Abstract
People who are exposed to horrific, life-threatening experiences are at risk for developing post-traumatic stress disorder (PTSD). Some of the symptoms of PTSD include persistent anxiety, exaggerated startle, cognitive impairments and increased sensitivity to yohimbine, an α2-adrenergic receptor antagonist. We have taken into account the conditions known to induce PTSD, as well as factors responsible for long-term maintenance of the disorder, to develop an animal model of PTSD. Adult male Sprague–Dawley rats were administered a total of 31 days of psychosocial stress, composed of acute and chronic components. The acute component was a 1-h stress session (immobilization during cat exposure), which occurred on Days 1 and 11. The chronic component was that on all 31 days the rats were given unstable housing conditions. We found that psychosocially stressed rats had reduced growth rate, reduced thymus weight, increased adrenal gland weight, increased anxiety, an exaggerated startle response, cognitive impairments, greater cardiovascular and corticosterone reactivity to an acute stressor and heightened responsivity to yohimbine. This work demonstrates the effectiveness of acute inescapable episodes of predator exposure administered in conjunction with daily social instability as an animal model of PTSD.
Introduction
Individuals exposed to horrific, life-threatening experiences are at substantial risk for developing post-traumatic stress disorder (PTSD). People who develop PTSD respond to an acute traumatic experience with intense feelings of fear, helplessness or horror (American Psychiatric Association Citation1994) and subsequently endure chronic psychological distress by repeatedly reliving their trauma through intrusive, flashback memories (Ehlers et al. Citation2004, Brewin Citation1998, Citation1999, Citation2007). These characteristics of the disorder foster the development of several other symptoms, including persistent anxiety, exaggerated startle, cognitive impairments and diminished extinction of conditioned fear (Brewin et al. Citation2000, Bremner Citation2002).
PTSD is characterized by a complex aberrant biological profile involving several physiological systems, including the sympathetic nervous system and the hypothalamus-pituitary-adrenal (HPA) axis (de Kloet et al. Citation2006; Nemeroff et al. Citation2006; Stam Citation2007a). For instance, relative to control subjects, PTSD patients display significantly greater cortisol levels in response to, and in anticipation of, acute laboratory stressors (Bremner et al. Citation2003; Elzinga et al. Citation2003). Although findings have been mixed, extensive work has also reported abnormally low baseline levels of cortisol in people with PTSD (Yehuda Citation2005).
PTSD patients also demonstrate greater baseline and stress-induced elevations of sympathetic activity (Buckley and Kaloupek Citation2001; Pole Citation2007). In response to traumatic reminders and standard laboratory stressors, people with PTSD display significantly greater increases in heart rate (HR), blood pressure (BP), skin conductance, epinephrine and norepinephrine than do control subjects (Blanchard et al. Citation1982, Citation1991, Mutalipassi Citation1992). In addition, PTSD patients exhibit elevated baseline HR, diastolic BP and systolic BP (Buckley and Kaloupek Citation2001; Pole Citation2007), findings that resonate with the recent work reporting an association between PTSD and increased risk for cardiovascular disease (Chang Citation1999).
Another indication of accentuated sympathetic activity in PTSD patients is the hyperresponsivity (i.e., greater increases in HR and BP, greater expression of anxiety-like behavior) they exhibit in response to the administration of yohimbine, an α2-adrenergic receptor antagonist which blocks noradrenergic autoreceptors and leads to increased central norepinephrine activity (Rasmusson et al. Citation2000, Citation1999a-c). These findings, along with those of greater baseline norepinephrine levels in PTSD patients (Geracioti et al. Citation2001), have implicated a major role of the noradrenergic system in the hyperarousal component of PTSD (Strawn and Geracioti Citation2007).
Preclinical researchers have used several types of stressors to model aspects of PTSD in rodents (Stam Citation2007b). Such stressors have included electric shock (Garrick et al. Citation2001, Citation2006, Wotjak Citation2007ab; Wakizono et al. Citation2007), underwater trauma (Cohen et al. Citation2004; Richter-Levin Citation1998), stress–restress and single prolonged stress paradigms (Harvey et al. Citation2003, Liberzon Citation2004) and exposure to predators (Adamec Citation1997, Citation2006, Citation2007, Shallow Citation1993) or predator-related cues (Cohen et al. Citation2000b, Citation2006, Citation2007, Zohar Citation2004). The stressors employed in these studies typically produced increased behavioral signs of anxiety, and in some cases, exaggerated startle, cognitive impairments, enhanced fear conditioning and reduced social interaction. Although these studies have reported physiological and behavioral changes resembling those observed in people with PTSD, most have utilized only a small set of assessments, such as stress-induced changes in anxiety, without assessing other measures common in people with PTSD, such as an impairment in cognitive measures. Moreover, many of these studies have evaluated stress-induced changes in responses for a relatively short period of time. Thus, while these studies have provided an insight into how stress or fear conditioning changes the aspects of behavior and physiology, the field will benefit from an animal model of PTSD that takes into account how traumatic stress produces long-lasting PTSD-like changes in rats given multiple behavioral and physiological diagnostic tests.
In the present set of experiments, we incorporated three factors in the development of our animal model of PTSD, each of which is known to influence the induction and maintenance of PTSD in people. First, we used cat exposure, a well-described, ethologically relevant stressor, which produces intense fear responses in rats (Blanchard et al. 1990, 2003, 2005; Hubbard et al. 2004). In addition, the element of control over a stressor exerts a great influence over the expression of the stress response (Kim and Diamond Citation2002). As a loss of control exacerbates behavioral and physiological responses to stress conditions (Amat et al. Citation2005, Citation2007, Watkins Citation2005), we immobilized the rats during predator exposure. The immobilization component of our animal model provides a rodent analogue of the sense of helplessness and a loss of control which feature prominently in the diagnostic and statistical manual of mental disorders (DSM-IV) criteria for PTSD (American Psychiatric Association Citation1994).
Second, a core symptom of PTSD is the repeated “re-experiencing” of the traumatic event that people with PTSD suffer from in response to activation of intense and intrusive memories of their trauma (Ehlers et al. Citation2004, Brewin Citation1998, Citation1999, Citation2007). We therefore included a re-experiencing component in our animal model of PTSD. Specifically, rats were given a second cat exposure session, 10 days after the first, to serve as a powerful reminder of their traumatic experience.
Third, it is well known that only a minority of traumatized individuals develop persistent PTSD symptoms. One factor that increases the likelihood that emotional trauma will develop into chronic PTSD is a lack of social support and stability (Andrews et al. Citation2003, Filipas Citation2001). Therefore, to increase the likelihood of producing long-lasting sequelae in the subjects, we included daily social instability in the model by randomizing the housing conditions of the rats. Three weeks after the second predator exposure, we gave the rats an extensive battery of diagnostic tests that had parallels to routine testing of patients with PTSD.
The overall goal of this work was to take into account that factors are known to be involved in the etiology and persistence of PTSD symptoms in people to study the persistent effects of traumatic stress on rat behavior and physiology. To accomplish this goal, we incorporated the use of powerful psychological stressors, including a life-threatening stressor, coupled with a re-experiencing of the trauma and chronic social instability, in the development of a novel animal model of PTSD.
Materials and methods
Animals
Adult male Sprague–Dawley rats of 225–250 g (Charles River Laboratories, Wilmington, MA) were housed on a 12:12 h light–dark schedule (lights on at 0700 h) in Plexiglas cages (two per cage) with food (Harlan Teklad Global 18% Protein Rodent Diet; Harlan Laboratories; Indianapolis, IN) and water provided ad libitum. Rats were given one week to acclimate to the housing room environment before any experimental manipulations took place. All experiments were carried out in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals, and the Institutional Animal Care and Use Committee at the University of South Florida approved all procedures.
In each study, rats were handled for three consecutive days (2–3 min each) prior to behavioral/physiological testing. Body weights were recorded on the day of the first stress session and on the first day of behavioral/physiological testing. Average growth rates (g/day) were calculated (total weight gained divided by the length of the experiment (i.e., 31 days)] for statistical analysis. The sequence of events in each experiment is described below and is illustrated in .
Figure 1 Timeline and procedures for Experiments 1–4. In Experiment 1, all rats were acclimated to the HR/BP apparatus prior to the experimental manipulations. Subsequently, rats were exposed to two acute stress sessions, comprised of immobilization during cat exposure (psychosocial stress; cat icon; n = 8) or home cage exposure (no psychosocial stress; rat cage icon; n = 10), that were separated by 10 days. We obtained blood samples and cardiovascular measurements from all rats following each of the two acute stress sessions. Psychosocially stressed rats were also given unstable housing conditions from Day 1 until the beginning of behavioral testing, which occurred three weeks after the second cat exposure. In Experiment 2, we repeated the explicit stress procedures from Experiment 1 (n = 10 per group) without performing blood sampling and cardiovascular measurements after each of the acute stress sessions. On Day 32, we assessed the effects of this paradigm on growth rate, as well as adrenal gland and thymus weight. In Experiment 3, groups of rats were given: (1) two inescapable cat exposures, in conjunction with daily social stress (psychosocial stress; n = 8), (2) two inescapable cat exposures, alone (cat only; n = 10), (3) two exposures to home cage, in conjunction with daily social stress (social stress only; n = 10) or (4) two exposures to home cage, alone (no psychosocial stress; n = 8) to determine whether both stress manipulations (i.e., inescapable cat exposure and daily social stress) were necessary to produce the observed effects on rat physiology and behavior. In Experiment 4, two groups of rats (psychosocial stress, n = 19 and no psychosocial stress, n = 20) were exposed to the same manipulations employed in Experiment 2. On Days 32 and 33, a subsection of each group of rats was injected with yohimbine (YOH: 10 and 12, respectively) or vehicle (VEH: 9 and 8, respectively) prior to behavioral testing.
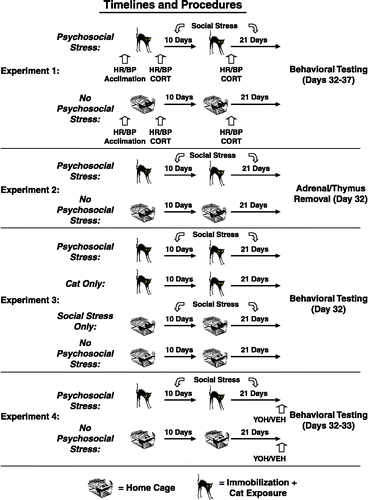
Experiment 1: Establishment of the animal model
Stress manipulations and physiological assessments
We took HR and BP measurements (described in detail below) at three different time points in Experiment 1. This procedure involved placing rats in a Plexiglas tube and a tail cuff sensor around their tails (IITC Life Science; Woodland Hills, CA). The tube was designed to inhibit gross movements during testing to minimize movement artifacts. Rats were brought to the laboratory one day prior to the first stress session and exposed to the HR and BP apparatus for 10 min each to habituate them to this procedure.
