
Abstract
Early life adversity and chronic inflammation have both been associated with cognitive impairment and neural compromise. In this study, we investigated the interactions between a history of chronic adolescent stress (CAS) and repeated endotoxin exposure on behavior, synaptic mitochondria, and microglia in adult male and female Wistar rats. Adult rats from chronic stress and control conditions were exposed to either repeated endotoxin (lipopolysaccharide; LPS) or saline injections every 3 days for 9 weeks. In both sexes, repeated LPS, regardless of stress history, impaired working memory in the Y maze. Regarding spatial memory, LPS impaired function for females; whereas, CAS altered function in males. Although males had an increase in anxiety-like behavior shortly after CAS, there were no long-term effects on anxiety-like behavior or social interaction observed in males or females. Stress did not alter synaptic mitochondrial function in either sex. Repeated LPS altered synaptic mitochondrial function such that ATP production was increased in females only. There were no observed increases in IBA-1 positive cells within the hippocampus for either sex. However, LPS and CAS altered microglia morphology in females. Impact of repeated LPS was evident at the terminal endpoint with increased spleen weight in both sexes and decreased adrenal weight in males only. Circulating cytokines were not impacted by repeated LPS at the terminal endpoint, but evidence of CAS effects on cytokines in females were evident. These data suggest a long-term impact of chronic stress and an impact of repeated endotoxin challenge in adulthood; however, not all physiological and behavioral metrics examined were impacted by the paradigm employed in this study and the two environmental challenges rarely interacted.
1. Introduction
Chronic stress during adolescence can serve as a risk factor for the development of neuropsychiatric disorders (Heim et al., Citation2008; Zisook et al., Citation2022) and women are twice as likely to suffer from these disorders compared to men (Kessler et al., Citation1994; Piccinelli & Wilkinson, Citation2000). Chronic stress during adolescence can shift the function of the hypothalamic pituitary adrenal (HPA) axis (Chaloner & Greenwood-Van Meerveld, Citation2013), leading to alterations in stress responsivity, neurocognitive performance, and plasticity (Bourke & Neigh, Citation2011; Chaby et al., Citation2015; Hyer et al., Citation2021) – outcomes that are also heavily influenced by sex. HPA axis function is tightly correlated with the immune response (Horowitz & Zunszain, Citation2015; Rohleder et al., Citation2001). Alterations of the balance between HPA axis function and immune responsivity following chronic stress increases the risk for dysregulation of immune function that can manifest in chronic inflammatory diseases (Cohen et al., Citation2012; Miller et al., Citation2002), exacerbating risk for psychiatric disease. The mechanisms that drive the relationship between stress and inflammation, their combined influence on physiology and behavior, and how sex influences these interactions remain unclear.
Chronic inflammation can originate from multiple sources and has been proposed as a primary mediator of the long-term effects of stress exposure (Liu et al., Citation2017). One source of chronic inflammation within the body is chronic exposure to endotoxin due to increases in gut permeability (Rudzki & Szulc, Citation2018). For instance, a history of three or more adverse childhood experiences has been associated with increased circulating lipopolysaccharide (LPS) binding protein in adult women (Keirns et al., Citation2023). Furthermore, repeated exposure to LPS has been linked to neurodegenerative disorders (Brown, Citation2019) and shown to accelerate pathology in rodent models of neurodegeneration (Frank-Cannon et al., Citation2008; Kitazawa et al., Citation2005; Noailles et al., Citation2018). We have previously demonstrated that a history of chronic adolescent stress augments the physiological and behavioral response to an acute exposure to LPS (Bekhbat et al., Citation2019, Citation2020; Hyer et al., Citation2023; Pyter et al., Citation2013). If chronic adolescent stress leads to an augmented response to LPS challenge, and individuals with a history of early life stress are potentially exposed to higher circulating LPS, this could create a feed-forward cycle to promote inflammation and adverse outcomes. Therefore, the current study leveraged an established mixed modality stress paradigm of psychosocial stress, restraint, and isolation during adolescence within rats (Bourke & Neigh, Citation2011; Hyer et al., Citation2021; Rowson et al., Citation2019) to test the hypothesis that a history of chronic stress would excarebate the behavioral and physiological response to repeated LPS exposure in male and female rats.
Synaptic performance following chronic stress or inflammation can be affected by altered metabolic capacity, which can contribute to synaptic decline and subsequent failures in cognitive performance (Guo et al., Citation2017). This is due to mitochondrial energy production contributing to the impact of stress (Eagleson et al., Citation2020; Picard et al., Citation2015; Shaw et al., Citation2020) and inflammation (Kim et al., Citation2019; Liu et al., Citation2017; Shaw et al., Citation2020) on cognition. Therapeutically targeting mitochondria alleviates inflammatory-induced memory deficits (Zhao et al., Citation2019), suggesting mitochondrial function may be a key responder to inflammatory mediators. In addition, sex hormones influence mitochondrial function (Gaignard et al., Citation2017; Shaw et al., Citation2021), positioning mitochondria as a pivotal player in elucidating sex differences in the response to stress and inflammation. Finally, we’ve previously reported that in mice the combination of a history of stress and repeated LPS exposure precipitates sex-specific changes in synaptic mitochondria (Shaw et al., Citation2020), suggesting that a similar phenomenon could be present in rats.
In addition, microglia are of particular relvance because activated microglia can release neurotoxins that disrupt synaptic function through disruption of neuronal mitochondrial at the level of the synapse (Harland et al., Citation2020) highlighting the value of examining both synaptic mitochondrial function and microglial endpoints. Given our previous report of chronic adolescent stress effects on microglia complexity (Bekhbat et al., Citation2020), we examined the impact of stress and repeated LPS exposure on microglia. Following exposure to an inflammatory challenge or stressor, microglia, the immune cells of the brain, become “activated” and present an overall change in morphology (Frank et al., Citation2007; Hinwood et al., Citation2012), synaptic pruning (Xin et al., Citation2019), and release of inflammatory mediators and neurotoxic factors to nearby neurons (Liddelow & Barres, Citation2017). Rats with a history of chronic adolescent stress display an exaggerated activation of the NF-κB pathway following an acute inflammatory insult as adults, but only females exhibit increased hippocampal microglial complexity (Bekhbat et al., Citation2020). Microglia work in tandem with the neuronal synapse and contribute to synaptic plasticity—a necessary function for learning and memory processes to occur (Basilico et al., Citation2019; Sanguino-Gómez et al., Citation2021). Inflammation-induced microglia activation contributes to deficits in spatial memory tasks (Hauss-Wegrzyniak et al., Citation2000). Similarly, chronic stress increases glial activation and decreases neural plasticity (Bourgognon & Cavanagh, Citation2020). As microglia express both glucocorticoid and sex steroid receptors (Sierra et al., Citation2008), they are positioned to be key regulators of sex differences in stress-induced synaptic plasticity along with their role in neuroinflammation.
We examined the combined impact of a history of chronic adolescent stress and repeated exposure to LPS on behavioral outcomes, synaptic mitochondrial respiration, and microglia morphology to understand potential consequences near the synapse that may co-occur or contribute to deficits in behavior. Collectively, we assessed the hypothesis that a history of chronic adolescent stress interacts with repeated endotoxin challenge to disrupt cognitive behavior and cause concomitant alterations in synaptic mitochondrial function and microglial activation.
2. Methods
2.1. Animals
Male and female Wistar rats were born to timed pregnant females (Charles River, NC) and culled to four males and four females per litter. Although shipping stress during puberty has been shown to impact later outcomes (Laroche et al., Citation2009), shipping while in utero has only been demonstrated to exert longterm effects if combined with a pharmacological challenge (Ogawa et al., Citation2007). Furthermore, we have previously shown effects of the chronic adolescent stress paradigm used here in both rats born from an in-house breeding colony and those born to timed pregnant rats shipped during gestation (Bekhbat et al., Citation2020; Rowson et al., Citation2019). Pups were weaned on postnatal day (PND) 21 and housed in same-sex pairs. Long-Evans retired breeder males (n = 12) and ovariectomized females (n = 12; Charles River, NC) were purchased to use as defeaters during the stress paradigm. These rats were housed in opposite sex pairs and allowed to acclimate to the colony for 1 week prior to use in the stress paradigm. Colony rooms were humidity controlled and maintained at 20–23 °C. Food and water were available ad libitum. All experiments were approved and performed in accordance with the Institutional Animal Care and Use Committee (IACUC) of Virginia Commonwealth University, as well as the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
2.2. Chronic adolescent stress (CAS)
CAS was performed as previously reported (Bourke & Neigh, Citation2011; Hyer et al., Citation2021; Rowson et al., Citation2019). In short, male (n = 18) and female (n = 16) rats assigned to the CAS groups were individually housed beginning on PND 35 for the duration of the experiment. On PND 38, all animals in the CAS group began 12 randomized, alternating days of physical restraint (6 total days) and social defeat (6 total days). For the physical restraint, each rat was placed in a Plexiglas restraint tube (BrainTree Scientific, Braintree, MA) such that it wasn’t compressed but could not turn 180° for 60 min. For the social defeat paradigm, experimental Wistar rats were placed into the home cage of a same-sex Long-Evans adult rat with a perforated plastic barrier dividing the cage but allowing for sensory detection between the two rats. After 2 min the barrier was lifted, and the animals interacted for five minutes or three pins, whichever came first. A pin was defined as the Long-Evans rat using both forepaws to restrain the experimental Wistar in a supine position. Following this period, the barrier was replaced for 25 min to allow for full sensory interaction but no physical contact. All CAS procedures occurred during the light cycle and all animals were returned to their home cages immediately upon completion of the stress paradigm each day. On PND 60, non-stress animals were individually housed, and all animals were given enrichment. Non-stressed groups were individually housed in adulthood to isolate the effect of the adolescent paradigm from individual housing in adulthood on the behavioral and physiological metrics within this study. Previous work has demonstrated that the CAS paradigm elicits effects beyond those of isolation housing (Bourke & Neigh, Citation2011).
2.3. Chronic LPS administration
Injections of either saline or LPS (7.5 × 104 Endotoxin units from Escherichia coli O111:B4; intraperitoneal; Sigma Aldrich cat. no. L4391) began on PND 62 and were administered every third day for 9 weeks. The regimen for repeated LPS treatment was adapted from previous work showing that chronic low-dose administration of LPS exacerbates Alzheimer’s Disease-related pathology (Kitazawa et al., Citation2005), Parkinson’s-related symptoms (Frank-Cannon et al., Citation2008), and degeneration associated with retinal dystrophy (Noailles et al., Citation2018) in rodent models. Behavioral assessments were collected beginning on PND 75 through PND 118 (see section 2.4 below). All tissues were collected on PND 122/125 (see section 2.6 below). Litters were counterbalanced across all groups and final groups sizes were as follows: non-stress, saline-treated males n = 10; non-stress, LPS-treated males n = 10; CAS, saline-treated males n = 9; CAS, LPS-treated males, n = 9; non-stress, saline-treated females n = 8; non-stress, LPS-treated females n = 8; CAS, saline-treated females n = 8; CAS, LPS-treated females, n = 9. Injection volume was calculated based on weight taken prior to each injection. Weight measurements were used to track weight gain across the course of the study. Health assessments were conducted on the day after each injection. Metrics were conducted via home cage visual observation by an individual blind to treatment group and included mobility, breathing, diarrhea, and porphyrin. Mobility and breathing were given a score between 1 and 3 with 1 being normal and 3 being immobile or labored breathing. Diarrhea and porphyrin were scored with a 1 for yes or 0 for no. Weights and health metrics were averaged by week to present a final weekly value for each variable across the course of the study. Injections were paused for 2 weeks during training on the Barnes Maze as any acute sickness effects could have altered performance on individual days, thus skewing the data for the behavioral task. Injections were reinstated following the second probe trial on PND 107 to allow for assessment of repeated LPS exposure on mitochondrial function and microglial morphology at the terminal endpoint.
