Abstract
We examined the ontogeny of the corticosterone response to (+)-methamphetamine in neonatal rats. In experiment-1, animals were injected with 10 mg/kg of (+)-methamphetamine or saline and plasma corticosterone levels were examined in separate groups 30 or 105 min later on postnatal day (P) 1, 3, 5, 7, 9, 11, 13, 15, 17, or 19. The adrenal response to methamphetamine was best described by a U-shaped function with the nadir of corticosterone release occurring between P7 and P13. Experiment-2 was similar except that the effect of four consecutive days of exposure to (+)-methamphetamine (four times daily at 2 h intervals with 10 mg/kg) was assessed with a single final dose early on the fifth day (i.e. P1–5, 3–7, 5–9, 7–11, 9–13, 11–15, 13–17, 15–19). The 30 min corticosterone response after multiple methamphetamine doses was augmented compared to single exposures, with the exception of the two earliest dosing intervals ending on P5 and P7, where the responses were lower. In addition, at 105 min, the levels of corticosterone were attenuated relative to a single drug administration. With the exception of animals receiving methamphetamine from P15 to P19, thymus weights were unaffected. The data demonstrate that (+)-methamphetamine is a robust activator of corticosterone release in developing animals and this release is extensively modified by age and previous drug exposure.
Introduction
Methamphetamine (MA) is a commonly used/abused psychostimulant of the phenethylamine family. Clinically, MA is prescribed for such disorders as attention deficit disorder, obesity, or narcolepsy, however the drug has a high abuse potential. Following administration of MA, monoamine transporters for dopamine, norepinephrine and serotonin are blocked and these transporters are reversed from the normal direction of action so that now monoamines are carried from the cytoplasm to the synaptic cleft, thus leading to increased release of monoamines (Kuczenski and Segal Citation1994). A similar reversal of the vesicular monoamine transporter occurs such that the synaptic vesicles are emptied into the cytoplasm (Seiden et al. Citation1993). Increased heart rate, blood pressure, respiration and adrenal cortical output has also been demonstrated in human users (Fehm et al. Citation1984; Seiden et al. Citation1993; Gouzoulis-Mayfrank et al. Citation1999). Interestingly, in humans it has been suggested that MA may not increase cortisol release through an ACTH-mediated mechanism (Fehm et al. Citation1984), although in rats increased ACTH has been observed concomitant with increased corticosterone levels following MA administration (Williams et al. Citation2000).
MA abuse, especially prevalent among young adults, is a worldwide problem (Chomchai et al. Citation2004; EMCDDA Citation2004; Johnston et al. Citation2004). While there is concern for the long-term neurological and cognitive effects produced by MA in the abusers (Ernst et al. Citation2000; Kalechstein et al. Citation2003; Thompson et al. Citation2004), another concern is that a proportion of the women abusing MA will become pregnant, expose their developing fetuses to the drug, and subsequently this exposure to MA will induce neurological and cognitive damage. The long-term effects of MA exposure in utero are only now being appreciated. For example, a preliminary magnetic resonance imaging study showed altered brain development in children exposed to MA that included reduced hippocampal volume, as well as associated cognitive deficiencies (Chang et al. Citation2004). Importantly, abuse of MA has been shown to continue throughout the entire pregnancy (Smith et al. Citation2003; Chomchai et al. Citation2004), similar to other abused psychostimulants, such as cocaine (Richardson et al. Citation1999) or nicotine (Lindqvist et al. Citation2002). In order to model the impact of late gestational exposure to MA in humans, we have used the neonatal rat since regional brain neurogenesis (e.g. dentate granule cells, cerebellum and olfactory bulbs) and development of some neurotransmitter systems is comparable between the two species at these ages (Bayer et al. Citation1993; Herlenius and Lagercrantz Citation2004). Furthermore, during the third trimester in humans, basal circulating cortisol levels are normal or below normal, but blunted following stimulation (Brosnan Citation2001). A similar attenuated adrenal response for corticosterone occurs in rats during approximately the first two weeks of the postnatal period and has been commonly referred to as the stress hyporesponsive period (SHRP) (Sapolsky and Meaney Citation1986; Vázquez Citation1998). These data when taken together demonstrate the developmental similarities between neonatal rats and fetal humans (Bayer et al. Citation1993; Clancy et al. Citation2001, also see, www.translatingtime.net).
We have demonstrated that MA administered during the neonatal period from postnatal day (P) 11–20 in rats produces long-term Morris water maze (MWM) deficits, a test of hippocampus-dependent learning and memory. No differences in MWM learning were detected when drug was administered from P1 to P10 (Vorhees et al. Citation1994). This is not to say that the P1–10 administration was without effect, since body weight deficits persisted in these animals into adulthood and they displayed enhanced acoustic startle reactivity relative to controls (Vorhees et al. Citation1994). The initial study of P11–20 dosing was performed with a two-dose per day (2 × ) drug administration paradigm. A later study showed greater deficits in MWM learning when a 4 dose per day (4 × ) drug administration regimen was used (Vorhees et al. Citation2000) even though total daily MA dosage was identical in the 2 × and 4 × groups. These cognitive deficits in the MWM are not the result of working memory deficits (Williams et al. Citation2003c) and generalize to other spatial tasks such as the Barnes maze (Williams et al. Citation2003a). In conjunction with the learning and memory deficits, we have also demonstrated that there is an altered hypothalamic-pituitary-adrenal (HPA) axis response in these animals in adulthood (Williams et al. Citation2003a).
The mechanism for these cognitive and physiological alterations is unknown. In adult animals, a 4x regimen of MA (10 mg/kg) on a single day has been demonstrated to induce hyperthermia, reduce dopamine and serotonin levels in the striatum, and produce neurotoxicity as demonstrated by reactive gliosis, silver staining, and Fluoro-Jade staining when examined 3 days after drug administration (O'Callaghan and Miller Citation2002). Contrary to adult animals, P20 neonates show neither hyperthermia, reductions in dopamine, nor neurotoxicity when examined 3 days later (Cappon et al. Citation1997). Animals dosed with MA from P11 to P20 did however show differences in the dopaminergic system in the striatum when examined in adulthood (Crawford et al. Citation2003). Nevertheless, these changes are unlikely to explain the long-term spatial learning and memory deficits, since small reductions in monoamines following neonatal drug exposure have not been correlated with learning and memory deficits (Broening et al. Citation2001). We have also shown that MA administration on P11 produces a protracted increase in corticosterone, even during the SHRP, although only short durations of corticosterone increases were observed after multiple days of drug administration when examined on P15 or P20 (Williams et al. Citation2000). Based on these observations, we hypothesized that the adrenal release of corticosterone may be an important determinant in the induction of learning and memory deficits after neonatal MA administration.
