
Abstract
Thirteen ichnogenera and seventeen ichnospecies were recognised in carbonate rocks from the Lower Permian Taiyuan Formation in North China Basin. Based on the composition, occurrence and diversity of ichnofossils, intensity of bioturbation, lithologic characteristics, especially on the variation of individual size and the filling colour diversities of the Zoophycos burrows, four types of ichnofabrics were recognised, that is (1) epigenic tiering ichnofabric – mainly appeared on or near the bedding plane and formed in restricted bay or lagoon sedimentary environments; (2) shallow tiering ichnofabric – developed less than 15 cm under the bedding plane and occurred in the upper shallow-marine environment; (3) medium tiering ichnofabric – mainly distributed from 15 to 20 cm under the bedding plane and generated in the mid-upper shallow sea (restricted–open platform); (4) deep tiering ichnofabric – distributed from 25 to 30 cm under the bedding plane and occurred in lower shallow sea (open platform) far from shore. Based on ichnological and sedimentological analyses above four types of ichnofabrics, the sedimentary environment model of the ichnofabrics in carbonate rocks from the Taiyuan Formation in North China Basin is proposed in this paper.
1. Introduction
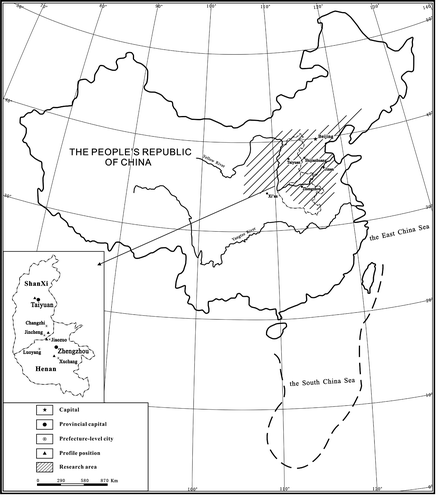
The past ichnological researches in Permian shallow-marine carbonates mainly related to the ichnofacies or ichnoassemblages (Knaust, Citation2008; Knaust & Bromley, Citation2012) and microbial fossils in Zoophycos (Gong et al., Citation2007; Gong, Shi, Weldon, Du, & Xu, Citation2008), but about ichnofabric studied results were reported rarely. In China, some studies about the ichnofossils in shallow-marine carbonate rocks of the Lower Permian Taiyuan Formation in North China Basin had been made formerly. Hu (Citation1985) found a lot of Coprolites of tidal flat deposits in the limestone of the upper part of the Taiyuan Formation in the west Xinggong coal field of western Henan Province. Wu et al. (Citation1987) illustrated the relationship between ichnofossils and sedimentary sequence of tempestites in shallow-marine carbonate rocks of Taiyuan Formation in Jiaozuo area. Hu and Qi (Citation2000) investigated the characteristics of Zoophycos ichnofabric in limestones of the Taiyuan Formation. Zhu and Li (Citation2005) found the Teichichnus-like ichnofossil of intertidal sedimentary environment in the limestone of this formation in Xinmi coal field. Hu, Zhou, and Song (Citation2010) and Song et al. (Citation2012) comprehensively studied the distribution characteristics of ichnofossils, types of ichnofabrics and their sedimentary environments of the Taiyuan Formation in Jiaozuo area. At the same time, four kinds of different colours of fillings in Zoophycos burrows were recognised by microstructure and ultrastructure analysis. Also, Hu et al. (Citation2013) recognised four types of ichnoassemblages and their sedimentary environments in the limestones of the Taiyuan Formation in south-eastern Shanxi Province, and 10 ichnogenera and 14 ichnospecies were distinguished. This work attempts to study systematically the taxonomy of ichnofossils and ichnofabric types and their tier features in the carbonate rocks of the Taiyuan Formation that was well exposed in Xishan of Taiyuan city and Fuchen of Lingchuan county in Shanxi Province and west Jiaozuo city and Longhuashan of Yuzhou county in western Henan Province (Figure ) and analyse the relationship between the changes of tiers and the sedimentary environments. The research result can provide a lot of important information in biogenic structure for exploring the sedimentary environment evolution of the carbonate rocks in the Taiyuan Formation of this study area.
Figure 1. Location map of study area and sections (▲: location of the study sections).
2. Geological background
From the regional structure, the Late Palaeozoic deposit areas of south-eastern Shanxi Province and western Henan Province are located in the middle-southern part of North China block within Sino-Korean plate. During the later period of the Caledonian movement, the western tectonic landform of North China blocks displays uplift, while the east part is decline (Shang, Citation1995). However, during the Carboniferous, the North China block became ‘south rise and north drop’, and then, to the Early Permian, the block transformed into ‘north rise and south drop’, so, the paleotopographic features of that time showed north-west higher and south-east lower, seawater frequently was transgressive and regressive approximately along northwest–southeast direction. Consequently, the Lower Permian Taiyuan Formation in North China block formed in such the sedimentary environments as shore, shallow sea, lagoon and tidal flat under the tectono-palaeogeographic setting afore mentioned.
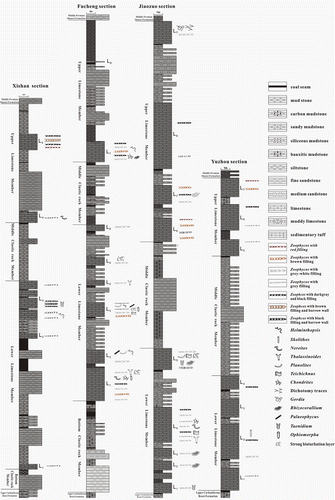
As a whole, the Lower Permian Taiyuan Formation of the North China Basin is a set of interactive deposits of carbonate rocks and clastic rocks, and mainly consists of limestone, sandstone, siltstone, mudstone and coal beds (Figure ). The bottom of the Taiyuan Formation in Henan Province is a thin layer limestone (L1), the uppermost limestone (L9) or siliceous rock layer is the top boundary of this Formation, it can be divided from bottom to top into the Lower Limestone, Middle Clastic rock and Upper Limestone Members. However, in Shanxi Province, the Taiyuan Formation can be divided from bottom to top into four members; they are Lower Clastic Rock, Lower Limestone, Middle Clastic Rock and Upper limestone Members, respectively. Because of finding some of the body fossil Pseudoschwagerina (Fusulinida) in the bottom limestone of the Lower Limestone member, strictly speaking, the geologic time of the Bottom Clastic Rock Member belongs to the Late Carboniferous. But, from lithostratigraphic classification, the bottom boundary of this Formation was determined as the bottom of the thick-bedded sandstone, and its top is also a bed of thick sandstone (Figure , left). In contrast, the Bottom Clastic Rock member is almost not developed in the realm of Henan Province (Figure , right).
Figure 2. Stratigraphic column of study sections and vertical distribution characteristics of ichnofossils from the Taiyuan Formation in North China Basin.