2.4. Behavioral assessments
To determine how stress history and chronic exposure to LPS might alter behavior and cognition, behavioral assessments began on PND 76 consisting of the Open Field Task (OFT), Y-Maze task, Social Interaction task, and Barnes Maze task. All behavioral assessments were recorded and analyzed using EthoVision XT 14.0 (Noldus, Leesburg, VA). Animals were habituated to transport between the colony room and the behavior room for 2 days prior to testing. A timeline of the experimental design including timing of behavioral tests is provided in .
Figure 1. (A) Schematic of the experiment timeline. Male and female Wistar rats underwent a mixed modality chronic adolescent stress paradigm consisting of social defeat and restraint or remained non-stressed. Rats received an intraperitoneal injection of saline or LPS every 3 days for 9 weeks. On PND 125, central and peripheral tissues were collected. Weight was monitored throughout the experiment. A history of chronic stress slowed weight gain in males regardless of LPS challenge (B) but did not alter weight in females (C). *p < 0.05. Symbols represent mean ± SEM. Panel A was created using Biorender.com.
2.4.1. Open field
Anxiety-like behavior was assessed in the Open Field task and conducted as reported in Kloster et al. (Citation2020). Animals were placed into the center of a square open field (90 cm × 90 cm) and allowed to freely explore the arena for 10 minutes before being returned to their home cage. Percent time spent in the center and corners of the maze as well as distance traveled were recorded.
2.4.2. Social Interaction
The Social Interaction task was used to assess anxiety-like and anhedonia-like behavior (File & Hyde, Citation1978). The task was conducted by placing an experimental animal into the open field arena with a same-sex, novel conspecific (Kloster et al., Citation2020). The animals were allowed to freely interact for 10 minutes. Latency to initial approach and duration of contact were measured. Additionally, velocity was recorded to verify no deficits in motor performance at time of assessment (La-Vu et al., Citation2020).
2.4.3. Barnes Maze
The Barnes Maze task was conducted as described in Hyer et al. (Citation2021) to assess spatial learning and memory. Briefly, all testing was done in a room lit to 2500 lux on a black, circular table (121 cm in diameter) elevated 91.5 cm from the floor with 20 equidistant holes (10 cm in diameter) around the perimeter fitted with 19 false boxes and one goal box. Acquisition training was conducted over 5 days with two trials per day and a three-minute inter-trial interval. Rats had up to 3 min per trial to locate the goal box location. The first probe was conducted 48 h after the final acquisition trial. The probe task consisted of a single five-minute trial without an open goal box. Reversal learning was conducted over three days with two trials per day and a 3-min inter-trial interval. The goal box was moved to the opposite quadrant and rats had up to 3 min to locate the new location. The second probe was conducted 48 h after the final reversal trial. Data recorded included summed latency to locate the goal box (seconds), number of incorrect holes visited prior to locating the goal box (errors), and distance traveled (cm).
2.4.4. Y-Maze
The Y-Maze task was used to assess spatial working memory (Kraeuter et al., Citation2019). Performance on the Y-Maze task was assessed three days after an LPS injection by placing the animal into one arm of a three-arm elevated maze with enclosed sides shaped like a Y. Animals had eight minutes to freely explore all three arms of the maze. The Y-Maze task assesses working memory performance by determining how often an animal visits all three arms sequentially—called spontaneous alternation (Kraeuter et al., Citation2019). The alternation index is calculated by taking the number of sequential arm visits divided by the total possible sequences given the total number of arms visited. This accounts for any differences in mobility between animals. We recorded the alternation index along with velocity for each animal.
2.5. Vaginal lavage
Estrus cycle stage was tracked using vaginal lavage (Cora et al., Citation2015). Female rats were gently restrained, and the vaginal canal was flushed with 200 μL of phosphate buffered saline (PBS) and the wet sample was imaged under a light microscope at 10×. The quantity of round cells, needle cells, and/or neutrophils was used to determine terminal cycle stage.
2.6. Tissue collection
Three days following the final injection, tissues were collected. Two collection days were necessary, on PND 122 or 125, due to the limited number of mitochondrial assessments that can be completed in one day. Experimental groups were counterbalanced across both collection days. Rats were rapidly decapitated without the use of anesthesia as this has been shown to alter corticosterone levels (Bekhbat et al., Citation2016), brains were extracted and rinsed in isotonic sucrose solution (0.32 M sucrose, 1 mM EDTA, 5 mM Tris, pH 7.4). For the synaptosome preparation, one hemisphere was diced and homogenized in a 7 mL glass Dounce homogenizer in isotonic homogenizing buffer using five strokes of the loose plunger and five strokes of the tight plunger. For immunohistochemistry, the second hemisphere was post-fixed in 4% paraformaldehyde for 48 h. In addition to the brain, spleen, adrenal, and gonads were collected and weighed at the terminal endpoint.
2.7. Synaptosome isolation
Synaptosome isolation was adapted from (Dunkley et al., Citation2008) and performed, in part, as described in Kloster et al. (Citation2020). Briefly, homogenized samples were spun at 3600 RPM for 10 minutes at 4 °C then the supernatant transferred to a clean tube and diluted to 7 mL with homogenizing buffer. The diluted supernatant was laid across a five-layer discontinuous Percoll gradient in a 26 mL polycarbonate ultracentrifuge tube then spun at 32,500 × g for 15 min to separate synaptosomes. The synaptosome layer was extracted from the interface between the 15% and 23% Percoll layers and placed in a clean polycarbonate tube with 20 mL of ionic media (20 mM HEPES, 10 mM D-Glucose, 1.3 mM Na2HPo4, 1 mM MgCl2, 5 mM NaHCO3, 5 mM KCl, 140 mM NaCl, pH 7.4) and spun for 35 min at 15,000 × g. The resulting synaptosome pellet was measured via Nanodrop for protein content then diluted to 40 µg/100 µL in Ionic media in a clean tube. 300 µL of the synaptosome sample was used to measure mitochondrial respiration and the remaining sample was stored at −20 °C for Western Blots.
2.8. Mitochondrial respiration
Respiration was assessed as reported in Shaw et al., Citation2020. Briefly, synaptosomes were plated in triplicate on a Seahorse XFe24 cell plate coated with poly-D-lysine (Agilent Technologies, Santa Clara, CA), the plate was spun to adhere the synaptosomes, ionic media was replaced with assay media, and plates were incubated at 37 °C in a non-CO2 incubator for 30 minutes. For the Cell Mitochondrial Stress Test assay (Agilent Technologies, PN: 103015-100), injected drugs were prepared and diluted per manufacturer’s instructions and added to the Seahorse XFe24 FluxPak (2.0 µM Oligomycin, 1.0 µM carbonyl cyanide-p-trifluoromethoxy-phenol-hydrazone (FCCP), 0.5 µM Rotenone/antimycin A per well). The plate and FluxPak were inserted into the Agilent Seahorse and oxygen consumption rate (OCR) was determined by sequential measurement cycles with three measurements following administration of each drug. As a complex V inhibitor, OCR following Oligomycin allowed for assessment of ATP-linked respiration (subtraction of baseline OCR) and proton leak (subtraction of non-mitochondrial respiration). FCCP was introduced as a protonophore that collapses the inner membrane gradient pushing the electron transport chain to the maximal rate where OCR displayed maximal respiration. Antimycin A and Rotenone were used to terminate the electron transport chain function by inhibition of complex III and I and demonstrated non-mitochondrial respiration. Spare capacity was calculated by subtraction of basal respiration from maximal respiration.
2.9. Western Blots
Western Blot analysis was conducted as reported in Shaw et al. (Citation2020). To determine the relative density of mitochondria within the synaptosomes, expression of Hexokinase I and SNAP-25 via western blot were assessed. Hexokinase I was chosen to label mitochondria due to its primary role in synaptic mitochondria as opposed the periphery (Robey & Hay, Citation2006) and is resistant to mitochondrial dissociation following inflammation (Han et al., Citation2021; Schindler & Foley, Citation2013). SNAP-25 is a pre-synaptic vesicle tethering protein that accurately reflects our pool of synaptosomes that consist of, at the minimum, the presynaptic terminal and post-synaptic density (Shaw et al., Citation2020). To assure adequate protein amounts for loading, samples were pooled according to group.
2.10. IBA-1 immunohistochemistry
Following post-fixation, tissue was moved to 30% sucrose in PBS for ∼7 days for cryoprotection, sectioned on a cryostat (Leica; CM 1950) at 40 µm, and sections were stored in PBS with 0.1% sodium azide at 4 °C until staining. Sections throughout the rostral-caudal extent of the hippocampus were mounted onto slides, allowed to dry, rinsed in PBS, and then incubated in 0.6% H2O2 in PBST for 30 min. Slides were rinsed with PBS and placed in a heated citrate buffer (10 mM Citric Acid, pH 6.0) for 20 minutes. After rinsing in PBS, tissue was blocked in 10% Normal Horse Serum (Vector, Cat # S-2012) and 0.4% Triton x-100 (Sigma, Cat # X100-100ML) in PBS used for the blocking buffer for 90 minutes. Following the blocking buffer, tissue was treated overnight with rabbit anti-IBA-1 antibody (1:2000, Wako cat. # 019-19741). Sections were rinsed with PBS and treated with secondary antibody goat anti-rabbit IgG (H + L) (1:250, Vector Laboratories, cat# BA-1000) in 10% Normal Goat Serum, 0.4% Triton x-100 in PBS for 1 h at room temperature. Sections were rinsed in PBS and incubated in an ABC solution (ThermoScientific, Cat # 32020) for 30 min, rinsed, and then treated with DAB (Sigma, D4293-50SET) for 3 min. After a final rinse, sections were dried overnight, rehydrated with dH2O, and immersed in Cresyl Violet. Sections were then cleared in a 70% ethanol (EtOH) solution with 6 drops of glacial acetic acid for 10 minutes followed by serial dehydration in 95% and 100% EtOH solution for 5 min each before final immersion in Xylene. Slides were cover-slipped with Permount.