The present experiment was designed to examine the ontogeny of the acute adrenal response to MA during the neonatal period and to determine if any differences in adrenal response existed between the two periods of drug administration used in previous studies (i.e. P1–10 or P11–20). Differences might provide some insight for the different patterns of behavioral changes following P1–10 MA administration compared to P11–20 MA administration. In the first experiment, the adrenal response to MA was examined every other day beginning on P1 until P19. The second experiment examined the effects of a chronic dosing regimen prior to a final acute dose of MA. We hypothesized that MA exposure would alter the response of the adrenal gland to MA on subsequent days. We chose a 5-day dosing regimen since in a previous study we demonstrated that the critical period for learning and memory deficits can be refined to a five-day window, P11–15 within the P11–20 administration period, with no learning effects after P16–20 exposure (Williams et al. Citation2003b).
Materials and methods
Subjects
Nulliparous female and male Sprague-Dawley CD IGS rats (Charles River Laboratories, Raleigh, NC) were mated in hanging wire cages following an acclimation period to the vivarium of at least one week. The day a sperm plug was detected was considered embryonic day 0 (EO). Fourteen days after being placed with the males, females were transferred to polycarbonate cages (45.7 × 23.8 × 20.3 cm). The offspring were the test subjects for these experiments. Food and water were freely available to the animals throughout the experiment. The vivarium was maintained on a 14 h light/10 h dark cycle (lights on at 0600 h) and was fully accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care (AAALAC). The Cincinnati Children's Research Foundation's Laboratory Animal Care and Use Committee approved all protocols for these experiments.
Beginning on E21 females were checked for a litter twice daily, once in the morning and once in the afternoon. The day a litter was detected was defined as postnatal day 0 (P0). On P1, the litters were standardized to 12 pups with at least four males in the litter. If a litter did not have 12 animals, up to two same-age pups were fostered to the litter if available. The identification of individual pups, when required for extended dosing paradigms, was by an India ink tattoo on the footpad prior to P7 or by an ear punch after P7. On the day of assessment, readily visible identifications were made with a permanent black marker at the time of drug administration to reduce the disruption to the litter when a pup was removed. All decapitations were done using large stainless steel scissors and no anesthesia was administered prior to decapitation (Holson Citation1992).
Drug administration
(+)-MA HCl (greater than 95% pure, National Institute of Drug Abuse, Bethesda, MD) was expressed as the freebase and dissolved in sterile physiological (0.9% w/vol) saline at a concentration of 10 mg/3 ml and was delivered at a dosage of 10 mg/kg. Isotonic saline (SAL) was administered at 3 ml/kg to controls. This dose of MA was used since it produces low mortality during the neonatal period and when administered over multiple days induces spatial learning and memory deficits (Vorhees et al. Citation2000; Williams et al. Citation2002, Citation2003b,Citationc) as well as neuronal cytoarchitectural changes in the dentate gyrus and nucleus accumbens (Williams et al. Citation2004a). As in our previous studies, MA administration was through a subcutaneous route in the dorsum, with injection sites alternated to prevent irritation to the dermis. All animals in a litter were treated identically. For younger animals, a Hamilton syringe was used to deliver drug and Vaseline was applied to the site of injection prior to administration to minimize leakage. The animals were weighed preceding each drug administration. In order to control for any potential circadian and ultradian variations during the sampling procedure, all drug administrations were between 1230 and 1330 h.
Experiment 1
Corticosterone responses of animals were tested every other day beginning on P1 and ending on P19 (e.g. P1, 3, 5, 7, 9, 11, 13, 15, 17 and 19). Samples were collected at one of two time points, 30 or 105 min, following either MA or SAL administration. On each day of testing, two animals were randomly selected from a litter and both were injected with either MA or SAL, marked for identification, and placed back in the litter until trunk blood was collected. The 105 min time point was selected based on a previous study where we demonstrated that after multiple doses of MA from P11 to P15, the corticosterone response of animals returned to baseline by this time (Williams et al. Citation2000). Only males were used on P17 and P19 as a precaution to reduce any potential sexually dimorphic variability in corticosterone response, although it should be noted that we have not observed any sex differences in corticosterone release on either P11, P15, or P20 following MA administration (Williams et al. Citation2000) or on P11 following MDMA administration (Williams et al. Citation2005). The use of both male and female pups has been used extensively before because of the lack of differences in the adrenal response at these ages to various manipulations (for example, see Walker et al. Citation1991; Walker and Dallman Citation1993; Kent et al. Citation1996; McCormick et al. Citation1998). Therefore, taken together with our previous data we used males and females in both experiments. For Experiment 1, on each day at least 10 litters were represented with the exception of P5, P9 and P11 that had 20 litters represented for a total of 260 pups.
Experiment 2
This experiment was designed to assess the corticosterone response following multiple exposures to MA. We used a five-day paradigm in this experiment since we have shown that P11–15 MA administration is effective at inducing learning and memory deficits, whereas P16–20 is not (Williams et al. Citation2003b). As in Experiment 1, animals were started on drug administration on either P1, 3, 5, 7, 9, 11, 13, or 15. However, rather than collecting trunk blood on the first day, it was collected on the fifth day following a single morning drug administration. For the first 4 days of each dosing paradigm, animals were administered either MA or SAL, four times daily at two hour intervals. On the last day of drug administration, only one dose of MA was given, comparable to Experiment 1. Trunk blood was then collected 30 or 105 min after the last dose. Only males were used when dosing began after P13 (i.e. tissue collected on P17 and P19) to be consistent with Experiment 1. In this experiment, on each day at least 8–10 litters were represented for a total of 146 pups.
Corticosterone assessment and thymus weights
All trunk blood was collected in polyethylene tubes containing 0.05 ml of 2% EDTA. The blood was centrifuged (1399 RCF) for 25 min at 4°C, plasma collected, and stored at − 80°C until assayed. Plasma was diluted either 2:1 or 3:1 in assay buffer prior to the determination of corticosterone, using a commercially available EIA (IDS, Inc., Fountain Hills, AZ). This EIA has little cross-reactivity with other steroidal hormones, precursors, or metabolites.
Previous studies have shown that long-term exposure to stressors in adult animals can cause involution of the thymus gland (Kioukia-Fougia et al. Citation2002), however it is not known if this occurs in neonatal rats following MA. Therefore, after trunk blood was collected, the thymus was removed, freed of fatty tissue, and weighed. Thymus weights were expressed as a percentage of body weight for analysis.