The lithologic characteristics of each member in the Taiyuan Formation are described in brief as follows:
| (1) | The Bottom Clastic Rock Member is made up of thin- to thick-bedded fine-grained sandstone, siltstone, sandy mudstone, carbonaceous shale and coal; | ||||
| (2) | The Lower Limestone Member contains 3–5 layer limestones, which is composed of dark grey, medium thick-bedded or thick-bedded limestone containing partly crumby flint, bioclastic limestone and micrite intercalated with thin-bedded fine-grained sandstone, siltstone, silty mudstone, mudstone and coal beds; | ||||
| (3) | The Middle Clastic Rock Member consists of off-white, medium thick-bedded or thick-bedded, medium- to fine-grained quartz sandstone and thin-bedded siltstone and sandy mudstone, containing 2–3 coal beds and 2–3 layers of unstable bioclastic limestones (frequently lenticular body); | ||||
| (4) | The Upper Limestone Member usually developed 2–4 layers of limestones, which is marked by grey or dark-grey bioclastic limestone, and fine- and medium-grained sandstone, siltstone, sandy mudstone and thin coal beds (or extremely thin coal layers). | ||||
This work researches four outcrop sections of the Taiyuan Formation, including Taiyuan Xishan (five layers of limestones) and Fucheng (eight layers of limestones) sections in Qinshui basin of Shanxi province, and Jiaozuo (nine layers of limestones) and Yuzhou (seven layers of limestones) sections in western Henan province, almost all the layers of limestones contain abundant ichnofossils, as shown in Figure .
3. Composition and description of ichnofossils
A lot of ichnofossils discovered in carbonate rocks of the Lower Permian Taiyuan Formation in North China Basin include 13 ichnogenera and 17 ichnospecies, and mainly are fodinichnia, pascichnia and domichnia based on the ethological classification. The common feeding traces mainly are Zoophycos brianteus, Zoophycos villae, Zoophycos aff. cauda-galli, Zoophycos isp. (1) Zoophycos isp. (2) Teichichnus rectus, Rhizocorallium isp., Chondrites isp., Planolites isp., Palaeophycus cf. heberti and Taenidium satanassi; some pascichnia are Gordia marina, Helminthopsis isp. and Nereites isp.; several kinds of dwelling burrows are Thalassinoides suevicus, Ophiomorpha isp. and Skolithos isp. and other dichotomy traces. The brief characteristics of these ichnofossils are described as follows:
Figure 3. Ichnofossils on the bedding plane from the Taiyuan Formation in Jiaozuo and Fucheng areas. (a) Chondrites isp. and Nereites isp. from the limestone (L4) of Jiaozuo; (b) Helminthopsis isp. from the limestone (L2) of Fucheng; (c) Dichotomy traces from the limestone (L9) of Jiaozuo; (d) Gordia marina from the limestone (L9) of Jiaozuo.

3.1. Chondrites isp. (Figure (a))
Description: The trace fossils here observed are the complex small-sized branching burrows, showing spindly ellipsoidal spotted aggregation in the longitudinal section, diameter of tunnels, .5–1.2 mm, but length of the burrow system is unknown. The burrow wall is smooth, only partly, with a paper-thin dark grey argillaceous lining. The filling of burrow is grey or grey white carbonaceous mudstone and obviously different from the host rock. These burrows generated in medium- to thick-bedded limestone and associated with Nereites constantly.
Horizon and location: L4 limestone bed from Jiaozuo section, west Henan province, L1 and L7 limestone beds from Fucheng section, Qinshui Basin, Shanxi province.
Discussion: The Chondrites is a regularly branched burrow system constructed by endobenthic deposit-feeding animals of unknown taxonomic affinity and is considered as an indicator of oxygen-poor conditions (Bromley & Ekdale, Citation1984). Jin, Li, and Fang (Citation2003) researched the ichnofossils developed in deep-sea deposits in China and discovered that Chondrites is a cross-facies ichnofossil. Gong (Citation2004) discussed detailedly the facies indication and tier characteristics of Chondrites and listed many research achievements, most of scholars considered that Chondrites is the facies fossil indicating oxygen-deficient or oxygen-low environment, and Chondrites is a kind of chemical symbiotic structure made by worms feeding on chemosynthetic bacteria in the oxygen-deficient environment.
3.2. Dichotomy traces (Figure (c))
Description: Slightly curved dichotomy traces (similar to ‘>’ form) observed on the bedding plane, smooth surface, one side of branch is 2–3 mm wide, another side 1–2 mm wide, 5–9 cm in length, less than 1 mm in depth and intersection angle between two branches is 15°–35°. The traces are often paired occurrence on the bedding plane and associated commonly with Gordia.
Horizon and location: L9 limestone bed from Jiaozuo section in Henan province.
Discussion: The traces occur on the bedding plane of some muddy limestones and are characterised by a dichotomy form and often associated with Gordia. Their trace maker is unclear, perhaps, relative to some gastropods.
3.3. Gordia marina Emmons, 1844 (Figure (d))
Description: The specimens here observed are a long, smooth, sinuous to meandering trails, or free curving groove marks, 2–7 mm in width, frequent crossing with each other and frequent forming irregular loops. These traces are preserved in epichnia groove on the bedding plane of muddy limestone.
Horizon and location: L9 limestone bed from Jiaozuo section in Henan province.
Discussion: This ichnogenus is a common wide-facies ichnofossil and developed in the calm shallow water and deepwater substrate (Gong, Citation1994; Gong & Si, Citation2002). Generally, its trace makers are worms (Aceñolaza & Buatois, Citation1993; McCann, Citation1993) or gastropods (Yang, Citation1984).
3.4. Helminthopsis isp. (Figure (b))
Description: Simple, smooth, unbranched, irregularly winding or loosely winding burrows preserved densely near bedding plane of muddy limestone. Observed ‘burrow segments’ is more than 4.5 cm long, with 2.6 mm in diameter, locally crossing with each other, with grey filling that is obviously different from its host rock.
Horizon and location: L2, L3 and L7 limestone beds from Fucheng section and L3 limestone bed from Taiyuan Xishan section in Shanxi province.
Discussion: The specimen here observed is characterised by its low degree of winding, narrow diameter and presence of ‘burrow segments’. It is well known that the Helminthopsis is a kind of wide-depth trace fossils, commonly generated in muddy sediment substrate, and its trace makers are explained mostly as polychaete or priapuloidea.
3.5. Nereites isp. (Figure (a))
Description: Simple, irregularly curved or meandering trail, both sides consisting clearly of numerous irregular, tiny oval lobes made by black mudstone. The width of the burrow is 2–4 mm and the length observed ‘burrow segments’ is more than 9 cm. Lobes either isolate or adjacent but never overlap, larger isolate lobes 3 × 2 mm in size, smaller lobes 2 × 1 mm in size. The burrows are preserved on bedding plane of medium thick-bedded limestone.
Horizon and location: L4 limestone bed from Jiaozuo section in Henan province; L1 limestone bed from Fucheng section and L4 limestone bed from Xishan section in Shanxi province.
Discussion: The described specimen is characterised by the irregularly curved trail associated with two lateral tiny lobes. Seilacher (Citation1983) explained Nereites as trails formed in substrate and made by infauna feeding on sediments and suggested that Nereites developed mostly on the bedding plane of straticulate turbidite that commonly occur in abysmal sea or moderate sea and is a typical ichnofossil of abysmal sea ichnofacies. However, Crimes and Anderson (Citation1985) thought that Nereites can also exist in shallow water environment and it is a wide-depth ichnofossil. Some scholars explained the trace makers of the Nereites probably are worms like organism such as polychaetes (Parcha & Pandey, Citation2011).
