation1980). The stereoplasm consists of an inner layer of small fibers (diameter ca. 2–3 μm) and an outer one of large fibers or fibroids (diameter ca. 10–15 μm, even up to 25 μm) (Figure ). These microstructural features are diagnostic of the genus Thamnopora Steininger, 1831 and clearly differ from Lecomptopora Tourneur, Citation1986.
In some cases, the wall exhibits a generally short duplication of the median plate which develops within the fibrous stereoplasm on one side of the median plate in the middle part of the wall or in a corner (Figure ). This very unusual structure (probably described for the first time) seems teratologic.
Another unusual feature is the replacement of the radial fibers of the inner layer by trabeculae (Figure ); it only occurs in a small area of the peripheral zone and this feature is probably linked with the calicinal ridges. In our opinion, this structure only seen in two specimens (LPB 19373 and 19374) does not question the assignment to the genus Thamnopora.
Increase. In tabulate corals, one of the more usual modes of reproduction belongs to the lateral type (or lateral increase, Oliver, Citation1968; Stel, Citation1978) also called intracalicular-visceral budding (Nowinski, Citation1976).
As far as we know, the only paper dealing with the increase in Thamnopora is that of Nowinski (Citation1976, Th. boloniensis from the Holy Cross Mountains, Poland). Using serial transverse sections, and taking into account the two layers of fibers of the stereoplasm, he described during the “preblastic stage” the development of a bulblike invagination becoming soon the incipient visceral cavity of a new individual. This stage is followed by the growth of a neotheca (“hystero-nepionic stage”) that separates the offset from the parent corallite; after complete separation, the young corallite is quadrangular in transverse section (“hystero-neanic stage”). Nowinski (Citation1976) assigns the increase to the intracalicular-intravisceral type.
The main features of the increase in Thamnopora as identified by Nowinski (Citation1976) are very similar to those described – for example – in Saouraepora gigantea (Plusquellec, Citation1991), except that in the latter, the “cella” (Plusquellec, 1976, p. 200; = polygonal bulge, Oliver, Citation1966, p. 449; = bulblike invagination, Nowinski, Citation1976, p. 37) is generally two-sided and subsequently, the offset is triangular (three-sided) in its youngest stage.
When lateral increase is seen in longitudinal section, the corresponding figure of the serial transverse sections clearly shows: (1) the basal pore, sometimes obstructed by a pore plate and (2) usually a change in the direction of the wall of the parent corallite at the basal pore level (Ohiopora cylindrica, Lafuste & Plusquellec, Citation1987, Figure 4; Parastriatopora cantabrica, Tourneur & Fernández-Martínez, Citation1991, Figure 11A–C; Saouraepora gigantea, Plusquellec, Tourneur, & Lafuste, Citation1993, Figure 8; Argentinella argentina, Fernández-Martínez, Plusquellec, & Tourneur, Citation2002, Figure 5; Praemichelinia steli, Fernández-Martínez & Plusquellec, Citation2006, Figure 10). It will be noticed that in these examples (as well as some unpublished ones), there are no significant morphological or structural changes in the wall of the parent corallite in the basal pore area, such as thickening, thinning down or something else.
The specimens of Thamnopora from Kozan have not been studied by serial sections, but the transverse and longitudinal thin sections and acetate peels undoubtedly show that the increase roughly belongs to the lateral type. Nevertheless, especially in longitudinal sections, numerous patterns of lateral increase exhibit an unusual feature that will be described first from the angle of longitudinal sections and secondly from transverse sections.
In longitudinal sections, the standard pattern of lateral increase is rare and somewhat doubtful when only seen on acetate peels. In some cases assigned to the standard type (and studied in thin section), the wall of the parent corallite shows on its “fertile” side a very weak thickening located below the basal pore (and below the pore plate) and immediately followed by a strong thinning displaying a concave outline to the apical part of the wall belonging now to the offset (Figure A).
Figure 20. Thamnopora sp. Lateral increase in longitudinal sections (A–C) and in transverse sections (E–F). Detailed explanations in text; asterisk as basal low wall. A. Standard type, basal pore (P0) closed by a pore plate (pp), LPB 19367, thin section Bb 1129. B1, B2. Respectively overview of lateral increase with reduced basal low wall and enlargement of the basal pore area, LPB 19366, thin section Bb 1133. C1, C2. Lateral view with typical basal low wall, black arrow as apical cul-de-sac, pore plate lacking, LPB 19367, thin section Bb 1129. D. typical basal low wall, but apical cul-de-sac not developed, LPB 19366, thin section Bb 1133. E. Section showing the parent corallite and its “cella,” boundary marked by re-entrant angles (black arrows), LPB 19374, thin section Bb 1123. F. Same caption, note the rounded section of the apical cul-de-sac bounded on its parent corallite side by the bar-like distal section of the basal low wall (open arrow), LPB 19374, thin section Bb 1123. G. Corallite parent and offset, section in the neotheca (grey arrow), LPB 19371, thin section Bb 1125.
