
Abstract
People who are exposed to life-threatening trauma are at risk of developing posttraumatic stress disorder (PTSD). In addition to psychological manifestations, PTSD is associated with an increased risk of myocardial infarction, arrhythmias, hypertension, and other cardiovascular problems. We previously reported that rats exposed to a predator-based model of PTSD develop myocardial hypersensitivity to ischemic injury. This study characterized cardiac changes in histology and gene expression in rats exposed this model. Male rats were subjected to two cat exposures (separated by a period of 10 d) and daily cage-mate changes for 31 d. Control rats were not exposed to the cat or cage-mate changes. Ventricular tissue was analyzed by RNA sequencing, western blotting, histology, and immunohistochemistry. Multifocal lesions characterized by necrosis, mononuclear cell infiltration, and collagen deposition were observed in hearts from all stressed rats but none of the control rats. Gene expression analysis identified clusters of upregulated genes associated with endothelial to mesenchymal transition, endothelial migration, mesenchyme differentiation, and extracellular matrix remodeling in hearts from stressed rats. Consistent with endothelial to mesenchymal transition, rats from stressed hearts exhibited increased expression of α-smooth muscle actin (a myofibroblast marker) and a decrease in the number of CD31 positive endothelial cells. These data provide evidence that predator-based stress induces myocardial lesions and reprograming of cardiac gene expression. These changes may underlie the myocardial hypersensitivity to ischemia observed in these animals. This rat model may provide a useful tool for investigating the cardiac impact of PTSD and other forms of chronic psychological stress.
Lay summary
Chronic predator stress induces the formation of myocardial lesions characterized by necrosis, collagen deposition, and mononuclear cell infiltration. This is accompanied by changes in gene expression and histology that are indicative of cardiac remodeling. These changes may underlie the increased risk of arrhythmias, myocardial infarction, and other cardiac pathologies in people who have PTSD or other forms of chronic stress.
Introduction
Posttraumatic stress disorder (PTSD) is a disabling mental health disorder that develops following wartime combat, assault, rape, motor vehicle accidents, terrorist attacks, and other traumatic experiences. People with PTSD experience psychological distress by repeatedly reliving their trauma through intrusive flashback memories (Ehlers, Hackmann, & Michael, Citation2004; Ehlers et al., Citation2002; Reynolds & Brewin, Citation1999). This is accompanied by debilitating symptoms such as nightmares, emotional numbing, avoidance of stimuli associated with the trauma, and persistent arousal and hypervigilance (Nemeroff et al., Citation2006; Pitman et al., Citation2012; Zoladz & Diamond, Citation2013).
In addition to psychological manifestations, PTSD is also associated with cardiovascular abnormalities including hypertension, increased risk of cardiac arrhythmias, atherosclerosis, and increased risk of myocardial infarction (Ahmadi et al., Citation2011; Beristianos, Yaffe, Cohen, & Byers, Citation2016; Khazaie et al., Citation2013; Vaccarino et al., Citation2013). However, little is known about the mechanisms that underlie the increased risk of cardiovascular disease in PTSD patients. The direct impact of PTSD on the heart is difficult to study in humans because PTSD is associated with increased rates of smoking, sedentary lifestyle, diabetes, depression, and other factors that influence cardiovascular function and potentially confound efforts to assess the direct impact of posttraumatic stress on the heart (Dedert, Calhoun, Watkins, Sherwood, & Beckham, Citation2010; Zen, Whooley, Zhao, & Cohen, Citation2012). Animal models provide a tool to examine the impact of posttraumatic stress on the heart in the absence of these confounding variables.
Cat exposure is a well described and ethologically relevant stressor that produces an intense fear response in rats (Blanchard, Griebel, & Blanchard, Citation2003; Hubbard et al., Citation2004; Wilson, McLaughlin, Ebenezer, Nair, & Francis, Citation2014a; Wilson et al., Citation2013, Citation2014b). Our laboratory uses a cat-based model of psychosocial stress that incorporates features that are known to promote the development of PTSD including acute intense stress (predator exposure), lack of control over the stress (immobilization during predator exposure), a lack of social support (daily cage mate changes), and a re-experiencing of the stressful event (second predator exposure). Exposure to this animal model results in physiological and behavioral abnormalities that are remarkably similar to those observed in people with PTSD including a robust fear-conditioned memory of the trauma exposure, increased anxiety, exaggerated startle response, impaired memory for new information, enhanced negative feedback of the hypothalamic-pituitary-adrenal axis, decreased basal corticosterone, and increased hormonal and cardiovascular reactivity to an acute stressor (Zoladz, Conrad, Fleshner, & Diamond, Citation2008; Zoladz, Fleshner, & Diamond, Citation2012, Citation2013; Zoladz, Park, Fleshner, & Diamond, Citation2015). Some of these changes are reversed by drugs such as sertraline, clonidine, and tianeptine that have efficacy in the treatment of patients who have PTSD (Wilson et al., Citation2014a; Zoladz et al., Citation2013).
Multiple studies have demonstrated an increased risk of myocardial infarction and other cardiovascular disorders in people with PTSD (Ahmadi et al., Citation2011; Beristianos et al., Citation2016; Khazaie et al., Citation2013; Vaccarino et al., Citation2013). We previously reported that rats exposed to predator stress develop myocardial hypersensitivity to ischemic injury (Rorabaugh et al., Citation2015, Citation2019). The goal of this study was to use this stress model to characterize changes in myocardial histology and gene expression that may make the heart more vulnerable to myocardial infarction and other cardiac disorders. Our data provide evidence that predator-based psychosocial stress induces the formation of cardiac lesions characterized by necrosis, collagen deposition, infiltration of immune cells, and changes in gene expression that are indicative of cardiac tissue remodeling. These stress-induced changes may underlie the increased risk of cardiac disorders associated with PTSD and other chronic stress disorders.
Methods
Animals
Male Sprague-Dawley rats (6–7 weeks of age at the beginning of the study) from an established breeding colony at Ohio Northern University were used for all experiments. The rat colony originated from Sprague-Dawley rats obtained from Charles River Laboratories (Boston, MA; Strain Code 001). The rats were housed on a 12-h light/dark cycle (lights on at 07:00) in standard Plexiglas cages (two per cage) with free access to food and water. The animals originated from four different litters. Siblings were divided between stressed and non-stressed experimental groups to avoid the potential for litter bias. All procedures were approved by the Institutional Animal Care and Use Committee and were performed in compliance with the recommendations published in the eighth edition of The Guide for the Care and Use of Laboratory Animals.