Figure 4. Ichnofossils in the Taiyuan Formation from Fucheng and Jiaozuo areas. (a) Ophiomorpha isp. from the limestone (L3) of Fucheng; (b) Palaeophycus cf. heberti from the limestone (L2) of Fucheng; (c) Planolites isp. from the limestone (L2) of Fucheng; (d) Rhizocorallium isp. from the limestone (L5) of Jiaozuo; (e) Rhizocorallium isp. from the limestone (L3) of Jiaozuo.

3.6. Ophiomorpha isp. (Figure (a))
Description: This specimen occurs as straight to gently curved branching and horizontal tunnel that is oval or rounded in cross section, 6–8 mm in diameter and the length observed ‘burrow segments’ is more than 11 cm. The branching style of the tunnel is mainly Y-shaped but also T-shaped, and the branching swell area is about 9–13 mm in diameter. The burrow exhibits a pelletal lining constituted by densely tiny mud nodules. The burrow is preserved in endichnia in medium thick-bedded limestone.
Horizon and location: L1 and L3 limestone beds from Fucheng section in Shanxi province.
Discussion: The burrow was found locally in a few of limestone beds of Taiyuan Formation and is closely similar to Ophiomorpha nodosa in the morphology and lining characteristics. Generally, Ophiomorpha is domichnia, or domichnia and fodinichnia simultaneously, and its distribution is controlled by such factors as salinity, substrate property and depth of water. Some scholars suggested that the main control factors are oxygen content in water and substrate property rather than substrate component, salinity or depth (Cui, Citation1999). In modern environment, main trace maker of this burrow is Crustacea that commonly occurs in tidal flat, estuary, lagoon environments; however, in geological history, the palaeoenvironments lived by trace maker of this burrow changed greatly, from tidal flat, shallow sea to outer shelf, even to deep sea (Uchman, Citation2007, 2009) and terrestrial environments; therefore, the ichnogenus is a kind of wide-facies ichnofossil.
3.7. Palaeophycus cf. heberti (Figure (b))
Description: Slightly curved, cylindrical burrow with circular cross section, 5–8 mm in diameter, 7 cm in the length observed, grey burrow lining 1–3 mm thick. The burrows are oriented nearly parallel to the bedding plane, earthy yellow filling is structureless and similar to host sediment. The burrows are preserved in muddy limestone.
Horizon and location: L2 and L3 limestone beds from Fucheng section in Shanxi province.
Discussion: The specimen is characterised by the slightly curved, cylindrical burrow with moderately thick lining and resembles Palaeophycus heberti described by Frey and Howard (Citation1985). Generally, this kind of burrow is domichnia (Pemberton & Frey, Citation1982), its trace makers are considered as benthic invertebrates, including worms, arthropods or insecta. Palaeophycus and Planolites are similar in shape, whereas there is the lining in the burrow of Palaeophycus. About the lining, Melchor et al. (Citation2006) thought that it is not for reinforcing burrows, but preventing the water run-off from the gradual drying sediments. The ichnogenus can exist in all kinds of marine environments including marginal sea and freshwater environments.
3.8. Planolites isp. (Figure (c))
Description: Straight to slightly curved, unbranched and unwalled smooth burrow about 10 mm in diameter and up to 10 cm in visible length, filling composition is a similar to the host rock. The burrows are uniform in diameter and are oriented parallel or oblique to the bedding plane.
Horizon and location: L5 limestone bed from Jiaozuo section in Henan province; L2 and L3 limestone beds from Fucheng section in Shanxi province.
Discussion: The burrows here studied mainly occur in the thin-bedded muddy limestone; therefore, it is probably made by a deposit-feeding worm. Generally, the ichnogenus Planolites is often interpreted to develop in various sedimentary environments.
3.9. Rhizocorallium isp. (Figure (d), (e))
Description: The first specimen (Figure (d)) shows the spreite burrow consisting of subhorizontal tunnel and oblique shaft in the longitudinal section of limestone bed, 5–12 mm in width. Another specimen (Figure (e)) occurs as the spreite tubular burrow and the U-shaped marginal tube with more or less parallel limbs, maximum length observed is 15 cm; interval between two side marginal tubes is 3.5–4 cm and the tube diameters are 1–1.3 cm; the spreite made up of the numerous tightly arranged tunnels (.5 cm in width) spreading between the limbs; the burrows are preserved as horizontal to subhorizontal flattened tubes along the bedding plane of the calcareous siltstone.
Figure 5. Ichnofossils from the Taiyuan Formation in Yuzhou, Jiaozuo and Fucheng areas. (a, b) Skolithos isp. from the limestone (L2) of Yuzhou; (c) Taenidium satanassi from the limestone (L5) of Jiaozuo; (d) Teichichnus rectus from the limestone (L3) of Fucheng; (e) Thalassinoides suevicus from the limestone (L4) of Jiaozuo.

Figure 6. Zoophycos on the bedding plane from the Taiyuan Formation of Fucheng and Jiaozuo areas. (a) Zoophycos brianteus from the limestone (L2) of Fucheng; (b) Zoophycos brianteus with black filling from the limestone (L2) of Fucheng; (c) Zoophycos villae from the limestone (L2) of Fucheng; (d) Zoophycos aff. cauda-galli from the limestone (L6) of Fucheng; (e) Zoophycos isp. 1 from the limestone (L7) of Jiaozuo; (f) Zoophycos isp. 2 from the limestone (L7) of Fucheng.

Figure 7. The composition and occurrence characteristics of ichnofossils in the epigenic tiering ichnofabric.
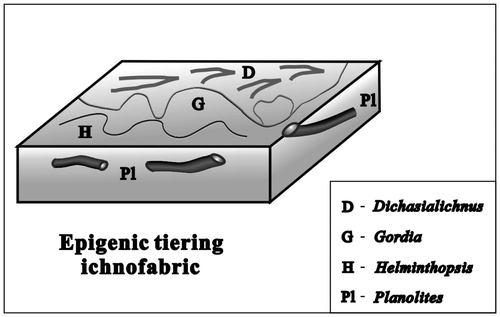
Figure 8. Gordia (a) and together with dichotomy traces (b) in the Taiyuan Formation L9 limestone of Jiaozuo area.

Figure 9. The composition and occurrence characteristics of ichnofossils in the shallow tiering ichnofabric.
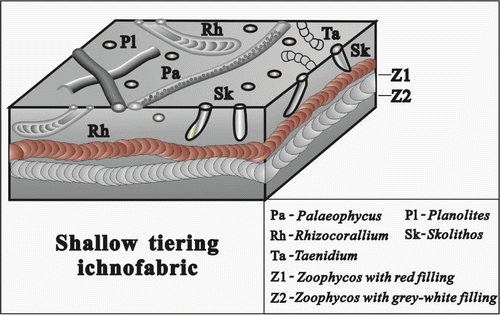
Figure 10. Zoophycos with red filling in the Taiyuan Formation L6 limestone of Jiaozuo area.

Figure 11. Zoophycos with grey-white filling in the Taiyuan Formation L4 limestone of Jiaozuo area.

Figure 12. The composition and occurrence characteristics of ichnofossils in the medium tiering ichnofabric.