Statistics
The corticosterone data were analyzed with a mixed-model analysis of variance (ANOVA) utilizing PROC GLM in SAS (SAS Institute, Cary, NC) for each experiment. The between-subject effects were Day of corticosterone assessment and Treatment (MA or SAL) and the within-subject factor was Time of sampling. Litter membership was treated as a matching factor, therefore, Time of sampling on each day of assessment was handled in the ANOVA as a repeated measures factor (Winer Citation1971; Kirk Citation1995). Body and thymus weights were assessed by between-subjects ANOVA and the mean values for animals from each litter on each day were used. On each day of assessment, litter was considered the unit of measurement (Holson and Pearce Citation1992) and a total of 8–20 litters were represented at each time point. When an analysis contained a within factor, the Greenhouse-Geisser correction was used when the symmetry of the variance-covariance matrices were significantly non-spherical. Simple effect ANOVAs were used to further analyze significant interactions. In rare cases where a value was over five standard deviations from the mean of the group, the data from that animal were removed from the analysis. Significance was set at p ≤ 0.05, and data are presented as means ± standard error of the mean (SEM). For simplicity of presentation, only the significant Treatment main effects or the highest order interactions with Treatment have F ratios presented.
Results
Experiment 1
An ANOVA of body weights of pups on each of the days revealed no difference prior to drug administration between the MA and SAL animals, as expected. An overall analysis of plasma corticosterone concentration was initially performed with Treatment, Day and Time. All main effects and interactions were significant for this analysis at p < 0.0001 with the exception of Time that was at p < 0.002. Since the Treat × Day × Time interaction was significant, F(9,239) = 12.83, p < 0.0001, simple effect analyses were performed on each day for Treatment and Time ().
Figure 1 Plasma concentrations of corticosterone following a single administration of 10 mg/kg of MA or SAL on the given postnatal days. The animals were decapitated at 30 min (left panel) or 105 min (right panel) after the administration of MA. The levels of corticosterone were dependent upon the treatment the animals received, the age that they received treatment, and the time the corticosterone levels were examined. * p < 0.05 for MA vs. SAL.
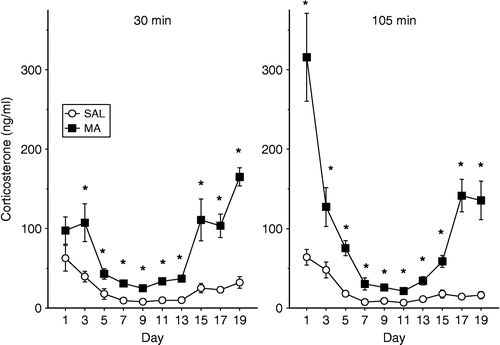
On P1, the interaction between Treatment and Time, F(1,18) = 19.43, p < 0.0003 was the result of no difference between treatments at 30 min, but at 105 min the MA-treated animals showed a significant increase in corticosterone. On P3, the MA-treated animals had higher levels of corticosterone than the SAL-treated animals, F(1,18) = 11.57, p < 0.004. On P5, even though there was a Treatment and Time interaction, F(1,38) = 19.43, p < 0.0003, regardless of the time point examined, MA-treated animals had higher levels of corticosterone than the SAL-treated animals.
On P7 and P9, the MA-treated animals had significantly increased corticosterone levels relative to SAL-treated animals, F(1,18) = 17.94, p < 0.0006 and F(1,38) = 33.47, p < 0.0001, respectively. Furthermore on P11, Treatment, F(1,38) = 70.69, p < 0.0001, and Time (p < 0.005) effects were significant. As observed on the other days, MA-treated animals had higher levels of corticosterone than SAL-treated animals. However, regardless of treatment, both the MA- and the SAL-treated animals had higher levels of corticosterone at 30 min compared to animals of the same treatment at 105 min.
For animals treated on P13 and P15 the corticosterone levels of the MA-treated animals were elevated over the SAL-treated animals, regardless of the time point measured, F(1,18) = 38.05, p < 0.0001 and F(1,17) = 19.29, p < 0.0004, respectively. On P17, MA administration increased corticosterone levels relative to animals receiving SAL, F(1,18) = 43.83, p < 0.0001. There was also a significant interaction of Treatment × Time, F(1,18) = 6.9, p < 0.02 since corticosterone levels decreased from the 30 min to the 105 min time point in the SAL-treated animals (p = 0.05) but not in the MA-treated animals (p < 0.06). On the final day of corticosterone examination, P19, the MA-treated animals had higher levels of corticosterone than the SAL-treated animals, F(1,18) = 55.11, p < 0.0001.
In summary, MA treatment produced increases in corticosterone regardless of the age when administration occurred. The corticosterone response to MA did however change over the ages examined, such that high levels of corticosterone were observed following early (i.e. P1, P3, or P5) or late (i.e. P15, P17, or P19) administration with low levels observed following P7–13 administration. Therefore, the response is best described as a U-shaped curve.
Experiment 2
As observed in our previous studies, when MA was administered to animals over multiple days, there was a reduction in body weight relative to animals that received SAL, F(1,133) = 110.67, p < 0.0001. The amount of reduction in body weight was dependent upon the day measured; interaction of Day × Treatment, F(7,133) = 4.54, p < 0.0001. For example, the greatest reduction in body weight occurred when animals were dosed with MA from P11 to P15 (27% reduction vs. SAL), while the least reduction occurred when animals were dosed with MA from P5 to 9 (14% reduction vs. SAL) ().
Table I. Body weights of animals on the last day of dosing receiving 10 mg/kg MA or SAL for five consecutive days with a dosing regimen of four administrations per day for the first four days and then a single administration on the fifth day. Administration began either on postnatal day 1 or every other day thereafter until postnatal day 15. Regardless of when drug administration began, body weights were reduced in the animals that received MA.
An ANOVA of Treatment, Day and Time was used for the corticosterone data. Similar to Experiment 1, all main effects and interactions were significant (p < 0.0001) with Treatment × Day × Time being the highest order significant interaction, F(7,133) = 17.16, p < 0.0001. Therefore, simple effects analyses were run on each Day using Treatment and Time as the main effects ().
Figure 2 Plasma concentrations of corticosterone following a single administration of 10 mg/kg of MA or SAL on the fifth day of administration following a dosing regimen of four administrations per day for the first four days. The animals were decapitated at 30 min (left panel) or 105 min (right panel) after the administration of MA. The levels of corticosterone were dependent on the treatment the animals received, the age that they received treatment, and the time the corticosterone levels were examined. * p < 0.05 for MA vs. SAL.