2.11. Microglial counting and reconstruction
Tissue sections sampled throughout the rostral-caudal axis of the hippocampus were used to assess microglia. Using the Paxinos and Watson brain atlas (Paxinos & Watson, Citation2007), hippocampal images anterior to Figure 70 were considered dorsal and images posterior considered ventral. For microglia morphology, cells were visualized on a widefield microscope (Zeiss Axio Imager, Carl Zeiss, Thornwood, NY) at 100x oil immersion, and microglia were reconstructed using Neurolucida software (MBF Biosciences, Williston, VT) by an individual blind to treatment groups. As the dorsal and ventral regions of the hippocampus mediate different neural functions (Fanselow & Dong, Citation2010), microglia were imaged and analyzed separately for each. Four randomly selected microglia were imaged for each: the apical and basal regions of the CA1 and CA3 regions of the hippocampus, two per dorsal and ventral sections. Six total microglia were imaged in the DG region, three per dorsal and ventral sections. An example image and reconstruction is provided in Supplemental Figure 8. Reconstructed cells were uploaded to Neurolucida Explorer and quantitative data was acquired using three dimensional Sholl analysis spheres with an initial diameter of 10 µm and additional concentric spheres every 5 µm. Cells were reconstructed to assess changes in morphology which has been shown to change following exposure to an environmental stressor (Hellwig et al., Citation2016; Tynan et al., Citation2010).
Images were taken of the hippocampus using a Keyence Microscope (BZ-X810) at 10× magnification. Using the Keyence software measurement tool, the DG, CA1, and CA3 were outlined, and the area (µm3) recorded. The outlined images were uploaded to Image J (Version 1.53o) (Schneider et al., Citation2012) and the counting tool was used to label all microglia within the outlined area. The density was calculated by dividing the number of microglia by the area of the region. Density was calculated for the dorsal and ventral DG, CA1, and CA3 by averaging all individual images for each region of interest (ROI) throughout the extent of the hippocampus.
2.12. Peripheral inflammation
Circulating cytokines were assessed from terminal trunk blood. Blood was collected, allowed to clot, the clot was removed and the sample was spun for 20 min at 1800 rcf. Serum was collected and analyzed using the V-Plex Pro-Inflammatory Cytokine Panel for rat on the Meso Scale Discovery system (Meso Scale Diagnostics, Rockville, MD, USA; cat. no. K15059D). Titration analysis against a standard curve indicated dilution factors of 1:3. MSD protocol was conducted per manufacturer’s instructions.
2.13. Statistical analysis
Direct statistical comparisons between males and females were not completed. We have previously observed sex differences in the physiological response to stress and acute LPS (Bekhbat et al., Citation2019), but the data analysis presented here is designed to assess the outcomes of repeated LPS and chronic stress through a multifaceted characterization of outcomes for each sex. As sex differences may fluctuate throughtout the time course of the study and we focused on a single endpoint for each metric assessed, the outcomes are analyzed separately for males and females. Data were analyzed using GraphPad Prism version 9. Non-parametric one-way repeated measures ANOVA (Friedman Test) was used to individually analyze health metrics across the study for each group. Three-way ANOVA was used to analyze the effects of stress history, treatment, and timepoint/week where appropriate. Where three-way ANOVA revealed significant interactions, data were collapsed across the non-significant variable and analyzed by two-way ANOVA. For analyses containing only two variables, two-way ANOVA was used. Post-hoc analyses were used to determine the directionality of effects where significant interactions were observed.
3. Results
3.1. Stress history and repeated LPS exposure altered physical health metrics
All males gained weight each week (F(9,333)=74.69; p < 0.0001; ) but males with a history of stress were slower to gain weight than controls (F(1,333)=63.83; p < 0.0001). LPS did not alter weight gain (p > 0.05). In females, there was a main effect of week (F(9,285)=35.74; p < 0.0001) on weight gain such that all females gained weight over the course of the study. Stress history (p > 0.05) and LPS exposure (p > 0.05) did not alter weight gain in females ().
Health metrics were tracked for 24 h following each injection across the course of the study resulting in weekly health metric scores (Supplemental Figure 1). Each group was individually analyzed along the 9 weeks to assess changes along the timecourse of the study. There were no effects of LPS (p > 0.05) or stress history (p > 0.05) on diarrhea, porphyrin, mobility or breathing in males or females.
LPS exposure increased terminal spleen weight (F(1,34)=17.15; p = 0.0002) and decreased adrenal weight (F(1,34)=4.56; p = 0.04) in males (Supplemental Figure 2(A,B)). Regardless of stress history (p > 0.05), males exposed to repeated LPS had a higher terminal spleen weight and a lower adrenal weight compared to saline controls. Terminal testicle weight was not altered by LPS (p > 0.05) or stress history (Supplemental Figure 2(C); p > 0.05). In females, LPS exposure increased terminal spleen weight (F(1,29)=12.48; p = 0.001). LPS treated females had heavier spleen weights compared to saline-injected controls regardless of stress history (Supplemental Figure 2(D); p < 0.05). There was no significant effect of LPS (p > 0.05) or stress history (p > 0.05) on terminal adrenal weight (Supplemental Figure 2(E)) or uterine weight of females (Supplemental Figure 2(F)). Similar uterine weight among groups suggests that there were no systematic group differences in estrous cycle stage among the groups at the terminal endpoint.
Circulating cytokines were measured in terminal trunk blood (). In males, levels of the pro-inflammatory cytokines IL-13, IL-1β, IFNγ, TNFα, or KC/GRO were not altered by stress history (p > 0.05) or repeated LPS exposure (p > 0.05). Similarly, there were no effects of stress history (p > 0.05) or repeated LPS exposure (p > 0.05) on the circulating anti-inflammatory cytokines IL-4, IL-5, and IL-10 in males. In females, stress history (F(1,25)=4.85; p = 0.04) lowered IL-13 levels with CAS females having less circulating IL-13 compared to non-stress controls regardless of LPS exposure. There were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on the remaining circulating pro-inflammatory cytokines IL-1β, IFNγ, TNFα, or KC/GRO in females. A history of stress (F(1,25)=7.51; p = 0.01) lowered the circulating anti-inflammatory cytokine IL-10 with CAS females having less IL-10 compared to non-stress control females. The remaining anti-inflammatory cytokines, IL-4 and IL-5, were not altered by stress history (p > 0.05) or repeated LPS (p > 0.05) in females.
Table 1. Circulating pro-inflammatory (IL-13, IL-1β, IFNγ, TNFα, KC/GRO) and anti-inflammatory (IL-4, IL-5, and IL-10) cytokines were assessed from terminal trunk blood.
3.2. Behavioral metrics were altered by stress history and repeated LPS exposure
3.2.1. Open field task
Males with a history of stress spent less time in the center of the open field compared to non-stress males (F(1,33)=5.59; p = 0.02; ) and increased time in the corners of the periphery (F(1,33)=4.83; p = 0.04; ) on the first open field exposure which was conducted 14 days after the initiation of repeated LPS exposure. There were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on distance traveled (). In females, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on percent time spent in the center, time in corners of the maze, or distance traveled (). To determine if there were changes in open field performance over time, we assessed these same metrics 5 weeks later after prolonged repeated LPS exposure. In males, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on time spent in the center or corners of the maze on the second trial. Males with a history of stress traveled more than non-stress controls (F(1,33)=8.61; p = 0.006; ). There were no effects of stress history (p > 0.05) or LPS (p > 0.05) on time spent in the center or corners of the maze or distance traveled in females on the second trial ().
Figure 2. In the first open field trial males with a history of chronic adolescent stress (CAS) (A) spent less time in the center and (B) more time in the periphery of the open field while C) mobility was not altered. This assessment was conducted on PND 76 which was over 3 weeks after the last stressor and 2 weeks into the chronic LPS paradigm. Behavioral assessments were conducted 72 h after LPS injection. A second open field assessment was conducted approximately 5 weeks after the first assessment on PND 113. Effects observed in trial 1 were attenuated in trial 2, however, CAS males moved more during trial 2 compared to non-stress males. D-F) Females did not show any changes in open field performance with stress history or LPS challenge in either open field trial. *p < 0.05. Bars represent mean ± SEM.
3.2.2. Social interaction task
In both males and females, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on the latency to approach the conspecific, duration of contact, or velocity (Supplemental Figure 3).
3.2.3. Barnes Maze
Acquisition learning on the Barnes Maze task was measured by latency to locate the goal box, the number of errors, and distance traveled. In males, stress history (p > 0.05) and repeated LPS (p > 0.05) did not alter the latency to locate the goal box (), total errors (Supplemental Figure 4(A)), or distance traveled during acquisition learning (Supplemental Figure 4(B)). Similarly, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on latency to locate the goal box (), total errors (Supplemental Figure 5(A)), or distance traveled during reversal learning observed in males (Supplemental Figure 5(B). In female rats, there were no effects of stress history (p > 0.05) or repeated LPS exposure (p > 0.05) on latency to locate the goal box () or primary errors during acquisition (Supplemental Figure 4(C)). However, stress did lower total distance traveled in females during acquisition (F(1,145)=5.55; p = 0.02; Supplemental Figure 4(D)). There was an effect of repeated LPS on total errors during the reversal learning in females only, such that LPS exposed females demonstrated fewer errors compared to saline treated females (F(1,87)=5.67; p = 0.02; Supplemental Figure 5(C)).
Figure 3. Barnes Maze was conducted starting on PND 95 after 3 weeks of repeated LPS injections. No LPS injections were given during acquisition, probe, or reversal. During acquisition training for the Barnes Maze task there were no changes in summed latency to target (A, B). stress history and LPS exposure history interacted to alter latency to locate the target position during Probe 1 in males (C). Males with a history of stress without LPS exposure history, showed an increased latency to locate the goal box position in Probe 1. In females, previous exposure to repeated LPS increased the latency to locate the goal box position in Probe 1 regardless of stress history. *p < 0.05. Bars represent mean ± SEM.
Figure 4. During reversal training for the Barnes Maze task there was not an influence of stress or history of repeated LPS on males (A) or females (B) in the summed latency to target. There were no effects of stress history or LPS history during Probe 2 in either sex (C). Bars represent mean ± SEM.
In males, there was a significant interaction between stress history and repeated LPS (F(1,33)=5.25; p = 0.03) on latency to locate the goal box location during probe 1 (). Post-hoc analysis indicated that males with a history of stress, but not repeated LPS, were slower to locate the goal box compared to non-stress males (p = 0.01 (saline exposed); p = 0.03 (LPS exposed)). In addition, males with a history of CAS and repeated LPS were also faster than CAS only males (p = 0.007). In females, repeated LPS exposure (F(1,27)=4.21; p = 0.05) increased latency to locate the goal box position such that LPS exposed females, regardless of stress history, took longer to locate the goal box compared to saline controls (). There were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on primary errors or distance traveled in probe 1 for males or females (Supplemental Figure 6). There were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on latency to locate the goal box position, primary errors, or distance traveled in probe 2 for either males or females (Supplemental Figure 6).
3.2.4. Y-Maze
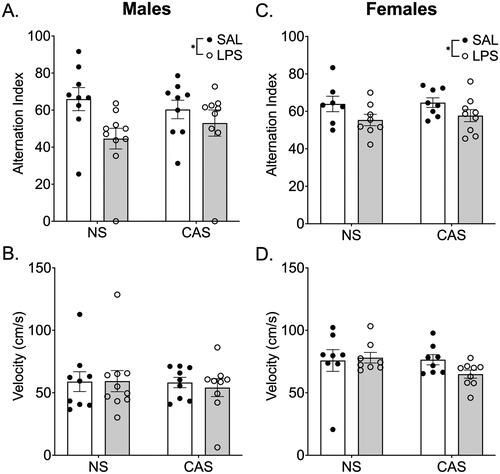
Repeated LPS (F(1,33)=5.69; p = 0.02), but not stress history (p > 0.05), impacted the alternation index during the Y-Maze task in males. Males repeatedly exposed to LPS had a lower alternation index score compared to saline-treated males suggesting impaired working memory performance (). Stress history (p > 0.05) and LPS (p > 0.05) did not alter velocity in males (). In females, there was an effect of LPS exposure (F(1,28)=5.67; p = 0.02) on alternation index such that LPS exposed females had a lower alternation index compared to saline treated females regardless of stress history (). There were no effects of repeated LPS (p > 0.05) or stress history (p > 0.05) on velocity in females ().