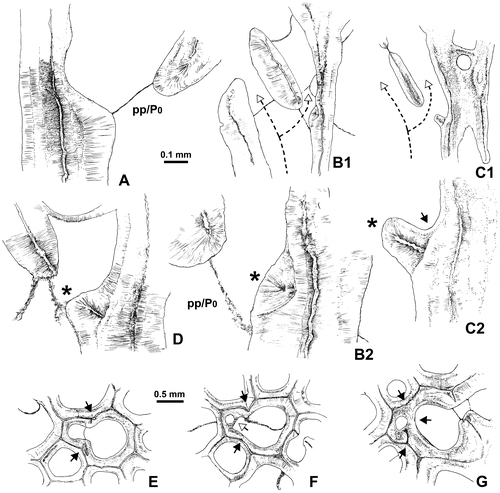
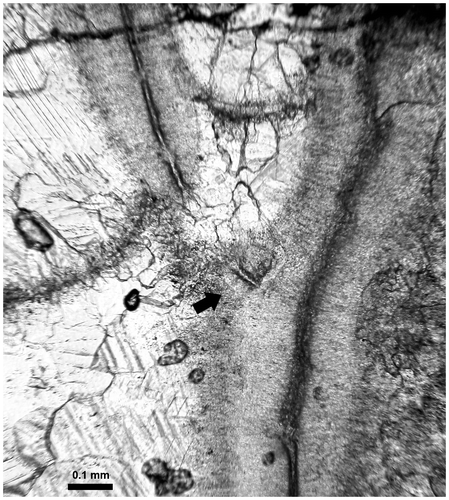
The other patterns of lateral increase show, on the “fertile” side of the wall of the parent corallite and forming the proximal margin of the basal pore, a small protuberance whose microstructure is similar to that of the corallite wall (Figures –D, ). Resting in unconformity on the outer layer of fibers of the stereoplasm of the wall of the parent corallite, this protuberance shows a wide light median plane flanked on each sides by a fibrous stereoplasm becoming fan-like on top. This structure has never been described and is called basal low wall. In some cases, the dipping of the distal side of this basal low wall is strong enough to create an apical cul-de-sac (Figure C2).
Figure 21. Thamnopora sp. Longitudinal section of the area of increase (see also Figure D and its caption), LPB 19366, thin section Bb 1133.
The basal low wall could be reduced to a low bundle of fibers devoid of true median plane, but clearly separated from the stereoplasm of the wall of the parent corallite by a line (in fact a plane) of unconformity (Figure B).
The comparison of Figure A (standard type) and Figure B (reduced basal low wall) shows that before the development of the reduced basal low wall, the morphology of the wall of the parent corallite in the basal pore area is very similar and exhibits in both cases a strong thinning above the basal pore and/or the pore plate. Thus, the reduced basal low wall clearly appears as secondary and/or optional. Moreover, it is noteworthy that the wall bearing typical basal low wall such as those illustrated (Figures C–D) does not show important thickening variations. This feature could be only a characteristic of the Turkish form because in Thamnopora sp. from the Givetian of Dollendorf (Eifel), the basal low wall is linked with a rather strong thickening of the parent corallite wall (unpublished data and sections not illustrated here).
In transverse sections, lateral increase at its preblastic stage is characterized by the presence of two opposite reentrant angles in the path of the median line of the wall, linked with a bulb-like extrusion of the main visceral chamber (Figures E–G). These features allow identifying the “cella” which is generally quadrangular in our material. The contracted connection between the two parts of the visceral chamber corresponds to the basal pore which is often closed by a pore plate.
When the transverse section is situated a little bit below the basal pore and when the basal low wall gives rise to a well-marked apical cul-de-sac, the transverse section of this latter structure appears as a blind lumen (Figure F). Remark: the pore plate cannot be involved in such a section because it is situated above it.
In transverse section crossing the offset barely above the basal pore, the neotheca appears between the two visceral chambers and, in some cases, its median line, outgrowing from the reentrant angle, is asymmetrically developed (Figure G). It can be noticed in some corallites, on both sides of the basal pore (i.e. very early during the increase), the presence of very short segments of the median line belonging to the neotheca and outgrowing as usual from the reentrant angles (Figure E).
The lateral increase with basal low wall is frequent in the branches of the Thamnopora from Kozan and thus it was interesting to do a research for the presence of this structure in other species.
A preliminary study of specimens from different localities and levels has been carried out and the structure has been recorded in Thamnopora sp. from Dollendorf (Eifel), Lower Givetian (in some cases with an obvious apical cul-de-sac), and in Th. cervicornis from Les Limites quarry, Ave-et-Auffe (Belgium), Trois-Fontaines Formation, Lower Givetian. In this species, the standard lateral increase coexists with the other one and the development of the apical cul-de-sac is variable. On the other hand, the basal low wall seems to be lacking in a large species of Thamnopora from Ria de Perán (Asturias), Candás Formation, Upper Givetian to Lower Frasnian.
We have not extended our study to all the genera known to have an asexual reproduction belonging to the basic lateral type and unfortunately, the illustrations are mainly unusable for the purpose.
Nevertheless, Swann (Citation1947, pl. 3, Figure 4) provides a very nice photograph of the increase in Favosites alpenensis calveri (Traverse Group, Middle Devonian of Michigan) showing in longitudinal section, below the basal pore, a protuberance that indisputably looks like the basal low wall of Thamnopora. This feature allows three remarks. First, following Sokolov (Citation1962), Favosites alpenensis calveri is assigned to Pachyfavosites (see legend of pl. 2, Figure 2). Secondly, Tourneur (Citation1985, p. 285), then Lafuste and Tourneur (Citation1991) pointed out that “in its axial part, the skeleton of Thamnopora is analogous to that of Pachyfavosites” and suggested that the systematic position of the genus Pachyfavosites within the Favositidae could be discussed. Thirdly, the occurrence of a basal pore with basal low wall in Pachyfavosites provides a new interesting argument concerning the issue raised by Lafuste and Tourneur (Citation1991). It indirectly proves the interest of detailed study of the microstructure and that, even submitted to some diagenesis, the main characteristics of the microstructure remain usable for systematics.
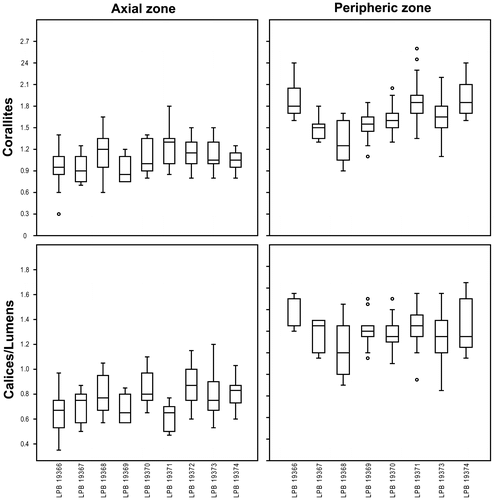
Biometric data – Overall dimensions of Thamnopora sp. are listed in Table ; in addition, some box plots (Figure ) are provided for the corallites and lumen diameters.