Psychosocial stress procedure
Rats were randomly assigned to “psychosocial stress” or “no stress” experimental groups. The stress regimen has been previously described and validated by multiple laboratories (Rorabaugh et al., Citation2015; Zoladz et al., Citation2008, Citation2013, Citation2015). Rats in the psychosocial stress group were given two acute stress sessions (exposure to a cat), which were separated by 10 d. The first stress session took place during the light cycle (between 08:00 and 13:00), and the second stress session took place during the dark cycle (between 19:00 and 21:00) so that the animals could not predict when they might re-experience the predator exposure.
On Day 1, rats in the psychosocial stress groups were immobilized in plastic DecapiCones (Braintree Scientific; Braintree, MA) and placed in a perforated wedge-shaped Plexiglas enclosure (Braintree Scientific; Braintree, MA; 20 × 20 × 8 cm). This enclosure was then taken to a cat housing room and placed in a metal cage (61 × 53 × 51 cm) with an adult female cat for 1 h. The Plexiglas enclosure prevented any contact between the cat and rats, but the rats were still exposed to all non-tactile sensory stimuli associated with the cat. Canned cat food was smeared on top of the enclosure to direct the cat’s attention toward the rats. An hour later, the rats were returned to their home cages. Rats in the no stress groups remained in their home cages during the 1-h stress period.
Social stress
Rats in the psychosocial stress groups were exposed to unstable housing conditions starting on the day of the first cat exposure (Day 1) and continuing throughout the paradigm until behavioral testing (Day 32). Rats in the psychosocial stress group were housed two animals per cage, but every day their cage mates were changed. Therefore, no rats in the psychosocial stress groups had the same cage mate on two consecutive days during the 31-day stress period. Rats in the no stress groups were also housed two animals per cage. However, non-stressed animals were cohoused with the same cage mate throughout the experiment. Non-stressed animals were handled daily to control for handling effects on stressed animals.
Behavioral testing
Three weeks following the second stress session (Day 32), rats were tested on the elevated plus maze (EPM) to assess anxiety-like behavior. Rats were placed on the EPM for 5 min, and their behavior was videotaped by a JVC hard disk camera hanging above the EPM and scored offline by two separate investigators who were blind to the experimental conditions of the animals. Time spent in the open arms of the maze was used as a measure of anxiety, and the total number of closed arm entries was used to assess locomotor activity.
RNA-sequencing
Stressed and non-stressed rats (5 animals/group) were anesthetized with sodium pentobarbital (100 mg/kg i.p.), and hearts were quickly isolated and perfused for 5 min on a Langendorff isolated heart system to flush blood from the tissue. The left ventricle was immediately flash frozen in liquid nitrogen and stored at −80 °C until it was used for RNA isolation. Total RNA was isolated using TRIzol reagent (Fisher Scientific (Waltham, MA, USA) Catalog # 15596026) according to the manufacturer’s instructions. RNA was DNase I digested with a Turbo DNA-Free kit from Fisher Scientific (Waltham, MA, USA) (Catalog # AM1907) according to the manufacturer’s protocol. The RNA was shipped on dry ice to the Michigan State University Research Technology Support Facility Genomics Core (East Lansing, MI, USA). The RNA from each rat was quantified using a Quibit dsDNA High Sensitivity Assay (Thermo Fisher (Waltham, MA, USA) Cat #Q32851) and analyzed on an Agilent 2100 Bioanalyzer RNA Pico Chip (Santa Clara, CA, USA). RNA sequencing libraries were made from each rat heart using the Illumina TruSeq Stranded mRNA library preparation kit. Equimolar amounts of libraries were pooled for multiplexed sequencing on a single Illumina NextSeq 500 lane in a 75 base pair single end format. Sequencing data were demultiplexed and converted to FastQ format.
Raw Fastq files underwent quality control processing with FastQC software (Babraham Institute, Bahbraham, England) and were further processed with TrimGalore software (Bahbraham Institute, Bahbraham, England) to remove low-quality bases and to trim adapter sequences. Reads passing quality controls were aligned to the Rn6 version (Wellcome Trust Institute, Hinxton, England) of the rat genome using STAR aligner software (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY) (Dobin et al., Citation2013). Gene count files on the reverse strand were generated from alignment files with FeatureCounts (Liao, Smyth, & Shi, Citation2014) software (Walter and Eliza Hall Institute, Parksville, Australia) using the RNor_6.0 ENSEMBL genome assembly (Wellcome Sanger Institute, Hinxton, England). Differential gene expression analysis was produced in R version 3.3.3 software (Vienna, Austria) with the DESeq2 package (Chapel Hill, NC, USA) (Love, Huber, & Anders, Citation2014). Adjusted p values (false discovery rate) were calculated in DESeq2 using the Benjamini-Hochberg equation (Benjamini & Hochberg, Citation1995). Log2-transformed, median-centered gene expression and genes within the defined gene ontology families were selected for heatmap generation using “pheatmap” package in R. Volcano plots to visualize transcriptional changes were produced with GraphPad 7 software (GraphPad Software, La Jolla, CA). Genes considered significant (adjusted p value < .05) were analyzed by gene ontology in R with “ClusterProfiler” software (Yu, Wang, Han, & He, Citation2012).
Data availability statement
Raw data from next-generation sequencing experiments were deposited in the National Center for Biotechnology Information Sequence Read Archive and are available under BioProject ID number PRJNA498684. Differential gene expression analysis of these data is shown at https://figshare.com/s/5f32d3c03635c9bf46f5.
Western blots
Frozen ventricular tissue (from the same rats described above for RNA sequencing) was used for western blotting with antibodies for alpha smooth muscle actin (Cell Signaling Technology, Danvers, MA; Catalog #19245), T-cadherin (Abcam, Cambridge, MA; Cat #ab167407), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Cell Signaling Technology, Danvers, MA; Catalog #2118). Alpha smooth muscle actin and T-cadherin bands were normalized to those of GAPDH.
Histology
Stressed and non-stressed rats (6 animals/group) were euthanized with sodium pentobarbital (100 mg/kg, i.p.) followed by opening of the chest cavity. Hearts were removed, immersed in 10% neutral buffered formalin, and submitted to the Comparative Pathology and Mouse Phenotyping Shared Resource at The Ohio State University for tissue trimming, processing, and staining. Hearts were trimmed by longitudinal bisection perpendicular to the plane of the interventricular septum (Berridge et al., Citation2016). Two 5-µm thick sections of paraffin-embedded heart tissue were stained with hematoxylin and eosin (H&E) or Masson’s trichrome stains and evaluated microscopically by a board certified veterinary pathologist who was blinded to treatment groups. Histology slides were digitally scanned at Charles River Laboratories in Spencerville, OH using a Leica ScanScope XT scanner at 20X magnification and Aperio ImageScope version 12.3.0.5056 (Buffallo Grove, IL, United States) viewing software. Microscopic evaluation included scoring of the following criteria on a scale of 1–10, where 1 represented no visible lesions and 2 through 10 represented ascending levels of severity of myofiber necrosis (1–10), mononuclear cell infiltrate (1–10), or fibrosis (1–10).