One day later, rats were randomly assigned to “psychosocial stress” (N = 8) or “no psychosocial stress” (N = 10) groups. Rats in the psychosocial stress group were exposed to two acute stress sessions; the first stress session was conducted during the light cycle (between 0800 and 1500 h), and the second stress session was conducted 10 days later during the dark cycle (between 1900 and 0200 h). The stress sessions were separated by 10 days based on the finding that pyramidal cells in the basolateral amygdala exhibited greater dendritic spine density 10 days after a single session of immobilization stress (Mitra et al. Citation2005). This form of dendritic “remodeling” in the amygdala is associated with increased fear- and anxiety-related behaviors (Fuchs et al. Citation2006; Mitra et al. Citation2005; Vyas et al. Citation2006). Thus, the strategy behind employing a second stress session 10 days after the first was: (1) to expose the rats to a traumatic re-experiencing of the original event, in a manner analogous to the intrusive reliving of traumatic memories by people with PTSD and (2) to reinforce the presumed changes in the amygdala that would have been initiated by the first stress session.
During each acute stress session, rats were immobilized in plastic DecapiCones (Braintree Scientific; Braintree, MA) and then individually placed in triangular-shaped wedges within a circular Plexiglas “pie” enclosure of 20 × 20 × 8 cm (l × w × h; Braintree Scientific; Braintree, MA). Fifteen minutes later, the rat pie enclosure (which could hold a maximum of 12 immobilized rats) was taken to the cat housing room (which was located in a different area of the animal facility than the laboratory) and placed in a metal cage (61 × 53 × 51 cm). An adult, gonadally intact, female cat (body weight = 3.7 kg) was placed inside the metal cage on top of the rat pie enclosure in full view of the rats. The Plexiglas pie enclosure prevented any contact between the cat and rats, but exposed the rats to all non-tactile sensory stimuli associated with the cat. Canned cat food was smeared on the top of the Plexiglas enclosure to direct cat activity toward the rats. The immobilized rats, inside the pie enclosure, were exposed to the cat for 45 min.
Rats in the no psychosocial stress group were brought to the laboratory (which never contained a cat), where they remained in their home cages for the 1-h stress period. They were returned to the laboratory for one hour once again 10 days later. The rats in the no psychosocial stress group were housed with the same cage cohort for the duration of the experiment.
Immediately after the inescapable cat exposure period (or the 1-h laboratory exposure period in control group), blood samples were obtained from the rats to determine their serum corticosterone levels. Rats were placed in a wire mesh restrainer and a 1-mm tail snip was made with a sterile razor blade. A 0.5 ml sample of blood was then collected in a microcentrifuge tube within 2–3 min. Once clotted, the blood was centrifuged, and the serum was extracted and stored at − 80°C until assayed for corticosterone with an Enzyme ImmunoAssay kit from Assay Design, Inc. (cat#901-097, Ann Arbor, Michigan). All samples were diluted 1:50 and assayed per kit instructions.
After obtaining the blood sample, the rats were placed in a Plexiglas tube within a warming test chamber (∼32°C) for 5 min. This manipulation increased blood flow to the tail, which enabled HR and BP to be measured using tail cuffs fitted with photoelectric sensors (IITC Life Science; Woodland Hills, CA). We obtained HR/BP measurements from all rats following each of the two stress sessions.
Beginning with the first stress session, the psychosocial stress group was exposed to unstable housing for the next 31 days. All rats were housed two per cage, but every day, the cohort pair combinations for the rats in the psychosocial stress group were changed. We utilized this manipulation because previous work from our group has demonstrated that it has significant deleterious effects on rat behavior (Gerges et al. Citation2004; Park et al. Citation2001).
Once behavioral testing began on Day 32, all rats remained housed with the same cohort for the remainder of the experiment. The daily unstable housing manipulation was terminated on Day 32 so that all rats were given identical housing conditions during behavioral testing. Therefore, group differences in performance during the behavioral testing days could be attributed to the psychosocial stress manipulations that occurred on Days 1 to 31.
Behavioral and physiological tests
Three weeks after the second stress session (beginning on Day 32), rats were given tests to measure anxiety, startle, learning and memory, cardiovascular activity and corticosterone activity. The three-week delay from the second inescapable cat exposure to behavioral testing was based on comparable time periods employed in other studies on the effects of stress on brain and behavior (Adamec and Shallow Citation1993-c). On each day of testing, rats were brought to the laboratory and left undisturbed for 30 min before testing began. This was done to acclimate rats to the environment and minimize any acute transport stress-induced effects on physiology and behavior. All testing took place between 0800 and 1500 h.
Elevated plus maze
The elevated plus maze (EPM) is a routine test of anxiety in rodents (Korte and De Boer Citation2003) and consists of two open arms (11 × 51 cm) and two closed arms (11 × 51 cm) that intersect each other to form the shape of a plus sign. On Day 32, rats were placed on the EPM for 5 min, and their behavior was scored by 48 infrared photobeams (located along the perimeter of the open and closed arms), which were connected to a computer program (Motor Monitor, Hamilton-Kinder, San Diego, CA). The primary dependent measures of interest were the amount of time rats spent in the open arms and the number of ambulations made by each rat. An arm entry was scored by the computer program only when the rat's entire body had moved from one arm into a new arm (e.g., the entire body of the rat moved from the closed arms into an open arm). Thus, the computer program would begin tallying open arm time only after a rat had completely entered an open arm. An ambulation was scored by the computer program each time a rat crossed a photobeam sensor. Thus, the ambulations score consisted of the total number of beam breaks made by each rat during the 5-min trial and served as a measure of motor activity.
Startle response
One hour after the EPM assessment, acoustic startle testing was administered. Rats were placed inside a small Plexiglas box (19 × 10 × 10 cm), which was inside a larger startle monitor cabinet (Hamilton-Kinder; San Diego, CA; 36 × 28 × 50 cm). The small Plexiglas box within this cabinet contained a sensory transducer on which rats were placed at the beginning of the trial. The sensory transducer was connected to a computer (Startle Monitor computer program; Hamilton-Kinder; San Diego, CA), which recorded rats' startle responses by measuring the maximum amount of force (in N) that rats exerted on the sensory transducer for a period of 250 ms after the presentation of each auditory stimulus. To control the differences in body weight, the sensitivity of the sensory transducer was adjusted prior to each trial via a vernier adjustment with a sensitivity range of 0–7 arbitrary units. The startle trial began with a 5-min acclimation period, followed by the presentation of 24 bursts of white noise (50 ms each), 8 from each of 3 auditory intensities (90, 100 and 110 dB). The noise bursts were presented in sequential order (i.e., 8 bursts at 90 dB, followed by 8 bursts at 100 dB, followed by 8 bursts at 110 dB), and the time between each noise burst varied pseudorandomly between 25 and 55 s. Upon the commencement of the first noise burst, the startle apparatus provided uninterrupted background white noise (57 dB).
Radial-arm water maze
On Day 33, rats underwent radial-arm water maze (RAWM) training to assess their learning and memory following methods described previously (Campbell et al. 2007; Diamond et al. 1999, 2006; Park et al. 2006; Sandi et al. 2005; Woodson et al. 2003; Zoladz et al. 2006). Briefly, the RAWM consists of a black, galvanized round tank (168 cm diameter, 56 cm height, 43 cm depth) filled with water (21–22°C). Six V-shaped stainless steel inserts (54 cm height, 56 cm length) were placed in the tank to produce six swim arms radiating from an open central area. A black, plastic platform (12 cm diameter) was located 1 cm below the surface of the water at the end of one arm (referred to as the “goal arm”). At the start of each trial, rats were released into one arm (referred to as the “start arm”) facing the center of the maze. If a rat did not locate the hidden platform within 60 s, it was guided to the platform by the experimenter. Once a rat found or was guided to the platform, it was left undisturbed for 15 s. Rats received 12 acquisition trials to learn the location of the hidden platform. Then, they were returned to their home cages for one hour, followed by a single memory test trial. To assess long-term memory, rats were brought to the laboratory 24 h later (Day 34) and given a single memory test trial. Arm entry errors (i.e., entries into arms that did not contain the hidden platform) served as an indicator of rats' memory for the hidden platform.
Novel object recognition
The novel object recognition (NOR) task was a modified version of that which was employed by Baker and Kim (Citation2002). On Day 35, rats were placed in an open field of 40 × 47 × 70 cm (Hamilton-Kinder, San Diego, CA) for 5 min to acclimate to the environment. Their behavior was scored by infrared photobeams connected to a computer program (Motor Monitor). Twenty-four hours later (Day 36), the rats were placed in the same open field with two identical (plastic/metal) objects for 5 min. The objects were in opposite corners of the open field and secured to the flooring to prevent rats from displacing them. The objects were counterbalanced across rats, as were the corners in which the objects were located. Three hours later, rats were returned to the open field, but this time it contained a replica of the object that had been there before and a novel object. To quantify the time spent with the objects during training and testing, we specified a 16-cm2 zone around each object to examine motor activity. During testing, greater time spent by the rats in proximity to the novel versus familiar object was an indication of intact memory for the familiar object.
Blood sampling and cardiovascular measurements
On Day 37, we obtained three blood samples and HR/BP measurements from all rats, following procedures described above. A single blood sample was collected after which the rats were restrained for 20 min. Then, another blood sample was collected to examine their corticosterone response to restraint stress. The rats were then removed from the restrainer and HR/BP measurements were obtained. The rats were then returned to their home cages for one hour, after which a sample of trunk blood was collected via rapid conscious decapitation to determine the recovery of corticosterone levels. The serum was processed as described above. Brains were rapidly removed and stored at − 80°C for later analysis (not reported here).
Experiment 2: Replication of Experiment 1 without physiological assessments at the time of acute predator stress effects
The psychosocial stress and no psychosocial stress groups in Experiment 1 had similar growth rates (see ). This null effect of psychosocial stress on body weight gain could have occurred because the no psychosocial stress group was adversely affected by the acute restraint for HR/BP measurements and by the tail bleed procedure on Days 1 and 11. A single exposure to restraint stress can induce a long-lasting reduction in food intake and, consequentially, body weight gain (Valles et al. Citation2000). To test our hypothesis directly, we repeated the explicit stress procedures from Experiment 1 without performing the cardiovascular measurements and blood sampling on Days 1 and 11. In Experiment 2, we examined the effects of inescapable cat exposure and daily social instability on growth rate, and on adrenal gland and thymus weight (N = 10 per group), since chronically stressed rodents exhibit hypertrophy of the adrenal glands and atrophy of the thymus (Karst and Joels Citation2003). On Day 32, at the conclusion of the chronic stress period, the rats were weighed, rapidly decapitated as in Experiment 1, adrenal glands and thymus were removed and weighed, and the brains collected as above. Organ weights were expressed as actual values and as mg/100 g body weight (Karst and Joels Citation2003).
Table I. Mean growth rate (g/day, mean±SEM) of rats during the 31-day period from stress session 1 to the first day of behavioral/physiological testing.