Table 1. Thamnopora sp.: biometric data (measurements in mm, corallite and lumen diameter for adults only).
Figure 22. Box plots for the corallites (top) and calices-lumens (bottom) diameters of Thamnopora sp. measured respectively in the axial (left) and peripheric (right) zones (measurements in mm).
Discussion –Systematics of the genus Thamnopora at the specific level is in great need of revision since Lecompte revitalized the genus in the 30s of last century by emphasizing the generic characters of this type of branching tabulate coral in comparison with the Silurian Pachypora Lindström, Citation1873 (mostly for reasons of microstructure), and with the “ordinary” Favositids. Even in the now classical paper of Lecompte (Citation1939), the distinction between the discrete species of Thamnopora is not obvious, when considering either biometric or morphologic characters, as well as their stratigraphical distribution, this latter being often based on isolated and somewhat uncertain occurrences. The collections of Goldfuss, mostly from the Eifel, which were used to define many species (Th. cervicornis, Th. reticulata and so on), are rather well known since the careful revision of Lecompte (Citation1936) and the investigations of Birenheide (Citation1985), but more ancient taxa (Th. cristata [Blumenbach, Citation1803] and Th. polyforata [von Schlotheim, Citation1820] a. o.) are in need of revision, as well as the species defined by Michelin (Citation1840–1847), Milne-Edwards & Haime (Citation1851), and Gosselet (Citation1877), mainly from western Europe. Since 50 years, a huge quantity of species (certainly more than 100) was created by Russian and Chinese authors, often with bad descriptions and poor illustrations and therefore very difficult to use! Thus, many studies are still to do, for example, to understand the biometric variation of large populations. Biometry is not easy, dealing with the relationships between corallites and lumen diameters, and their relative expansion towards the periphery of branches, in the same time that walls are thickening. Other morphological characters, such as occurrence of spines or more frequently of squamulae, distributions of tabulae and so on, are not well understood; whether they depend of ecology or are of systematic interest, remains questionable.
In this context, the rather small population from the Kozan section, although rather well preserved, is unfortunately not easy to identify at specific level. The biometric data are rather close to those of species collected in the Frasnian of Belgium, northern France and western Germany. But, it is quite impossible to assign an exact specific name to the Turkish material. So, we propose herein to let these specimens in open nomenclature, underlying their similarities with rather well-known species from Western countries.
A set of Middle Devonian species, around the classical Th. reticulata (de Blainville, Citation1830) and Th. cervicornis (de Blainville, Citation1830), well known from western part of Germany and south of Belgium, is characterized by smaller corallites, rather numerous in the axial zone of the branches, and slightly expanding distally.
Another group from the same area, including Th. polyforata (von Schlotheim, Citation1820) – that has been shown to be synonymous of the more frequently used name of Th. dubia (de Blainville, Citation1830) – was identified in the vicinity of the Givetian/Frasnian boundary. It is characterized by larger, not so numerous corallites in the somewhat slender branches and opening in slightly oblique calices. Moreover, it should be noticed that in the specimens of Th. dubia illustrated by Birenheide (Citation1985, pl. 19, Figure 1A), the lower lip of the calices exhibit a very tiny median tooth.
In the Frasnian strata of Belgium, at least three different species can be easily distinguished. The first one, with rather thick branches and larger corallites, was traditionally described as Th. boloniensis (Gosselet, Citation1877) (name created for material of the Boulonnais area), but the older name Th. cristata (Blumenbach, Citation1803) was proposed as a valid synonym. It was originally used for specimens from the Frasnian reefal limestones (Weyer, personal communication, 2013) of Iberg-Winterberg (Harz), but the latter population is not so well known as the Franco-Belgian one and these species globally need a revision with restudy of type materials.
Another species, Th. micropora Lecompte, Citation1939, with smaller corallites, is known from Late Frasnian and was collected in the red reefal limestones and in the surrounding shales. Finally, thin branches with rather large corallites, opening in oblique calices, are observed in the somewhat younger red limestones of the Rance area (western part of South Belgium), well known as “decorative marbles”; they are until now unnamed. From all these populations, in great need of revision, the Turkish specimens are closer to the Middle Frasnian group Th. cristata/Th. boloniensis.
The material formerly described by several authors as “Thamnopora” from different regions of Turkey – Th. vermicularis from Yahyali (Flügel, Citation1955) and Th. reticulata reticulata from Feke (Flügel & Flügel, Citation1961) – is not sufficiently described, lack illustrations and thus cannot be compared with our material. Hubmann (Citation1992) identified specimens from Feke as Th. reticulata, assigning to this species a wide geographical distribution and a rather long stratigraphic life (“Middle/Late Devonian” …). Our studies of Belgian material allowed the identification of this species in Upper Eifelian and Lower Givetian strata only. The rather small figures of Hubmann (Citation1992, pl.2, Figure 3–4) show specimens closely related to the material herein described.
In conclusion, we assigned our specimens without any doubt to the genus Thamnopora, close to the Frasnian group of Th. polyforata/Th. boloniensis. In addition, the presence of very unusual features such as calicinal ridges and trabeculae in the peripheral stereoplasm are to be underlined although their systematic value needs further investigations.
Family Alveolitidae Duncan, Citation1872
Genus Alveolites Lamarck, Citation1801
Type species. Alveolites suborbicularis Lamarck, Citation1801
Diagnosis. See Hill (Citation1981, p. F591).
Alveolites sp.
Material. 1 specimen, 2 thin sections, 3 acetate peels. Level PK66 B, catalogue number LPB 19365.
Description. The morphology of this small and incomplete specimen, in contact with the colony of Wapitiphyllum sp. described above, is unknown (however corallum not fingerlike). Calices not seen.