Hearts from stressed and non-stressed rats (6 animals/group) were analyzed by immunohistochemistry using a CD31-specific antibody (Abcam, Cambridge, MA; Catalog # ab-28364). Five randomly chosen regions of the stained ventricular myocardium were photographed at 400X magnification and the number of CD31 positive cells in each field was averaged to determine the number of CD31 cells for each heart. This process was repeated for each heart by three independent investigators who were blinded to treatment groups. The mean value of the three investigators was used to represent the number of CD31 positive cells for each heart.
Statistical analyses
Data are presented as the mean ± SEM. The unpaired Student’s t test was used to compare data from stressed and non-stressed experimental groups in the EPM, western blots, histology, and immunohistochemistry. These analyses were performed using GraphPad Prism 7 software (GraphPad Software, La Jolla, CA). p Values < .05 were regarded as statistically significant. p Values for normalized counts from RNA sequencing were adjusted using R version 3.3.3 software with DESeq2 as previously described (Love et al., Citation2014; Benjamini & Hochberg, Citation1995) to minimize the frequency of false discovery. Adjusted p values < .05 were regarded as statistically significant.
Results
Anxiety-related behavior in the elevated plus maze
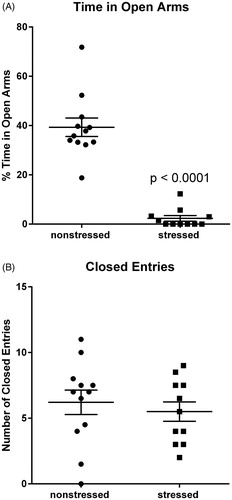
Previous work demonstrated that rats subjected to this stress model display anxiety-related behavior in the EPM (Rorabaugh et al., Citation2015; Zoladz et al., Citation2008, Citation2013). Consistent with previous studies, rats exposed to 31 d of psychosocial stress (n = 11) spent significantly (t(21) = 9.13, p < .0001) less time than non-stressed rats (n = 12) in the open arms of the maze (). The number of closed arm entries was similar (p=.47) for stressed and non-stressed rats (), indicating that stress had no significant effect on overall locomotor activity. These data provide behavioral validation of the efficacy of the stress regimen in rats that were subsequently used for cardiac analyses. Data from one stressed rat was excluded because the amount of time that it spent in the open arms of the EPM was more than five standard deviations from the mean.
Figure 1. Stressed rats exhibit anxiety-related behavior in the elevated plus maze. Rats exposed to 31 d of psychosocial stress spent significantly less time than non-stressed rats in the open arms of the elevated plus maze (A). Stressed and non-stressed rats had similar numbers of entries into the closed arms, indicating that stress had no significant effect on locomotor activity (B). Data represent the mean ± SEM of 11–12 animals per group.
Body weight
Body weights of control and stressed rats were similar on Day 1 of the stress protocol (278 ± 19 g and 290 ± 14 g for control and stressed animals, respectively; p=.61), and both experimental groups gained similar amounts of weight over the course of the 31-d stress regimen (187 ± 12 g and 185 ± 9 g; p=.92).
Stress-induced changes in cardiac gene expression
RNA sequencing analysis was performed using RNA isolated from the left ventricular tissue from five stressed and five non-stressed rats. Principal component analysis (PCA) indicated that ventricles from non-stressed rats had variable baseline gene expression patterns that separated on principal component 1 (PC1), while ventricular tissue from all 5 stressed rats clustered closely and were separated by principal component 2 (PC2) (). These data indicate that stress-induced distinct and reproducible transcriptional changes in rat ventricles.
Figure 2. RNA-sequencing of ventricular tissue from stressed and non-stressed rats. Principal component analysis of gene expression profiles indicated that hearts from non-stressed rats have variable baseline gene expression that separated on principal component 1 (PC1), while hearts from all five stressed rats clustered closely and were separated by principal component 2 (PC2) (A). A volcano plot illustrating gene expression alterations and p values for 12,218 genes. Genes of interest are annotated in red (B). Gene ontology analysis identified significantly enriched gene family clusters for upregulated (red) or downregulated (black) genes (C). Genes were ranked in order of most to least enriched in stressed hearts and analyzed by Geneset Enrichment Analysis (GSEA). The enrichment plot shows the normalized enrichment score (NES) for genes associated with endothelial-to-mesenchymal transition (D). Heatmap showing median-centered, log2-fold changes for select upregulated genes identified from Gene Ontology families in panel C (E).
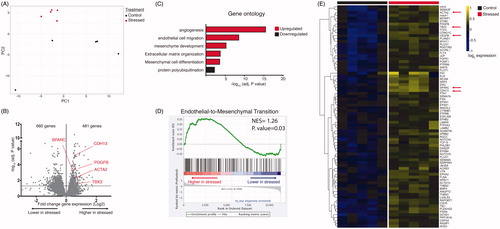
In light of our finding that stressed hearts undergo transcriptional alterations, we next sought to define these transcriptional programs. Out of 12,218 detected genes, 481 genes were significantly (adjusted p value < .05) upregulated and 660 genes were significantly downregulated (; the differential gene expression analysis for the entire data set is available at https://figshare.com/s/5f32d3c03635c9bf46f5). Gene ontology analysis indicated that upregulated genes were primarily associated with angiogenesis (GO:0001525) and endothelial cell migration (GO:0043542) (, ), mesenchyme development (GO:0060485) and mesenchymal differentiation (GO:0048762) (, ), and extracellular matrix organization (GO:0030198) (, ). Interestingly, downregulated genes were only associated with one gene cluster (protein polyubiquitination; GO:0000209) (), suggesting that only upregulated genes fall within defined biologically relevant processes. Additionally, we ranked genes in order from increasing to decreasing log2-fold expression changes in stressed hearts and performed gene set enrichment analysis (GSEA) to identify pathways associated with stress. We found that “Endothelial to Mesenchymal Transition” was among the top enriched gene sets (Normalized Enrichment Score = 1.26, p=.03) (). Thus both analyses (gene ontology and GESA) are consistent with increased formation of mesenchymal tissue. These data suggest that stressed hearts may have an increase in fibrotic tissue. Notably, many genes that were significantly upregulated have been associated with cardiac fibrosis, including PDGFB (Gallini, Lindblom, Bondjers, Betsholtz, & Andrae, Citation2016), TBX2 (Shirai, Imanaka-Yoshida, Schneider, Schwartz, & Morisaki, Citation2009), SPARC (Vaughan, Marley, Miellet, & Hartley, Citation2018), and ACTA2 (Teekakirikul et al., Citation2010; Zeisberg et al., Citation2007) (). Differential gene expression analysis data for the entire data set is available at https://figshare.com/s/5f32d3c03635c9bf46f5.