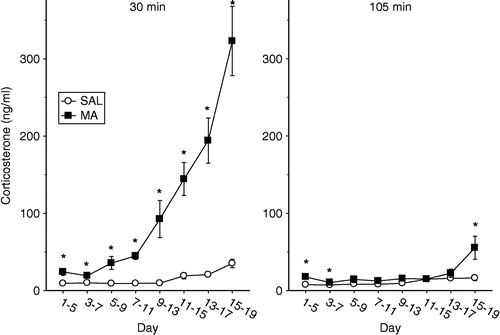
When animals were dosed from P1 to P5 the MA-treated animals had an increase in plasma corticosterone at both the 30 and 105 min time points compared to SAL-treated animals, F(1,18) = 15.93, p < 0.0009. Following MA administration from P3 to P7, the MA-treated animals had slightly elevated levels of corticosterone relative to SAL-treated animals, F(1,14) = 16.00, p < 0.002. For animals that were dosed with MA from P5 to P9, P7–11, or P9–13, only at the 30 min time point did MA administration on the last day of dosing produce a significant increase in corticosterone compared to animals treated with SAL (Treatment × Time, F(1,15) = 17.58, p < 0.0008, F(1,15) = 50.67, p < 0.0001, and F(1,18) = 11.21, p < 0.004, respectively). No differences between MA- and SAL-treated animals were detected at 105 min.
MA-treated animals dosed from P11 to P15, the dosing period that induces cognitive deficits in adulthood (Williams et al. Citation2003b), had elevated levels of corticosterone at 30 min compared to SAL-treated, however by 105 min no differences in corticosterone were observed between the treatments (Treatment × Time, F(1,18) = 35.73, p < 0.0001); supporting our previous findings (Williams et al. Citation2000).
Slightly different patterns were observed in animals that received MA from P13 to P17 or P15–19, although there was an interaction of Treatment and Time for both dosing periods, F(1,17) = 38.06, p < 0.0001 and F(1,18) = 40.65, p < 0.0001, respectively. The MA-treated animals had higher levels of corticosterone at 30 min compared to SAL-treated animals. Animals treated from P13 to 17 with MA had similar levels of corticosterone at 105 min compared to SAL-treated animals, however, animals dosed from P15 to P19 with MA had increased levels of corticosterone at 105 min compared to SAL-treated animals.
In summary for the corticosterone data, MA treatment once again produced increases in plasma corticosterone concentration. Nonetheless, the response was quite different in comparison to an acute MA dose. For example, in this experiment, corticosterone increases were transient following MA administration. At 30 min, corticosterone was increased but by 105 min the response was gone or greatly attenuated at most ages examined. The greatest corticosterone response was following P15–19 MA administration.
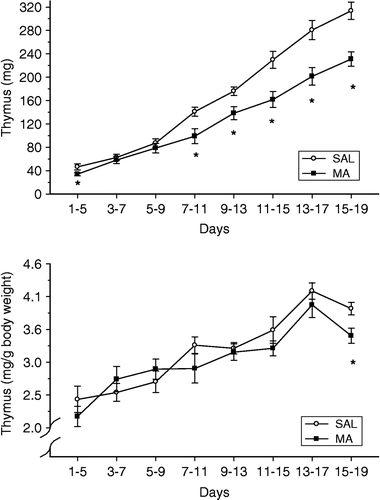
The thymus glands were examined, since in adult animals it is known that chronic exposure to glucocorticoids produces glandular involution. Because there were differences in corticosterone output on each of the days examined, a Student's t-test was performed on each day to examine the animals that received MA compared with those that received SAL (time was not considered a factor for this measure, therefore the means of each litter were used). Thymus weights are presented as either the raw weight or as a function of body weight (, top and bottom panels, respectively). For the raw thymus weights (, top panel), with the exception of animals dosed with MA from P3 to P7 and P5–9, the thymus weights were reduced relative to animals that received SAL. In contrast to the raw weights, examination of (bottom panel) shows that only animals receiving MA from P15 to P19 had smaller thymus weights as percentage of body weight than SAL animals (p < 0.05), whereas no differences were noted after the other dosing periods. There was a general increase in both raw thymus weights and percent thymus weights with development.
Figure 3 Thymus weights of animals following either 10 mg/kg MA or SAL for five consecutive days with a dosing regimen of four administrations per day for the first four days and then a single administration on the fifth day. The weights of pups and thymus from a litter were averaged on each day and a t-test performed to determine significance. The top panel represents the raw thymus weights of the animals, whereas the bottom graph represents the thymus weights as a percent of body weight (). Examination of the raw thymus weights demonstrates that a decrease in weight relative to SAL treatment was observed following most dosing regimens, with the exception of days 3–7 and 5–9. Only animals receiving MA from postnatal day 15–19 had smaller thymus weight expressed as percent body weight, relative to SAL treatment. *p < 0.05.
Comparison of corticosterone response from experiment 1 and experiment 2
In order to determine if there were significant differences in the corticosterone response to MA following an acute or chronic exposure (Regimen), the MA-treated values from both Experiments 1 and 2 were expressed as percent of the SAL-treated values and subjected to ANOVA (Regimen × Day) for the 30 min time point and for the 105 min time point (). At 30 min there was a significant Regimen and Day main effect (p < 0.0001 for both) and a Regimen × Day interaction, F(7,185) = 4.24, p < 0.001. Analysis of the interaction demonstrated that starting on P11, animals that received chronic dosing had increased plasma levels of corticosterone compared to animals that received only an acute dose at the 30 min time point (p < 0.05 on P11, P13, P17 and P19, p < 0.1 on P15). No differences in the corticosterone response were observed on P5 or P9, and a decreased corticosterone response was observed in the chronically-treated animals on P7 compared to the acutely-treated animals (p < 0.05).
Figure 4 Comparison of an acute (Experiment 1) or chronic (Experiment 2) MA-induced increase in plasma corticosterone concentration, expressed as percent of the SAL control values at both 30 (left) and 105 (right) min following MA administration. Chronic administration of MA altered the corticosterone response to acute MA. At 30 min generally the animals that received chronic administration of MA had a larger corticosterone response than animals that received only an acute administration of MA. However, at the 105 min time point, the animals that received only an acute dose of MA had a protracted corticosterone response relative to animals that received prior chronic MA. *p < 0.05, †p < 0.10.