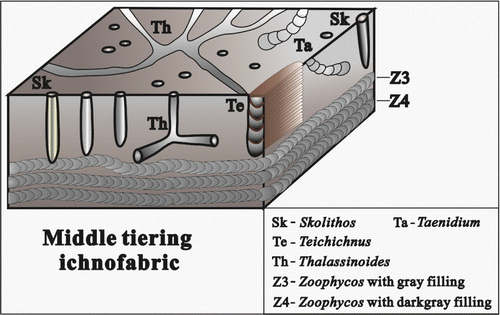
Figure 13. Bioturbation (spotted) structure in the Taiyuan Formation L5 limestone of Jiaozuo area.

Figure 14. Zoophycos with grey or dark-grey filling in the Taiyuan Formation L5 limestone of Jiaozuo area.

Figure 15. The composition and occurrence characteristics of ichnofossils in the deep tiering ichnofabric.
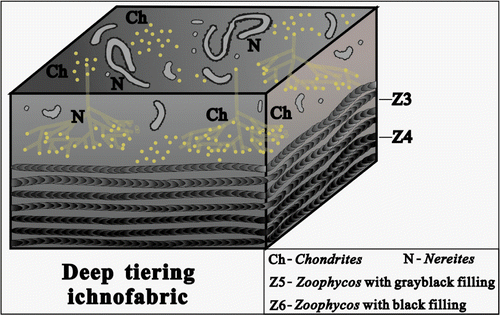
Figure 16. The small type of Zoophycos with dark-grey filling in the Taiyuan Formation L3 limestone from Taiyuan Xishan area.

Figure 17. Zoophycos with black filling in the Taiyuan Formation L4 limestone of Jiaozuo area.

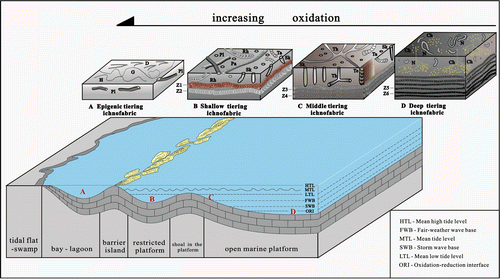
Figure 18. Sedimentary environment model of the ichnofabrics in the Taiyuan Formation of North China Basin.
Horizon and location: L2, L3, L5 and L7 limestone beds from Jiaozuo section in Henan province; L7 limestone bed from Fucheng section in Shanxi province.
Discussion: Rhizocorallium is one of the oldest known trace fossils, with wide distribution through the Phanerozoic and all over the world (Knaust, Citation2013) and is generally attributed to deposit-feeding Crustaceans or annelids (Pemberton & Frey, Citation1982; Sarkar, Sanjoy Kumar, & Chandan, Citation2009). The burrows can be divided into horizontal-type (parallel or oblique to the bedding plane) and oblique-type. The former represent subtidal zone with low energy (Seilacher, Citation1967); however, some scholars suggested that they can also exist in tidal flat environment, what is more, horizontal Rhizocorallium forming in this condition is often associated with some other burrows perpendicular to the bedding plane (Xia et al., Citation1993).
3.10. Skolithos isp. (Figure (a), (b))
Description: Straight, unbranched tube burrow nearly perpendicular to the bedding plane, cylindrical or sub-cylindrical in section, diameter ranges usually from 3 to 8 mm but some reach 10 mm, length is more variable, it averages 20 cm but reaches 35 cm (Figure (a)). Burrow filling mainly consists of the light-grey calcareous and muddy sediments, but their host rock is dark-grey finely crystalline limestone with many bioclastics. The burrows distributed dispersively on the bedding plane (Figure (b)).
Horizon and location: L2 limestone bed from Yuzhou section in Henan province.
Discussion: The studied specimens are consistent with the ichnogenus Skolithos, and similar to Skolithos linearis in size. Usually, Skolithos is quite common in sandstone and siltstone deposits of shallow-marine palaeoenvironment (Fitzgerald & Barrett, Citation1986), but less common in marine carbonate rock deposits. It may be considered that most Skolithos generated in higher energy sedimentary environment (Knaust & Bromley, Citation2012).
3.11. Taenidium satanassi D’ Alessandro et Bromley, 1987 (Figure (c))
Description: Straight or slightly curved, unbranched, unlined, backfilled burrow spreading along the bedding plan of limestone. Observed specimen associated with Zoophycos and the burrows are 2–6 mm in width and 20–24 mm in visible length, with distinct crescent backfill structure with .8 mm in width and about .4 mm space between the adjacent backfilling lamellae. The width of crescent backfilling lamellae in the burrows is almost equal and much smaller than the burrow width. The filling presents the same composition as the host rock.
Horizon and location: L1 and L5 limestone beds from Jiaozuo section in Henan province; L3 limestone bed from Taiyuan Xishan section in Shanxi province.
Discussion: Main characteristics of the ichnospecies display that the width of every backfilling lamellae is almost equal and much smaller than the width of the burrow. Previous studies had shown that Taenidium can exist in shallow sea (Buckman, Citation2001), river and lake deposits (Melchor et al., Citation2006; Savrda, Citation2000).
3.12. Teichichnus rectus Seilacher, 1955 (Figure (d))
Description: The trace fossils appear as a near-vertical or steeply inclined retrusive spreiten structures or U-shaped burrow in longitudinal section, 1.5 cm in width, 7 cm in length, containing numerous concave-upwards crescent-shaped or very low-curvature laminae, and 4–8 mm in interval between adjacent lamina. Their host rock is micrite.
Horizon and location: L4 and L5 limestone layers from Jiaozuo section in Henan province; L3 limestone bed from Fucheng section in Shanxi province.
Discussion: The trace fossil is consistent to Teichichnus rectus Seilacher, 1955 in the characteristics of morphology, size and occurrence and is usually interpreted as possible feeding-dwelling/equilibrium structures. The spreiten structures formerly found mostly generated in the clastic rock deposits formed in shoreface, intertidal zone, lagoon, estuary, delta, deep-sea fan and deep-sea plain sedimentary environments and their trace makers are commonly explained as worms, annulata or arthropod (Chamberlain, Citation1977; Chisholm, Citation1970; Corner & Fjalstad, Citation1993; Loughlin & Hillier, Citation2010).
3.13. Thalassinoides suevicus Kennedy, 1966 (Figure (e))
Description: The Thalassinoides studied here is the compound burrow systems with straight to slightly curved, cylindrical tunnels and is smooth-walled and unlined (i.e. without a pelleted burrow wall). The burrows mostly have Y- and T-shaped branches, 7–12 mm in diameter, and burrow junctions form swollen areas larger than 24 mm in size. The colour of burrow fillings is dark grey and visibly darker than the host rock, in which a part of the burrow fillings was weathered to change into yellowish-brown calcareous mudstone. The tunnels are oriented parallel to the bedding plane of the grey argillaceous limestone beds.
Horizon and location: L4 limestones from Jiaozuo section in Henan province; L1 and L4 limestones from Fucheng section in Shanxi province.