Table 1. Selected changes in the number of mRNA transcripts (normalized counts) for genes in the angiogenesis (GO:0001525) and endothelial cell migration (GO:0043542) gene ontology clusters.
Table 2. Selected changes in the number of mRNA transcripts (normalized counts) for genes in the mesenchyme development (GO:0060485) and mesenchyme differentiation (GO:0048762) gene ontology clusters.
Table 3. Selected changes in the number of mRNA transcripts (normalized counts) for genes in the extracellular matrix assembly (GO:0085029) and extracellular matrix organization (GO:0030198) gene ontology clusters.
Stress-induced cardiac fibrosis and immune cell infiltration
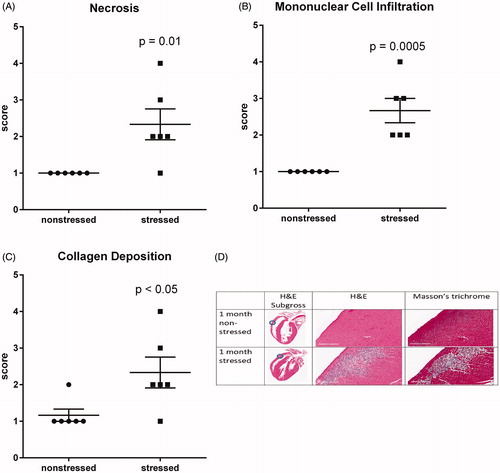
RNA sequencing indicated significant upregulation of genes involved in angiogenesis, cell migration, differentiation into mesenchymal tissue, and assembly/organization of extracellular matrix (, ). Thus, H&E and Masson’s trichrome staining were used to determine whether these changes in gene expression reflected histological changes in the myocardium. Multifocal lesions characterized by myofiber necrosis (), infiltration of mononuclear immune cells (), and collagen deposition () were identified in the ventricular walls of all (n = 6) stressed rats. These lesions were absent from the hearts of all (n = 6) non-stressed rats with the exception of 1 animal that exhibited minor fibrosis (score = 2) without necrosis or mononuclear cell infiltration.
Figure 3. Stress induces myocardial lesions characterized by fibrosis, necrosis, and mononuclear cell infiltration. Hearts from stressed and non-stressed rats were evaluated by H&E and Masson’s trichrome staining. Hearts from stressed rats developed multifocal lesions characterized by necrosis (A,D), infiltration by mononuclear immune cells (B,D), and collagen deposition (C,D). Data represent the mean ± SEM of six animals. Representative photographs of heart sections are shown in panel D. Blue circles in photographs of subgross heart images indicate heart regions that were photographed at higher (15.8X) magnification. Scale bars in panel D represent 200 µm.
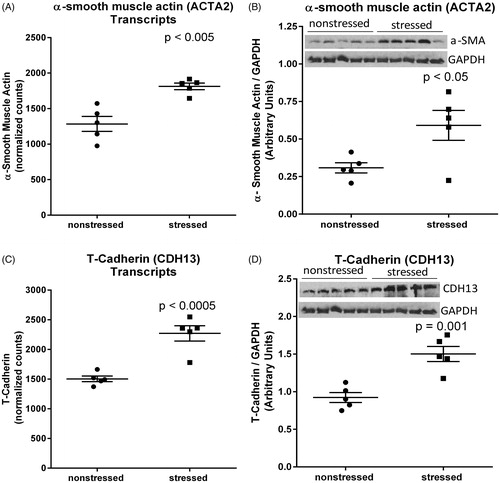
RNA sequencing and western blots indicated that α-smooth muscle actin was upregulated at both the mRNA transcript (t(8)=4.60, p < .005) () and protein (t(8)=2.69, p < .05) () levels in stressed heart. Increased expression of this myofibroblast marker is consistent with the observed deposition of collagen in stressed hearts () (van Putten, Shafieyan, & Hinz, Citation2016). We also observed that T-cadherin (CDH13) was significantly upregulated at the mRNA transcript (t(8)=5.56, p < .0005) () and protein (t(8)=4.80, p=.001) () levels in hearts from stressed rats. These stress-induced changes are consistent with fibrosis and extracellular matrix remodeling (Asada et al., Citation2007; Frismantiene et al., Citation2014).
Figure 4. Stress up-regulates myocardial expression of α-smooth muscle actin and T-cadherin. RNA sequencing identified upregulation of mRNA transcripts encoding α-smooth muscle actin (encoded by ACTA2) (A) and T-cadherin (encoded by CDH13) (C) in hearts from rats exposed to 31 d of psychosocial stress compared to hearts from non-stressed rats. Western blots confirmed significant increases in the expression of α-smooth muscle actin (B) and T-cadherin (D) at the protein level. Data in panels A and C represent the mean ± SEM of six hearts. Data in panels B and D represent the mean ± SEM of five hearts.
Impact of stress on CD31 positive cells
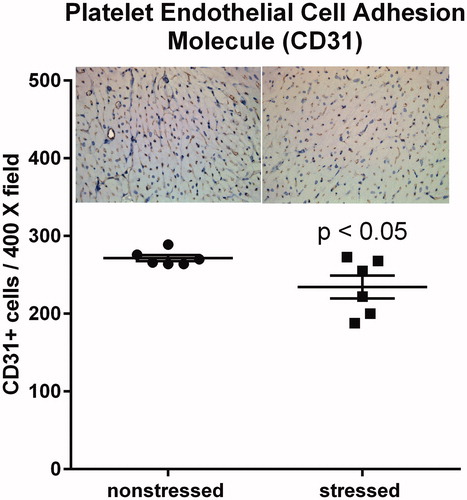
RNA sequencing indicated that hearts from stressed rats exhibited upregulation of genes involved in angiogenesis, blood vessel morphogenesis, and endothelial cell migration (). CD31 is commonly used as an endothelial marker (Lv et al., Citation2018; Tian et al., Citation2019; Wang et al., Citation2018). Thus, we compared the number of CD31 positive cells in high magnification (400X) images of ventricles from stressed and non-stressed hearts. Immunohistochemistry indicated a significant (t(10)=2.43, p < .05) decrease in the number of CD31 positive cells in ventricles from stressed rats compared to ventricles from non-stressed rats ().