Experiment 3: Is the combination of predator exposure and social instability necessary for PTSD-like effects?
Experiment 3 assessed whether the combination of two inescapable cat exposures and daily social instability was necessary to produce the persistent effects on physiology and behavior observed in Experiment 1. Rats were randomly assigned to one of four experimental groups: two inescapable cat exposures and daily social stress (“cat + social stress,” N = 8), two inescapable cat exposures alone (“cat only,” N = 10), daily social stress alone (“social stress only,” N = 10) or no manipulation (“no stress,” N = 8). On Day 32, rats were given EPM and startle testing, following the procedures from Experiment 1.
Experiment 4: Effects of psychosocial stress on behavioral and corticosterone responses to yohimbine
Experiment 4 examined the effects of two inescapable cat exposures, in conjunction with daily social instability (as in Experiments 1–3), on responsivity to yohimbine. On Day 32, psychosocial stress and no psychosocial stress rats were injected with yohimbine hydrochloride (1 mg/kg, i.p.) or vehicle (distilled H2O, 1 ml/kg) 30 min prior to testing. Pilot work indicated that 1 mg/kg was a threshold dose of yohimbine for producing anxiogenic behavior. The group sample sizes were: psychosocial stress-vehicle (S-VEH; N = 9), psychosocial stress-yohimbine (S-YOH; N = 10), no psychosocial stress-vehicle (NS-VEH; N = 8), no psychosocial stress-yohimbine (NS-YOH; N = 12). Thirty minutes after the injections, rat behavior in the open field was monitored for 10 min. One hour after the open field assessment, rats were tested on the EPM for 5 min, followed one hour later by acoustic startle testing. On the next day (Day 33), the rats were brought to the laboratory and left undisturbed for 30 min and then were injected with the same agent that they had received one day earlier. Thirty minutes later, three blood samples were obtained as in Experiment 1.
Statistical analyses
Alpha was set at 0.05 for all analyses, and Holm–Sidak post hoc comparisons (Sigmastat, SPSS) were employed when appropriate. In Experiments 1 and 2, independent samples t-tests and mixed-model ANOVAs (for repeated measures variables) were used to compare the psychosocial stress and no psychosocial stress groups. In Experiment 3, two-way and mixed-model ANOVAs (for repeated measures variables) were used to compare the groups on all measures. In Experiment 4, growth rate data were analyzed with an independent samples t-test, while all other behavioral and physiological data were analyzed with two-way and mixed-model ANOVAs (for repeated measures variables). Outlier data points greater than three standard deviations from exclusive group means were eliminated from analyses (less than 1% of the data were outliers). In all figures, values are expressed as means ± SEM.
Results
Experiment 1: Establishment of the animal model
Physiological measurements after each acute stress session
Heart rate and blood pressure
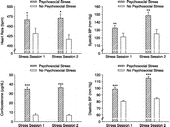
We employed mixed-model ANOVAs to analyze cardiovascular and corticosterone responses after each stress session (). For HR, there was a significant main effect of stress, F(1,15) = 7.89, p < 0.05, indicating that the psychosocial stress group had significantly greater HR than the no psychosocial stress group after each stress session. There was no significant main effect of session, F(1,15) = 0.11, and the Stress × Session interaction was not significant, F(1,15) = 0.85 (all p>0.05). For systolic BP, there was a significant main effect of stress, F(1,13) = 15.80, p < 0.01, and a significant Stress × Session interaction, F(1,13) = 4.94, p < 0.05. The psychosocial stress group demonstrated significantly greater systolic BP than the no psychosocial stress group after each stress session, and only the psychosocial stress group displayed significantly greater systolic BP after the second stress session, relative to the first stress session (all p < 0.05). There was no significant main effect of session, F(1,13) = 1.48, p>0.05. For diastolic BP, there were significant main effects of stress, F(1,14) = 118.03, p < 0.0001, and session, F(1,14) = 30.81, p < 0.001, and a significant Stress × Session interaction, F(1,14) = 9.40, p < 0.01. The psychosocial stress group demonstrated significantly greater diastolic BP than the no psychosocial stress group after each stress session, and only the psychosocial stress group displayed significantly greater diastolic BP after the second stress session, relative to the first stress session (all p < 0.05).
Figure 2 Cardiovascular activity and serum corticosterone following each stress session in Experiment 1. The psychosocial stress group displayed greater HR (top left), systolic BP (top right), diastolic BP (bottom right) and serum corticosterone concentrations (bottom left) than the no psychosocial stress group following each stress session. Additionally, the psychosocial stress group demonstrated significantly greater systolic and diastolic BP after the second stress session, relative to the first stress session. *p < 0.05; **p < 0.01; ***p < 0.0001 versus the no psychosocial stress group. Data are group means ± SEM.
Serum corticosterone
For serum corticosterone concentration, there was a significant main effect of stress, F(1,18) = 189.90, p < 0.0001, indicating that the psychosocial stress group had significantly greater corticosterone levels than the no psychosocial stress group after each stress session. There was no significant main effect of session, F(1,18) = 0.11, and the Stress × Session interaction was not significant, F(1,18) = 2.54 (all p>0.05).
Behavioral and physiological testing (Days 32–37)
Body weight
There was no significant difference between the psychosocial stress and no psychosocial stress groups in terms of growth rate, t(14) = 0.69, p>0.05 ().
Elevated plus maze
On EPM, the psychosocial stress group spent significantly less time in the open arms than the no psychosocial stress group, t(16) = 5.01, p < 0.001 (). There was no significant difference between the two groups in terms of ambulations on the EPM, t(16) = 1.92, p>0.05.
Figure 3 Elevated plus maze and startle response data from Experiment 1. The psychosocial stress group spent significantly less time in the open arms of the elevated plus maze than the no psychosocial stress group (top left). This was despite finding that the two groups demonstrated comparable motor activity on the elevated plus maze (top right). The psychosocial stress group also demonstrated greater startle responses than the no psychosocial stress group to the 90 and 110 dB, but not 100 dB, auditory stimuli (bottom). *p < 0.05; **p < 0.001 versus the no psychosocial stress group. Data are group means ± SEM.

Startle response
We employed mixed-model ANOVAs to analyze startle responses to the 90, 100 and 110 dB auditory stimuli (). There was a significant main effect of psychosocial stress, F(1,14) = 13.03, p < 0.01, indicating that, overall, the psychosocial stress group demonstrated significantly greater startle responses than the no psychosocial stress group. There was also a significant main effect of auditory stimulus intensity, F(2,28) = 74.57, p < 0.0001, indicating that rats exhibited significantly greater startle responses at each successive increase in stimulus intensity (i.e., 110 dB > 100 dB > 90 dB) (all p < 0.001). The Psychosocial Stress × Auditory Stimulus Intensity interaction was also significant, F(2,28) = 5.78, p < 0.01. Post hoc analyses revealed that the psychosocial stress group exhibited significantly greater startle responses than the no psychosocial stress group to the 90 and 110 dB (all p < 0.05), but not 100 dB, auditory stimuli.
Radial-arm water maze
For RAWM training, there was a significant main effect of trials, F(11,176) = 11.81, p < 0.0001, indicating that rats made significantly fewer errors as trials progressed. There was no significant main effect of psychosocial stress, F(1,15) = 0.16, and the Psychosocial Stress × Trial interaction was not significant, F(11,176) = 1.13 (all p>0.05). There were no significant group differences on the 1-h, t(16) = 0.00, or 24-h, t(14) = 0.13, memory test trials (all p>0.05; data not shown).
Novel object recognition
In the NOR task, the psychosocial stress and no psychosocial stress groups demonstrated comparable behavior during acclimation to the open field. There were no significant group differences for distance traveled, t(15) = 1.35, rearing, t(16) = 0.80, time in the center area, t(16) = 0.12, or time in the perimeter area, t(16) = 0.13 (all p>0.05; data not shown). During NOR training, the psychosocial stress, t(8) = 0.28, and no psychosocial stress, t(8) = 0.52, groups spent an equivalent amount of time with Object 1 and Object 2, indicating no preference for one object over the other (all p>0.05; ). During the memory test, however, there were differences between the psychosocial stress and no psychosocial stress groups. While the no psychosocial stress group spent significantly more time with the novel than the familiar object, t(8) = 3.05, p < 0.05, the psychosocial stress group failed to discriminate between the objects and spent an equivalent amount of time with each object, t(8) = 0.62, p>0.05.
Figure 4 Novel object recognition training and testing data from Experiment 1. During training, the psychosocial stress and no psychosocial stress groups spent a comparable amount of time with Object 1 and Object 2, indicating no preference for one object over the other (left). Three hours later, the no psychosocial stress group spent more time with the novel than the familiar object, but the psychosocial stress group spent a comparable amount of time with each object (right). *p < 0.05 versus familiar object. Data are group means ± SEM.
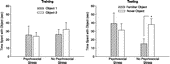
Heart rate and blood pressure
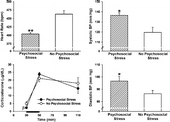
On Day 36, the psychosocial stress group displayed significantly lower HR, t(13) = 4.57, p < 0.001, but significantly greater systolic BP, t(12) = 2.92, p < 0.05, and significantly greater diastolic BP, t(14) = 2.15, p < 0.05, than the no psychosocial stress group following 20 min of acute restraint stress ().
Figure 5 Cardiovascular responses and serum corticosterone on the final day of behavioral testing in Experiment 1. The psychosocial stress group displayed reduce HR (top left), greater systolic BP (top right) and greater diastolic BP (bottom right) than the no psychosocial stress group following 20 min of restraint stress. The psychosocial stress group exhibited marginally reduced baseline levels of corticosterone than the no psychosocial stress group, and both groups demonstrated elevated corticosterone levels following 20 min of acute restraint stress, which is indicated by the dark black line from 30–50 min(bottom left). βp < 0.09; *p < 0.05; **p < 0.001 versus the no psychosocial stress group. Data are group means ± SEM.
Serum corticosterone
For serum corticosterone concentrations on Day 36, there was a significant main effect of time, F(2,26) = 30.61, p < 0.0001. Both groups (i.e., psychosocial stress and no psychosocial stress) showed a significant increase in corticosterone following 20 min of acute restraint (50 min time point in ), and these levels remained elevated one hour later (110 min time point in ) (all p < 0.05). There was no significant main effect of psychosocial stress, F(1,13) = 0.33, and the Psychosocial Stress × Time interaction was not significant, F(2,26) = 1.20 (all p>0.05). Since a majority of the PTSD literature has indicated that PTSD patients have abnormally low baseline levels of cortisol, we compared the baseline levels of corticosterone in the psychosocial stress and no psychosocial stress groups. A planned comparison indicated that the difference in baseline levels of corticosterone between these two groups was marginally significant, t(16) = 1.82, p = 0.087, indicating that the psychosocial stress group tended to have lower corticosterone levels than the no psychosocial stress group before the acute restraint.