Discussion: The trace fossils are consistent to the Thalassinoides suevicus in the characteristics and size of branching systems. The Thalassinoides is generally considered as dwelling and feeding traces of decapoda crustaceans (Bromley, Citation1996; Ekdale, Citation1992; Miller & Curran, Citation2001; Wetzel, Citation1991), and deposit-feeding crabs, shrimps and lobsters can create extensive Thalassinoides-like burrow systems in modern tidal flat and marine environments (Ekdale & Bromley, Citation2003). In geological record, most of the Thalassinoides are usually interpreted to form in intertidal zone, brackish water lagoon and shallow-marine environments (Miller, Citation2001; Sharafi et al., Citation2012; Yang et al., Citation2004).
3.14. Zoophycos brianteus Massalongo, 1855 (Figure (a), (b))
Description: This ichnospecies is a kind of intensive spreiten structures consisting of J-shaped protrusive burrows that are spiral lamellae around a thicker centre column tube burrow. Numerous primary and few secondary lamellae can be observed on the bedding plane. In the specimen, an apical axis occurs on the centre of this specimen, and a lot of lamellae extend at a low angle from the centre to margin. The primary lamellae are about 1 mm (Figure (a)) wide or 3–6 mm wide and 16 cm × 15 cm in the whole contour (Figure (b)). Also, a series of clear crescented backfilling lamina appear and are oriented from oblique to parallel to the bedding plane on the longitudinal section. This ichnofossil is preserved in endochnia in thin- to medium thick-bedded limestone or chert band. In the specimen, spreiten structures are protrusive on the weathered bedding plane and the burrow fillings are dark limy mudstone or silica mudstone.
Horizon and location: Every limestone layer of the Taiyuan Formation.
Discussion: Villa (Citation1844) described some large-scale rounded trace fossils with a compacted centre spirally surrounded by some lamellae, they were found in Brianza, Italy and were considered as phytolite and named Fucoides brianteus, but no picture afforded in the text. Massalongo (Citation1855) originally named Zoophycos brianteus found in the limestone of turbidite facies from upper Cretaceous in Livorno, Italy, and showed typical Zoophycos characteristic. Based on the lectotype of Zoophycos brianteus Massalongo, 1855 found in Verona museum, Olivero (Citation2007) determined Zoophycos brianteus Massalongo, 1855 as a type species of Zoophycos Massalongo, 1855. The specimen found here is almost the same as the type species in characteristics.
3.15. Zoophycos villae Massalongo, 1855 (Figure (c))
Description: This ichnospecies is characterised by several set of parabolic and J-shaped lamellae with a big radian at the end of marginal tube on the bedding plane. The primary lamellae with .3–.5 mm wide are branched two or three smaller lamellae with .1–.2 mm wide. This specimen is 6 cm long and 5 cm wide. The primary lamellae radiate and curve from the centre to end. Centre column tube and marginal tube are not clear, but the backfill structures of slab lamina are clear on the longitudinal section. The burrow filling is red-brown silty mudstone and different from the host rock apparently.
Horizon and location: L6 limestone bed from Jiaozuo section in Henan province; L2 limestone bed from Fucheng section in Shanxi province.
Discussion: Through comparative analyses, above observed characteristics indicate that this specimen is almost the same as Zoophycos villae Massalongo, 1855.
3.16. Zoophycos aff. cauda-galli Vanuxem, 1842 (Figure (d))
Description: This specimen is an elliptic spreiten lamina consisting of clear primary lamellae .3 mm wide and secondary lamellae .15 mm wide, with 10 × 14 cm in the overall size. Middle part shows U-shaped spiral lamina on the bedding plane, its two sides are radiate lamellae with two or three branches from medium part to margin.
Horizon and location: L6 limestone bed from Fucheng section in Shanxi province.
Discussion: The Zoophycos cauda-galli found in calcareous and clayish sandstone from the Devonian Hamilton Group in New York and Ohio, USA (Vanuxem, Citation1842). The specimen shows an elliptic lamina consisting of clear primary and secondary lamellae. The specimen here is similar to Zoophycos cauda-galli in overall shape.
3.17. Zoophycos isp. 1 (Figure (e))
Description: This specimen shows the lobose spiral lamina with 9 × 8 cm in the overall size on the bedding plane, with 3–4 sets of lobed spreiten lamina. Marginal tube is clear in part, and the primary lamellae are compact, with 4–13 mm in space between spreite lobes. The burrow filling is black and is different greatly from the host rock.
Horizon and location: L7 limestone bed from Fucheng section in Shanxi province.
3.18. Zoophycos isp. 2 (Figure (f))
Description: This specimen shows a clear spreiten structure like a cocktail or palmate on the whole. The primary lamellae with .5–1.6 mm wide are clear on the bedding plane, and branches in the tail end are clearly visible, the secondary lamellae with .5 mm wide, but marginal tube is unsighted. The spiral burrows are parallel to the bedding plane, with 14 cm wide and 8.6 cm long, showing crescent-shaped backfilling lamellae on the longitudinal section.
Horizon and location: L7 limestone bed from Fucheng section in Shanxi province.
Above described the Zoophycos trace fossils are a large, well-preserved spreiten burrow system exhibiting numerous morphologic variations and extraordinary abundant in the Taiyuan Formation of this study area, which mainly formed in early Permian shallow-marine carbonate platform environments. In general, Palaeozoic Zoophycos occurs mainly in shallow-marine deposits, whereas Mesozoic and Cenozoic Zoophycos are common only in shelfal and deeper-marine deposits (Knaust, Citation2004, 2008; Olivero, Citation2003; Seilacher, Citation2007). Most workers agree that Zoophycos is the organised deposit-feeding burrow system of an infaunal animal and probably formed by chemobiosynthetic-feeding activity of annelids or worms (Bromley, Citation1991; Sarkar et al., Citation2009).
4. Ichnofabric types and tiering characteristics
Based on the composition, occurrence, abundance and diversity of ichnofossils specially, size and buried depth of Zoophycos, intensity of bioturbation and the colour of Zoophycos burrow filling in carbonate rock of the Taiyuan Formation in study area, meanwhile, combined with lithologic characteristics of host rock, four types of ichnofabrics can be recognised, they are epigenic tiering ichnofabric, shallow tiering ichnofabric, medium tiering ichnofabric and deep tiering ichnofabric, respectively.
4.1. Epigenic tiering ichnofabric
This ichnofabric (Figure ) mainly is characterised by Gordia and together with dichotomy traces (Figure ), sometimes, changing to Helminthopsis and Planolites. Generally, the ichnofossils in this ichnofabric, with higher abundance and lower diversity, mainly composed of trails on the bedding plane and a few fodinichnia near the bedding plane, and crosscut each other occasionally. This ichnofabric shows a medium to low intensity of bioturbation, bioturbation index (short for BI) is 2–3. Its host rocks are mostly argillaceous limestone and marlstone, and always overlaid by tidal flat deposits containing coal bed. This ichnofabric may be suggested to form in restricted bay or lagoon environment that is weak hydrodynamic and lower energy.