Figure 5. Stressed hearts exhibit a decrease in the number of CD31 positive cells. Predator-based psychosocial stress decreased the number of CD31-positive cells in the ventricular myocardium. Ventricular tissue was stained with a CD31-specific antibody, and the number of CD31 positive cells were counted by in 5 high (400X) magnification fields for each heart by three independent investigators that were blinded to animal treatment. Data represent the mean ± SEM of six hearts for each group.
Discussion
PTSD and other forms of chronic psychosocial stress are associated with an increased risk of cardiovascular abnormalities including hypertension, endothelial dysfunction, atherosclerosis, cardiac arrhythmias, and myocardial infarction (Ahmadi et al., Citation2011; Beristianos et al., Citation2016; Grenon et al., Citation2016; Khazaie et al., Citation2013; Vaccarino et al., Citation2013). We previously reported that rats exposed to the predator-stress model used in this study develop myocardial hypersensitivity to ischemic injury (Rorabaugh et al., Citation2015, Citation2019). The present work demonstrated that these animals also develop multifocal cardiac lesions characterized by myofiber necrosis, fibrosis, and infiltration by mononucleated immune cells (). This was accompanied by changes in gene expression associated with endothelial cell migration, mesenchymal development, and extracellular matrix organization (, ). In addition, changes in the expression of markers for endothelial to mesenchymal transition were observed at both the mRNA () and protein levels ( and ). These data provide evidence of cardiac stress and tissue remodeling in rats exposed to predator stress. These changes in histology and gene expression may underlie our previous observation that rats exposed to this model develop myocardial hypersensitivity to ischemic injury (Rorabaugh et al., Citation2015, Citation2019).
Cho et al. previously used DNA microarray analysis to assess changes in gene expression in the mouse heart following 1–10 d of exposure to a social defeat model (Cho et al., Citation2014). Similar to this study, these investigators identified changes in gene expression that were indicative of extracellular matrix remodeling, angiogenesis, and the formation of mesenchymal tissue. This prior work differed from this study in fundamental ways including the species (mouse vs. rat), stress model (stress induced by an aggressor mouse vs. a predator), the duration of stress (1–10 d vs. 31 d), and the method of assessing gene expression (DNA microarray vs. RNA sequencing). Despite these differences, similar stress-induced changes in gene expression were observed, suggesting that increased expression of genes related to these ontologies represent a fundamental cardiac response to psychosocial stress.
Rats exposed to the psychosocial stress model used in this study exhibit significant increases in superoxide, peroxynitrite, and total reactive oxygen species in circulating blood, brain, and adrenal tissue (Ebenezer, Wilson, Wilson, Nair, & Francis, Citation2016; Wilson et al., Citation2013). These animals also exhibit increased expression of pro-inflammatory cytokines (interleukin-1β and interleukin-18), decreased expression of anti-inflammatory cytokines (interleukin-10 and interleukin-4), and increased expression of the NALP3 inflammasome in the brain (Wilson et al., Citation2013). However, these proinflammatory markers have not previously been assessed in hearts of animals exposed to this model. RNA sequencing did not identify significant changes in the presence of mRNA transcripts encoding pro-inflammatory cytokines in the heart. However, the fact that hearts from stressed rats were infiltrated with mononucleated immune cells provides evidence that this form of stress produces an inflammatory response in the heart.
The gene expression profile, increased secretion of collagen, increased expression of α-smooth muscle actin, and the decrease in the number of CD31 positive cell is consistent with endothelial to mesenchymal transition in hearts from stressed rats. In addition to these phenotypic markers, stressed hearts also exhibited upregulation of genes (Zeb1, bone morphogenetic protein-4, notch 1, and β-catenin) (Cho, Lee, Chang, Lee & Kim, Citation2018) that are known to mediate endothelial to mesenchymal transition (). However, we did not trace fibroblast lineage to determine whether stressed hearts contain an increase in the number of fibroblasts of endothelial origin. Thus, we cannot exclude the possibility that the decrease in endothelial cells and the increase in mesenchymal markers (collagen and α-smooth muscle actin) occurred independent of each other rather than through the process of endothelial to mesenchymal transition.
T-cadherin was upregulated in stressed hearts at both the mRNA transcript and protein levels. Previous work demonstrated that T-cadherin is involved in the organization of collagen fibrils in vitro (Frismantiene et al., Citation2014) and that it promotes remodeling of the vascular wall following vascular injury (Kudrjashova et al., Citation2002). Asada et al. (Citation2007) reported that T-cadherin was upregulated in a rat model of liver fibrosis. Thus, it is not surprising that T-cadherin was upregulated in the heart under conditions that promote collagen deposition and fibrosis. However, we are unaware of any previous studies in which T-cadherin has been associated with extracellular matrix remodeling or fibrosis in the heart. Further work is needed to better understand the role of T-cadherin in myocardial fibrosis.
The observed decrease in the number of CD31 positive cells in stressed hearts () seemed contradictory to the outcome of gene ontology analysis () which suggested that stressed hearts exhibit a gene expression profile indicative of increased angiogenesis. However, it should be noted that there is overlap in the content of these gene clusters. Several genes that were significantly upregulated (FLNA, NRP2, BMP4, FGFR1, and EFNA1) at the mRNA level are part of both the angiogenesis () and mesenchyme development gene clusters (). We have performed western blots on ventricular homogenates from both stressed and non-stressed rats for a number of proteins that are associated with angiogenesis including endothelial nitric oxide synthase, vascular endothelial growth factor receptor 3, and vascular cell adhesion molecule 1 (data not shown) and found no stress-induced effect on the expression of these angiogenesis markers at the protein level. These data suggest that overlap between gene clusters may have caused gene ontology analysis to falsely identify myocardial angiogenesis as a stress-induced response in these animals.