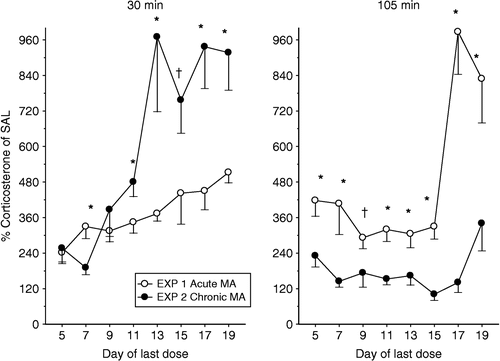
As shown in , right panel, for the 105 min time point, there was a significant Regimen and Day main effect (p < 0.0001 for both) and a Regimen × Day interaction, F(7,185) = 5.85, p < 0.0001. Analysis of the interaction showed that animals that received an acute dose of MA had a greater corticosterone response relative to animals that received the chronic dosing (all p < 0.05 except on P9, p < 0.1).
Discussion
These data demonstrate that (1) MA is a potent stimulator of corticosterone release during development in rats, (2) the adrenal response is dependent upon the age of the rat and is best represented as a U-shaped function when describing the acute response between P1 and P19, (3) a prior history of MA exposure alters the subsequent corticosterone response of the animals, and (4) MA has little influence on the growth of the thymus gland prior to P18 when expressed as a percentage of body weight (since no differences were observed when dosing was from P13 to P17 or prior intervals), however thymus raw weight was decreased on all days except P7 and P9.
We have shown previously that MA administered to neonates from P11 to P20 produces deficits in spatial learning and memory (Vorhees et al. Citation1994, Citation2000; Williams et al. Citation2002, Citation2003c, Citation2004b). The current data therefore suggest that the corticosterone response following MA should be examined in future studies to determine if it is involved in altering neural development and subsequent spatial learning and memory.
The effect of MA on the HPA axis has been studied previously almost exclusively in adults, rather than in developing animals. Adult humans administered MA have increased levels of cortisol (Slade and Checkley Citation1980; Fehm et al. Citation1984), and users of MA that experience MA-induced psychosis can have these psychotic events augmented by stressful situations (Yui et al. Citation1999, Citation2000). Conversely, animal data suggest that MA may potentiate the stress response (Tsuchiya et al. Citation1996a,Citationb) or induce behavioral changes similar to those that occur during or as a result of exposure to stressors. For example, cliff avoidance behavior in adult rats is affected similarly by MA or stressor administration; an effect dependent on an animal's genetic predisposition to stressors (Yoshida et al. Citation1998). MA administration can also act as a zeitgeber to entrain the circadian rhythm of corticosterone release, however, while the rhythm may be shifted, it remains constant in total time (Asano and Moroji Citation1974; Honma et al. Citation1988); weaker effects on bodily rhythms are observed following some stressors as well (Meerlo et al. Citation2002). The present data suggest that neonatal rats also are affected by multiple exposures to MA and have an altered corticosterone response to the drug that is dependent upon the prior exposure to MA as well as age. As can be seen in , 30 min following MA, animals that had previous exposure to MA had augmented corticosterone release when dosing was from P7 to P11, or during the later intervals, compared to animals that only had a single acute dose. Importantly, these augmented increases occurred even during the SHRP. This augmented increase in adrenal output beginning on P11 may indicate a precocious development of the adrenal gland and therefore an early exit from the SHRP. Contrary to the augmented corticosterone increase seen at 30 min for animals that had prior exposure to MA, at 105 min these rats had lower levels of corticosterone, regardless of age, compared to an acute dose. Therefore, this may be indicative of negative feedback processes being more efficient following chronic MA exposure or it may be the result of the higher levels of corticosterone produced following chronic MA being able to bind to more receptors to shut down the system faster. Taken together with the adult data, these data demonstrate that MA alters adrenal and/or HPA axis reactivity throughout an animal's lifetime. In the current study we cannot rule out a potential shift or induction of circadian function to explain the augmented response at 30 min in the chronic MA-exposed animals and this would have to be addressed in future experiments. Because of the altered HPA axis reactivity, it is likely that MA produces alterations to areas of the brain important in the regulation of the body's response to stress.
Neuroanatomical changes related to MA exposure have been noted in both human and animal studies. In human users, magnetic resonance imaging has shown long-term changes in various brain regions, including the hippocampus, following MA exposure either in adulthood or during in utero development (Smith et al. Citation2001; Chang et al. Citation2004; Thompson et al. Citation2004). In animal studies, we have shown that the dentate gyrus and nucleus accumbens have a reduction in dendritic spines in adulthood when the animals were exposed to MA as neonates (Williams et al. Citation2004a). Adult rats administered a neurotoxic regimen of MA have dose-dependent reductions in glucocorticoid receptor binding in the hippocampus and striatum but no changes in mineralocorticoid receptor binding (Lowy and Novotney Citation1994); comparable studies in the neonate are not available. The hippocampus has a rich supply of mineralocorticoid and glucocorticoid receptors that are important in inhibiting the stress response through negative feedback (Mizoguchi et al. Citation2003). Further, the hippocampus is particularly vulnerable to high concentrations of glucocorticoids throughout an animal's lifetime (McEwen et al. Citation1992; Sapolsky Citation1994; CitationMagariños and McEwen Citation1995a,Citationb Sapolsky Citation1996; McEwen Citation1999; Huang and Herbert Citation2005). The adult data suggest that MA might induce a deficit in the negative feedback process of the HPA axis. However, the changes that occur in adult animals are not representative of what occurs in developing animals since we demonstrate that rats with previous exposure to MA also evidently have a rapid negative feedback response relative to animals that received only a single dose with no previous experience with the drug, although as pointed out earlier this may be a function of the amount of corticosterone present in relation to the number of receptors present.
It is well known that neonatal rats respond differently to stressors than adults, going through a period of adrenal quiescence (Jailer Citation1950), or the SHRP (Sapolsky and Meaney Citation1986; Vázquez Citation1998). This period of blunted adrenal response has been hypothesized to be important in protecting neurons during development (Sapolsky and Meaney Citation1986) since high circulating levels of corticosterone can inhibit neurogenesis (Gould et al. Citation1991b). Nonetheless, a complete removal of adrenal steroids is deleterious as well, and therefore, it is important to maintain an optimal level of corticosterone without large increases or decreases during the neonatal period (Woolley et al. Citation1990; Gould et al. Citation1991a; Liu et al. Citation2003). During the SHRP there is rapid growth of neurons within the hippocampus (Bayer et al. Citation1993), an area important for not only negative feedback processes during stress, but also for certain types of learning and memory, such as spatial learning (Morris et al. Citation1982). Furthermore, peak levels of BrdU-labeled cells in the dentate gyrus occur between P9–11 (Liu et al. Citation2003) and P11 is the day at the beginning of the critical period for MA-induced long-term cognitive deficits (Vorhees et al. Citation1994, Citation2000; Williams et al. Citation2002, Citation2003b,Citationc, Citation2004b). Similar to the effects of MA, other factors such as corticotropin-releasing factor (Walker et al. Citation1986) or various stressors (Schoenfeld et al. Citation1980) administered during the neonatal period can be described as producing a U-shaped curve response pattern of corticosterone release. In the current study we also demonstrated this U-shaped corticosterone response to MA treatment.