Figure 5. Y-Maze was conducted on PND 116, over 2 months after stressors concluded and 72 h after LPS injection during the 4th week of LPS injections every 3rd day. In both males (A) and females (C) repeated LPS decreased the alternation index in the Y-Maze task regardless of stress history suggesting an impairment in working memory performance. The alternation index is the number of actual alternations based on the number of possible alternations given the total number of arms visited. There were no changes in velocity (B, D). *p < 0.05. Bars represent mean ± SEM.
3.3. Stress history and repeated LPS altered mitochondrial respiration
3.3.1. OCR metrics
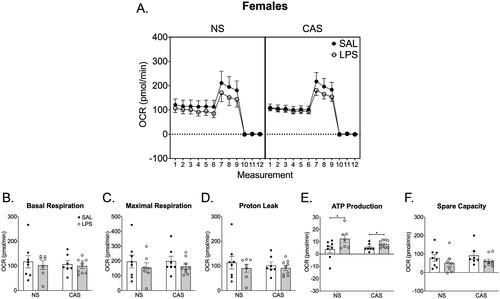
In males, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on mitochondrial dynamics including basal respiration, maximal respiration, proton leak, ATP-linked respiration, and spare capacity in males (). LPS exposed females, regardless of stress history, had increased ATP-linked respiration (F(1,27)=7.60; p = 0.01; ). There were no effects of stress history or repeated LPS exposure on basal respiration (p > 0.05), maximal respiration (p > 0.05), proton leak (p > 0.05), or spare capacity (p > 0.05) in females.
Figure 6. Synaptosomes were collected on PND 122 and 125. This was over 10 weeks after the final stressor and 72 h following the final repeated LPS injection. We collected tissue 72 h after LPS injection in order to reflect mitochondrial function at a timepoint relative to LPS that was similar to the final open field and Y maze assessments. Mitochondrial respiration was not altered by stress history or chronic LPS history in males. A) Overall oxygen consumption rate (OCR) across the measurements of the Cell Mito stress Test. Mitochondrial dynamics, including basal respiration (B), maximal respiration (C), proton leak (D), ATP production (E), and spare capacity (F), were not altered by stress history or chronic LPS exposure. *p < 0.05. Symbols or bars represent mean ± SEM.
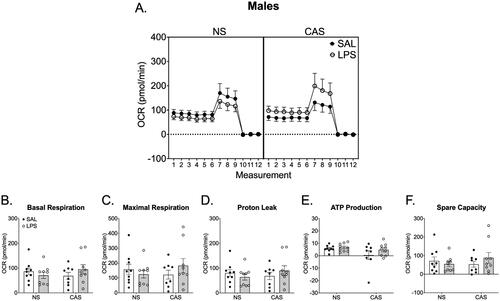
Figure 7. Synaptosomes were collected on PND 122 and 125. This was over 10 weeks after the final stressor and 72 h following the final repeated LPS injection. We collected tissue 72 h after LPS injection in order to reflect mitochondrial function at a timepoint relative to LPS that was similar to the final open field and Y maze assessments. (A) Overall oxygen consumption rate (OCR) across the measurements of the Cell Mito stress Test. (E) ATP production was increased in females following chronic LPS regardless of stress history. Basal respiration (B), maximal respiration (C), proton leak (D), and spare capacity (F) were not altered by repeated LPS exposure or stress history. *p < 0.05. Symbols or bars represent mean ± SEM.
3.3.2. Western Blots
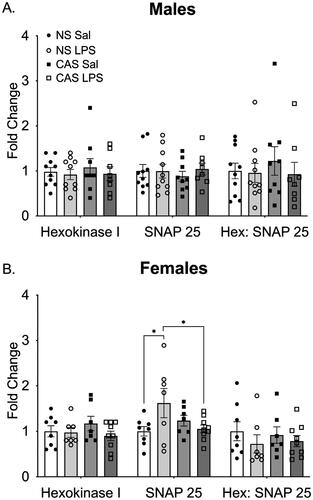
Synaptosome preparations were probed for mitochondria using Hexokinase I and presynaptic terminals using SNAP-25 (Supplemental Figure 7). In males, there were no effects of stress history (p > 0.05) or repeated LPS (p > 0.05) on fold change in Hexokinase I, SNAP-25, or the ratio between the two (). In females, there was no effect of stress history or repeated LPS on Hexokinase I fold change (). For SNAP-25, there was a significant interaction between stress history and repeated LPS (F(1,27)=5.49; p = 0.03). Post-hoc analysis indicated that non-stress females exposed to LPS had significantly higher SNAP-25 levels compared to saline controls (p = 0.02). Among the LPS exposed females, non-stress females had higher SNAP-25 compared to CAS females (p = 0.03). There were no effects of stress history (p > 0.05) or LPS (p > 0.05) on the ratio between Hexokinase I and SNAP-25.
Figure 8. (A) In males, neither stress history nor repeated LPS exposure altered density of the mitochondrial marker Hexokinase 1 or the synaptic marker SNAP25. The ratio of the two markers was similarly not altered. (B) In females, there was an interaction between stress history and repeated LPS exposure on SNAP25 such that non-stress females treated with LPS showed an increase in SNAP25. However, this was not reflected in the ratio of Hexokinase 1 to SNAP25. *p < 0.05. Bars represent mean ± SEM.
3.4. Microglial morphology was altered by stress history and repeated LPS
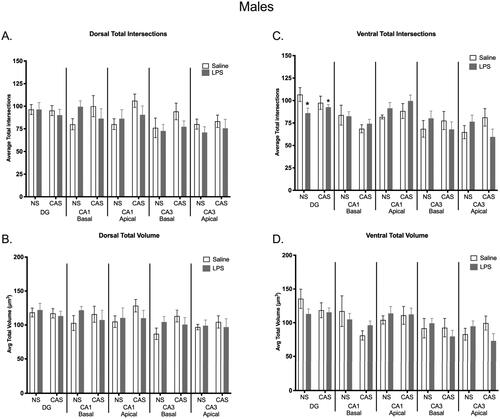
The total number of microglia counted, and overall density was not altered within any hippocampal subregion analyzed for either sex (Supplemental Figure 8). There were subregion specific effects of stress and LPS on microglia complexity and volume in the hippocampus. Within the ventral DG, LPS decreased total number of intersections in males (F(1,19)=4.450; p = 0.048; ). There were no other observed significant differences within hippocampal subregions in the males.
Figure 9. There was no effect of chronic adolescent stress or repeated LPS on microglia total process intersections within the dorsal hippocampus in males (a). There were no group differences observed in total microglia volume in the dorsal (B) or ventral (D) portions of the hippocampus within males. Within the ventral DG region, repeated LPS exposure lowered average total intersections regardless on stress history (C). *p < 0.05. Bars represent mean ± SEM.
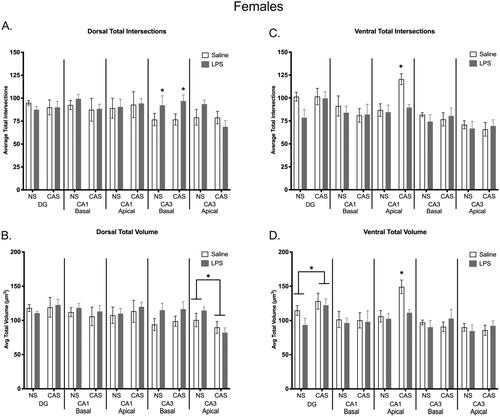
In the basal portion of the dorsal CA3, LPS exposure increased the total number of intersections in females (F(1,20)=5.5; p = 0.029; ). Within the apical portion of the dorsal CA3 region, stress decreased average total volume (F(1,20)=7.4; p = 0.013; ) in females. There were no significant differences in the ventral CA3 microglia within females. In the apical ventral CA1, intersections were altered by stress (F(1,20)=9.569; p = 0.006), LPS (F(1,20)=7.106; p = 0.015), and an interaction between stress and repeated LPS in females (F(1,20)=5.252; p = 0.033; ). Total volume in females was also altered by stress (F(1,20)=10.74; p = 0.004), LPS (F(1,20)=6.905; p = 0.016), and an interaction between the two variables (F(1,20)=4.741; p = 0.042; ). There were no significant differences in the dorsal CA1 microglia for females. A history of stress increased average total volume in the ventral DG of females only (F(1,20)=4.834; p = 0.040; ). No parameters were altered in the dorsal DG for either sex.
Figure 10. Microglia morphology was shifted in the dorsal and ventral regions of the hippocampus within females. Repeated exposure to LPS increased total number of intersections within the dorsal basal CA3 region (a). A history of chronic adolescent stress increased total intersections within the ventral apical CA1 region, but this effect of stress was not observed when females were also exposed to repeated LPS (C). A history of stress decreased average total volume in the dorsal apical CA3 region (B). Within the ventral hippocampus, stress history increased overall total volume in the DG and stress increased volume in the ventral apical CA1, which was again absent in stressed females with repeated LPS exposure (D). Bars represent mean ± SEM.
3.5. Estrus cycle stage was not altered by stress history or repeated LPS and did not correlate with any metrics
All females displayed a cyclic pattern of estrus stage across the experiment. Cycle stage did not correlate with terminal endpoints including basal respiration, ATP production, hexokinase, SNAP-25 or the HEX:SNAP ratio (p > 0.05 for all metrics tested).
4. Discussion
Here we demonstrate that while both a history of CAS and repeated exposure to the immune stimulus of LPS can alter physiology and behavior, the two environmental exposures generally do not interact to modify outcomes in the endpoints assessed, for either sex. Collectively, the outcomes of this study do not support our overarching hypothesis that a history of CAS interacts with repeated endotoxin challenge to disrupt cognitive behavior and cause concomitant alterations in synaptic mitochondrial function and microglial activation. However, the results of the study demonstrate nuanced influences of each environmental exposure, CAS and repeated LPS, for both males and females. Males with a history of CAS exhibited an increase in anxiety-like behavior in the open field, however this effect was transient and was not observed in the second exposure to the open field. Neither sex demonstrated an effect of CAS or LPS on social interaction. In both sexes, repeated LPS, regardless of stress history, impaired working memory. Spatial memory in males and females was differentially impacted by environmental exposure such that LPS impaired function for females; whereas, CAS altered function in males. Stress did not alter synaptic mitochondrial function in either sex, but repeated LPS altered synaptic mitochondrial function such that ATP production was increased in females only. There were no observed increases in IBA-1 positive cells within the hippocampus for either sex. However, LPS and CAS altered microglia morphology in females. Impact of repeated LPS was evident at the terminal endpoint with increased spleen weight in both sexes suggesting long-term effects of repeated LPS exposure. Circulating cytokines were not impacted by repeated LPS at the terminal endpoint, but evidence of CAS effects on cytokines in females were evident.