PTSD is associated with an increased risk of developing cardiac disorders (Ahmadi et al., Citation2011; Beristianos et al., Citation2016; Khazaie et al., Citation2013; Vaccarino et al., Citation2013). The identification of pharmacotherapies that can prevent the development of cardiac disorders in patients with PTSD or other forms of chronic psychological stress would be of great clinical value. Increased sympathetic signaling is a hallmark feature of PTSD, and sympatholytics such as clonidine and propranolol have demonstrated efficacy in treating trauma-related nightmares, intrusive thoughts, and hyperarousal in some patients with PTSD (Alao, Selvarajah, & Razi, Citation2012; Belkin & Schwartz, Citation2015; Boehnlein & Kinzie, Citation2007; Kinzie & Leung, Citation1989; Lindgren et al., Citation2013; Onder, Tural, & Aker, Citation2006). Clonidine prevents the development of some of the PTSD-like physiological (increased systolic and diastolic blood pressure, tachycardia, and adrenal hypertrophy) and behavioral (anxiety in EPM and increased startle response) effects induced by this animal model (Zoladz et al., Citation2013). However, neither clonidine nor propranolol prevent the development of myocardial hypersensitivity to ischemia in this rat model (Rorabaugh et al., Citation2019), indicating that this stress-induced effect is not the result of excessive sympathetic signaling. The current data suggest that anti-inflammatory agents or pharmacotherapies that inhibit endothelial to mesenchymal transition and the deposition of collagen may be more effective approaches to protecting the heart from the adverse effects of psychosocial stress.
In conclusion, our data indicate that predator stress promotes myocardial necrosis, collagen deposition, infiltration of immune cells, and reprograming of myocardial gene expression. These effects are consistent with the observation that animals exposed to this model develop myocardial hypersensitivity to ischemic injury (Rorabaugh et al., Citation2015, Citation2019). They are also consistent with previous work demonstrating that people who develop PTSD and other forms of chronic stress are at increased risk of developing cardiac disorders such as myocardial infarction and arrhythmias (Ahmadi et al., Citation2011; Beristianos et al., Citation2016; Khazaie et al., Citation2013; Vaccarino et al., Citation2013). This predator-based model of psychosocial stress may provide a useful tool to understand the mechanism by which PTSD impacts the heart and to identify therapeutic strategies to prevent the development of heart diseases in people who suffer from PTSD and other chronic stress disorders.
Acknowledgments
The authors thank Dr. Kevin Childs (Michigan State University Research Technology Support Facility Genomics Core) for help with the RNA sequencing experiments and Dr. Krista LePerle (The Ohio State University Comparative Pathology and Mouse Phenotyping core facility) for preparing hearts for histological analysis.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Notes on contributors
Boyd R. Rorabaugh
Boyd R. Rorabaugh is Professor of Pharmacology and Cell Biology at the Ohio Northern University College of Pharmacy. He contributed his expertise in the impact of predator stress on the heart and also served as PI of this study.
Nathaniel W. Mabe
Nathaniel W. Mabe was a postdoctoral fellow in the Department of Pharmacology and Cancer Biology at Duke University when the mansucrtipt was submitted. He is now a postdoctoral fellow at Harvard University’s Dana Farber Cancer Institute. He contributed his expertise in genomics and gene expression analysis.
Sarah L. Seeley
Sarah L. Seeley is a research technician in Dr. Rorabaugh’s laboratory. She contributed to the evaluation and analysis of histology data in this study.
Thorne S. Stoops
Thorne S. Stoops is a Pharm.D. student at the Ohio Northern University School of Pharmacy. He contributed to the evaulation and analysis of histology data.
Kasey E. Mucher
Kasey E. Mucher is a Pharm.D. student at the Ohio Northern University College of Pharmacy. She was a member of a team of students that performed the chronic stress manipulations.
Connor P. Ney
Connor P. Ney is an undergraduate student majoring in Biology at Ohio Northern University. He was a member of a team of students who performed the chronic stress manipulations.
Cassandra S. Goodman
Cassandra S. Goodman is a Pharm.D. student at Ohio Northern University. She was a member of a team of students that performed the chronic stress manipulations in this study.
Brooke J. Hertenstein
Brook J. Hertenstein is an undergraduate student majoring in Biology at Ohio Northern University. She was a member of a team of students that conducted elevated plus maze experiments.
Austen E. Rush
Austen Rush is an undergraduate student majoring in Biology at Ohio Northern University. He was a member of a team of students that conducted elevated plus maze experiments.
Charis D. Kasler
Charis D. Kasler is an undergraduate student majoring in Chemistry at Ohio Northern University. She was a member of a team of students that conducted elevated plus maze experiments.
Aaron M. Sargeant
Aaron M. Sargeant is a board certified veterinary pathologist at Charles River Laboratories. He performed histological analyses of myocardial tissue in this study and contributed his expertise in myocardial pathology.
Phillip R. Zoladz
Phillip R. Zoladz is Associate Professor of Psychology at Ohio Northern University.
Dr. Zoladz contributed his expertise in animal models of PTSD and stress and also managed the team of undergraduate students who performed the chronic stress maniulations in this study.
References
- Ahmadi, N., Hajsadeghi, F., Mirshkarlo, H.B., Budoff, M., Yehuda, R., & Ebrahimi, R. (2011). Post-traumatic stress disorder, coronary atherosclerosis, and mortality. The American Journal of Cardiology, 108, 29–33. doi:10.1016/j.amjcard.2011.02.340
- Alao, A., Selvarajah, J., & Razi, S. (2012). The use of clonidine in the treatment of nightmares among patients with co-morbid PTSD and traumatic brain injury. International Journal of Psychiatry in Medicine, 44, 165–169. doi:10.2190/PM.44.2.g