Experiment 2: Replication of Experiment 1 without physiological assessments at the time of acute predator stress effects
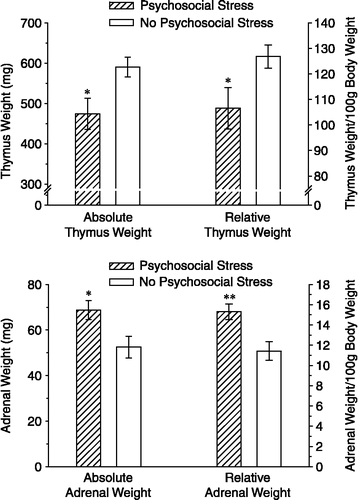
The psychosocial stress group exhibited a significantly reduced growth rate, t(16) = 2.42, p < 0.05, and had significantly larger adrenal glands, [raw weight: t(17) = 2.57, p < 0.05; mg/100 g body weight: t(17) = 3.24, p < 0.01) and a significantly smaller thymus, [raw weight: t(17) = 2.59, p < 0.05; mg/100 g body weight: t(17) = 2.24, p < 0.05], then the no psychosocial stress group ( and ). Therefore, eliminating the acute stress of cardiovascular measurements on Days 1 and 11 from the control group revealed a psychosocial stress effect on body, and organ weights comparable to those observed in other laboratories (Karst and Joels Citation2003).
Figure 6 Adrenal gland and thymus weights from Experiment 2. Data are shown as actual weights (left side of each figure) and relative weights expressed as mg/100 g body weight (right side of each figure). Three weeks after the second cat exposure, the psychosocial stress group demonstrated a smaller thymus (top) and larger adrenal glands (bottom) than the no psychosocial stress group. *p < 0.05; **p < 0.01 versus the no psychosocial stress group. Data are group means ± SEM.
Experiment 3: Is the combination of predator exposure and social instability necessary for PTSD-like effects?
Body weight
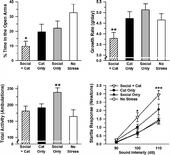
Two-way ANOVAs were used to analyze the growth rate and EPM data (). Analysis of growth rate revealed a significant main effect of inescapable cat exposure, F(1,36) = 4.59, p < 0.05. There was no significant main effect of daily social stress, F(1,36) = 0.58, p>0.05, but the Inescapable Cat Exposure × Daily Social Stress interaction was significant, F(1,36) = 5.79, p < 0.05. Only the two manipulations combined led to a significant reduction in growth rate, relative to the no stress group (p < 0.05).
Figure 7 Elevated plus maze growth rate and startle response data from Experiment 3. Only the cat exposure + social stress group spent significantly less time in the open arms of the elevated plus maze (top left) and exhibited significantly reduced growth rate (top right), relative to the no stress group. The social stress only group displayed greater motor activity on the elevated plus maze than all other groups (bottom left). Only the social stress + cat exposure group exhibited greater startle responses than the no stress group to the 100 and 110 dB auditory stimuli (bottom right). *p < 0.05 versus the no stress group; **p < 0.05 versus all other groups; ***p < 0.01 versus the no stress group. Data are group means ± SEM.
Elevated plus maze
Analysis of time spent in the open arms of the EPM revealed a significant main effect of inescapable cat exposure, F(1,32) = 4.56, p < 0.05, indicating that the groups given inescapable cat exposure on Days 1 and 11 of the experiment spent significantly less time in the open arms of the EPM than non-exposed groups. There was no significant main effect of daily social stress, F(1,32) = 2.69, and the Inescapable Cat Exposure × Daily Social Stress interaction was not significant, F(1,32) = 0.09 (all p>0.05). An inspection of the data suggested that the effects of inescapable cat exposure and daily social stress on EPM behavior were additive, so we used Bonferroni-corrected t-tests to compare each of the experimental groups to the no stress controls. We found that groups given inescapable cat exposure in conjunction with daily social stress, t(14) = 3.35, p < 0.05, but not inescapable cat exposure, t(16) = 1.67, p>0.05, or daily social stress, t(16) = 1.40, p>0.05, alone, spent significantly less time in the open arms than the no stress group.
An analysis of ambulations on the EPM revealed no significant main effect of inescapable cat exposure, F(1,32) = 1.00, p>0.05. There was a significant main effect of daily social stress, F(1,32) = 4.42, p < 0.05, indicating that groups exposed to daily social stress made significantly more ambulations on the EPM than the non-exposed groups. The Inescapable Cat Exposure × Daily Social Stress interaction was also significant, F(1,32) = 7.60, p < 0.01. Post hoc tests revealed that daily social stress, alone, significantly increased ambulatory activity on the EPM, but when combined with inescapable cat exposure, daily social stress had no effect.
Startle response
We used a mixed-model ANOVA to analyze the startle data (). The analysis revealed a significant main effect of auditory stimulus intensity, F(2,62) = 120.7, p < 0.0001. Post hoc tests indicated that rats exhibited significantly greater startle responses at each successive increase in stimulus intensity (i.e., 110 dB > 100 dB > 90 dB) (all p < 0.001). There was also a significant main effect of inescapable cat exposure, F(1,31) = 20.71, p < 0.001, and a significant Auditory Stimulus Intensity × Inescapable Cat Exposure interaction, F(2,62) = 10.85, p < 0.001. Post hoc tests indicated that the groups given inescapable cat exposure on Days 1 and 11 of the experiment demonstrated significantly greater startle responses to the 100 and 110 dB, but not 90 dB, auditory stimuli than non-exposed groups. There was no significant main effect of daily social stress, F(1,31) = 2.62, p>0.05. The Inescapable Cat Exposure × Daily Social Stress interaction was marginally significant, F(1,31) = 3.50, p = 0.071. Similar to behavior on the EPM, an inspection of the data suggested that the effects of inescapable cat exposure and daily social stress on startle were additive, so we used Bonferroni-corrected t-tests to compare each of the experimental groups' overall startle responses (i.e., average startle responses to all three stimulus intensities) to those of the no stress controls. We found that groups given inescapable cat exposure in conjunction with daily social stress, t(14) = 2.97, p < 0.05, but not inescapable cat exposure, t(16) = 1.57, p>0.05, or daily social stress, t(16) = 0.18, p>0.05, alone, exhibited significantly greater startle responses than the no stress group. The Auditory Stimulus Intensity × Daily Social Stress, F(2,62) = 1.67, p>0.05, and Auditory Stimulus Intensity × Inescapable Cat Exposure × Daily Social Stress, F(2,62) = 1.83, p>0.05, interactions were not significant. Furthermore, we used Bonferroni-corrected t-tests to compare each of the experimental groups to the no stress controls for these 100 and 110 dB auditory stimuli. We found that groups given inescapable cat exposure in conjunction with daily social stress [100 dB: t(14) = 2.47, p < 0.05; 110 dB: t(14) = 4.99, p < 0.01] but not inescapable cat exposure [100 dB: t(16) = 1.17, p>0.05; 110 dB: t(16) = 1.80, p>0.05] or daily social stress [100 dB: t(16) = 0.06, p>0.05; 110 dB: t(16) = 0.27, p>0.05], alone, exhibited significantly greater startle responses to the 100 and 110 dB auditory stimuli than the no stress group.
Experiment 4: Effects of psychosocial stress on behavioral and corticosterone responses to yohimbine
Body weight
The psychosocial stress groups (i.e., S-VEH and S-YOH) exhibited a significantly reduced growth rate, when compared with the no psychosocial stress groups (i.e., NS-VEH and NS-YOH), t(37) = 3.13, p < 0.01 ().
Open field
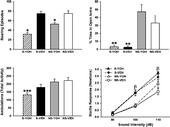
Two-way ANOVAs were used to analyze the open field and EPM data (). For a number of rearing episodes in the open field there were significant main effects of psychosocial stress, F(1,32) = 6.00, p < 0.05, and yohimbine, F(1,32) = 55.63, p < 0.0001, indicating that the psychosocial stress (S-VEH, S-YOH) reduced rearing relative to the no psychosocial stress groups (NS-VEH, NS-YOH) and yohimbine treatment (NS-YOH, S-YOH) reduced rearing relative to the vehicle-treated groups (NS-VEH, S-VEH). There was also a significant Psychosocial Stress × Yohimbine interaction, F(1,32) = 5.59, p < 0.05. Post hoc tests indicated that the yohimbine-treated psychosocial stress group (S-YOH) displayed significantly fewer rearing episodes than all other groups (NS-VEH, NS-YOH, S-VEH), revealing that yohimbine exerted a greater behavioral effect on the psychosocial stress group than on the no psychosocial stress group (all p < 0.01).
Figure 8 Open field, elevated plus maze and startle findings from Experiment 4. In open field testing, yohimbine treatment (1 mg/kg, i.p.) suppressed rearing in the no psychosocial stress group (NS-YOH), an effect that was significantly exacerbated by prior stress experience (S-YOH) (top left). On the elevated plus maze, both psychosocial stress groups (S-YOH, S-VEH) spent significantly less time in the open arms than the no psychosocial stress groups (NS-YOH, NS-VEH) (top right). In addition, only the yohimbine-treated psychosocial stress group (S-YOH) exhibited significantly less motor activity on the elevated plus maze than the vehicle-treated no psychosocial stress group (NS-VEH) (bottom left). Psychosocial stress significantly increased startle responses to the 100 and 110 dB auditory stimuli (bottom right). *p < 0.01 versus all other groups; **p < 0.0001 versus the no psychosocial stress groups; ***p < 0.01 versus the vehicle-treated no psychosocial stress group; βp < 0.05 versus the respective no psychosocial stress groups. Data are group means ± SEM.
Elevated plus maze
Analysis of time spent in the open arms of the EPM revealed a significant main effect of psychosocial stress, F(1,32) = 70.29, p < 0.0001, indicating that the psychosocial stress groups (S-VEH, S-YOH) spent significantly less time in the open arms than the no psychosocial stress groups (NS-VEH, NS-YOH). There was no significant main effect of yohimbine, F(1,32) = 0.18, and the Psychosocial Stress × Yohimbine interaction was not significant, F(1,32) = 0.34 (all p>0.05). An analysis of ambulations made on the EPM revealed a significant main effect of psychosocial stress, F(1,33) = 10.93, p < 0.01, indicating that the psychosocial stress groups (S-VEH, S-YOH) made significantly fewer ambulations than no psychosocial stress groups (NS-VEH, NS-YOH). There was no significant main effect of yohimbine, F(1,33) = 2.32, and the Psychosocial Stress × Yohimbine interaction was not significant, F(1,33) = 1.10 (all p>0.05). An inspection of the data suggested that the effects of psychosocial stress and yohimbine on EPM behavior were additive. Therefore, we used Bonferroni-corrected t-tests to compare each of the experimental groups to the vehicle-treated no psychosocial stress group. We found that the yohimbine-treated psychosocial stress group (S-YOH), t(15) = 3.85, p < 0.01, but not the yohimbine-treated no psychosocial stress group (NS-YOH), t(18) = 0.31, p>0.05, or vehicle-treated psychosocial stress group (S-VEH), t(14) = 1.52, p>0.05, made significantly fewer ambulations on the EPM than the no psychosocial stress group (NS-VEH).