The transverse section shows crescentic corallites. The presence of septal elements is difficult to establish (boundary between skeleton and cement not sharp); nevertheless, some corallites show a solitary spine in the middle of the lower calicinal side. The width (= large diameter) of the corallites is ca. 0.7-0.8 mm, the width of the calices ca. 0.45-0.55 mm, the small diameter of corallites ca. 0.45 mm, the small diameter of calices ca. .25–.28 mm and the thickness of the wall ca. 0.15 mm.
In longitudinal sections, the thin, complete and slightly concave tabulae seem closely spaced (ca. 0.2 mm). Scarce tangential sections of mural pores, probably P2, can be seen.
Discussion. Owing to the few biometric data available and the rather poor preservation, the species is left in open nomenclature, but it is interesting to point out the presence – in the Gümüşali Fm. – of the genus Alveolites which is very common in the coral-bearing facies of Givetian and especially Frasnian. It could be noticed that some species of Alveolites, ranging from Upper Eifelian to Frasnian, have been mentioned in the same tectono-stratigraphic unit (eastern Taurus, Feke area) by Hubmann (Citation1992).
Palaeobiogeography
The more or less strong affinities of the Frasnian species of the Rugosa and Tabulata identified in the eastern Taurus (Kozan section) with those known elsewhere are summarized on a palaeobiogeographic map (Figure ; late Devonian base map after Scotese and Golonka [Citation1992] modified). The paleontological data concerning Rugosa (Disphyllum and Wapitiphyllum) are based on Coen-Aubert and Plusquellec (this paper) and those concerning Tabulata (Thamnopora) on this paper, Tourneur (Citation1985), Tourneur et al. (Citation1989) and Zapalski (Citation2012).
Figure 23. Palaeobiogeographic map of the Frasnian (ancient landmasses in light grey) showing the location of the areas considered in the present paper. Turkey in dark grey. 1: North Devon (Great Britain). 2: Boulonnais (France). 3: Rhenish Mountains (Germany) and Ardenne (Belgium). 4: Upper Harz Mountains (Germany). 5: Holy Cross Mountains (Poland). 6: Moravia (Slovakia). 7: Central Iran (Iran). 8: District of Mackenzie (Canada). 9: North Vietnam (Vietnam). 10: Canning Basin (Australia). 11: Russian platform and Oural (Russia). 12: Kuznetsk Basin (Russia). Genera and species taken into account- Disphyllum rugosum: Ardenne (Middle Frasnian), Rhenish Mountains/Bergisches Land (Middle Frasnian). D. curtum: Canning Basin (Frasnian). D. grabaui : Ardenne (Lower Frasnian). Disphyllum sp. 1 sensu Rohart (Citation2000, p. 68): Central Iran (Middle Frasnian). Wapitiphyllum defectum: Boulonnais (Middle Frasnian). Thamnopora polyforata/dubia: Boulonnais (Givetian or lower to Middle Frasnian), Ardenne (Givetian to Middle Frasnian), Rhenish Mountains (Givetian to Frasnian), North Vietnam (Givetian-Frasnian), Russian platform and Oural (Lower to Middle Frasnian). Th. near Th. polyforata: District of Mackenzie (probably lower or Middle Frasnian). Th. boloniensis /cristata: Boulonnais (Middle Frasnian), Ardenne (Givetian to Middle Frasnian), Upper Harz Mountains (Frasnian), North Devon (Givetian to Frasnian), Moravia (predominantly Lower Frasnian), Canning Basin (Upper Givetian to Lower Frasnian), North Vietnam (Givetian-Frasnian), Kuznetsk Basin (Frasnian). Th. e.g. boloniensis: Holy Cross Mountains (Givetian to Lower Frasnian).
The map clearly shows that the Disphyllum and Thamnopora are mainly located between 0° and 30° South latitudes, i.e. in warm to rather warm epicontinental seas, on the margin of Laurussia and Gondwana, and in the Eastern area, the latter is probably present in the vicinity of South China (North Vietnam). A strange exception is the spot of Th. boloniensis in the Kuznetsk Basin (Siberia). However, May (Citation1995), dealing with Givetian Thamnopora, points out close palaeobiogeographical relations between Asturias and Siberia.
Moreover, it can be noticed that Thamnopora e.g. polyforata/boloniensis seems to be missing in Spain and North Africa during Frasnian times while, at least, Th. boloniensis is recorded in the Givetian of Asturias (May, Citation1995). In addition, the genus Disphyllum seems to be unknown in the Lower to Middle Frasnian of these regions whereas some species are described from the Givetian of Asturias; nevertheless, Disphyllum appears again – as well as Thamnopora sp.– in the Upper Frasnian of the Piñeres Formation in Asturias (Fernández, Fernández-Martínez, Mendez-Bedia, Rodríguez, & Soto, Citation1995). Thus, it appears that the distribution of the species (and probably the genera) taken into account in this paper is more confined during the Lower to Middle Frasnian period than during the Givetian, especially on the northwestern margin of the Gondwana.
Conclusions
Despite the rather few specimens of rugose and tabulate corals collected in the Kozan section, their study provides some interesting new data dealing with various aspect of paleontology such as systematics, biostratigraphy, paleobiogeography, structure and microstructure.
The Kozan section has yielded an assemblage of cosmopolitan genera: Disphyllum – Wapitiphyllum – Thamnopora – Alveolites which is very common in the Givetian to Frasnian limestones, especially in the southeastern Laurussia (Boulonnais, Ardenne). Though the Rugosa and the Tabulata remain in open nomenclature, it is interesting to outline that the specimens of Disphyllum are referred to species known in the Frasnian (base of the Middle Frasnian and Frasnian sensu lato) whereas the specimens of Thamnopora show affinities with species mainly known in the Upper Givetian and Lower to Middle Frasnian. In addition, the genus Wapitiphyllum although recorded in the Givetian, is mainly known in the Frasnian. Thus, the probably Early Frasnian age assigned to the Gümüşali Formation exposed in the Kozan section by Gourvennec (Citation2006) cannot be disputed by the data provided by the corals.