- Asada, K., Yoshiji, H., Noguchi, R., Ikenaka, Y., Kitade, M., Kaji, K., … Fukui, H. (2007). Crosstalk between high-molecular-weight adiponectin and t-cadherin during liver fibrosis development in rats. International Journal of Molecular Medicine, 20, 725–729. doi:10.3892/ijmm.20.5.725
- Belkin, M.R., & Schwartz, T.L. (2015). Alpha-2 receptor agonists for the treatment of posttraumatic stress disorder. Drugs in Context, 4, 212286. doi:10.7573/dic.212286
- Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. Series B (Methodological), 57, 289–300. doi:10.1111/j.2517-6161.1995.tb02031.x
- Beristianos, M.H., Yaffe, K., Cohen, B., & Byers, A.L. (2016). PTSD and risk of incident cardiovascular disease in aging veterans. The American Journal of Geriatric Psychiatry: Official Journal of the American Association for Geriatric Psychiatry, 24, 192–200. doi:10.1016/j.jagp.2014.12.003
- Berridge, B.R., Mowat, V., Nagai, H., Nyska, A., Okazaki, Y., Clements, P.J., … Wells, M.Y. (2016). Non-proliferative and proliferative lesions of the cardiovascular system of the rat and mouse. Journal of Toxicologic Pathology, 29, 1S–47S. doi:10.1293/tox.29.3S-1
- Blanchard, D.C., Griebel, G., & Blanchard, R.J. (2003). Conditioning and residual emotionality effects of predator stimuli: Some reflections on stress and emotion. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 27, 1177–1185. doi:10.1016/j.pnpbp.2003.09.012
- Boehnlein, J.K., & Kinzie, J.D. (2007). Pharmacologic reduction of CNS noradrenergic activity in PTSD: The case for clonidine and prazosin. Journal of Psychiatric Practice, 13, 72–78. doi:10.1097/01.pra.0000265763.79753.c1
- Cho, J.G., Lee, A., Chang, W., Lee, MS &Kim J. (2018). Endothelial to mesenchymal transition represents a key link in the interaction between inflammation and endothelial dysfunction. Frontiers in Immunology, 9, 294. doi:10.3389/fimmu.2018.00294
- Cho, J.H., Lee, I., Hammamieh, R., Wang, K., Baxter, D., Scherler, K., … Hood, L. (2014). Molecular evidence of stress-induced acute heart injury in a mouse model simulating posttraumatic stress disorder. Proceedings of the National Academy of Sciences of the United States of America, 111, 3188–3193. doi:10.1073/pnas.1400113111
- Dedert, E.A., Calhoun, P.S., Watkins, L.L., Sherwood, A., & Beckham, J.C. (2010). Posttraumatic stress disorder, cardiovascular, and metabolic disease: A review of the evidence. Annals of Behavioral Medicine:A Publication of the Society of Behavioral Medicine, 39, 61–78. doi:10.1007/s12160-010-9165-9
- Dobin, A., Davis, C.A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., … Gingeras, T.R. (2013). Star: Ultrafast universal RNA-seq aligner. Bioinformatics, 29, 15–21. doi:10.1093/bioinformatics/bts635
- Ebenezer, P.J., Wilson, C.B., Wilson, L.D., Nair, A.R., & Francis, J. (2016). The anti-inflammatory effects of blueberries in an animal model of post-traumatic stress disorder (PTSD). PLoS One, 11, e0160923. doi:10.1371/journal.pone.0160923
- Ehlers, A., Hackmann, A., & Michael, T. (2004). Intrusive re-experiencing in post-traumatic stress disorder: Phenomenology, theory, and therapy. Memory, 12, 403–415. doi:10.1080/09658210444000025
- Ehlers, A., Hackmann, A., Steil, R., Clohessy, S., Wenninger, K., & Winter, H. (2002). The nature of intrusive memories after trauma: The warning signal hypothesis. Behaviour Research and Therapy, 40, 995–1002. doi:10.1016/S0005-7967(01)00077-8
- Frismantiene, A., Pfaff, D., Frachet, A., Coen, M., Joshi, M.B., Maslova, K., … Philippova, M. (2014). Regulation of contractile signaling and matrix remodeling by t-cadherin in vascular smooth muscle cells: Constitutive and insulin-dependent effects. Cellular Signalling, 26, 1897–1908. doi:10.1016/j.cellsig.2014.05.001
- Gallini, R., Lindblom, P., Bondjers, C., Betsholtz, C., & Andrae, J. (2016). PDGF-A and PDGF-B induces cardiac fibrosis in transgenic mice. Experimental Cell Research, 349, 282–290. doi:10.1016/j.yexcr.2016.10.022
- Grenon, S.M., Owens, C.D., Alley, H., Perez, S., Whooley, M.A., Neylan, T.C., … Cohen, B.E. (2016). Posttraumatic stress disorder is associated with worse endothelial function among veterans. Journal of the American Heart Association, 5, e003010. doi:10.1161/JAHA.115.003010
- Hubbard, D.T., Blanchard, D.C., Yang, M., Markham, C.M., Gervacio, A., Chun-I, L., & Blanchard, R.J. (2004). Development of defensive behavior and conditioning to cat odor in the rat. Physiology and Behavior, 80, 525–530. doi:10.1016/j.physbeh.2003.10.006
- Khazaie, H., Saidi, M.R., Sepehry, A.A., Knight, D.C., Ahmadi, M., Najafi, F., … Tahmasian, M. (2013). Abnormal ECG patterns in chronic post-war PTSD patients: A pilot study. International Journal of Behavioral Medicine, 20, 1–6. doi:10.1007/s12529-011-9197-y
- Kinzie, J.D., & Leung, P. (1989). Clonidine in Cambodian patients with posttraumatic stress disorder. The Journal of Nervous and Mental Disease, 177, 546–550. doi:10.1097/00005053-198909000-00005
- Kudrjashova, E., Bashtrikov, P., Bochkov, V., Parfyonova, Y., Tkachuk, V., Antropova, J., … Resink, T.J. (2002). Expression of adhesion molecule t-cadherin is increased during neointima formation in experimental restenosis. Histochemistry and Cell Biology, 118, 281–290. doi:10.1007/s00418-002-0463-6
- Liao, Y., Smyth, G.K., & Shi, W. (2014). Featurecounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics, 30, 923. doi:10.1093/bioinformatics/btt656
- Lindgren, M.E., Fagundes, C.P., Alfano, C.M., Povoski, S.P., Agnese, D.M., Arnold, M.W., … Kiecolt-Glaser, J.K. (2013). Beta-blockers may reduce intrusive thoughts in newly diagnosed cancer patients. Psychooncology, 22, 1889–1894. doi:10.1002/pon.3233
- Love, M.I., Huber, W., & Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biology, 15, 550. doi:10.1186/s13059-014-0550-8
- Lv, Y.X., Zhong, S., Tang, H., Luo, B., Chen, S.J., Chen, L., … Tang, J.M. (2018). VEGF-A and VEGF-B coordinate the arteriogenesis to repair the infarcted heart with vagus nerve stimulation. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology, 48, 433–449. doi:10.1159/000491775
- Nemeroff, C.B., Bremner, J.D., Foa, E.B., Mayberg, H.S., North, C.S., & Stein, M.B. (2006). Posttraumatic stress disorder: A state-of-the-science review. Journal of Psychiatric Research, 40, 1–21. doi:10.1016/j.jpsychires.2005.07.005