Startle response
A mixed-model ANOVA was used to analyze the startle () and corticosterone () data. There was a significant main effect of auditory stimulus intensity, F(2,60) = 191.65, p < 0.0001, indicating that rats exhibited significantly greater startle responses at each successive increase in stimulus intensity (i.e., 110 dB > 100 dB > 90 dB) (all p < 0.001). There was also a significant main effect of psychosocial stress, F(1,30) = 21.14, p < 0.001, and a significant Auditory Stimulus Intensity × Psychosocial Stress interaction, F(2,60) = 6.54, p < 0.01. Post hoc tests indicated that the psychosocial stress groups (S-YOH, S-VEH) exhibited significantly greater startle responses to the 100 and 110 dB, but not 90 dB, auditory stimuli than the no psychosocial stress groups (NS-YOH, NS-VEH, all p < 0.05). There was no significant main effect of yohimbine, F(1,30) = 1.30, p>0.05, but the Auditory Stimulus Intensity × Yohimbine interaction was marginally significant, F(2,60) = 3.06, p = 0.054. Post hoc tests indicated that the yohimbine-treated groups (S-YOH, NS-YOH) displayed significantly greater startle responses to the 110 dB, but not 90 or 100 dB, auditory stimuli than the vehicle-treated groups (S-VEH, NS-VEH, p < 0.05). The Psychosocial Stress × Yohimbine, F(1,30) = 0.14, and Auditory Stimulus Intensity × Psychosocial Stress × Yohimbine, F(2,60) = 0.04, interactions were not significant (all p>0.05).
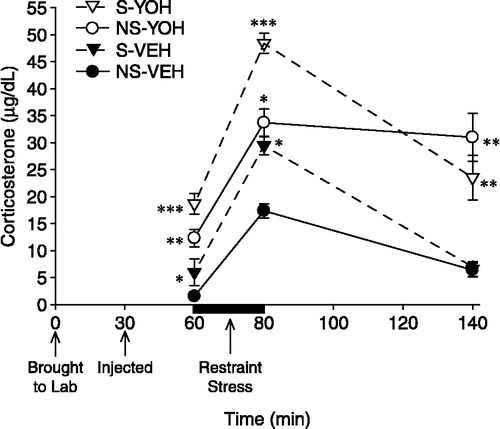
Figure 9 Serum corticosterone responses in Experiment 4. On Day 33 of Experiment 4, rats were given yohimbine (1 mg/kg, i.p.) or vehicle. The psychosocial stress and yohimbine-treated groups exhibited greater serum corticosterone concentrations 30 min post-injection (60 min time point) than the no psychosocial stress and vehicle-treated groups, respectively. Corticosterone levels were increased in all groups following 20 min of restraint (indicated by the dark black line between 60–80 min); the increases were greater in groups that had been psychosocially stressed and/or pre-treated with yohimbine. Psychosocial stress and yohimbine treatment had an additive effect on corticosterone levels at each of the first two time points: the group that had been psychosocially stressed and pre-treated with yohimbine (S-YOH) exhibited the greatest corticosterone levels at both time points. While corticosterone levels of the two vehicle-treated groups (S-VEH, NS-VEH) significantly declined 60 min following the cessation of immobilization, those of the two yohimbine-treated groups (S-YOH, NS-YOH) remained elevated. *p < 0.05 versus the no psychosocial stress-vehicle group; **p < 0.05 versus psychosocial stress-vehicle and no psychosocial stress-vehicle groups; ***p < 0.05 versus all other groups. Data are group means ± SEM.
Serum corticosterone
For corticosterone concentrations on Day 33, there was a significant main effect of time, F(2,46) = 71.18, p < 0.0001. Corticosterone levels increased significantly after 20 min of restraint (80 min time point in ) and declined 60 min later (140 min time point in ), but remained elevated when compared with the first time point (i.e., 60 min time point in ) (all p < 0.05). The Time × Psychosocial Stress interaction was significant, F(2,46) = 10.65, p < 0.001. The psychosocial stress groups (S-YOH, S-VEH) exhibited significantly greater corticosterone levels of 30 min post-injection (60 min time point in ) and following 20 min of restraint than the no psychosocial stress groups (NS-YOH, NS-VEH) (p < 0.05). Corticosterone levels of the psychosocial stress and no psychosocial stress groups did not differ 60 min later (p>0.05). The Time × Yohimbine interaction was significant, F(2,46) = 3.82, p < 0.05. Yohimbine led to significantly greater corticosterone levels 30 min post-injection and following 20 min of restraint. While corticosterone levels of vehicle-treated rats significantly declined 60 min later, those of yohimbine-treated rats remained elevated (all p < 0.05).
Discussion
The primary finding of this series of experiments is that rats exposed to psychosocial stressor composed of two 1-h periods of inescapable cat exposure, in conjunction with daily unstable housing conditions, exhibited reduced growth rate, reduced thymus weight, greater adrenal gland weight, increased anxiety, an exaggerated startle response, impaired memory, greater cardiovascular and corticosterone reactivity to an acute stressor and an exaggerated physiological and behavioral response to the α2-adrenergic receptor antagonist, yohimbine. These psychosocial stress-induced changes in rat behavior and pharmacological responses are comparable to commonly observed sequelae in people with PTSD, which strengthens the validity of this psychosocial stress paradigm as an animal model of PTSD.
Rationale for the PTSD model paradigm
The design of our animal model of PTSD was developed to expose rats to conditions which, based on DSM-IV criteria, are analogous to conditions that produce PTSD in people. Specifically, a subset of the DSM-IV criteria for the diagnosis of PTSD includes the following three conditions: (1) PTSD can be triggered by an event that involves threatened death or a threat to one's physical integrity, (2) a person's response to the event involves intense fear, helplessness or horror and (3) in the aftermath of the trauma, the person feels as if the traumatic event is recurring, including a sense of reliving the experience (American Psychiatric Association Citation1994).
The behaviors that rats exhibit in response to forced exposure to a cat are consistent with the first two components of the DSM-IV criteria for PTSD. That is, rats exhibit an intense fear response when exposed to a predator, clearly a condition that is a threat to their survival. In addition, we have observed that rats typically direct their posture away from the cat's gaze, which provides the rat with an element of control over its confrontation with the cat. As control critically influences the expression of the stress response, in general (Kim and Diamond Citation2002), and a loss of control exacerbates behavioral and physiological responses to stress conditions (Amat et al. Citation2005, Citation2007, Watkins Citation2005), we immobilized the rats during predator exposure. The immobilization component of our animal model, therefore, provides a rodent analogue to the sense of helplessness and a loss of control which feature prominently in the DSM-IV criteria for PTSD.
Another component of our model was that rats were exposed to the cat on two occasions, separated by 10 days. There were four reasons for this. First, PTSD develops in some people only after they have repeated traumatic experiences (Resnick et al. Citation1995; Taylor and Cahill Citation2002) and prolonged exposure to trauma increases the likelihood of developing symptoms of PTSD (Gurvits et al. Citation1996). Therefore, the repeated inescapable cat exposure was designed. Therefore, the repeated inescapable cat exposure was designed to increase the likelihood that the manipulations would produce effects in the rats that could be broadly applied to people who develop PTSD as a result of multiple traumatic experiences.
Second, people who develop PTSD in response to only a single trauma experience powerful episodes of anxiety and panic as a result of their repeated reliving of the trauma through intrusive, flashback memories (Reynolds and Brewin Citation1999). In a similar way, the second exposure of the rats to the cat forced them to re-experience the original stress experience.
The third reason why the rats were re-exposed to the cat pertained to the issue of predictability. The first predator exposure occurred during the light cycle and the second predator exposure occurred during the dark cycle, thereby adding an element of unpredictability as to when the rats might re-experience the traumatic event. A lack of predictability in one's environment is a major factor in the development and expression of PTSD, at least in a subset of susceptible people (Orr et al. 1990; Regehr et al. 2000; Solomon et al. 1988, 1989).
Fourth, McEwen and colleagues observed increased spine density on dendritic arbors of amygdala neurons 10 days after a single immobilization experience (Mitra et al. Citation2005). Therefore, the second stress session, 10 days after the first, reinforced stress-induced changes in brain and behavior, which were presumably initiated by the first stress session. Any reinforcement of morphological plasticity in the amygdala through a reminder of the original experience might augment the PTSD-like syndrome in psychosocially stressed rats. The presumed predator stress-induced strengthening of plasticity in the amygdala, which might include dendritic hypertrophy (Fuchs et al. Citation2006, Chattarji Citation2004, Citation2003, Citation2006) or stress-induced LTP (Richter-Levin Citation2006, Citation2006), lends itself to experimentation via pharmacological manipulations of the reconsolidation process, which is likely to occur in response to traumatic memory recall (Cai et al. Citation2006, LeDoux Citation2004, Citation2006).
In addition to the two acute cat exposures, we included chronic unstable housing conditions in the psychosocial stress paradigm to mimic the lack of social support and chronic mild stress experienced by people with PTSD (Andrews et al. Citation2003, Filipas Citation2001). We hypothesized that the daily anxiety produced by unstable housing would exacerbate any adverse effects on the rats induced by predator exposure alone. This hypothesis was supported by our finding that the combination of two cat exposures and social instability produced greater anxiogenic effects on rat behavior than either manipulation in isolation. Indeed, chronic social instability, alone, had no negative effects on behavior and may have even been beneficial for rats, as it led to a small increase in growth rate and significantly greater motor activity on the EPM. Therefore, the physiological and behavioral effects that we have observed here occurred only when rats experienced two episodes of an acute life-threatening stress (cat exposure) in conjunction with chronic social instability.
Finally, a methodological difference between our PTSD model and the approaches of other groups is that we kept the rats separated from the cat by a transparent barrier. Adamec and colleagues (Adamec Citation1997, Citation2006, Citation2007, Shallow Citation1993) have utilized a physical attack by the cat on the rat. Our approach was designed to induce a persistent state of fear and a loss of control in the rats by maintaining them in close, but unavoidable, proximity to the cat for 45 min, without introducing confounds that may arise from physical alterations (i.e. immune response to a wound). Since direct predator attack and indirect threat are both effective at producing lasting changes in rodent anxiety behavior, the two strategies can serve as complementary models of PTSD.