The genera Dispyhyllum, Alveolites and Thamnopora have already been mentioned since the Middle Devonian in the eastern Taurus (Flügel and Flügel, Citation1961; Hubmann, Citation1992), but the genus Wapitiphyllum is for the first time reported from the Frasnian of this area.
Within the Gümüşali Formation exposed in the Kozan section, the coral fauna shows a trend to be more diversified and more abundant towards the top and it can be noticed that the level PK66 C – devoid of corals – is also the one in which the brachiopods (Gourvennec, Citation2006) are both more numerous and diversified.
The coral fauna of the Kozan section shows affinities with species known both in the south Laurussia (Disphyllum rugosum, Wapitiphyllum defectum) and in the northeast Gondwana (D. curtum). Thus, these new data reinforce – as noticed by Gourvennec (Citation2006) – “the faunal evidence for a narrow oceanic separation between North Gondwana and Euramerica” (=Laurussia) during the Frasnian, and illustrate the possibility of free movement of the faunas between Turkey and Ardenne/Rhenish Mountains area.
The septal insertion of the genus Disphyllum has been studied and the protosepta identified in some cases. It will be noticed that such interpretations are scarce in papers dealing with the genus.
Thanks to the use of “LFP” thin sections, new data on the microstructure of the wall (cf. epithecate wall) and dissepiments (lamellar microstructure) of the genus Disphyllum are provided.
In the calices of Thamnopora, unusual morphological features (septal ridges, median teeth, pseudopercula) are recorded; microstructural feature, i.e. the replacement of the radial fibers of the inner layer by trabeculae is described for the first time; and a new pattern of lateral increase characterized by the development of a “basal low wall” and an “apical cul-de-sac” is amply illustrated.
Acknowledgments
We are grateful to the anonymous reviewers for their constructive remarks. O.M. thanks the Turkish Petroleum consortium (TPAO) for providing all the field assistance and is grateful to Dr H. Kozlu for numerous and helpful discussions during fieldwork. Thanks are due to Pr E. Poty for sending to one of us (Y.P.) specimens of Cyathaxonia cornu and to Drs B. Mistiaen, D. Weyer and M. Zapalski for their help concerning stratigraphy and references. The photographs of rugose corals were prepared by W. Miseur. This paper is a contribution to the IGCP 596 on Mid-Palaeozoic Climate and Biodiversity.
References
- Birenheide, R. (1985). Chaetetida und tabulate Korallen. Leitfossilien, 3, 1–249.
- de Blainville, H. (1830). Zoophytes. Dictionnaire des Sciences Naturelles, 60, 1–546. Paris.
- Blumenbach, J. F. (1803). Specimen Archaeologiae telluris terrarumque inprimis Hannoveranarum.1–28, Goettingae.
- Boulvain, F., & Coen-Aubert, M. (2006). A fourth level of Frasnian carbonate mounds along the south side of the Dinant Synclinorium (Belgium). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 76, 31–51.
- Brownlaw, R. L. S., & Jell, J. S. (2008). Middle and upper Devonian rugose corals from the Canning Basin, Western Australia. Memoir of the Association of Australasian Palaeontologists, 35, 1–126.
- Bulvanker, E. Z. (1941). Rugosa glavnogo devonskogo polya [Rugosa of the main Devonian field]. In M. A. Batalina & al. (Ed.): Fauna glavnogo devonskogo polya, tom 1 [Fauna of the main Devonian field], (pp. 133–137). Paleontologicheskiy Institut, Akademiya Nauk SSSR, Izdatel’stvo Nauka, Moskow and Leningrad.
- Coen-Aubert, M. (1989). Les Rugueux dévoniens du sondage de Nieuwkerke (extrémité occidentale du Synclinorium de Namur, Belgique). Annales de la Société Géologique du Nord, 108, 100–102.
- Coen-Aubert, M. (1994). Stratigraphie et systématique des Rugueux de la partie moyenne du Frasnien de Frasnes-lez-Couvin (Belgique). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 64, 21–56.
- Coen-Aubert, M. (2008). Fasciculate Disphyllids (Rugosa) from the Early Givetian Trois-Fontaines Formation in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 78, 31–50.
- Coen-Aubert, M. (2009). Fasciculate rugose corals across the Early-Middle Frasnian boundary in Belgium. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 79, 55–86.
- Dean, W. T. (1967). The correlation and trilobite fauna of the Bedinan Formation (Ordovician) in southeastern Turkey. Bulletin of the British Museum (Natural History) Geology, 15, 81–123.
- Dean, W. T. (1983). Additional faunal data for the Bedinan Formation (Ordovician) of south-eastern Turkey. Special Paper in Palaeontology, 30, 89–105.
- Dean, W. T. (2005). Trilobites from the Çal Tepe Formation (Cambrian), near Seydişehir, Central Taurides, southwestern Turkey. Turkish Journal of Earth Sciences, 13, 1–71.
- Dean, W. T., & Monod, O. (1970). The Lower Palaeozoic stratigraphy and faunas of the Taurus Mountains near Beyşehir, Turkey. I. Stratigraphy. Bulletin of the British Museum (Natural History). Geology, 19, 411–426.
- Dean, W. T., Monod, O. Ghienne, J.-F. Kozlu, H., & Günay, Y. (2003). TPAO report 4461, 54 p. ( unpublished).
- Duncan, P. M. (1872). Third report on the British fossil corals. Report of the 41st Meeting of the British Association for the Advancement of Science, Edinburgh (1871), 116–137.
- Fernández, L. P. Fernández-Martínez, E. Mendez-Bedia, I. Rodríguez, S., & Soto, F. (1995). Field trip A. Devonian and Carboniferous reefal facies from the Cantabrian Zone (NW Spain). In A. Perejon (Ed.), VII International Symposium on Fossil Cnidaria and Porifera (pp. 1–76). Salamanca: Graficas Varona.