Early developmental influences are known to alter the response of the HPA axis later in life (Grota Citation1981; Andrews and Matthews Citation2004). These changes are thought to be the result of the early increases in glucocorticoids, as well as perturbations of the serotonergic system, however the basis of these changes is not fully understood. The HPA axis, and especially glucocorticoids, modulates learning and memory as well as a number of physiological and behavioral responses (Grota Citation1981; Lupien and McEwen Citation1997; de Kloet et al. Citation1998; Lupien and Lepage Citation2001). Therefore, any deviation in the development of this system will likely lead to long-term changes in the animal's response to later stressors and how the animal performs when learning. In this regard, we have demonstrated that cliff avoidance in animals exposed neonatally to MA and challenged as adults with 20 mg/kg MA avoided a cliff longer than their control littermates (Williams et al. Citation2003a). MA-treated animals also demonstrated reduced levels of corticosterone immediately following a 15 min forced swim. Interestingly, when corticosterone levels were examined 3 days after a neurotoxic administration of MA (20 mg/kg on a single day), females exposed to MA during development and in adulthood displayed increased basal levels of corticosterone. These data demonstrate that MA administered during the neonatal period produces long-lasting effects on stressor reactivity, both behaviorally and physiologically in the adult animals and physiologically in the neonatal animals as demonstrated in this study.
Other factors that could influence the development of the rats are maternal care, malnutrition, or sickness. In this study, rats that received chronic MA lost weight over time as we have noted before (Vorhees et al. Citation1994, Citation2000; Williams et al. Citation2003b), but they usually do not show signs of lethargy or other sickness-type behaviors. These rats are all nursed as evident by the milk bands observed in their stomachs. Furthermore, it should be noted that long-term body weight changes are not predictive of long-term learning and memory changes since administration of MA from P1 to P10 produced persistent changes in body weights, whereas administration from P11 to P20 produced transient changes (Vorhees et al. Citation1994). Changes in the raw weights of the thymus gland that were noted became significant when body weight changes are greatest (i.e. beginning after P7–P11 dosing). Even though expressing the thymus weight as a percent of body weight does not show any differences until dosing was from P15 to P19, it may be that the smaller thymus is unable to cope with immunological challenges. We have not examined maternal behaviour in detail, but the pups tend to be in a huddle, suggesting maternal care. One factor that was of some concern for the present design was that the litter reductions might influence the adrenal response. Examination of the corticosterone response from pups on P11 that had either 2 animals or 6 animals left in the litter showed no differences at the 105 time point, although slightly lower levels of corticosterone were observed in animals with 6 animals left in a litter, compared to animals with only 2 left in the litter at the 30 min time point. Nonetheless, the percent increase in this study compared to our previous study (c.f. (Williams et al. Citation2000)) where animals were raised in a litter of 10 and killed on P11 are comparable, suggesting that litter size has minimal effects on corticosterone release following MA administration.
The current study is the foundation for examining the impact of MA-induced corticosterone release on hippocampal development early in life, with the possibility of a causal link between MA-induced corticosterone increases and later hippocampus- dependent learning and memory deficits. Furthermore, these data demonstrate that the dynamic corticosterone response to MA is dependent upon age and previous MA exposure.
Acknowledgements
Some of these data were presented in abstract form at the annual meeting for the Society of Neuroscience in 2002 and 2003. Support for this work was from National Institutes of Health grants DA014269 (MTW) and DA006733 (CVV) and training grant ES07051 (LAE, TLS).
References
- Andrews MH, Matthews SG. Programming of the hypothalamo-pituitary-adrenal axis: Serotonergic involvement. Stress 2004; 7: 15–27
- Asano Y, Moroji T. Effects of methamphetamine on daily rhythms of hypothalamic norepinephrine, serotonin and plasma corticosterone levels in the rat. Life Sci 1974; 14: 1463–1472
- Bayer SA, Altman J, Russo RJ, Zhang X. Timetables of neurogenesis in the human brain based on experimentally determined patterns in the rat. Neurotoxicology 1993; 14: 83–144
- Broening HW, Morford LL, Inman-Wood SL, Fukumura M, Vorhees CV. 3,4-methylenedioxymethamphetamine (Ecstasy)-induced learning and memory impairments depend on the age of exposure during early development. J Neurosci 2001; 21: 3228–3235
- Brosnan PG. The hypothalamic pituitary axis in the fetus and newborn. Semin Perinatol 2001; 25: 371–384
- Cappon G, Morford LL, Vorhees CV. Ontogeny of methamphetamine-induced neurotoxicity and associated hyperthermic response. Dev Brain Res 1997; 103: 155–162
- Chang L, Smith LM, LoPresti C, Yonekura ML, Kuo J, Walot I, Ernst T. Smaller subcortical volumes and cognitive deficits in children with prenatal methamphetamine exposure. Psychiatry Res 2004; 132: 95–106
- Chomchai C, Na MN, Watanarungsan P, Yossuck P, Chomchai S. Methamphetamine abuse during pregnancy and its health impact on neonates born at Siriraj hospital, Bangkok, Thailand. Southeast Asian J Trop Med Public Health 2004; 35: 228–231
- Clancy B, Darlington RB, Finlay BL. Translating developmental time across mammalian species. Neurosceince 2001; 105: 7–17
- Crawford CA, Williams MT, Newman ER, McDougall SA, Vorhees CV. Early postnatal methamphetamine exposure causes prolonged changes in dorsal striatal protein kinase A activity, dopamine D2-like binding sites, and dopamine content. Synapse 2003; 48: 131–137
- de Kloet ER, Vreugdenhil E, Oitzl MS, Jodls M. Brain corticosteroid receptor balance in health and disease. Endocr Rev 1998; 19: 269–301
- EMCDDA. 2004. Annual Report 2004: The state of the drugs problem in the European Union and Norway, European Monitoring Centre for Drugs and Drug Addiction, Luxembourg.