Psychosocial stress-induced changes in anxiety-like behavior and startle
As with other animal models of PTSD (Adamec Citation1997, Citation2007, Liberzon Citation2004), we found that psychosocially stressed rats spent significantly less time in exploring the open arms of the EPM and demonstrated greater startle responses than control rats. These findings indicate that the present psychosocial stress paradigm produced persistent heightened anxiety and hyperarousal, two important components of PTSD. In Experiment 4, we replicated these effects, but we did not observe an exacerbation of either effect by the administration of yohimbine. Since yohimbine induces flashbacks and panic attacks in PTSD patients, we expected an even greater anxiogenic response on these tasks in psychosocially stressed rats. Nevertheless, it is important to consider three issues related to the EPM and startle data from Experiment 4. First, psychosocial stress, alone, had such a potent effect on EPM behavior that it would have been difficult to observe any exacerbation. For instance, the vehicle-treated psychosocial stress group almost completely avoided the open arms of the EPM (spending < 3% of their time in the open arms). Second, the same issue observed for the EPM data may have contributed to the startle data. Psychosocial stress, alone, may have led to a maximal startle response which could not be further intensified. Nevertheless, there was a clear yohimbine-mediated enhancement of the neuroendocrine (corticosterone) response to acute restraint in these rats. Thus, the multiple behavioral tests administered may have been relatively insensitive to reveal a yohimbine-mediated enhancement of anxiety, but at an endocrine level, the psychosocial stress group exhibited a synergy between their prior stress experiences and reactivity to yohimbine, which is consistent with yohimbine effects in people with PTSD.
Task-specific effects of psychosocial stress on memory
In Experiment 1, we found no effect of chronic psychosocial stress on learning and memory in the RAWM, a hippocampus-dependent task. Although this lack of effect of chronic stress on hippocampus-dependent memory was surprising, methodological considerations can explain why the psychosocial stress group did not exhibit impaired performance in the water maze. Extensive research has reported cognitive deficits in PTSD patients, but these deficits have primarily been found in people tested on emotionally neutral information, such as word lists (Bremner et al. 2004; Gilbertson et al. 2001; Yehuda et al. 2005, 2006). In contrast, people with PTSD exhibit greater conditioned fear (Orr et al. Citation2000; Peri et al. Citation2000) and enhanced attention and memory for emotionally arousing information (Golier et al. 2003; McNally 1997; Moradi et al. 1999a,b; Paunovic et al. 2002). Thus, it is possible that we did not observe impaired learning and memory in psychosocially stressed rats because water maze training is a highly arousing task.
In other animal models of PTSD, the delayed effects of intense, acute stressors on water maze performance have been inconsistent. While some studies have reported slower rates of acquisition, as well as impaired memory, in previously stressed animals (Cohen et al. Citation2003; Harvey et al. Citation2003; Kohda et al. Citation2007), others have found that intense, aversive stressors applied only within the context of the learning environment result in a delayed impairing effect on water maze performance (Richter-Levin Citation1998). Further, the aforementioned animal models of PTSD reporting water maze deficits involved training rats in a pool of water that was warmer (23–26°C) than the water employed in the present study (21–22°C). Previous work has shown water temperature influences the stress response in rats and subsequent performance in the water maze (Sandi et al. Citation1997; Wright and Conrad Citation2007). In addition, animal studies have shown that prior stress enhances learning in stressful tasks (Conrad et al. Citation1999; Cordero et al. Citation2003; Sandi et al. Citation2001), and chronic stress can even enhance spatial learning and memory in a Morris water maze filled with cold (18–22°C) water (Liu et al. Citation2004). Therefore, the lack of an impairing effect of psychosocial stress in rats trained in the water maze, an emotionally arousing task, is actually consistent with studies employing similar methods in the rodent literature.
Importantly, psychosocially stressed rats in the current study did exhibit impaired memory when they were trained in an emotionally neutral task involving their memory of novel objects. This finding is consistent with work reporting chronic stress-induced impairments of memory for a novel arm in the Y-maze, another low stress hippocampus-dependent memory task (Conrad Citation2005; Wright and Conrad Citation2005). It also coincides with numerous others studies reporting chronic stress-induced impairments of object recognition memory (Luine Citation2002). Our findings, therefore, are consistent with the findings from studies on rodents and people indicating that chronic stress exerts a greater adverse effect on new memory processing for emotionally neutral, and not arousing information.
Psychosocial stress and cardiovascular reactivity
Experiment 1 confirmed that acute immobilization during cat exposure significantly increased HR, BP and corticosterone release. We also found that psychosocially stressed rats exhibited a significant elevation of systolic and diastolic BP in response to acute restraint on the final day of behavioral testing, three weeks after the last predator exposure. This finding is potentially relevant to cardiovascular sequelae common in people with PTSD. Chronic anxiety and elevations of BP are significant risk factors for cardiovascular disease (Haider et al. Citation2003; Izzo et al. Citation2000), and PTSD has been associated with increased risk for cardiovascular disorders (Chang Citation1999). Therefore, our model may provide insight into how traumatic stress contributes to the development and progression of cardiovascular disease.
Surprisingly, rats given psychosocial stress exhibited lower HR, but higher BP, than previously unstressed rats in response to acute restraint on the final day of behavioral testing. Bruijnzeel et al. (Citation2001) similarly found that exposing rats to a single session of scrambled electric footshocks led to long-term sensitization of their cardiovascular stress response. Two weeks after footshock exposure, previously shocked rats displayed significantly greater elevations of BP, but not HR, than non-shocked rats in response to a novel stressor. Comparable research in people (Halligan et al. Citation2006) showed that individuals with assault-related PTSD exhibited a smaller HR response to voluntary recollections of their trauma than trauma-exposed controls.
Psychosocially stressed rats in the present study had lower HR in response to restraint perhaps because their multiple prior stressor experiences protected them against responding as strongly as naïve animals, specifically with regard to HR. However, despite a lower HR, the psychosocially stressed rats still exhibited significantly greater systolic and diastolic BP than controls. This finding suggests that chronic psychosocial stress may have lasting effects on regulatory systems that modulate cardiovascular functioning, such as the parasympathetic nervous system. Extensive work has suggested that people with PTSD exhibit reduced parasympathetic tone, which contributes to heightened baseline and stress-induced elevations of sympathetic activity (Cohen et al. 1998, 2000a; Hopper et al. 2006; Sack et al. 2004; Sahar et al. 2001). In addition, some animal models of PTSD have been reported to show reduced parasympathetic tone in maladapted, stress-exposed rodents (Cohen et al. Citation2003). Thus, the failure of the psychosocially stressed group to demonstrate comparable HR and BP to the no psychosocial stress group may reflect pathological effects of stress on interactions between parasympathetic and sympathetic activity, as well as stress-induced changes in blood vessel integrity and elasticity (Consoli Citation1993, Kolarova Citation1993).
Corticosterone and psychosocial stress in rats
On the final day of testing in Experiment 1, psychosocially stressed rats exhibited baseline corticosterone levels that were marginally (p = 0.087) lower than those of control rats. This finding is consistent with extensive work finding lower baseline cortisol levels in people with PTSD (Yehuda Citation2005) and suggests that the chronic psychosocial stress employed in the present study may have long-term effects on basal HPA axis functioning. Comparable findings have been reported in some animal studies examining the effects of stress–restress or single prolonged stress paradigms on rat physiology (e.g., Harvey et al. Citation2003). In Experiment 4, psychosocially stressed rats exhibited significantly greater corticosterone levels than controls 30 min after injection with vehicle and following 20 min of acute restraint stress. The finding of greater corticosterone levels in psychosocially stressed rats following injection and acute restraint is consistent with findings from PTSD patients who exhibit greater cortisol levels in anticipation of anxiety-provoking situations (Bremner et al. Citation2003) and in response to trauma-related stimuli (Elzinga et al. Citation2003).
We did observe differences in corticosterone levels, both at baseline and in response to acute restraint in Experiments 1 and 4. Unlike the findings of Experiment 4, we did not observe significant group differences in corticosterone levels in Experiment 1. We suggest three reasons for the different findings between the two experiments. First, in Experiment 1, rats in the psychosocial stress, as well as the no psychosocial stress, group were exposed to the cardiovascular measurement apparatus, with blood sampling in all groups on Days 1 and 11 of the paradigm. These manipulations appear to have produced a lasting negative influence on the control animals, as indicated by their reduced weight gain. The control group in Experiment 2 that was not restrained or exposed to blood sampling on Days 1 and 11 gained significantly more weight than the psychosocial stress group. Thus, the acute stress on Days 1 and 11 given to all animals may have influenced baseline and acute stress-evoked corticosterone levels at the conclusion of behavioral testing. Second, in Experiment 1, the rats had undergone several days of behavioral testing prior to the collection of pre- and post-stress blood samples. Less behavioral testing was conducted in Experiment 4 before acute stress and blood sampling was performed. Third, in Experiment 4 the rats were injected with either drug (yohimbine) or vehicle (saline) before the initial blood sampling. Thus, the injections in Experiment 4 likely induced a stress response in rats before the first blood sample was obtained, which may have affected corticosterone levels before, as well as after, acute restraint 30 min later. In future work, baseline and acute stress-evoked levels of corticosterone are needed from rats that have minimal exposure to these extraneous influences.
In Experiment 4, yohimbine increased corticosterone levels in both stressed and unstressed rats within 30 min. This finding is consistent with human and rodent research indicating that yohimbine increases HPA activity and anxiety-like behavior (Grunhaus et al. Citation1989; McDougle et al. Citation1995; Myers et al. Citation2005; Vythilingam et al. Citation2000). We also found that psychosocially stressed rats treated with yohimbine showed greater increases in corticosterone level following acute restraint than control rats treated with yohimbine, which is consistent with well-documented evidence of greater yohimbine-induced increases in physiological activity in PTSD patients (Southwick et al. Citation1993). Overall, our findings, in conjunction with studies on people with PTSD, indicate that yohimbine sensitivity can serve as a diagnostic of behavioral and physiological abnormalities produced by traumatic stress.
Summary
We have found that two episodes of inescapable cat exposure, in conjunction with chronic social instability, produced a cluster of physiological and behavioral changes in rats (e.g., heightened anxiety, exaggerated startle, impaired cognition, heightened physiological reactivity to an acute stressor, exaggerated response to yohimbine), which are comparable to those observed in people with PTSD. There was also an indication that this paradigm results in reduced baseline HPA axis functioning. Future studies are needed to substantiate this preliminary finding and should also test the hypothesis that psychosocially stressed rats would exhibit enhanced negative feedback of the HPA axis. As PTSD is a memory disorder characterized by an abnormally powerful, often invasive, memory for a traumatic event, future work with the present rat model will evaluate memory for the traumatic stress experiences.
An increase in the magnitude of the trauma is related to a greater incidence of PTSD (Maercker et al. Citation2000; Pynoos et al. Citation1993; Sutker et al. Citation1993). The psychosocial stress paradigm employed here produced a powerful and homogeneous response in rats, which can be viewed as a model of how the most intense and horrific traumatic experiences affect people. As such, our psychosocial stress model may provide insight into the etiology and maintenance of PTSD, as well as to serve in the development of treatment strategies to ameliorate PTSD symptoms.