Both neuroinflammation and stress can impair memory performance. We tested the combined impact of CAS and repeated LPS on two memory behavioral tasks that rely on intact hippocampal connectivity (Gawel et al., Citation2019; Kraeuter et al., Citation2019), a region sensitive to stress and involved in cognitive decline (Sampath et al., Citation2017). Consistent with previous reports in male rats (Mamunur et al., Citation2023) and mice (Fronza et al., Citation2022), both male and female rats in this study demonstrated a LPS-induced impairment in working memory as assessed in the Y-maze. This effect was not modified by stress history in either sex. While neither sex exhibited a deficit in learning, nor cognitive rigidity, there were sex differences in spatial memory measured by the Barnes Maze. Males with a history of stress took longer to locate the goal box during the probe trial suggesting a spatial memory deficit – an effect not observed when stressed males were exposed to repeated LPS. This result was unexpected as we expected LPS to have a compounding effect on stress-induced memory deficits. It is possible for LPS administered prior to a stressor to result in anti-inflammatory phenotype in the hippocampus (Gu et al., Citation2021). However, in the current study, LPS exposure was initiated after completion of the adolescent stress paradigm. Interestingly, females that received repeated LPS, regardless of stress history, exhibited deficits in spatial memory during the probe trial. The results of the probe trial suggest spatial memory is differentially affected by stress and repeated LPS exposure in males and females, respectively. The lack of a compounding effect of stress and repeated LPS exposure in either sex was contrary to our hypothesis, however, the stressor was experienced during adolescence and the inflammatory stimulus was designed to model a chronic low-grade immune challenge, thus it is possible that presentation of an acute stressor just before testing or a more robust immune challenge could compound the deficits observed on memory performance in either males or females.
Mitochondrial function is sensitive to inflammatory insults (Picard et al., Citation2015; Zhao et al., Citation2019) and stress (Shaw et al., Citation2020; Tagliari et al., Citation2010). Considering that synaptic mitochondria deficits contribute to cognitive decline (Olesen et al., Citation2020), in this study, we measured mitochondrial respiration in isolated synaptosomes. We did not observe differences in mitochondrial respiration within the males, suggesting that the function of the electron transport chain itself remained intact across all groups. Despite observing deficits in memory performance, we were unable to detect alterations in mitochondrial respiration suggesting that this behavioral outcome may be influenced by another unknown neural mechanism or we were unable to detect a mitochondrial change due to lack of regional mitochondrial assessement. Synaptic respiration in females was vulnerable to the effects of LPS but not CAS. The proportion of mitochondrial respiration contributing to ATP production was increased in LPS treated females, suggesting that mitochondria in these groups increased efforts to produce ATP, perhaps in response to the negative effects of LPS. This aligns with assessment of memory in the Barnes Maze in which females that received repeated LPS exhibited longer latency to the goal box during the probe trial, with no effects of stress on ability to find the goal box. The presynaptic protein SNAP-25 is required for neurotransmitter release and neuronal plasticity (Antonucci et al., Citation2016), and we observed an increase in SNAP-25 in non-stressed females following repeated LPS. As both inflammation and stress can lead to neuronal atrophy and a decrease in dendritic spines (Haroon et al., Citation2017; Woolley et al., Citation1990), the increased SNAP-25 expression may be reflective of a neuroprotective mechanism to maintain plasticity within the non-stressed females, but this is not reflected in any behavioral outcomes for this group. Thus, increased SNAP-25 expression seems to have subtle influence on physiological outcomes within the non-stressed LPS exposed females. Neither stress or LPS altered the proportion of mitochondria at the synapse in males or females suggesting there were no deficits in mitochondria trafficking and only shifts in overall respiration. These data are reflective of a global response to stress and inflammation. Glucocorticoids have regionally specific influences on neuronal morphology (Shors et al., Citation2001) and can influence synaptic loss (Tata et al., Citation2006). As such, regional sensitivity that may have occurred throughout the hippocampus was not detectable here due to the volume of tissue necessary to complete the OCR assays.
Activated microglia can release cytokines to nearby neurons which can induce mitochondrial dysfunction. We assessed terminal neuroinflammatory tone by measuring microglia morphology throughout the hippocampus. Total volume and total intersections were quantified, as stress induces dynamic motility (Liu et al., Citation2021) and hyper-ramification of microglial processes (Bekhbat et al., Citation2020; Hinwood et al., Citation2012; Walker et al., Citation2013). Subregion-specific changes in morphology were exhibited along the dorsal-ventral axis in both sexes, which is consistent with the dorsal and ventral hippocampus exhibiting different stress responses (Ivens et al., Citation2019; Nasca et al., Citation2017). Within the dorsal hippocampus of females, repeated LPS increased basal CA3 intersections, while CA3 apical volume was decreased in stressed females. Sustained effects of CAS on the ventral apical CA1 region of stressed females were observed with increases in total microglia intersections and volume. Interestingly, this shift in morphology was lost in stressed females that received LPS, suggesting the long-term effects of CAS influenced the microglial response to LPS within the CA1 subregion for this endpoint. Considering that apical and basal dendrites can exhibit different mechanisms of synaptic plasticity (Brzdak et al., Citation2019) and exhibit a sex difference following stress (Galea et al., Citation1997; Shors et al., Citation2001), it may be expected that microglia exhibit dissimilar morphological responses near apical and basal dendrites. Further, changes in microglia morphology may be reflective of hippocampal neuronal remodeling following stress (Bollinger et al., Citation2017; Wohleb et al., Citation2018).
The complex region-specific responses to stress and LPS described here for females were not observed in the males where LPS resulted in a decrease in total ventral DG intersections only. This is consistent with other research that demonstrates a sex- and region-specific effect of stress on microglia state (Bollinger et al., Citation2017). Microglia morphology alone does not indicate changes in functional state (Woodburn et al., Citation2021), but the work presented herein reveals morphological changes along the trisynaptic circuit of the hippocampus. As we observed more changes in females, we would expect to observe a stronger influence of adolescent stress and LPS on memory performance in females. Absence of this observation may be attributed to the timepoint at which memory was assessed versus a terminal microglia assessment. Viral techniques that allow for continuous monitoring of microglia dynamics during physical manipulation may be useful in measuring microglia state throughout an expanded observation window in future studies.
The absence of effects of repeated LPS on behavioral and central endpoints could be reflective of an insufficient LPS dose or development of tolerance to LPS exposure (Mamunur et al., Citation2023). We monitored peripheral signs of LPS-induced changes in physiology and behavior throughout the study and assessed peripheral markers to provide insight on the longitudinal effects of repeated LPS exposure. While we observed some variability in health metrics, no significant changes were observed throughout the study timeline. Chronic stress triggers hypertrophy of the adrenal glands (Horowitz & Zunszain, Citation2015; Ulrich-Lai et al., Citation2006), and LPS can also act directly on adrenal cells (Kanczkowski et al., Citation2013; Vakharia & Hinson, Citation2005). To this end, we observed a decrease in normalized terminal adrenal weight in males that received LPS, regardless of stress history. While we cannot infer a relationship between adrenal function and behavioral outcomes, the data suggest males were sensitive to the impact of repeated LPS on adrenal function. We observed an increase in normalized spleen weight in both non-stressed and stressed males and females that received repeated LPS. This could be due to the known influence of LPS to increase inflammatory mediators within the spleen (Laukova et al., Citation2018). In addition, increased spleen weight at the terminal endpoint suggests that rats were still reponsive to LPS exposure at the end of the 9 week injection scheme. We did not observe increased spleen weight due to stress alone.
Peripheral cytokines may fluctuate in response to stress (Hodes et al., Citation2015), but within the current study there were few changes in terminal peripheral cytokines, with the only significant differences being a decrease in IL-10 and IL-13 observed in stressed compared to non-stressed females. IL-10 is an anti-inflammatory cytokine that is increased in the plasma by LPS (Yan & Kentner, Citation2017) and is necessary to prevent microglia hyperactivation following an LPS challenge (Laffer et al., Citation2019; Shemer et al., Citation2020). Thus, IL-10 may have a neuroprotective role that is lost in the stressed females, however, due to the highly cyclic nature of cytokines, it is unclear if this is reflected in our behavior or microglia data. The pro-inflammatory cytokine IL-13 is upregulated in microglia following LPS administration (Shin et al., Citation2004) and is proposed to contribute to microglia death following injury to prevent chronic neuroinflammation (Yang et al., Citation2002). Decreased circulation of terminal IL-10 and IL-13 plasma levels suggests stressed females were less likely to benefit from the neuroprotective influence of these two cytokines. Investigation of multiple timepoints to fully characterize the impact of repeated LPS on circulating cytokines is needed to draw further conclusions regarding IL-10 or IL-13.
Of note, LPS injections were paused during execution of the Barnes Maze assessment. It is possible that repeated low dose LPS injections resulted in development of endotoxin tolerance (Fraker et al., Citation1988). LPS is recognized by toll-like receptor 4 (TLR4) as part of the innate immune system (Raetz & Whitfield, Citation2002) and in vitro studies demonstrate a macrophage tolerance to repeated LPS challenge demonstrated by a decrease in cytokine release (Biswas & Lopez-Collazo, Citation2009). However, there is limited research on the timeline of tolerance and resurgence of inflammation that can occur upon resuming systemic injections in a rat along the time course used in the current study. Previous work suggests that the stage of estrous cycle during the initial and subsequent LPS injections can influence if tolerance occurs (Engeland et al., Citation2006). In this study, we did not determine estrous stage on each injection day and did not collect blood samples throughout the timeline due to the added stress both procedures would place on the animals. It is possible that the rats did experience tolerance to the repeated LPS administration which could explain the lack of differences within the peripheral cytokines. However, we did observe an effect of LPS on alternation index within the Y-maze which was performed over 50 days after the initiation of the LPS injections and an effect on spleen weights collected at the end of the 9 week exposure period. This suggests that a complete tolerance was not present in males or females. While we cannot definitively determine if a graded endotoxin tolerance may have occurred or if sensitization was evident following the pause in LPS exposures, these caveats should be taken into consideration when interpreting the data presented as assessments were collected over a 9 week longitudinal exposure.
5. Limitations and future studies
Males and females were both vulnerable to CAS and repeated LPS exposure but diverged along the behavioral, neural, and peripheral metrics assessed. The variability in physiology and behavior outcomes may be explained by the temporal response to LPS across the 9 week paradigm. While we did not observe any correlations between terminal estrous stage and mitochondrial outcomes, LPS can influence circulating levels of hormones (Shaw et al., Citation2021) and this could influence the behavior throughout the paradigm. Due to the terminal requirements for assessing many dependent measures within the study, a full interpretation of repeated LPS and hormonal interactions on the outcomes was not feasible. A separate characterization of endotoxin tolerance, peripheral cytokine mediators, and estrous cycling in the absence of a stress paradigm may provide further insight into the overall influence of these factors following repeated LPS in a study in which stressful repeated handling and behavioral outcomes is not of concern. Due to insufficient sample quantity, it was not feasible to analyze synaptosomal mitochondria within the subregions of the hippocampus. As assays become more sensitive, future assessments of regional mitochondrial respiration may better allude to changes across the trisynaptic circuit of the hippocampus. Dynamic in vivo recording of microglial processes over time within the hippocampus will further elucidate the mechanisms by which sexes differ in responses to developmental stress and endotoxin exposures. While out of the scope of the current study, in vivo imaging would allow for simultaneous monitoring of microglia immediately following behavioral assays, increasing interpretability of the relationship.