- Ernst T, Chang L, Leonido-Yee M, Speck O. Evidence for long-term neurotoxicity associated with methamphetamine abuse: A 1H MRS study. Neurology 2000; 54: 1344–1349
- Fehm HL, Holl R, Steiner K, Klein E, Voigt KH. Evidence for ACTH-unrelated mechanisms in the regulation of cortisol secretion in man. Klin Wochenschr 1984; 62: 19–24
- Gould E, Woolley CS, Cameron HA, Daniels DC, McEwen BS. Adrenal steroids regulate postnatal development of the rat dentate gyrus: II. Effects of glucocorticoids and mineralocorticoids on cell birth. J Comp Neurol 1991a; 313: 486–493
- Gould E, Woolley CS, McEwen BS. Adrenal steroids regulate postnatal development of the rat dentate gyrus: I. Effects of glucocorticoids on cell death. J Comp Neurol 1991b; 313: 479–485
- Gouzoulis-Mayfrank E, Thelen B, Habermeyer E, Kunert HJ, Kovar K-A, Lindenblatt H, Hermle L, Spitzer M, Sass H. Psychopathological, neuroendocrine and autonomic effects of 3,4-methylenedioxyethylamphetamine (MDE), psilocybin and d-methamphetamine in healthy volunteers: Results of an experimental double-blind placebo-controlled study. Psychopharmacology 1999; 142: 41–50
- Grota LJ. Serotonin regulation of corticoid secretion in infant rats. Dev Psychobiol 1981; 14: 221–228
- Herlenius E, Lagercrantz H. Development of neurotransmitter systems during critical periods. Exp Neurol 2004; 190(Suppl 1)S8–21
- Holson RR. Euthanasia by decapitation: Evidence that this technique produces prompt, painless unconsciousness in laboratory rodents. Neurotoxicol Teratol 1992; 14: 253–257
- Holson RR, Pearce B. Principles and pitfalls in the analysis of prenatal treatment effects in multiparous species. Neurotoxicol Teratol 1992; 14: 221–228
- Honma S, Honma K-I, Shirakawa T, Hiroshige T. Rhythms in behaviors, body temperature and plasma corticosterone in SCN leasioned rats given methamphetamine. Physiol Behav 1988; 44: 247–255
- Huang GJ, Herbert J. Serotonin modulates the suppressive effects of corticosterone on proliferating progenitor cells in the dentate gyrus of the hippocampus in the adult rat. Neuropsychopharmacology 2005; 30: 231–241
- Jailer JW. The maturation of the pituitary-adrenal axis in the newborn rat. Endocrinology 1950; 46: 420–425
- Johnston LD, O'Malley PM, Bachman JG, Schulenberg JE. Monitoring the future national survey results on drug abuse, 1975–2003. Vol. II College students and adult ages 19–45 (NIH Publication No. 04-5508). National Institue on Drug Abuse, Bethesda, MD 2004
- Kalechstein AD, Newton TF, Green M. Methamphetamine dependence is associated with neurocognitive impairment in the initial phases of abstinence. J Neuropsychiatry Clin Neurosci 2003; 15: 215–220
- Kent S, Kernahan SD, Levine S. Effects of excitatory amino acids on the hypothalamic-pituitary-adrenal axis of the neonatal rat. Dev Brain Res 1996; 94: 1–13
- Kioukia-Fougia N, Antoniou K, Bekris S, Liapi C, Christofidis I, Padadopoulou-Daifoti Z. The effects of stress exposure on the hypothalamic-pituitary-adrenal axis, thymus, thyroid hormones and glucose levels. Prog Neuropsychopharmacol Biol Psychiatry 2002; 26: 823–830
- Kirk RE. Experimental design: Procedures for the behavioral sciences. Brooks/Cole Publishing Co, Pacific Grove 1995
- Kuczenski R, Segal DS. Neurochemistry of amphetamine. Amphetamine and its analogs: Psychopharmacology, toxicology, and abuse, AK Cho, DS Segal. Academic Press, San Diego, CA 1994; 81–113
- Lindqvist R, Lendahls L, Tollbom O, Aberg H, Hakansson A. Smoking during pregnancy: Comparison of self-reports and cotinine levels in 496 women. Acta Obstet Gynecol Scand 2002; 81: 240–244
- Liu H, Kaur J, Dashtipour K, Kinyamu R, Ribak CE, Friedman LK. Suppression of hippocampal neurogenesis is associated with developmental stage, number of perinatal seizure episodes, and glucocorticosteroid level. Exp Neurol 2003; 184: 196–213
- Lowy MT, Novotney S. Methamphetamine-induced decrease in neural glucocorticoid receptors: Relationship to monoamine levels. Brain Res 1994; 638: 175–181
- Lupien SJ, Lepage M. Stress, memory, and the hippocampus: Can't live with it, can't live without it. Behav Brain Res 2001; 127: 137–158
- Lupien SJ, McEwen BS. The acute effects of corticosteroids on cognition: Integration of animal and human model studies. Brain Res Rev 1997; 24: 1–27
- Magariños AM, McEwen BS. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: Comparison of stressors. Neuroscience 1995a; 69: 83–88
- Magariños AM, McEwen BS. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: Involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 1995b; 69: 89–98
- McCormick CM, Kehoe P, Kovacs S. 1998. Corticosterone release in response to repeated, short episodes of neonatal isolation: Evidence of sensitization, p 175–185.