4.2. Shallow tiering ichnofabric
This ichnofabric mainly consists of large-sized Zoophycos associating often with Rhizocorallium, Taenidium or Planolites and Palaeophycus, sometimes, Skolithos found locally (Figure ), which is characterised by fodinichnia in substrata, but sometimes, domichnia occurred only. Most of these ichnofossils are common in the Cruziana ichnofacies, with medium–high abundance and diversity. Commonly, these ichnofossils occur in the upper part of substrata, that is the depth of bioturbation is less than 15 cm under the bedding plane. In this ichnofabric, intensity of bioturbation is medium or high (BI = 3–5), and the host rocks are mainly micrite and silt-crystal limestone.
Ichnofossil Zoophycos displays larger in individual size, its slablike spreiten is more than 4 mm wide and 30 cm long observed, extending paralleled to the bedding plane on the outcrop section. Its burrow filling is red (Figure ) or off-white (Figure ), and the red filling tested by energy disperse spectroscopy shows abundant iron (Fe2O3), indicating that these Zoophycos generated in oxygen-rich substrata, that is bottom water and pore water in the substrate were all oxygen-rich.
Based on the above-mentioned characteristics, this ichnofabric was explained to form in the upper shallow-marine environment between mean low tide level and fair-weather wave base, the sedimentary environment is relatively calm with shallow water.
4.3. Medium tiering ichnofabric
This ichnofabric is marked by fodinichnia in substrata, typically, the medium-sized Zoophycos associated with Teichichnus and Taenidium (Figure ); however, sometimes, only domichnia Skolithos and Thalassinoides found in some limestones. These are all the common ichnofossils of Cruziana ichnofacies in the Late Palaeozoic shallow-marine deposits, with medium–high abundance and diversity. These burrows penetrate from bedding plane to the middle part of substrata, usually, the depth of bioturbation is 15–25 cm under the bedding plane, showing medium or high intensity of bioturbation (BI = 3–5, Figure ), their host rock is mostly micrite. Ichnofossil Zoophycos is medium in individual size and slablike spreiten burrows on the outcrop longitudinal section are 2–3 mm wide and less than 30 cm long, extending almost paralleled to the bedding plane. Their burrow fillings are grey to dark grey (Figure ), indicating these ichnofossils developed in oxygenous or oxygen-low substratum condition.
Compared with Zoophycos in the above shallow tiering ichnofabric, the medium-sized Zoophycos in this ichnofabric shows smaller slablike spreiten and indicates that substrata containing the Zoophycos are better cohesiveness of carbonate sediment and the substrata are unsuitable to trace makers to build bigger burrows. Sometimes, vertical burrow Skolithos and other irregular near-vertical burrows are developed only in some limestone layers, showing that they formed in the shoal with higher energy in the shallow-marine carbonate platforms. In addition, some piles of fasciculate coral fossils are associated with these ichnofossils in same host rock, indicating that the sea water is clear and the sedimentary area is far from the coast.
According to the analysis of above characteristic, it may be considered that this ichnofabric forms in middle-upper shallow sea between mean low tide level and storm wave base, with a certain water depth change.
4.4. Deep tiering ichnofabric
The ichnofabric is mainly composed of many small- and medium-sized Zoophycos with dark-grey or black filling, and commonly associating with Chondrites and Nereites (Figure ). These ichnofossils, with high abundance and low diversity, mainly are fodinichnia in substrata. Their host rock is grey-dark grey, thin to medium thick-bedded microcrystalline limestone with more organic matter. The burrows in the ichnofabric penetrate from bedding plane to the middle or lower part of substrata, commonly, the depth of bioturbation is 25–35 cm under the bedding plane, and intensity of bioturbation is medium or high (BI = 2–5).
In this ichnofabric, ichnofossil Zoophycos is small and medium in individual size, showing two morphological characteristics on the outcrop section. Slablike spreiten burrow of small-sized Zoophycos with dark grey filling is 2–3 mm wide (Figure ), while the burrow with black filling is 3–4 mm wide (Figure ), and both the lateral extended length is bigger than 15 cm. Microscopic analysis indicates that the dominant composition of black filling is black mudstone containing more organic matter and silicious component. Through comparative analysis and combined with the relevant data (Frey & Howard, Citation1985), it may be suggested that this kind of Zoophycos is connected with low energy and poor oxygen environment. Nereites and Chondrites are the most common traces generated in the oxygen-poor substrata and generally present in the bottom tier bed. In the geological history records, Chondrites commonly appeared in black carbonaceous shale or argillaceous rock containing only few other ichnofossils. The characteristic can also prove that trace maker of Chondrites can tolerate the oxygen-low environment. Some scholars considered Chondrites, Zoophycos and Nereites with other tiny tubular burrows are the indicating mark of oxygen-poor environment (Ekdale & Mason, Citation1988; Jin et al., Citation2003). Therefore, it may be suggested that the deep tiering ichnofabric formed in a kind of substrata connected with oxygen-low or oxygen-poor environment, and so, this ichnofabric can be explained to form in the lower zone of shallow sea far from shore, the deposit substrata located between the storm wave base and the near redox interface.
5. Sedimentary environment model of ichnofabric types
The past studies of sedimentology and sequence stratigraphy had demonstrated that the limestones of the Taiyuan Formation in North China Basin formed in the epicontinental sea carbonate platform sedimentary environment (Chen, Xu, & Liu, Citation2008). On basis of above analysis in the composition, occurrence and sedimentary characteristics of ichnofabrics, specially, variation of the individual size, buried depth and the filling colour diversities of the Zoophycos burrows, it may be suggested that the four types of ichnofabrics generated, respectively, in different sedimentary environments of the epicontinental sea carbonate platform, including the gulf, lagoon, restricted platform, shoal in platform and open platform (Figure ). (1) Epigenic tiering ichnofabric mainly appeared in restricted bay or lagoon environment of low energy hydrodynamic conditions (Figure (A)); (2) shallow tiering ichnofabric occurred in the upper shallow-marine environment between mean low tide level and storm wave base, which is relatively calm shallow sea (Figure (B)); (3) medium tiering ichnofabric generated in the middle-upper shallow sea (restricted–open platform) between mean low tide level and storm wave base (Figure (C)); (4) deep tiering ichnofabric distributed in lower shallow sea (open platform) located under the storm wave base and the near redox interface (Figure (D)).
In addition, the substrate oxygen content has close relationship with water depth, hydrodynamic conditions, restricted and open degree of sedimentary environments and organic matter contents, and different trace maker has different tolerance towards the oxygen content; therefore, the oxygen content changes of above four types of ichnofabrics (from epigenic to deep tiers) are in accordance with ranging from the aerobic to anoxic conditions (Figure (A)–(D)). On these grounds, it may be deduced that the vertical variation trend of oxygen content in all beds of limestones of the Lower Permian Taiyuan Formation in North China Basin.