6. Summary
While we report effects of each a history of CAS and repeated injection of LPS, the data indicate only subtle interactions between these environmental exposures with differential manifestation of phenotype and underlying biological processes for males and females. The metrics investigated were far removed from the experience of the adolescent stressor, suggesting a long-lasting influence of adolescent stress on modifiers of the endocrine and immune system where effects of CAS were observed. Given the wide array of parameters evaluated, there was not a consistent directional influence of stress or repeated LPS exposure in either sex across the study. It is possible that the mechanisms affected are altered along a differential timeline for each metric, even within sex. Collectively, the data suggest that while both sexes demonstrated a deficit in working memory following repeated LPS, females were more impacted by repeated LPS and demonstrated effects in spatial memory, ATP production and microglial morphology. Conversely, males exhibited a greater degree of modification as a result of the history of CAS with evidence of transient increases in anxiety-like behavior and a deficit in spatial memory. The data herein contribute to the growing field of sex variability in response to adolescent stress and vulnerability to repeated inflammatory-related insults, while identifying opportune investigational strategies into repeated immune challenge paradigms.
Supplemental Material
Download MS Word (9.3 MB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
A. J. Wegener
Amy. J. Wegener is a Ph.D. candidate in Neuroscience at Virginia Commonwealth University specializing in the neural consequences of pair bond disruption and inflammation.
M. M. Hyer
Molly M. Hyer, Ph.D., is director of Research Development and Innovation for the Institute of Women’s Health at Virginia Commonwealth University and Program Administrator for Building Interdisciplinary Research Careers in Women’s Health (BIRCWH).
I. Targett
Imogen Targett, completed degrees in biology and neuroscience and is now working as an industry Scientist.
A. Kloster
Alix Kloster completed aher Master of Science in Anatomy & Neurobiology, then obtained her Doctor of Dental Surgery (DDS) from Virginia Commonwealth University.
G. A. Shaw
Gladys Shaw, Ph.D., completed aher Ph.D. in Neuroscience at Virginia Commonwealth University and is currently a Senior Scientist.
A. M. M. Rodriguez
A. M. M. Rodriguez completed aher Bachelor of Science in biology at Virginia Commonwealth University.
S. K. Dyer
Samya K. Dyer, MP H, completed degrees in biology and psychology at Virginia Commonwealth University and holds a master’s in public health.
G. N. Neigh
Gretchen N. Neigh, Ph.D., is a Professor of Anatomy & Neurobiology. Dr. Neigh is director of Translational Research for the VCU Institute of Women’s Health, co-director of research for BIRCWH, and co-director for the Clinical and Translational Sciences PhD Program at Virginia Commonwealth University.
References
- Antonucci, F., Corradini, I., Fossati, G., Tomasoni, R., Menna, E., & Matteoli, M. (2016). SNAP-25, a Known presynaptic protein with emerging postsynaptic functions. Frontiers in Synaptic Neuroscience, 8(MAR), 1. https://doi.org/10.3389/fnsyn.2016.00007
- Basilico, B., Pagani, F., Grimaldi, A., Cortese, B., Di Angelantonio, S., Weinhard, L., Gross, C., Limatola, C., Maggi, L., & Ragozzino, D. (2019). Microglia shape presynaptic properties at developing glutamatergic synapses. Glia, 67(1), 53–18. https://doi.org/10.1002/glia.23508
- Bekhbat, M., Howell, P. A., Rowson, S. A., Kelly, S. D., Tansey, M. G., & Neigh, G. N. (2019). Chronic adolescent stress sex-specifically alters central and peripheral neuro-immune reactivity in rats. Brain, Behavior, and Immunity, 76, 248–257. https://doi.org/10.1016/j.bbi.2018.12.005
- Bekhbat, M., Merrill, L., Kelly, S. D., Lee, V. K., & Neigh, G. N. (2016). Brief anesthesia by isoflurane alters plasma corticosterone levels distinctly in male and female rats: Implications for tissue collection methods. Behavioural Brain Research, 305(404), 122–125. https://doi.org/10.1016/j.bbr.2016.03.003
- Bekhbat, M., Mukhara, D., Dozmorov, M., Stansfield, J., Benusa, S., Hyer, M. M., Rowson, S. A., Kelly, S., Qin, Z., Dupree, J., Tharp, G., Tansey, M. G., & Neigh, G. N. (2020). Adolescent stress sensitizes the adult neuroimmune transcriptome and leads to sex-specific microglial and behavioral phenotypes. Neuropsychopharmacology, 46(5), 949–958. https://doi.org/10.1038/s41386-021-00970-2
- Biswas, S. K., & Lopez-Collazo, E. (2009). Endotoxin tolerance: New mechanisms, molecules and clinical significance. Trends in Immunology, 30(10), 475–487. https://doi.org/10.1016/j.it.2009.07.009
- Bollinger, J. L., Collins, K. E., Patel, R., & Wellman, C. L. (2017). Behavioral stress alters corticolimbic microglia in a sex- and brain region-specific manner. PloS One, 12(12), e0187631. https://doi.org/10.1371/journal.pone.0187631
- Bourgognon, J.-M., & Cavanagh, J. (2020). The role of cytokines in modulating learning and memory and brain plasticity. Brain and Neuroscience Advances, 4, 2398212820979802. https://doi.org/10.1177/2398212820979802
- Bourke, C. H., & Neigh, G. N. (2011). Behavioral effects of chronic adolescent stress are sustained and sexually dimorphic. Hormones and Behavior, 60(1), 112–120. https://doi.org/10.1016/j.yhbeh.2011.03.011
- Brown, G. C. (2019). The endotoxin hypothesis of neurodegeneration. Journal of Neuroinflammation, 16(1), 180. https://doi.org/10.1186/s12974-019-1564-7
- Brzdak, P., Wójcicka, O., Zareba-Koziol, M., Minge, D., Henneberger, C., Wlodarczyk, J., Mozrzymas, J. W., & Wójtowicz, T. (2019). Synaptic potentiation at basal and apical dendrites of hippocampal pyramidal neurons involves activation of a distinct set of extracellular and intracellular molecular cues. Cerebral Cortex (New York, N.Y.: 1991), 29(1), 283–304. https://doi.org/10.1093/cercor/bhx324
- Chaby, L. E., Sheriff, M. J., Hirrlinger, A. M., Lim, J., Fetherston, T. B., & Braithwaite, V. A. (2015). Does chronic unpredictable stress during adolescence affect spatial cognition in adulthood? PloS One, 10(11), e0141908. https://doi.org/10.1371/journal.pone.0141908
- Chaloner, A., & Greenwood-Van Meerveld, B. (2013). Early life adversity as a risk factor for visceral pain in later life: Importance of sex differences. Frontiers in Neuroscience, 7(7 FEB), 13. https://doi.org/10.3389/fnins.2013.00013
- Cohen, S., Janicki-Deverts, D., Doyle, W. J., Miller, G. E., Frank, E., Rabin, B. S., & Turner, R. B. (2012). Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proceedings of the National Academy of Sciences of the United States of America, 109(16), 5995–5999. https://doi.org/10.1073/pnas.1118355109
- Cora, M. C., Kooistra, L., & Travlos, G. (2015). Vaginal cytology of the laboratory rat and mouse: Review and criteria for the staging of the estrous cycle using stained vaginal smears. Toxicologic Pathology, 43(6), 776–793. https://doi.org/10.1177/0192623315570339
- Dunkley, P. R., Jarvie, P. E., & Robinson, P. J. (2008). A rapid percoll gradient procedure for preparation of synaptosomes. Nature Protocols, 3(11), 1718–1728. https://doi.org/10.1038/nprot.2008.171
- Eagleson, K. L., Villaneuva, M., Southern, R. M., & Levitt, P. (2020). Proteomic and mitochondrial adaptations to early-life stress are distinct in juveniles and adults. Neurobiology of Stress, 13, 100251. https://doi.org/10.1016/j.ynstr.2020.100251
- Engeland, C. G., Kavaliers, M., & Ossenkopp, K.-P. (2006). Influence of the estrous cycle on tolerance development to LPS-induced sickness behaviors in rats. Psychoneuroendocrinology, 31(4), 510–525. https://doi.org/10.1016/j.psyneuen.2005.11.007
- Fanselow, M. S., & Dong, H.-W. (2010). Are the dorsal and ventral hippocampus functionally distinct structures? Neuron, 65(1), 7–19. https://doi.org/10.1016/j.neuron.2009.11.031
- File, S. E., & Hyde, J. R. G. (1978). Can social interaction be used to measure anxiety? British Journal of Pharmacology, 62(1), 19–24. https://doi.org/10.1111/j.1476-5381.1978.tb07001.x
- Fraker, D. L., Stovroff, M. C., Merino, M. J., & Norton, J. A. (1988). Tolerance to tumor necrosis factor in rats and the relationship to endotoxin tolerance and toxicity. The Journal of Experimental Medicine, 168(1), 95–105. https://doi.org/10.1084/jem.168.1.95
- Frank, M. G., Baratta, M. V., Sprunger, D. B., Watkins, L. R., & Maier, S. F. (2007). Microglia serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine responses. Brain, Behavior, and Immunity, 21(1), 47–59. https://doi.org/10.1016/j.bbi.2006.03.005
- Frank-Cannon, T. C., Tran, T., Ruhn, K. A., Martinez, T. N., Hong, J., Marvin, M., Hartley, M., Treviño, I., O’Brien, D. E., Casey, B., Goldberg, M. S., & Tansey, M. G. (2008). Parkin deficiency increases vulnerability to inflammation-related nigral degeneration. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 28(43), 10825–10834. https://doi.org/10.1523/JNEUROSCI.3001-08.2008
- Fronza, M. G., Baldinotti, R., Fetter, J., Rosa, S. G., Sacramento, M., Nogueira, C. W., Alves, D., Praticò, D., & Savegnago, L. (2022). Beneficial effects of QTC-4-MeOBnE in an LPS-induced mouse model of depression and cognitive impairments: The role of blood-brain barrier permeability, NF-κB signaling, and microglial activation. Brain, Behavior, and Immunity, 99, 177–191. https://doi.org/10.1016/j.bbi.2021.10.002
- Gaignard, P., Liere, P., Thérond, P., Schumacher, M., Slama, A., & Guennoun, R. (2017). Role of sex hormones on brain mitochondrial function, with special reference to aging and neurodegenerative diseases. Frontiers in Aging Neuroscience, 9(DEC), 406. https://doi.org/10.3389/fnagi.2017.00406