- McEwen BS. Stress and the aging hippocampus. Front Neuroendocrinol 1999; 20: 49–70
- McEwen BS, Angulo J, Cameron H, Chao HM, Daniels D, Gannon MN, Gould E, Mendelson S, Sakai R, Spencer R, Woolley C. Paradoxical effects of adrenal steroids on the brain: Protection versus degeneration. Biol Psychiatry 1992; 31: 177–199
- Meerlo P, Sgoifo A, Turek FW. The effects of social defeat and other stressors on the expression of circadian rhythms. Stress 2002; 5: 15–22
- Mizoguchi K, Ishige A, Aburada M, Tabira T. Chronic stress attenuates glucocorticoid negative feedback: Involvement of the prefrontal cortex and hippocampus. Neuroscience 2003; 119: 887–897
- Morris RGM, Garrud P, Rawlins JNP, O'Keefe J. Place navigation impaired in rats with hippocampal lesions. Nature 1982; 297: 681–683
- O'Callaghan JP, Miller DB. Neurotoxic effects of substituted amphetamines in mice and rats: Challenges to the current dogma. Handbook of neurotoxicity, EJ Massaro. Humana Press, Totowa, NJ 2002; 269–301
- Richardson GA, Hamel SC, Goldschmidt L, Day NL. Growth of infants prenatally exposed to cocaine/crack: Comparison of prenatal care and no prenatal care sample. Pediatrics 1999; 104: e18
- Sapolsky RM. The physiological relevance of glucocorticoid endangerment of the hippocampus. Ann N Y Acad Sci 1994; 746: 294–307
- Sapolsky RM. Stress, glucocorticoids, and damage to the nervous system: The current state of confusion. Stress 1996; 1: 1–19
- Sapolsky RM, Meaney MJ. Maturation of the adrenocortical stress response: Neuroendocrine control mechanisms and the stress hyporesponsive period. Brain Res Rev 1986; 11: 65–76
- Schoenfeld NM, Leathem JH, Rabii J. Maturation of adrenal stress responsiveness in the rat. Neuroendocrinology 1980; 31: 101–105
- Seiden LS, Sabol KE, Ricaurte GA. Amphetamine: Effects on catecholamine systems and behavior. Annu Rev Pharmacol Toxicol 1993; 33: 639–677
- Slade AP, Checkley SA. A neuroendocrine study of the mechanism of action of ECT. Br J Psychiatry 1980; 137: 217–221
- Smith LM, Chang L, Yonekura ML, Grob C, Osborn D, Ernst T. Brain proton magnetic resonance spectroscopy in children exposed to methamphetamine in utero. Neurology 2001; 57: 255–260
- Smith L, Yonekura ML, Wallace T, Berman N, Kuo J, Berkowitz C. Effects of prenatal methamphetamine exposure on fetal growth and drug withdrawal symptoms in infants born at term. J Dev Behav Pediatr 2003; 24: 17–23
- Thompson PM, Hayashi KM, Simon SL, Geaga JA, Hong MS, Sui Y, Lee JY, Toga AW, Ling W, London ED. Structural abnormalities in the brains of human subjects who use methamphetamine. J Neurosci 2004; 24: 6028–6036
- Tsuchiya K, Inoue T, Izumi T, Hashimoto S, Koyama T. Effects of footshock stress on regional brain monoamine metabolism and the acquisition of conditioned freezing in rats previously exposed to repeated methamphetamine. Prog Neuropsychopharmacol Biol Psychiatry 1996a; 20: 1239–1250
- Tsuchiya K, Inoue T, Koyama T. Effect of repeated methamphetamine pretreatment on freezing behavior induced by conditioned fear stress. Pharmacol Biochem Behav 1996b; 54: 687–691
- Vázquez DM. Stress and the developing limbic-hypothalamic-pituitary-adrenal axis. Psychoneuroendocrinology 1998; 23: 663–700
- Vorhees CV, Ahrens KG, Acuff-Smith KD, Schilling MA, Fisher JE. Methamphetamine exposure during early postnatal development in rats: I. Acoustic startle augmentaion and spatial learning deficits. Psychopharmacology 1994; 114: 392–401
- Vorhees CV, Inman-Wood SL, Morford LL, Broening HW, Fukumura M, Moran MS. Adult learning deficits after neonatal exposure to d-methamphetamine: Selective effects on spatial navigation and memory. J Neurosci 2000; 20: 4732–4739
- Walker C-D, Dallman MF. Neonatal facilitation of stress-induced adrenocorticotropin secretion by prior stress: Evidence for increased central drive to the pituitary. Endocrinology 1993; 132: 1101–1107
- Walker C-D, Perrin M, Vale W, Rivier C. Ontogeny of the stress response in the rat: Role of the pituitary and the hypothalamus. Endocrinology 1986; 118: 1445–1451
- Walker C-D, Scribner KA, Cascio CS, Dallman MF. The pituitary-adrenocortical system of neonatal rats is responsive to stress throughout development in a time-dependent and stressor-specific fashion. Endocrinology 1991; 128(3)1385–1395
- Williams MT, Inman-Wood SL, Morford LL, McCrea AE, Ruttle AM, Moran MS, Rock SL, Vorhees CV. Preweaning treatment with methamphetamine induces increases in both corticosterone and ACTH in rats. Neurotoxicol Teratol 2000; 22: 751–759
- Williams MT, Vorhees CV, Boon F, Saber AJ, Cain DP. Methamphetamine exposure from postnatal day 11 to 20 causes impairments in both behavioral strategies and spatial learning in adult rats. Brain Res 2002; 958: 312–321
- Williams MT, Blankenmeyer TL, Schaefer TL, Brown CA, Gudelsky GA, Vorhees CV. Long-term effects of neonatal methamphetamine exposure in rats on spatial learning in the Barnes maze and on cliff avoidance, corticosterone release, and neurotoxicity in adulthood. Dev Brain Res 2003a; 147: 163–175
- Williams MT, Moran MS, Vorhees CV. Refining the critical period for methamphetamine-induced spatial deficits in the Morris water maze. Psychopharmacology 2003b; 168: 329–338
- Williams MT, Morford LL, Wood SL, Wallace TL, Fukumura M, Broening HW, Vorhees CV. Developmental d-methamphetamine treatment selectively induces spatial navigation impairments in reference memory in the Morris water maze while sparing working memory. Synapse 2003c; 48: 138–148
- Williams MT, Brown RW, Vorhees CV. Neonatal methamphetamine administration induces region-specific long-term neuronal morphological changes in the rat hippocampus, nucleus accumbens, and parietal cortex. Eur J Neurosci 2004a
- Williams MT, Moran MS, Vorhees CV. Behavioral and growth effects induced by low dose methamphetamine administration during the neonatal period in rats. Int J Dev Neurosci 2004b; 22: 273–283
- Williams MT, Schaefer TL, Ehrman LA, Able JA, Gudelsky GA, Sah R, Vorhees CV. 3,4-Methylenedioxymethamphetamine administration on postnatal day 11 in rats increases pituitary-adrenal output and reduces striatal and hippocampal serotonin without altering SERT activity. Brain Res 2005; 1039: 97–107
- Winer BS. Statistical principles in experimental design. McGraw-Hill, New York 1971
- Woolley CS, Gould E, McEwen BS. Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Res 1990; 531: 225–231
- Yoshida S, Numachi Y, Matsuoka H, Sato M. Impairment of cliff avoidance reaction induced by subchronic methamphetamine administration and restraint stress: Comparison between two inbred strains of rats. Prog Neuropsychopharmacol Biol Psychiatry 1998; 22: 1023–1032
- Yui K, Goto K, Ikemoto S, Ishiguro T, Angrist B, Duncan GE, Sheitman BB, Lieberman JA, Bracha SH, Ali SF. Neurobiological basis of relapse prediction in stimulant-induced psychosis and schizophrenia: The role of sensitization. Mol Psychiatr 1999; 4: 512–523
- Yui K, Goto K, Ikemoto S, Ishiguro T. Stress induced spontaneous recurrence of methamphetamine psychosis: The relation between stressful experiences and sensitivity to stress. Drug Alcohol Depend 2000; 58: 67–75