6. Conclusion
On basis of the detail study of ichnofossils and ichnofabrics in carbonate rocks of the Lower Permian Taiyuan Formation in North China Basin, the main results are obtained as follows:
| (1) | Thirteen ichnogenera and seventeen ichnospecies were recognised and descripted in the Lower Permian Taiyuan Formation of this study area and are common composition in the carbonate platform deposits of epicontinental sea. Common ichnofossils are attributed to fodinichnia, pascichnia and domichnia, mainly including Zoophycos, Teichichnus, Rhizocorallium, Chondrites, Palaeophycus, Planolites, Taenidium, Gordia, Helminthopsis, Nereites, Thalassinoides, Ophiomorpha and Skolithos, and other dichotomy traces. | ||||
| (2) | Four types of ichnofabrics were recognised in the carbonate rocks of the Taiyuan Formation, including epigenic tiering ichnofabric mainly appeared in restricted bay or lagoon sedimentary environments, shallow tiering ichnofabric occurred in the upper shallow-marine environment, medium tiering ichnofabric formed in the mid-upper shallow sea (restricted–open platform) and deep tiering ichnofabric distributed in lower shallow sea (open platform) far from shore. | ||||
| (3) | Based on the occurrence characteristics of ichnofossils, the vertical tier change characteristics of the ichnofabric types are summarised, i.e. epigenic tiering ichnofabric is mainly developed on or near the bedding plane, shallow tiering ichnofabric is produced in the top of deposit substrata within depth of 15 cm under the bedding plane, medium tiering ichnofabric are formed in the middle-upper part of deposit substrata within depth of 15–25 cm under the bedding plane, and the deep tiering ichnofabric penetrates down deeper substrata within depth of 25–35 cm under the bedding plane. At the same time, these ichnofabric types showed the change rule from oxygen to hypoxia, which is in accordance with ranging from the epigenic tiering to deep tiering ichnofabric types. | ||||
| (4) | According to the change law of the composition, occurrence of ichnofossils and associated with sedimentary characteristics, the sedimentary environment model of ichnofabric types is proposed (Figure ). | ||||
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Aceñolaza, F. G., & Buatois, L. (1993). Nonmarine perigondwanic trace fossils from the late Paleozoic of Argentina. Ichnos, 2, 183–201.10.1080/10420949309380092
- Bromley, R. G. (1991). Zoophycos: Strip mine, refuse dump, cache or sewage farm?. Lethaia, 24, 460–462.10.1111/let.1991.24.issue-4
- Bromley, R. G. (1996). Trace fossils: Biology, taphonomy and applications (2nd ed., 361pp). London: Chapman and Hall.10.1007/978-1-4899-2875-7
- Bromley, R. G., & Ekdale, A. A. (1984). Trace fossil preservation in Flint in the European Chalk. Journal of Paleontology, 58, 298–311.
- Buckman, J. O. (2001). Parataenidium, a new Taenidium‐like ichnogenus from the carboniferous of Ireland. Ichnos, 8, 83–97.10.1080/10420940109380177
- Chamberlain, C. K. (1977). Ordovician and Devonian trace fossils from Nevada. Nevada Bureau of Mines and Geology Bulletin, 90, 1–24.
- Chen, S., Xu, F., & Liu, H. 2008. The late paleozoic sequence stratigraphy and coal accumulation law in North China (pp. 18–23). Shandong: Petroleum University Press ( in Chinese).
- Chisholm, J. I. (1970). Teichichnus and related trace-fossils in the lower carboniferous at St. Monance, Scotland. Bulletion of the Geological Survey of Great Britain, 23, 21–51.
- Corner, G. D., & Fjalstad, A. (1993). Spreite trace fossils (Teichichnus) in a raised Holocene fjord‐delta, Breidvikeidet, Norway. Ichnos, 2, 155–164.10.1080/10420949309380085
- Crimes, T. P., & Anderson, M. M. (1985). Trace fossils from late precambrian-early cambrian strata of Southeastern Newfoundland (Canada): Temporal and environmental implications. Journal of Paleontology, 59, 310–343.
- Cui, Z. (1999). The environmental implication of the trace fossil Ophiomorpha from the upper permian of the Yily Basin, Xinjiang. Acta Sedimentologica Sinica, 17, 209–213 (in Chinese).
- Ekdale, A. A. (1992). Muckraking and mudslinging: The joys of deposit-feeding. In C. G. Maples & R. R. West (Eds.), Trace fossils, short courses in paleontology (Vol. 5, pp. 143–171). Knoxville, TN: Paleontological Society.
- Ekdale, A. A., & Bromley, R. G. (2003). Paleoethologic interpretation of complex Thalassinoides in shallow-marine limestones, Lower Ordovician, southern Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology, 192, 221–227.10.1016/S0031-0182(02)00686-7
- Ekdale, A. A., & Mason, T. R. (1988). Characteristic trace-fossil associations in oxygen-poor sedimentary environments. Geology, 16, 720–723.10.1130/0091-7613(1988)016<0720:CTFAIO>2.3.CO;2
- Fitzgerald, P. G., & Barrett, P. J. (1986). Skolithos in a Permian braided river deposit, southern Victoria Land, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology, 52, 237–247.10.1016/0031-0182(86)90049-0
- Frey, R. W., & Howard, J. D. (1985). Trace fossils from the Panther Member, Star Point Formation (Upper Cretaceous), Coal Creek Canyon, Utah. Journal of Paleontology, 59, 370–404.
- Gong, Y. (1994). Association relationships among the devonian trace fossils in Northern Xinjiang and their paleoenvironmental and palaeoecological significances. Geoscience, 2, 16–27 (in Chinese).
- Gong, Y. (2004). Facies characteristics and tiering distributions of Chondrites. Acta Palaeontologica Sinica, 43, 94–10 (in Chinese).
- Gong, Y. M., Shi, G. R., Weldon, E. A., Du, Y. S., & Xu, R. (2008). Pyrite framboids interpreted as microbial colonies within the Permian Zoophycos spreiten from southeastern Australia. Geological Magazine, 145, 95–103.
- Gong, Y. M., Xu, R., Xie, S. C., Huang, X. Y., Hu, B., Qi, Y. A., & Zhang, G. C. (2007). Microbial and molecular fossils from the Permian Zoophycos in South China. Science in China Series D: Earth Sciences, 50, 1121–1127.10.1007/s11430-007-0080-2
- Gong, Y., & Si, Y. (2002). Classification and evolution of metazoan traces at a topological level. Lethaia, 35, 263–274.
- Hu, B., Hu, L., Song, H. B., & Yang, L. C. (2013). Ichnoassemblages and their sedimentary environments in limestones of the upper carboniferous-lower permian Taiyuan Formation, Southeastern Shanxi Province. Journal of Paleogeography, 15, 809–818 (in Chinese).
- Hu, B., & Qi, Y. 2000. Zoophycos ichnofabric in limestones of the upper carboniferous Taiyuan Formation North China. Coal Geology & Exploration, 28, 12–15 ( in Chinese).
- Hu, B., Zhou, F., & Song, H. B. (2010). Ichnofossils and their sedimentary environments in the Lower Permian Taiyuan Formation in Jiaozuo area, Henan Province. Journal of Paleogeography, 12, 577–588 (in Chinese).
- Hu, Y. (1985). A type of rare trace fossils-Arenicola coprolites in the Taiyuan Formation (C3), Western Henan Province. Earth Science, 10, 21–26 (in Chinese).
- Jin, H., Li, Y., & Fang, G. (2003). Oxygen-dependent Ichnocoenose in Paleo-pelagic Substrata. Acta Sedimentologica Sinica, 21, 75–79 (in Chinese).
- Knaust, D. (2004). The oldest mesozoic nearshore Zoophycos: Evidence from the German Triassic. Lethaia, 37, 297–306.10.1080/00241160410002045
- Knaust, D. (2008). Complex behavioural pattern as an aid to identify the producer of Zoophycos from the middle Permian of Oman. Lethaia, 42, 146–154.