- Galea, L. A. M., McEwen, B. S., Tanapat, P., Deak, T., Spencer, R. L., & Dhabhar, F. S. (1997). Sex differences in dendritic atrophy of CA3 pyramidal neurons in response to chronic restraint stress. Neuroscience, 81(3), 689–697. https://doi.org/10.1016/S0306-4522(97)00233-9
- Gawel, K., Gibula, E., Marszalek-Grabska, M., Filarowska, J., & Kotlinska, J. H. (2019). Assessment of spatial learning and memory in the Barnes maze task in rodents—methodological consideration. Naunyn-Schmiedeberg’s Archives of Pharmacology, 392(1), 1–18. https://doi.org/10.1007/s00210-018-1589-y
- Gu, Y., Ye, T., Tan, P., Tong, L., Ji, J., Gu, Y., Shen, Z., Shen, X., Lu, X., & Huang, C. (2021). Tolerance-inducing effect and properties of innate immune stimulation on chronic stress-induced behavioral abnormalities in mice. Brain, Behavior, and Immunity, 91, 451–471. https://doi.org/10.1016/j.bbi.2020.11.002
- Guo, L., Tian, J., & Du, H. (2017). Mitochondrial dysfunction and synaptic transmission failure in Alzheimer’s disease. Journal of Alzheimer’s Disease: JAD, 57(4), 1071–1086. https://doi.org/10.3233/JAD-160702
- Han, S., He, Z., Jacob, C., Hu, X., Liang, X., Xiao, W., Wan, L., Xiao, P., D’Ascenzo, N., Ni, J., Liu, Q., & Xie, Q. (2021). Effect of increased il-1β on expression of hk in Alzheimer’s disease. International Journal of Molecular Sciences, 22(3), 1–17. https://doi.org/10.3390/ijms22031306
- Haroon, E., Miller, A. H., & Sanacora, G. (2017). Inflammation, glutamate, and glia: A trio of trouble in mood disorders. Neuropsychopharmacology: official Publication of the American College of Neuropsychopharmacology, 42(1), 193–215. https://doi.org/10.1038/npp.2016.199
- Hauss-Wegrzyniak, B., Vraniak, P. D., & Wenk, G. L. (2000). LPS-induced neuroinflammatory effects do not recover with time. Neuroreport, 11(8), 1759–1763. https://doi.org/10.1097/00001756-200006050-00032
- Heim, C., Newport, D. J., Mletzko, T., Miller, A. H., & Nemeroff, C. B. (2008). The link between childhood trauma and depression: Insights from HPA axis studies in humans. Psychoneuroendocrinology, 33(6), 693–710. https://doi.org/10.1016/j.psyneuen.2008.03.008
- Hellwig, S., Brioschi, S., Dieni, S., Frings, L., Masuch, A., Blank, T., & Biber, K. (2016). Altered microglia morphology and higher resilience to stress-induced depression-like behavior in CX3CR1-deficient mice. Brain, Behavior, and Immunity, 55, 126–137. https://doi.org/10.1016/j.bbi.2015.11.008
- Harland, M., Torres, S., Liu, J., & Wang, X. (2020). Neuronal mitochondria modulation of LPS-induced neuroinflammation. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 40(8), 1756–1765. https://doi.org/10.1523/JNEUROSCI.2324-19.2020
- Hinwood, M., Morandini, J., Day, T. A., & Walker, F. R. (2012). Evidence that microglia mediate the neurobiological effects of chronic psychological stress on the medial prefrontal cortex. Cerebral Cortex (New York, N.Y.: 1991), 22(6), 1442–1454. https://doi.org/10.1093/cercor/bhr229
- Hodes, G. E., Kana, V., Menard, C., Merad, M., & Russo, S. J. (2015). Neuroimmune mechanisms of depression. Nature Neuroscience, 18(10), 1386–1393. https://doi.org/10.1038/nn.4113
- Horowitz, M. A., & Zunszain, P. A. (2015). Neuroimmune and neuroendocrine abnormalities in depression: Two sides of the same coin. Annals of the New York Academy of Sciences, 1351(1), 68–79. https://doi.org/10.1111/nyas.12781
- Hyer, M. M., Wegener, A. J., Targett, I., Dyer, S. K., & Neigh, G. N. (2023). Chronic stress beginning in adolescence decreases spatial memory following an acute inflammatory challenge in adulthood. Behavioural Brain Research, 442, 114323. https://doi.org/10.1016/j.bbr.2023.114323
- Hyer, M. M., Shaw, G. A., Goswamee, P., Dyer, S. K., Burns, C. M., Soriano, E., Sanchez, C. S., Rowson, S. A., McQuiston, A. R., & Neigh, G. N. (2021). Chronic adolescent stress causes sustained impairment of cognitive flexibility and hippocampal synaptic strength in female rats. Neurobiology of Stress, 14, 100303. https://doi.org/10.1016/j.ynstr.2021.100303
- Ivens, S., Çalışkan, G., Papageorgiou, I., Cesetti, T., Malich, A., Kann, O., Heinemann, U., Stork, O., & Albrecht, A. (2019). Persistent increase in ventral hippocampal long-term potentiation by juvenile stress: A role for astrocytic glutamine synthetase. Glia, 67(12), 2279–2293. https://doi.org/10.1002/glia.23683
- Kanczkowski, W., Chatzigeorgiou, A., Samus, M., Tran, N., Zacharowski, K., Chavakis, T., & Bornstein, S. R. (2013). Characterization of the LPS-induced inflammation of the adrenal gland in mice. Molecular and Cellular Endocrinology, 371(1-2), 228–235. https://doi.org/10.1016/j.mce.2012.12.020
- Keirns, B. H., Keirns, N. G., Tsotsoros, C. E., Layman, H. M., Stout, M. E., Medlin, A. R., Sciarrillo, C. M., Teague, T. K., Emerson, S. R., & Hawkins, M. A. W. (2023). Adverse childhood experiences and obesity linked to indicators of gut permeability and inflammation in adult women. Physiology & Behavior, 271, 114319. https://doi.org/10.1016/j.physbeh.2023.114319
- Kessler, R. C., McGonagle, K. A., Zhao, S., Nelson, C. B., Hughes, M., Eshleman, S., Wittchen, H. U., & Kendler, K. S. (1994). Lifetime and 12-month prevalence of DSM-III-R psychiatric disorders in the United States. Archives of General Psychiatry, 51(1), 8–19. https://doi.org/10.1001/archpsyc.1994.03950010008002
- Kim, J. D., Yoon, N. A., Jin, S., & Diano, S. (2019). Microglial UCP2 mediates inflammation and obesity induced by high-fat feeding. Cell Metabolism, 30(5), 952–962.e5. https://doi.org/10.1016/j.cmet.2019.08.010
- Kitazawa, M., Oddo, S., Yamasaki, T. R., Green, K. N., & LaFerla, F. M. (2005). Lipopolysaccharide-induced inflammation exacerbates Tau pathology by a cyclin-dependent kinase 5-mediated pathway in a transgenic model of Alzheimer’s disease. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 25(39), 8843–8853. https://doi.org/10.1523/JNEUROSCI.2868-05.2005
- Kloster, A., Hyer, M. M., Dyer, S., Salome-Sanchez, C., & Neigh, G. N. (2020). High fructose diet induces sex-specific modifications in synaptic respiration and affective-like behaviors in rats. Neuroscience, 454, 40–50. https://doi.org/10.1016/j.neuroscience.2019.11.039
- Kraeuter, A.-K., Guest, P. C., & Sarnyai, Z. (2019). The Y-maze for assessment of spatial working and reference memory in mice. In Methods in molecular biology (Clifton, N.J.) (pp. 105–111). https://doi.org/10.1007/978-1-4939-8994-2_10
- Laffer, B., Bauer, D., Wasmuth, S., Busch, M., Jalilvand, T. V., Thanos, S., Meyer zu Hörste, G., Loser, K., Langmann, T., Heiligenhaus, A., & Kasper, M. (2019). Loss of IL-10 promotes differentiation of microglia to a M1 phenotype. Frontiers in Cellular Neuroscience, 13(October), 430. https://doi.org/10.3389/fncel.2019.00430
- Laroche, J., Gasbarro, L., Herman, J. P., & Blaustein, J. D. (2009). Reduced behavioral response to gonadal hormones in mice shipped during the peripubertal/adolescent period. Endocrinology, 150(5), 2351–2358. https://doi.org/10.1210/en.2008-1595
- Laukova, M., Vargovic, P., Rokytova, I., Manz, G., & Kvetnansky, R. (2018). Repeated stress exaggerates lipopolysaccharide-induced inflammatory response in the rat spleen. Cellular and Molecular Neurobiology, 38(1), 195–208. https://doi.org/10.1007/s10571-017-0546-5
- La-Vu, M., Tobias, B. C., Schuette, P. J., & Adhikari, A. (2020). To approach or avoid: An introductory overview of the study of anxiety using rodent assays. Frontiers in Behavioral Neuroscience, 14, 145. https://doi.org/10.3389/fnbeh.2020.00145
- Liddelow, S. A., & Barres, B. A. (2017). Reactive astrocytes: Production, function, and therapeutic potential. Immunity, 46(6), 957–967. https://doi.org/10.1016/j.immuni.2017.06.006
- Liu, Y.-Z., Wang, Y.-X., & Jiang, C.-L. (2017). Inflammation: The common pathway of stress-related diseases. Frontiers in Human Neuroscience, 11, 316. https://doi.org/10.3389/fnhum.2017.00316
- Liu, T., Lu, J., Lukasiewicz, K., Pan, B., & Zuo, Y. (2021). Stress induces microglia-associated synaptic circuit alterations in the dorsomedial prefrontal cortex. Neurobiology of Stress, 15(May), 100342. https://doi.org/10.1016/j.ynstr.2021.100342
- Liu, T., Zhang, L., Joo, D., & Sun, S. C. (2017). NF-κB signaling in inflammation. Signal Transduction and Targeted Therapy, 2(1), 17023–17023. https://doi.org/10.1038/sigtrans.2017.23
- Mamunur, R., Hashioka, S., Azis, I. A., Jaya, M. A., Jerin, S. J. F., Kimura-Kataoka, K., Fujihara, J., Inoue, K., Inagaki, M., & Takeshita, H. (2023). Systemic administration of porphyromonas gingivalis lipopolysaccharide induces glial activation and depressive-like behavior in rats. Journal of Integrative Neuroscience, 22(5), 120. https://doi.org/10.31083/j.jin2205120
- Miller, G. E., Cohen, S., & Ritchey, A. K. (2002). Chronic psychological stress and the regulation of pro-inflammatory cytokines: A glucocorticoid-resistance model. Health Psychology: official Journal of the Division of Health Psychology, American Psychological Association, 21(6), 531–541. https://doi.org/10.1037/0278-6133.21.6.531
- Nasca, C., Bigio, B., Zelli, D., de Angelis, P., Lau, T., Okamoto, M., Soya, H., Ni, J., Brichta, L., Greengard, P., Neve, R. L., Lee, F. S., & McEwen, B. S. (2017). Role of the astroglial glutamate exchanger xCT in ventral hippocampus in resilience to stress. Neuron, 96(2), 402–413.e5. https://doi.org/10.1016/j.neuron.2017.09.020
- Noailles, A., Maneu, V., Campello, L., Lax, P., & Cuenca, N. (2018). Systemic inflammation induced by lipopolysaccharide aggravates inherited retinal dystrophy. Cell Death & Disease, 9(3), 350. https://doi.org/10.1038/s41419-018-0355-x