Acknowledgements
The authors thank Edward Levine, Kristen Salomon and Paula Bickford for their constructive input regarding the design of the experiments. We also appreciate the constructive criticisms of an earlier version of this work by the three anonymous reviewers and the editor. This work was supported by a VA Merit Review Award to DMD and an NIMH grant to CDC.
References
- Adamec R. Transmitter systems involved in neural plasticity underlying increased anxiety and defense—implications for understanding anxiety following traumatic stress. Neurosci Biobehav Rev 1997; 21: 755–765
- Adamec RE, Shallow T. Lasting effects on rodent anxiety of a single exposure to a cat. Physiol Behav 1993; 54: 101–109
- Adamec RE, Burton P, Shallow T, Budgell J. NMDA receptors mediate lasting increases in anxiety-like behavior produced by the stress of predator exposure: Implications for anxiety associated with posttraumatic stress disorder. Physiol Behav 1999; 65: 723–737
- Adamec RE, Blundell J, Burton P. Relationship of the predatory attack experience to neural plasticity, pCREB expression and neuroendocrine response. Neurosci Biobehav Rev 2006; 30: 356–375
- Adamec R, Muir C, Grimes M, Pearcey K. Involvement of noradrenergic and corticoid receptors in the consolidation of the lasting anxiogenic effects of predator stress. Behav Brain Res 2007; 179: 192–207
- Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci 2005; 8: 365–371
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders: DSM-IV4th ed. American Psychiatric Association, Washington, D.C. 1994
- Andrews B, Brewin CR, Rose S. Gender, social support, and PTSD in victims of violent crime. J Trauma Stress 2003; 16: 421–427
- Baker KB, Kim JJ. Effects of stress and hippocampal NMDA receptor antagonism on recognition memory in rats. Learn Mem 2002; 9: 58–65
- Beck KD, Luine VN. Sex differences in behavioral and neurochemical profiles after chronic stress: Role of housing conditions. Physiol Behav 2002; 75: 661–673
- Blanchard EB, Kolb LC, Pallmeyer TP, Gerardi RJ. A psychophysiological study of post traumatic stress disorder in Vietnam veterans. Psychiatry Q 1982; 54: 220–229
- Blanchard RJ, Blanchard DC, Rodgers J, Weiss SM. The characterization and modeling of antipredator defensive behavior. Neurosci Biobehav Rev 1990; 14: 463–472
- Blanchard EB, Kolb LC, Prins A, Gates S, McCoy GC. Changes in plasma norepinephrine to combat-related stimuli among Vietnam veterans with posttraumatic stress disorder. J Nerv Ment Dis 1991; 179: 371–373
- Blanchard RJ, Nikulina JN, Sakai RR, McKittrick C, McEwen B, Blanchard DC. Behavioral and endocrine change following chronic predatory stress. Physiol Behav 1998; 63: 561–569
- Blanchard DC, Griebel G, Blanchard RJ. Conditioning and residual emotionality effects of predator stimuli: Some reflections on stress and emotion. Prog Neuro-Psychopharmacol Biol Psychiatry 2003; 27: 1177–1185
- Blanchard DC, Canteras NS, Markham CM, Pentkowski NS, Blanchard RJ. Lesions of structures showing FOS expression to cat presentation: Effects on responsivity to a cat, cat odor, and nonpredator threat. Neurosci Biobehav Rev 2005; 29: 1243–1253
- Bland ST, Schmid MJ, Greenwood BN, Watkins LR, Maier SF. Behavioral control of the stressor modulates stress-induced changes in neurogenesis and fibroblast growth factor-2. Neuroreport 2006; 17: 593–597
- Bland ST, Tamlyn JP, Barrientos RM, Greenwood BN, Watkins LR, Campeau S, Day HE, Maier SF. Expression of fibroblast growth factor-2 and brain-derived neurotrophic factor mRNA in the medial prefrontal cortex and hippocampus after uncontrollable or controllable stress. Neuroscience 2007; 144: 1219–1228
- Boscarino JA. Post-traumatic stress and associated disorders among Vietnam veterans: The significance of combat exposure and social support. J Trauma Stress 1995; 8: 317–336
- Boscarino JA, Chang J. Electrocardiogram abnormalities among men with stress-related psychiatric disorders: Implications for coronary heart disease and clinical research. Ann Behav Med 1999; 21: 227–234
- Bowman RE, Beck KD, Luine VN. Chronic stress effects on memory: Sex differences in performance and monoaminergic activity. Horm Behav 2003; 43: 48–59
- Bremner JD, Vythilingam M, Vermetten E, Adil J, Khan S, Nazeer A, Afzal N, McGlashan T, Elzinga B, Anderson GM, Heninger G, Southwick SM, Charney DS. Cortisol response to a cognitive stress challenge in posttraumatic stress disorder (PTSD) related to childhood abuse. Psychoneuroendocrinology 2003; 28: 733–750
- Bremner JD, Vermetten E, Afzal N, Vythilingam M. Deficits in verbal declarative memory function in women with childhood sexual abuse-related posttraumatic stress disorder. J Nerv Ment Dis 2004; 192: 643–649
- Brewin CR, Andrews B, Valentine JD. Meta-analysis of risk factors for posttraumatic stress disorder in trauma-exposed adults. J Consult Clin Psychol 2000; 68: 748–766
- Bruijnzeel AW, Stam R, Croiset G, Wiegant VM. Long-term sensitization of cardiovascular stress responses after a single stressful experience. Physiol Behav 2001; 73: 81–86
- Buckley TC, Kaloupek DG. A meta-analytic examination of basal cardiovascular activity in posttraumatic stress disorder. Psychosom Med 2001; 63: 585–594
- Cai WH, Blundell J, Han J, Greene RW, Powell CM. Postreactivation glucocorticoids impair recall of established fear memory. J Neurosci 2006; 26: 9560–9566
- Campbell AM, Park CR, Zoladz PR, Munoz C, Fleshner M, Diamond DM. Pre-training administration of tianeptine, but not propranolol, protects hippocampus-dependent memory from being impaired by predator stress. Eur Neuropsychopharmacol 2007, In Press
- Casada JH, Amdur R, Larsen R, Liberzon I. Psychophysiologic responsivity in posttraumatic stress disorder: Generalized hyperresponsiveness versus trauma specificity. Biol Psychiatry 1998; 44: 1037–1044
- Cohen H, Zohar J. An animal model of post-traumatic stress disorder: The use of cut-off behavioral criteria. Ann N Y Acad Sci 2004; 1032: 167–178
- Cohen H, Kotler M, Matar MA, Kaplan Z, Loewenthal U, Miodownik H, Cassuto Y. Analysis of heart rate variability in posttraumatic stress disorder patients in response to a trauma-related reminder. Biol Psychiatry 1998; 44: 1054–1059
- Cohen H, Benjamin J, Geva AB, Matar MA, Kaplan Z, Kotler M. Autonomic dysregulation in panic disorder and in post-traumatic stress disorder: Application of power spectrum analysis of heart rate variability at rest and in response to recollection of trauma or panic attacks. Psychiatry Res 2000a; 96: 1–13
- Cohen H, Benjamin J, Kaplan Z, Kotler M. Administration of high-dose ketoconazole, an inhibitor of steroid synthesis, prevents posttraumatic anxiety in an animal model. Eur Neuropsychopharmacol 2000b; 10: 429–435
- Cohen H, Zohar J, Matar M. The relevance of differential response to trauma in an animal model of posttraumatic stress disorder. Biol Psychiatry 2003; 53: 463–473
- Cohen H, Zohar J, Matar MA, Zeev K, Loewenthal U, Richter-Levin G. Setting apart the affected: The use of behavioral criteria in animal models of post traumatic stress disorder. Neuropsychopharmacology 2004; 29: 1962–1970
- Cohen H, Kaplan Z, Matar MA, Loewenthal U, Kozlovsky N, Zohar J. Anisomycin, a protein synthesis inhibitor, disrupts traumatic memory consolidation and attenuates posttraumatic stress response in rats. Biol Psychiatry 2006; 60: 767–776
- Cohen H, Kaplan Z, Matar MA, Loewenthal U, Zohar J, Richter-Levin G. Long-lasting behavioral effects of juvenile trauma in an animal model of PTSD associated with a failure of the autonomic nervous system to recover. Eur Neuropsychopharmacol 2007; 17: 464–477
- Conrad CD. The relationship between acute glucocorticoid levels and hippocampal function depends upon task aversiveness and memory processing stage. Nonlinearity Biol Toxicol Med 2005; 3: 57–78
- Conrad CD, LeDoux JE, Magarinos AM, McEwen BS. Repeated restraint stress facilitates fear conditioning independently of causing hippocampal CA3 dendritic atrophy. Behav Neurosci 1999; 113: 902–913
- Consoli SM. Stress and the cardiovascular system. Encephale 1993; 19 Spec No 1: 163–170
- Cook SC, Wellman CL. Chronic stress alters dendritic morphology in rat medial prefrontal cortex. J Neurobiol 2004; 60: 236–248
- Cordero MI, Venero C, Kruyt ND, Sandi C. Prior exposure to a single stress session facilitates subsequent contextual fear conditioning in rats: Evidence for a role of corticosterone. Horm Behav 2003; 44: 338–345
- Debiec J, LeDoux JE. Disruption of reconsolidation but not consolidation of auditory fear conditioning by noradrenergic blockade in the amygdala. Neuroscience 2004; 129: 267–272
- Debiec J, LeDoux JE. Noradrenergic signaling in the amygdala contributes to the reconsolidation of fear memory: Treatment implications for PTSD. Ann N Y Acad Sci 2006; 1071: 521–524
- Debiec J, LeDoux JE, Nader K. Cellular and systems reconsolidation in the hippocampus. Neuron 2002; 36: 527–538
- Diamond DM, Park CR, Heman KL, Rose GM. Exposure to a predator impairs spatial working memory in the radial arm water maze. Hippocampus 1999; 9: 542–552
- Diamond DM, Campbell AM, Park CR, Woodson JC, Conrad CD, Bachstetter AD, Mervis RF. Influence of predator stress on the consolidation versus retrieval of long-term spatial memory and hippocampal spinogenesis. Hippocampus 2006; 16: 571–576
- Ehlers A, Hackmann A, Michael T. Intrusive re-experiencing in post-traumatic stress disorder: Phenomenology, theory, and therapy. Memory 2004; 12: 403–415
- Elzinga BM, Bremner JD. Are the neural substrates of memory the final common pathway in posttraumatic stress disorder (PTSD)?. J Affect Disord 2002; 70: 1–17
- Elzinga BM, Schmahl CG, Vermetten E, van Dyck R, Bremner JD. Higher cortisol levels following exposure to traumatic reminders in abuse-related PTSD. Neuropsychopharmacology 2003; 28: 1656–1665