- Knaust, D. (2013). The ichnogenus Rhizocorallium: Classification, trace makers, palaeoenvironments and evolution. Earth-Science Reviews, 126, 1–47.10.1016/j.earscirev.2013.04.007
- Knaust, D., & Bromley, R. G. (Eds.). 2012. Trace fossils as indicators of sedimentary environments ( 924pp). Amsterdam: Elsevier.
- Loughlin, N. J. D., & Hillier, R. D. (2010). Early cambrian Teichichnus-dominated ichnofabrics and palaeoenvironmental analysis of the Caerfai Group, Southwest Wales, UK. Palaeogeography, Palaeoclimatology, Palaeoecology, 297, 239–251.10.1016/j.palaeo.2010.07.030
- Massalongo, A. (1855). Zoophycos, novum genus Plantarum fossilium (pp. 45–52). Typis Antonellianis, Verona: typis Antonellianis.
- McCann, T. A. (1993). Nereites ichnofacies from the Ordovician‐Silurian Welsh Basin. Ichnos, 3, 39–56.10.1080/10420949309386372
- Melchor, R. N., Bedatou, E., de Valais, S., & Genis, J. F. (2006). Lithofacies distribution of invertebrate and vertebrate trace-fossil assemblages in an early mesozoic ephemeral fluvio-lacustrine system from Argentina: Implications for the Scoyenia ichnofacies. Palaeogeography, Palaeoclimatology, Palaeoecology, 239, 253–285.10.1016/j.palaeo.2006.01.011
- Miller, M. F., & Curran, H. A. (2001). Behavioral plasticity of modern and Cenozoic burrowing thalassinidean shrimp. Palaeogeography, Palaeoclimatology, Palaeoecology, 166, 219–236.10.1016/S0031-0182(00)00210-8
- Miller, W. III. (2001). Thalassinoides-Phycodes compound burrow systems in Paleocene deep-water limestone, Southern Alps of Italy. Palaeogeography, Palaeoclimatology, Palaeoecology, 170, 149–156.10.1016/S0031-0182(01)00234-6
- Olivero, D. (2003). Early jurassic to late cretaceous evolution of Zoophycos in the French Subalpine Basin (southeastern France). Palaeogeography, Palaeoclimatology, Palaeoecology, 192, 59–78.10.1016/S0031-0182(02)00679-X
- Olivero, D. (2007). Zoophycos and the role of type specimens in ichnotaxonomy. In W. Miller Ш (Ed.), Trace fossils concepts, problems, prospects (pp. 219–231). Amsterdam: Elsevier.10.1016/B978-044452949-7/50139-X
- Parcha, S. K., & Pandey, S. (2011). Ichnofossils and their significance in the Cambrian successions of the Parahio Valley in the Spiti Basin, Tethys Himalaya, India. Journal of Asian Earth Sciences, 42, 1097–1116.10.1016/j.jseaes.2011.04.028
- Pemberton, S. G., & Frey, R. W. (1982). Trace fossil nomenclature and the Planolites-Palaeophycus dilemma. Journal of Paleontology, 56, 843–881.
- Sarkar, S., Sanjoy Kumar, G., & Chandan, C. (2009). Ichnology of a late palaeozoic ice-marginal shallow marine succession: Talchir Formation, Satpura Gondwana basin, central India. Palaeogeography, Palaeoclimatology, Palaeoecology, 283, 28–45.10.1016/j.palaeo.2009.09.001
- Savrda, C. E. (2000). Taenidium and associated ichnofossils in fluvial deposits, Cretaceous Tuscaloosa Formation, Eastern Alabama, Southeastern U.S.A. Ichnos, 7, 227–242.10.1080/10420940009380162
- Seilacher, A. (1967). Bathymetry of trace fossils. Marine Geology, 5, 413–428.10.1016/0025-3227(67)90051-5
- Seilacher A. (1983). Paleozoic sandstones in southern Jordan: Trace fossils, depositional environments and biogeography. In A. M. Abedm & H. M. Khafed (Eds.), Geology of Jordan, Proceedings of the First Jordean Geological Conference of the Jordanian Geological Association (pp. 209–222). Jordan: Cruziana ichnostratigraphy.
- Seilacher, A. (2007). Trace fossil analysis (pp. 1–226). Berlin: Springer-Verlag.
- Shang, G. (1995). An outline of basining structures of North China late palaeozoic coal accumulation basin. Coal Geology of China, 7(2), 1–7 (in Chinese).
- Sharafi, M., Ashuri, M., Mahboubi, A., & Moussavi-Harami, R. (2012). Stratigraphic application of Thalassinoides ichnofabric in delineating sequence stratigraphic surfaces (Mid-Cretaceous), Kopet-Dagh Basin, northeastern Iran. Palaeoworld, 21, 202–216.10.1016/j.palwor.2012.06.001
- Song, H. B., Jin, Y., Hu, L., & Hu, B. (2012). Ichnofabrics and their sedimentary environment of the lower permian Taiyuan Formation, Western Henan. Acta Geologica Sinica, 86, 972–984 (in Chinese).
- Uchman, A. (2007). Deep-sea trace fossils from the mixed carbonate-siliciclastic flysch of the Monte Antola Formation (Late Campanian-Maastrichtian), North Apennines, Italy. Cretaceous Research, 28, 980–1004.10.1016/j.cretres.2007.01.005
- Uchman, A. (2009). The Ophiomorpha rudis ichnosubfacies of the Nereites ichnofacies: Characteristics and constraints. Palaeogeography, Palaeoclimatology, Palaeoecology, 276, 107–119.10.1016/j.palaeo.2009.03.003
- Vanuxem, L. (1842). Geology of New York Part III survey of the third geological district. White and Visscher, 1–306.
- Villa, A. (1844). Memoria sulla costituzione geologica e geognostica della Brianza [Memory on geological constitution and surveys of Brianza]. Milano, 44, 1–46 ( in Italian).
- Wetzel, A. (1991). Ecologic interpretation of deep-sea trace fossil communities. Palaeogeography, Palaeoclimatology, Palaeoecology, 85, 47–69.10.1016/0031-0182(91)90025-M
- Wu, X. T., Hu, B., Wang, G. Z., & Zhang, G. C. (1987). Storm deposits in upper carboniferous shallow marine carbonates of Jiaozuo, Henan. Acta Sedimentologica Sinica, 5(4), 1–14 (in Chinese).
- Xia, B., Zhong, L., Fang, Z., et al. (1993). Discovery of Rhizocorallium in Wanghucun Formation in Xuancheng, Southern Anhui and its environment significance. Oil & Gas Geology, 14, 61–63 (in Chinese).
- Yang, S. (1984). The discovery of Zoophycos in the carboniferous of China and its environmental significance. Oil & Gas Geology, 5, 228–235 (in Chinese).
- Yang, S. F., Zhang, J. P., & Yang, M. F. 2004. Trace fossils in China (pp. 245–247). Beijing: Science Press (in Chinese).
- Zhu, S., & Li, G. 2005. A discovery of cf. Teichichnus rectusin in the upper Carboniferous strata in Xinmi coalfield. Oil Geology & Exploration, 33, 6–7 ( in Chinese).