- Ogawa, T., Kuwagata, M., Hori, Y., & Shioda, S. (2007). Valproate-induced developmental neurotoxicity is affected by maternal conditions including shipping stress and environmental change during early pregnancy. Toxicology Letters, 174(1–3), 18–24. https://doi.org/10.1016/j.toxlet.2007.08.006
- Olesen, M. A., Torres, A. K., Jara, C., Murphy, M. P., & Tapia-Rojas, C. (2020). Premature synaptic mitochondrial dysfunction in the hippocampus during aging contributes to memory loss. Redox Biology, 34(May), 101558. https://doi.org/10.1016/j.redox.2020.101558
- Paxinos, G., & Watson, C. (2007). The rat brain in stereotaxic coordinates. Elsevier. https://books.google.com/books?id=Tp5QlAEACAAJ
- Picard, M., McManus, M. J., Gray, J. D., Nasca, C., Moffat, C., Kopinski, P. K., Seifert, E. L., McEwen, B. S., & Wallace, D. C. (2015). Mitochondrial functions modulate neuroendocrine, metabolic, inflammatory, and transcriptional responses to acute psychological stress. Proceedings of the National Academy of Sciences of the United States of America, 112(48), E6614–E6623. https://doi.org/10.1073/pnas.1515733112
- Piccinelli, M., & Wilkinson, G. (2000). Gender differences in depression. The British Journal of Psychiatry: The Journal of Mental Science, 177(6), 486–492. https://doi.org/10.1192/bjp.177.6.486
- Pyter, L. M., Kelly, S. D., Harrell, C. S., & Neigh, G. N. (2013). Sex differences in the effects of adolescent stress on adult brain inflammatory markers in rats. Brain, Behavior, and Immunity, 30, 88–94. https://doi.org/10.1016/j.bbi.2013.01.075
- Raetz, C. R. H., & Whitfield, C. (2002). Lipopolysaccharide endotoxins. Annual Review of Biochemistry, 71(1), 635–700. https://doi.org/10.1146/annurev.biochem.71.110601.135414
- Robey, R. B., & Hay, N. (2006). Mitochondrial hexokinases, novel mediators of the antiapoptotic effects of growth factors and Akt. Oncogene, 25(34), 4683–4696. https://doi.org/10.1038/sj.onc.1209595
- Rohleder, N., Schommer, N. C., Hellhammer, D. H., Engel, R., & Kirschbaum, C. (2001). Sex differences in glucocorticoid sensitivity of proinflammatory cytokine production after psychosocial stress. Psychosomatic Medicine, 63(6), 966–972. https://doi.org/10.1097/00006842-200111000-00016
- Rowson, S. A., Bekhbat, M., Kelly, S. D., Binder, E. B., Hyer, M. M., Shaw, G., Bent, M. A., Hodes, G., Tharp, G., Weinshenker, D., Qin, Z., & Neigh, G. N. (2019). Chronic adolescent stress sex-specifically alters the hippocampal transcriptome in adulthood. Neuropsychopharmacology: official Publication of the American College of Neuropsychopharmacology, 44(7), 1207–1215. https://doi.org/10.1038/s41386-019-0321-z
- Rudzki, L., & Szulc, A. (2018). “Immune Gate” of psychopathology—the role of gut derived immune activation in major psychiatric disorders. Frontiers in Psychiatry, 9, 205. https://doi.org/10.3389/fpsyt.2018.00205
- Sampath, D., Sathyanesan, M., & Newton, S. S. (2017). Cognitive dysfunction in major depression and Alzheimer’s disease is associated with hippocampal–prefrontal cortex dysconnectivity. Neuropsychiatric Disease and Treatment, 13, 1509–1519. https://doi.org/10.2147/NDT.S136122
- Sanguino-Gómez, J., Buurstede, J. C., Abiega, O., Fitzsimons, C. P., Lucassen, P. J., Eggen, B. J. L., Lesuis, S. L., Meijer, O. C., & Krugers, H. J. (2021). An emerging role for microglia in stress-effects on memory. The European Journal of Neuroscience, 55(9–10), 2491–2518. https://doi.org/10.1111/ejn.15188
- Schindler, A., & Foley, E. (2013). Hexokinase 1 blocks apoptotic signals at the mitochondria. Cellular Signalling, 25(12), 2685–2692. https://doi.org/10.1016/j.cellsig.2013.08.035
- Schneider, C. A., Rasband, W. S., & Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 Years of image analysis. Nature Methods, 9(7), 671–675. https://doi.org/10.1038/nmeth.2089
- Shaw, G. A., Hyer, M. M., Dustin, E., Dyer, S. K., Targett, I. L., & Neigh, G. N. (2021). Acute LPS exposure increases synaptosomal metabolism during estrus but not diestrus. Physiology & Behavior, 239, 113523. https://doi.org/10.1016/j.physbeh.2021.113523
- Shaw, G. A. (2021). Mitochondria as the target for disease related hormonal dysregulation. Brain, Behavior, & Immunity - Health, 18(September), 100350. https://doi.org/10.1016/j.bbih.2021.100350
- Shaw, G. A., Hyer, M. M., Targett, I., Council, K. R., Dyer, S. K., Turkson, S., Burns, C. M., & Neigh, G. N. (2020). Traumatic stress history interacts with sex and chronic peripheral inflammation to alter mitochondrial function of synaptosomes. Brain, Behavior, and Immunity, 88, 203–219. https://doi.org/10.1016/j.bbi.2020.05.021
- Shemer, A., Scheyltjens, I., Frumer, G. R., Kim, J. S., Grozovski, J., Ayanaw, S., Dassa, B., Van Hove, H., Chappell-Maor, L., Boura-Halfon, S., Leshkowitz, D., Mueller, W., Maggio, N., Movahedi, K., & Jung, S. (2020). Interleukin-10 prevents pathological microglia hyperactivation following peripheral endotoxin challenge. Immunity, 53(5), 1033–1049.e7. https://doi.org/10.1016/j.immuni.2020.09.018
- Shin, W. H., Lee, D. Y., Park, K. W., Kim, S. U., Yang, M. S., Joe, E. H., & Jin, B. K. (2004). Microglia expressing interleukin-13 undergo cell death and contribute to neuronal survival in vivo. Glia, 46(2), 142–152. https://doi.org/10.1002/glia.10357
- Shors, T. J., Chua, C., & Falduto, J. (2001). Sex differences and opposite effects of stress on dendritic spine density in the male versus female hippocampus. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 21(16), 6292–6297. https://doi.org/10.1523/jneurosci.21-16-06292.2001
- Sierra, A., Gottfried-Blackmore, A., Milner, T. A., McEwen, B. S., & Bulloch, K. (2008). Steroid hormone receptor expression and function in microglia. Glia, 56(6), 659–674. https://doi.org/10.1002/glia.20644
- Tagliari, B., Noschang, C. G., Ferreira, A. G. K., Ferrari, O. A., Feksa, L. R., Wannmacher, C. M. D., Dalmaz, C., & Wyse, A. T. S. (2010). Chronic variable stress impairs energy metabolism in prefrontal cortex and hippocampus of rats: Prevention by chronic antioxidant treatment. Metabolic Brain Disease, 25(2), 169–176. https://doi.org/10.1007/s11011-010-9194-x
- Tata, D. A., Marciano, V. A., & Anderson, B. J. (2006). Synapse loss from chronically elevated glucocorticoids: Relationship to neuropil volume and cell number in hippocampal area CA3. The Journal of Comparative Neurology, 498(3), 363–374. https://doi.org/10.1002/cne.21071
- Tynan, R. J., Naicker, S., Hinwood, M., Nalivaiko, E., Buller, K. M., Pow, D. V., Day, T. A., & Walker, F. R. (2010). Chronic stress alters the density and morphology of microglia in a subset of stress-responsive brain regions. Brain, Behavior, and Immunity, 24(7), 1058–1068. https://doi.org/10.1016/j.bbi.2010.02.001
- Ulrich-Lai, Y. M., Figueiredo, H. F., Ostrander, M. M., Choi, D. C., Engeland, W. C., & Herman, J. P. (2006). Chronic stress induces adrenal hyperplasia and hypertrophy in a subregion-specific manner. American Journal of Physiology. Endocrinology and Metabolism, 291(5), E965–973. https://doi.org/10.1152/ajpendo.00070.2006
- Vakharia, K., & Hinson, J. P. (2005). Lipopolysaccharide directly stimulates cortisol secretion by human adrenal cells by a cyclooxygenase-dependent mechanism. Endocrinology, 146(3), 1398–1402. https://doi.org/10.1210/en.2004-0882
- Walker, F., Nilsson, M., & Jones, K. (2013). Acute and chronic stress-induced disturbances of microglial plasticity, phenotype and function. Current Drug Targets, 14(11), 1262–1276. https://doi.org/10.2174/13894501113149990208
- Wohleb, E. S., Terwilliger, R., Duman, C. H., & Duman, R. S. (2018). Stress-induced neuronal colony stimulating factor 1 provokes microglia-mediated neuronal remodeling and depressive-like behavior. Biological Psychiatry, 83(1), 38–49. https://doi.org/10.1016/j.biopsych.2017.05.026
- Woodburn, S. C., Bollinger, J. L., & Wohleb, E. S. (2021). The semantics of microglia activation: Neuroinflammation, homeostasis, and stress. Journal of Neuroinflammation, 18(1), 258. https://doi.org/10.1186/s12974-021-02309-6
- Woolley, C. S., Gould, E., & McEwen, B. S. (1990). Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Research, 531(1–2), 225–231. https://doi.org/10.1016/0006-8993(90)90778-A
- Xin, Y. R., Jiang, J. X., Hu, Y., Pan, J. P., Mi, X. N., Gao, Q., Xiao, F., Zhang, W., & Luo, H. M. (2019). The immune system drives synapse loss during lipopolysaccharide-induced learning and memory impairment in mice. Frontiers in Aging Neuroscience, 11(November), 279. https://doi.org/10.3389/fnagi.2019.00279
- Yan, S., & Kentner, A. C. (2017). Mechanical allodynia corresponds to Oprm1 downregulation within the descending pain network of male and female rats exposed to neonatal immune challenge. Brain, Behavior, and Immunity, 63, 148–159. https://doi.org/10.1016/j.bbi.2016.10.007
- Yang, M. S., Park, E. J., Sohn, S., Kwon, H. J., Shin, W. H., Pyo, H. K., Jin, B., Choi, K. S., Jou, I., & Joe, E. H. (2002). Interleukin-13 and -4 induce death of activated microglia. Glia, 38(4), 273–280. https://doi.org/10.1002/glia.10057
- Zhao, W., Xu, Z., Cao, J., Fu, Q., Wu, Y., Zhang, X., Long, Y., Zhang, X., Yang, Y., Li, Y., & Mi, W. (2019). Elamipretide (SS-31) improves mitochondrial dysfunction, synaptic and memory impairment induced by lipopolysaccharide in mice. Journal of Neuroinflammation, 16(1), 230. https://doi.org/10.1186/s12974-019-1627-9
- Zisook, S., Planeta, B., Hicks, P. B., Chen, P., Davis, L. L., Villarreal, G., Sapra, M., Johnson, G. R., & Mohamed, S. (2022). Childhood adversity and adulthood major depressive disorder. General Hospital Psychiatry, 76(February), 36–44. https://doi.org/10.1016/j.genhosppsych.2022.03.008