- Fernández-Martínez, E., & Plusquellec, Y. (2006). A new species of Praemichelinia (Tabulata) from the Emsian (Lower Devonian) of Colle (Cantabrian Mountains, NW Spain) and the occurrence of solenia-like pores in the genus. Revista Española de Paleontologia, 21, 43–59.
- Fernández-Martínez, E., Plusquellec, Y., & Tourneur, F. (2002). Revisión de Favosites argentina Thomas, 1905, especie tipo de Argentinella nov. gen. Coral tabulado del Devónico inferior de Argentina. Revista Española de Paleontologia, 17, 101–116.
- Flügel, H. (1955). Zur Paläontologie des anatolischen Paläozoikums. II. Mitteldevonfauna von Yahyalí (NO Ala Dağ, Taurus). Neues Jahrbuch für Geologie und Paläontologie, Abh, 101, 267–282.
- Flügel, E., & Flügel, H. (1961). Stromatoporen und Korallen aus dem Mittel-Devon von Feke (Anti-Taurus). Senckenbergiana lethaea, 42, 377–409.
- de Fromentel, E. (1861). Introduction à l’étude des polypiers fossiles. Paris: Savy. 1–357.
- Gerth, H. (1921). Die Anthozoën der Dyas von Timor. Paläontologie von Timor, 9(16), 65–147.
- Ghienne, J.-F., Monod, O., Kozlu, H., & Dean, W. T. (2010). Cambrian-Ordovician depositional sequences in the Middle East: A perspective from Turkey. Earth-Science Reviews, 101, 101–146.
- Goldfuss, A. (1826). Petrefacta Germaniae, t.1, Düsseldorf: Arnz & Comp. 1–76
- Gosselet, J. (1877). Le calcaire dévonien supérieur dans le N.-E. de l’arrondissement d’Avesnes. Annales de la Société Géologique du Nord, 4, 238–272.
- Gourvennec, R. (2006). Upper Devonian Brachiopods from Eastern Taurus (Turkey). Geologia Croatica, 59(1), 1–17.
- Gürich, G. (1896). Das Palaeozoicum im Polnischen Mittelgebirge. Verhandlungen der Russisch-kaiserlichen Mineralogischen Gesellschaft zu St. Petersburg, Series 2, 32, 1–539.
- Harris, W. J. (1924). Victorian graptolites (new series), Part 1. Proceedings of the Royal Society of Victoria, 36, 92–106.
- He, Y. X. (1978). Subclass Rugosa. In Chengdu Institute of Geology and Mineral Resources (Ed.), Atlas of fossils of Southwest China. Sichuan volume. Part I. From Sinian to Devonian (pp. 98–178). Beijing: Geological Publishing House.
- Hill, D. (1939). The Devonian rugose corals of Lilydale and Loyola, Victoria. Proceedings of the Royal Society of Victoria, new series, 51, 219–256.
- Hill, D. (1954). Coral faunas from the Silurian of New South Wales and the Devonian of Western Australia. Bulletin, Bureau of Mineral Ressources, Geology and Geophysics, 23, 1–51.
- Hill, D. (1981). Part F, Coelenterata, Supplement 1, Rugosa and Tabulata, 2 vols. In C. Teichert (Ed.), Treatise on invertebrate paleontology (p. 1–762). The Geological Society of America and The University of Kansas, Boulder, CO and Lawrence, KS.
- Hill, D., & Jell, J. S. (1970). Devonian corals from the Canning Basin Western Australia. Geological Survey of Western Australia, Bulletin, 121, 1–158.
- Hinde, G. J. (1890). Notes on the palaeontology of Western Australia. 2. Corals and Polyzoa. The Geological Magazine, new series 3, 7, 194–204.
- Hubmann, B. (1992). Die Korallenfauna aus dem Devon von Feke (Antitaurus, SE-Türkei) I. Tabulata. Mitteillungen der Österreichischen geologischen Gesellschaft, 84, 355–372.
- Hubmann, B., & Messner, F. (2011). Pseudopercula in Thamnopora orthostachys; Plabutsch Formation (Eifelian), Graz Palaeozoic. Kölner Forum für Geologie und Paläontologie, 19, 55–56 ( 11th Symposium on Fossil Cnidaria and Porifera Liège, August 19-29, Abstracts).
- Kozlu, H., Sachanski, V., & Göncüoglu, M. C. (2012). Silurian. In M. C. Göncüoglu & N. Bozdogan (Eds.), Guide Book of the Palaeozoic in the Eastern Taurides (pp. 61–78). Ankara: Turkish Association of Petroleum Geologists. Spec. Pub. 7.
- Lafuste, J. (1970). Lames ultra-minces à faces polies. Procédé et application à la microstructure des Madréporaires fossiles. Comptes rendus Académie des Sciences, Paris, 270, 679–681.
- Lafuste, J., & Plusquellec, Y. (1987). Structure et microstructure de Favosites cylindrica Michelin 1847, espèce-type de Ohiopora n. gen. (Tabulata, Dévonien). Canadian Journal of Earth Sciences, 24, 1465–1477.
- Lafuste, J., & Tourneur, F. (1991). Biocristaux et éléments foncés de la muraille chez Thamnopora Steininger, 1831 (Tabulata, Dévonien). Annales de Paléontologie, 77, 3–20.
- Lafuste, J., & Plusquellec, Y. (1980). Les polypiers – Tabulata. In Y. Plusquellec (Ed.), Les Schistes et calcaires de l’Armorique (Dévonien inférieur. Massif Armoricain). Mémoires de la Société géologique et minéralogique de Bretagne, 23, 143–173.
- Lamarck J. B. (1801). Systême des animaux sans vertèbres. Paris, France, Libraire Deterville, viii+1–432.