- Onder, E., Tural, U., & Aker, T. (2006). A comparative study of fluoxetine, moclobemide, and tianeptine in the treatment of posttraumatic stress disorder following an earthquake. European Psychiatry: The Journal of the Association of European Psychiatrists, 21, 174–179. doi:10.1016/j.eurpsy.2005.03.007
- Pitman, R.K., Rasmusson, A.M., Koenen, K.C., Shin, L.M., Orr, S.P., Gilbertson, M.W., … Liberzon, I. (2012). Biological studies of post-traumatic stress disorder. Nature Reviews. Neuroscience, 13, 769–787. doi:10.1038/nrn3339
- Reynolds, M., & Brewin, C.R. (1999). Intrusive memories in depression and posttraumatic stress disorder. Behaviour Research and Therapy, 37, 201–215. doi:10.1016/S0005-7967(98)00132-6
- Rorabaugh, B.R., Bui, A.D., Seeley, S.L., Eisenmann, E.D., Rose, R.M., Johnson, B.L., … Zoladz, P.R. (2019). Myocardial hypersensitivity to ischemic injury is not reversed by clonidine or propranolol in a predator-based rat model of posttraumatic stress disorder. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 89, 117–124. doi:10.1016/j.pnpbp.2018.09.003
- Rorabaugh, B.R., Krivenko, A., Eisenmann, E.D., Bui, A.D., Seeley, S., Fry, M.E., … Zoladz, P.R. (2015). Sex-dependent effects of chronic psychosocial stress on myocardial sensitivity to ischemic injury. Stress (Amsterdam, Netherlands), 18, 645–653. doi:10.3109/10253890.2015.1087505
- Shirai, M., Imanaka-Yoshida, K., Schneider, M.D., Schwartz, R.J., & Morisaki, T. (2009). T-box 2, a mediator of Bmp-Smad signaling, induced hyaluronan synthase 2 and Tgfbeta2 expression and endocardial cushion formation. Proceedings of the National Academy of Sciences of the United States of America, 106, 18604–18609. doi:10.1073/pnas.0900635106
- Teekakirikul, P., Eminaga, S., Toka, O., Alcalai, R., Wang, L., Wakimoto, H., … Seidman, J.G. (2010). Cardiac fibrosis in mice with hypertrophic cardiomyopathy is mediated by non-myocyte proliferation and requires Tgf-β. The Journal of Clinical Investigation, 120, 3520–3529. doi:10.1172/JCI42028
- Tian, D., Zeng, X., Wang, W., Wang, Z., Zhang, Y., & Wang, Y. (2019). Protective effect of rapamycin on endothelial-to-mesenchymal transition in HUVECs through the Notch signaling pathway. Vascular Pharmacology, 113, 20–26. doi:10.1016/j.vph.2018.10.004
- Vaccarino, V., Goldberg, J., Rooks, C., Shah, A.J., Veledar, E., Faber, T.L., … Bremner, J.D. (2013). Post-traumatic stress disorder and incidence of coronary heart disease: A twin study. Journal of the American College of Cardiology, 62, 970–978. doi:10.1016/j.jacc.2013.04.085
- van Putten, S., Shafieyan, Y., & Hinz, B. (2016). Mechanical control of cardiac myofibroblasts. Journal of Molecular and Cellular Cardiology, 93, 133–142. doi:10.1016/j.yjmcc.2015.11.025
- Vaughan, L., Marley, R., Miellet, S., & Hartley, P.S. (2018). The impact of SPARC on age-related cardiac dysfunction and fibrosis in drosophila. Experimental Gerontology, 109, 59–66. doi:10.1016/j.exger.2017.10.011
- Wang, W., Wang, Z., Tian, D., Zeng, X., Liu, Y., Fu, Q., … Wang, Y. (2018). Integrin β3 mediates the endothelial-to-mesenchymal transition via the Notch pathway. Cellular Physiology and Biochemistry: International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology, 49, 985. doi:10.1159/000493229
- Wilson, C.B., McLaughlin, L.D., Ebenezer, P.J., Nair, A.R., & Francis, J. (2014a). Valproic acid effects in the hippocampus and prefrontal cortex in an animal model of post-traumatic stress disorder. Behavioural Brain Research, 268, 72–80. doi:10.1016/j.bbr.2014.03.029
- Wilson, C.B., McLaughlin, L.D., Nair, A., Ebenezer, P.J., Dange, R., & Francis, J. (2013). Inflammation and oxidative stress are elevated in the brain, blood, and adrenal glands during the progression of post-traumatic stress disorder in a predator exposure animal model. PLoS One, 8, e76146. doi:10.1371/journal.pone.0076146
- Wilson, C.B., McLaughlin, L.D., Ebenezer, P.J., Nair, A.R., Dange, R., & Harre, J.G. (2014b). Differential effects of sertraline in a predator exposure animal model of post-traumatic stress disorder. Frontiers in Behavioral Neuroscience, 8, 256. doi:10.3389/fnbeh.2014.00256
- Yu, G., Wang, L.G., Han, Y., & He, Q.Y. (2012). Clusterprofiler: An R package for comparing biological themes among gene clusters. Omics, 16, 284–287. doi:10.1089/omi.2011.0118
- Zeisberg, E.M., Tarnavski, O., Zeisberg, M., Dorfman, A.L., McMullen, J.R., Gustafsson, E., … Kalluri, R. (2007). Endothelial-to-mesenchymal transition contributes to cardiac fibrosis. Nature Medicine, 13, 952–961. doi:10.1038/nm1613
- Zen, A.L., Whooley, M.A., Zhao, S., & Cohen, B.E. (2012). Post-traumatic stress disorder is associated with poor health behaviors: Findings from the heart and soul study. Health Psychology: Official Journal of the Division of Health Psychology, American Psychological Association, 31, 194–201. doi:10.1037/a0025989
- Zoladz, P.R., Conrad, C.D., Fleshner, M., & Diamond, D.M. (2008). Acute episodes of predator exposure in conjunction with chronic social instability as an animal model of post-traumatic stress disorder. Stress (Amsterdam, Netherlands), 11, 259–281. doi:10.1080/10253890701768613
- Zoladz, P.R., & Diamond, D.M. (2013). Current status on behavioral and biological markers of PTSD: A search for clarity in a conflicting literature. Neurosci Biobehav Rev, 37, 860–895. doi:10.1016/j.neubiorev.2013.03.024
- Zoladz, P.R., Fleshner, M., & Diamond, D.M. (2012). Psychosocial animal model of PTSD produces a long-lasting traumatic memory, an increase in general anxiety and PTSD-like glucocorticoid abnormalities. Psychoneuroendocrinology, 37, 1531–1545. doi:10.1016/j.psyneuen.2012.02.007
- Zoladz, P.R., Fleshner, M., & Diamond, D.M. (2013). Differential effectiveness of tianeptine, clonidine and amitriptyline in blocking traumatic memory expression, anxiety and hypertension in an animal model of PTSD. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 44, 1–16. doi:10.1016/j.pnpbp.2013.01.001
- Zoladz, P.R., Park, C.R., Fleshner, M., & Diamond, D.M. (2015). Psychosocial predator-based animal model of PTSD produces physiological and behavioral sequelae and a traumatic memory four months following stress onset. Physiology and Behavior, 147, 183–192. doi:10.1016/j.physbeh.2015.04.032