- Lang, W. D., & Smith, S. (1934). Ludwig’s “Corallen aus Paläolitischen Formationen” and the genotype of Disphyllum de Fromentel. The Annals and Magazine of Natural History, (10), 13, 78–81.
- Lang, W. D., & Smith, S. (1939). Some new generic names for Palaeozoic corals. The Annals and Magazine of Natural History, (11) 3, 152–156.
- Lecompte, M. (1936). Révision des Tabulés dévoniens décrits par Goldfuss. Mémoires du Musée Royal d’Histoire Naturelle de Belgique, 75, 1–111.
- Lecompte, M. (1939). Les Tabulés du Dévonien moyen et supérieur du bord sud du Bassin de Dinant. Mémoires du Musée Royal d’Histoire Naturelle de Belgique, 90, 1–213.
- Lindström, G. (1873). Nagra anteckningar om Anthozoa tabulata. Öfversigt af Kongl. Vetenskaps-Akademiens Föfhandlingar, 4, 3–20.
- McLean, R. A., & Pedder, A. E. H. (1984). Frasnian rugose corals of Western Canada. Part 1: Chonophyllidae and Kyphophyllidae. Palaeontographica A, 185, 1–38.
- McLean, R. A. (2007). Kyphophyllid rugose corals from the Frasnian (Upper Devonian) of Canada and their biostratigraphic significance. Palaeontographica Canadiana, 26, 1–109.
- McLean, R. A. (2010). Frasnian (Upper Devonian) colonial disphyllid corals from Western Canada. Ottawa, Ontario: NRC Research Press.
- May, A. (1995). Thamnopora (Anthozoa; Tabulata) aus dem Givetium bis Frasnium von Asturien (Devon; Nord-Spanien). Münstersche Forschungen für Geologie und Paläontologie, 77, 479–491.
- Michelin, H. (1840-1847). Iconographie zoophytologique, description par localités et terrains des polypiers fossiles de France et pays environnants Paris, Bertrand (Ed.), 1–348.
- Milne-Edwards, H., & Haime, J. (1851). Monographie des polypiers fossiles des terrains paléozoïques. Archives du Muséum d’Histoire Naturelle, Paris, 5, 1–502.
- Milne-Edwards, H., & Haime, J. (1850). A monograph of the British fossil corals. London, GB, Palaeontographical Society Monography, i-lxxxv + 1–71.
- Monod, O. (1967). Présence d’une faune ordovicienne dans les schistes de Seydişehir à la base des calcaires du Taurus occidental. Bulletin of the Mineral Resource and Exploration Institute (MTA, Ankara), 69, 78–89.
- Monod, O., Kozlu, H., Ghienne, J. F., Dean, W. T., Günay, Y., Le Hérissé, A., Paris, F., & Robardet, M. (2003). Late Ordovician glaciation in southern Turkey. Terra Nova, 15, 249–257.
- Nothdurft, L. D., & Webb, G. E. (2009). Clypeotheca, a new skeletal structure in scleractinian corals: a potential stress indicator. Coral Reefs, 28, 143–153.
- Nowinski, A. (1976). Tabulata and Chaetetida from the Devonian and Carboniferous of southern Poland. Palaeontologica Polonica, 55, 1–125.
- Oliver, W. A., Jr (1966). Description of dimorphism in Striatopora flexuosa Hall. Palaeontology, 9, 448–454.
- Oliver, W. A., Jr (1968). Some aspects of colony development in corals. Journal of Paleontology, 42, 16–34.
- Özgül, N., Metin, S., Erdoğan, B., Göğer, E., Bingöl, I., & Baydar, O. (1973). Tufanbeyli dolayinin Kambryien ve Tersiyer kayalari [Cambrian-Tertiary rocks of the Tufanbeyli region, Eastern Taurus, Turkey]. Bulletin of the geological Society of Turkey, 16, 82–100. ( in Turkish, English abstract).
- Paris, F., Le Hérissé, A., Monod, O., Kozlu, H., Ghienne, J.-F., Dean, W. T., Vecoli, M., & Günay, Y. (2007). Ordovician chitinozoans and acritarchs from southern and southeastern Turkey. Revue de Micropaléontologie, 50, 81–107.
- Plusquellec Y. (1976). Les polypiers - Tabulata. In H. Lardeux (Ed.), Les schistes et calcaires éodévoniens de Saint-Céneré (Massif armoricain, France). Mémoires de la Société géologique et minéralogique de Bretagne, 19, 183–215.
- Plusquellec, Y. (1991). Bourgeonnement chez Saouraepora gigantea et Praemichelinia homofavosa, Tabulata du Dévonien d’Algérie. Geobios, 24, 47–57.
- Plusquellec, Y., Tourneur, F., & Lafuste, J. (1993). Saouraepora nouveau genre de Micheliniidae (Tabulata), du Dévonien du Nord Gondwana et du Carbonifère d’Amérique du Nord. Palaeontographica A, 227(1/3), 1–86.
- Powell, J. H., & Scrutton, C. T. (1978). Variation in the Silurian tabulate coral Paleofavosites asper, and the status of Mesofavosites. Palaeontology, 21, 307–319.
- Rohart, J. C. (1988). Rugueux givetiens et frasniens de Ferques (Boulonnais-France). In D. Brice (Ed.), Le Dévonien de Ferques. Bas-Boulonnais (N. France). Biostratigraphie du Paléozoïque (Vol. 7, pp. 231–297).
- Rohart, J. C. (1999). Palaeozoic rugose corals from central and eastern Iran (A.F. De Lapparent and M. Zahedi collections). Annales de la Société Géologique du Nord, 7, 47–70.
- Rohart, J. C. (2000). Frasnian rugose corals from Chahriseh (Esfahan Province, Central Iran). Annales de la Société Géologique du Nord, 8, 67–71.
- Roniewicz, E., & Stolarski, J. (1999). Evolutionary trends in the epithecate scleractinian corals. Acta Palaeontologica Polonica, 44, 131–166.
- Rozkowska, M. (1979). Contribution to the Frasnian Tetracorals from Poland. Palaeontologica Polonica, 40, 3–56.
- Schlotheim , E. F.von (1820). Die Petrefaktenkunde auf ihren jetzigen Standpunkt durch die Beschreibung seiner Sammlung versteinerter und fossiler Überreste des Thier- und Pflanzenreichs der Vorwelt erläutert. LXII + 1-437, Gotha.
- Schröder, S. (2005). Stratigraphie und Systematik rugoser Korallen aus dem Givetium und Unter-Frasnium des Rheinischen Schiefergebirges (Sauerland/Bergisches Land). Zitteliana, B25, 39–116.
- Scotese, C. R., & Golonka, J. (1992). PALEOMAP paleogeographic atlas, PALEOMAP progress report #20. Arlington: Dept. of Geology, University of Texas.
- Smith, S. (1945). Upper Devonian corals of the Mackenzie River region, Canada. Special Papers Geological Society of America, 59, 1–126.
- Sokolov, B. S. (1962). Osnovy paleontologii; Podclass Tabulata [Fundamentals of paleontology; Subclass Tabulata], 192–265. Moskova: Izdatelstvo Akademii Nauk SSSR.
- Sorauf, J. E. (1967). Massive Devonian Rugosa of Belgium. The University of Kansas Paleontological Contributions, 16, 1–41.
- Sorauf, J. E. (1998). Frasnian (Upper Devonian) rugose corals from the Lime Creek and Shell Rock Formations of Iowa. Bulletins of American Paleontology, 113 (355), 1–159.
- Soshkina, E. D. (1949). Devonskie korally Rugosa Urala [Devonian corals Rugosa of the Urals]. Trudy Paleontologicheskogo Instituta Akademiya Nauk SSSR, 15(4), 1–160.
- Soshkina, E. D. (1951). Pozdnedevonskie korally Rugosa, ikh sistematika i evolyutsiya [Late Devonian corals Rugosa, their systematics and evolution]. Trudy Paleontologicheskogo Instituta Akademiya Nauk SSSR, 34, 1–122.
- Stel, J. H. (1978). Studies on the paleobiology of favositids. Goningen:Rijksuniversiteit te Groningen..
- Stolarski, J. (1995). Ontogenetic development of the thecal structures in caryophylliine scleractinian corals. Acta Palaeontologica Polonica, 40, 19–44.
- Stolarski, J. (1996). Gardinera – a scleractinian living fossil. Acta Palaeontologica Polonica, 41, 339–367.
- Swann, D. H. (1947). The Favosites alpenensis lineage in the Middle Devonian Traverse Group of Michigan. Contributions from the Museum of Paleontology, University of Michigan, 6, 235–318.
- Tourneur, F. (1985). Contribution à l’étude des tabulés du Dévonien moyen de la Belgique. Systématique et stratigraphie (unpublished thesis). Belgium: Université catholique de Louvain, 568 pp.
- Tourneur, F. (1986). Microstructure des genres Thamnopora Steininger, 1831 et Lecomptopora nov. gen., Tabulés branchus du Dévonien moyen. Comptes Rendus Académie des Sciences Paris, 303, 1255–1258.
- Tourneur, F., Babin, C., Bigey, F., Boulvain, F., Brice, D., Coen-Aubert, M., Dreesen, R., Dusar, M., Loboziak, S., Loy, W., & Streel, M. (1989). Le Dévonien du sondage de Nieuwkerke (Flandre Occidentale, Belgique – extrémité occidentale du Synclinorium de Namur). Annales de la Société Géologique du Nord, 108, 85–112.
- Tourneur, F., & Fernández-Martínez, E. (1991). Parastriatopora cantabrica, nueva especie de tabulado del Devónico inferior (Emsiense) de la Cordillera cantabrica (NO de España). Revista Española de Paleontologia, 6, 3–19.
- Tsien, H. H. (1970). Espèces du genre Disphyllum (Rugosa) dans le Dévonien moyen et le Frasnien de la Belgique. Annales de la Société Géologique de Belgique, 93, 159–182.
- Verrill, A. E. (1865). Classification of polyps (Extract condensed from a synopsis of the polypi of the North Pacific Exploring Expedition, under captains Ringgold and Rodgers, U.S.N.). Essex Institute Proceedings, 4, 145–149.
- Wedekind, R. (1922). Zur Kenntnis der Stringophyllen des oberen Mitteldevon. Sitzungsberichte der Gesellschaft zur Beförderung der gesamten Naturwissenschaften zu Marburg, 19211, 1–16.
- Wedekind, R. (1937). Einfürung in die Grundlagen der historischen Geologie, II. Band. Microbiostratigraphie die Korallen- und Foraminiferenzeit. Stuttgart, Germany: Ferdinand Enke.
- Wehrmann, A., Yılmaz, I., Yalçın, M. N., Wilde, V., Schindler, E., Weddige, K., Saydam Demirtas, G., Özkan, R., Nazik, A., Nalcıoğlu, G., Kozlu, H., Karslıoğlu, Ö., Jansen, U., Ertuğ, K., Brocke, R., & Bozdoğan, N. (2010). Devonian shallow-water sequences from the North Gondwana coastal margin (Central and Eastern Taurides, Turkey): Sedimentology, facies and global events. Gondwana Research, 17, 546–560.
- Weyer, D. (1975). Combophyllidae (Anthozoa, Rugosa) im Mitteldevon des Thüringer Schiefergebirges. Freiberger Forschungshafte C, 304, 7–31.
- Yalçın, M. N., & Yılmaz, I. (2010). Devonian in Turkey – a review. Geologica Carpathica, 61, 235–253.
- Zapalski, M. (2012). Tabulate corals from the Givetian and Frasnian of the southern region of the Holy Cross Mountains (Poland). Special Papers in Paleontology, 87, 1–100.