- Faye K, Heng LH, Collomp R, Peroux E. Hypertension and stress. J Mal Vasc 2003; 28: 4–8
- Flaa A, Mundal HH, Eide I, Kjeldsen S, Rostrup M. Sympathetic activity and cardiovascular risk factors in young men in the low, normal, and high blood pressure ranges. Hypertension 2006; 47: 396–402
- Fuchs E, Flugge G, Czeh B. Remodeling of neuronal networks by stress. Front Biosci 2006; 11: 2746–2758
- Garrick T, Morrow N, Shalev AY, Eth S. Stress-induced enhancement of auditory startle: An animal model of posttraumatic stress disorder. Psychiatry 2001; 64: 346–354
- Geracioti TD, Jr., Baker DG, Ekhator NN, West SA, Hill KK, Bruce AB, Schmidt D, Rounds-Kugler B, Yehuda R, Keck PE, Jr., Kasckow JW. CSF norepinephrine concentrations in posttraumatic stress disorder. Am J Psychiatry 2001; 158: 1227–1230
- Gerges NZ, Alzoubi KH, Park CR, Diamond DM, Alkadhi KA. Adverse effect of the combination of hypothyroidism and chronic psychosocial stress on hippocampus-dependent memory in rats. Behav Brain Res 2004; 155: 77–84
- Gilbertson MW, Gurvits TV, Lasko NB, Orr SP, Pitman RK. Multivariate assessment of explicit memory function in combat veterans with posttraumatic stress disorder. J Trauma Stress 2001; 14: 413–432
- Golier JA, Yehuda R, Lupien SJ, Harvey PD. Memory for trauma-related information in Holocaust survivors with PTSD. Psychiatry Res 2003; 121: 133–143
- Grunhaus L, Tiongco D, Zelnik T, Flegel P, Hollingsworth PJ, Smith CB. Intravenous yohimbine: Selective enhancer of norepinephrine and cortisol secretion and systolic blood pressure in humans. Clin Neuropharmacol 1989; 12: 106–114
- Gurvits TV, Shenton ME, Hokama H, Ohta H, Lasko NB, Gilbertson MW, Orr SP, Kikinis R, Jolesz FA, McCarley RW, Pitman RK. Magnetic resonance imaging study of hippocampal volume in chronic, combat-related posttraumatic stress disorder. Biol Psychiatry 1996; 40: 1091–1099
- Hackmann A, Ehlers A, Speckens A, Clark DM. Characteristics and content of intrusive memories in PTSD and their changes with treatment. J Trauma Stress 2004; 17: 231–240
- Haider AW, Larson MG, Franklin SS, Levy D. Systolic blood pressure, diastolic blood pressure, and pulse pressure as predictors of risk for congestive heart failure in the Framingham Heart Study. Ann Intern Med 2003; 138: 10–16
- Halligan SL, Michael T, Wilhelm FH, Clark DM, Ehlers A. Reduced heart rate responding to trauma reliving in trauma survivors with PTSD: Correlates and consequences. J Trauma Stress 2006; 19: 721–734
- Harvey BH, Naciti C, Brand L, Stein DJ. Endocrine, cognitive and hippocampal/cortical 5HT 1A/2A receptor changes evoked by a time-dependent sensitisation (TDS) stress model in rats. Brain Res 2003; 983: 97–107
- Henry JP. The induction of acute and chronic cardiovascular disease in animals by psychosocial stimulation. Int J Psychiatry Med 1975; 6: 147–158
- Hopper JW, Spinazzola J, Simpson WB, Van der Kolk BA. Preliminary evidence of parasympathetic influence on basal heart rate in posttraumatic stress disorder. J Psychosom Res 2006; 60: 83–90
- Hubbard DT, Blanchard DC, Yang M, Markham CM, Gervacio A, Chun I, Blanchard RJ. Development of defensive behavior and conditioning to cat odor in the rat. Physiol Behav 2004; 80: 525–530
- Ivanovich E, Kolarova D. An estimation of the prevalence of risk factors for the cardiovascular system among female telephone operators. Probl Khig 1993; 18: 79–88
- Izzo JL, Levy D, Black HR. Importance of systolic blood pressure in older Americans. Hypertension 2000; 35: 1021–1024
- Karst H, Joels M. Effect of chronic stress on synaptic currents in rat hippocampal dentate gyrus neurons. J Neurophysiol 2003; 89: 625–633
- Kavushansky A, Richter-Levin G. Effects of stress and corticosterone on activity and plasticity in the amygdala. J Neurosci Res 2006; 84: 1580–1587
- Kavushansky A, Vouimba RM, Cohen H, Richter-Levin G. Activity and plasticity in the CA1, the dentate gyrus, and the amygdala following controllable vs. uncontrollable water stress. Hippocampus 2006; 16: 35–42
- Khan S, Liberzon I. Topiramate attenuates exaggerated acoustic startle in an animal model of PTSD. Psychopharmacology (Berl) 2004; 172: 225–229
- Kim JJ, Diamond DM. The stressed hippocampus, synaptic plasticity and lost memories. Nat Rev Neurosci 2002; 3: 453–462
- de Kloet CS, Vermetten E, Geuze E, Kavelaars A, Heijnen CJ, Westenberg HG. Assessment of HPA-axis function in posttraumatic stress disorder: Pharmacological and non-pharmacological challenge tests, a review. J Psychiatr Res 2006; 40: 550–567
- Kohda K, Harada K, Kato K, Hoshino A, Motohashi J, Yamaji T, Morinobu S, Matsuoka N, Kato N. Glucocorticoid receptor activation is involved in producing abnormal phenotypes of single-prolonged stress rats: A putative post-traumatic stress disorder model. Neuroscience 2007; 148: 22–33
- Kolb LC, Mutalipassi LR. The conditioned emotional response: A sub-class of the chronic delayed post-traumatic stress disorder. Psychiatric Ann 1992; 12: 979–987
- Korte SM, De Boer SF. A robust animal model of state anxiety: Fear-potentiated behaviour in the elevated plus-maze. Eur J Pharmacol 2003; 463: 163–175
- Kubzansky LD, Koenen KC, Spiro A, III, Vokonas PS, Sparrow D. Prospective study of posttraumatic stress disorder symptoms and coronary heart disease in the Normative Aging Study. Arch Gen Psychiatry 2007; 64: 109–116
- Liberzon I, Krstov M, Young EA. Stress–restress: Effects on ACTH and fast feedback. Psychoneuroendocrinology 1997; 22: 443–453
- Liu NB, Li H, Liu XQ, Sun CY, Cheng SR, Zhang MH, Liu SC, Wang WX. Chronic multiple stress enhances learning and memory capability in rats. Sheng Li Xue Bao 2004; 56: 615–619
- Li S, Murakami Y, Wang M, Maeda K, Matsumoto K. The effects of chronic valproate and diazepam in a mouse model of posttraumatic stress disorder. Pharmacol Biochem Behav 2006; 85: 324–331
- Luine V. Sex differences in chronic stress effects on memory in rats. Stress 2002; 5: 205–216
- Maercker A, Beauducel A, Schutzwohl M. Trauma severity and initial reactions as precipitating factors for posttraumatic stress symptoms and chronic dissociation in former political prisoners. J Trauma Stress 2000; 13: 651–660
- Magarinos AM, McEwen BS, Flugge G, Fuchs E. Chronic psychosocial stress causes apical dendritic atrophy of hippocampal CA3 pyramidal neurons in subordinate tree shrews. J Neurosci 1996; 16: 3534–3540
- Maier SF, Watkins LR. Stressor controllability and learned helplessness: The roles of the dorsal raphe nucleus, serotonin, and corticotropin-releasing factor. Neurosci Biobehav Rev 2005; 29: 829–841
- Maier SF, Grahn RE, Kalman BA, Sutton LC, Wiertelak EP, Watkins LR. The role of the amygdala and dorsal raphe nucleus in mediating the behavioral consequences of inescapable shock. Behav Neurosci 1993; 107: 377–388
- Malloy PF, Fairbank JA, Keane TM. Validation of a multimethod assessment of posttraumatic stress disorders in Vietnam veterans. J Consult Clin Psychol 1983; 51: 488–494
- Manzanares PAR, Isoardi NA, Carrer HF, Molina VA. Previous stress facilitates fear memory, attenuates GABAergic inhibition, and increases synaptic plasticity in the rat basolateral amygdala. J Neurosci 2005; 25: 8725–8734
- Maroun M, Akirav I. Arousal and stress effects on consolidation and reconsolidation of recognition memory. Neuropsychopharmacology 2007, In Press
- McDougle CJ, Krystal JH, Price LH, Heninger GR, Charney DS. Noradrenergic response to acute ethanol administration in healthy subjects: Comparison with intravenous yohimbine. Psychopharmacology 1995; 118: 127–135
- McEwen BS, Chattarji S. Molecular mechanisms of neuroplasticity and pharmacological implications: The example of tianeptine. Eur Neuropsychopharmacol 2004; 14 Suppl 5: S497–S502
- McFall ME, Murburg MM, Ko GN, Veith RC. Autonomic responses to stress in Vietnam combat veterans with posttraumatic stress disorder. Biol Psychiatry 1990; 27: 1165–1175
- McLaughlin KJ, Gomez JL, Baran SE, Conrad CD. The effects of chronic stress on hippocampal morphology and function: An evaluation of chronic restraint paradigms. Brain Res 2007; 1161: 56–64
- McNally RJ. Implicit and explicit memory for trauma-related information in PTSD. Ann N Y Acad Sci 1997; 821: 219–224
- Milde AM, Sundberg H, Roseth AG, Murison R. Proactive sensitizing effects of acute stress on acoustic startle responses and experimentally induced colitis in rats: Relationship to corticosterone. Stress 2003; 6: 49–57
- Mitra R, Jadhav S, McEwen BS, Vyas A, Chattarji S. Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc Natl Acad Sci USA 2005; 102: 9371–9376
- Moradi AR, Neshat-Doost HT, Teghavi R, Yule W, Dalgleish T. Performance of children of adults with PTSD on the stroop color-naming task: A preliminary study. J Trauma Stress 1999a; 12: 663–671
- Moradi AR, Taghavi MR, MoradiAR AR, Taghavi MR, Neshat Doost HT, Yule W, Dalgleish T. Performance of children and adolescents with PTSD on the Stroop colour-naming task. Psychol Med 1999b; 29: 415–419
- Myers EA, Banihashemi L, Rinaman L. The anxiogenic drug yohimbine activates central viscerosensory circuits in rats. J Comp Neurol 2005; 492: 426–441
- Nader K, Schafe GE, Le Doux JE. Fear memories require protein synthesis in the amygdala for reconsolidation after retrieval. Nature 2000; 406: 722–726
- Nemeroff CB, Bremner JD, Foa EB, Mayberg HS, North CS, Stein MB. Posttraumatic stress disorder: A state-of-the-science review. J Psychiatr Res 2006; 40: 1–21
- Newport DJ, Nemeroff CB. Neurobiology of posttraumatic stress disorder. Curr Opin Neurobiol 2000; 10: 211–218
- Orr SP, Claiborn JM, Altman B, Forgue DF, de Jong JB, Pitman RK, Herz LR. Psychometric profile of posttraumatic stress disorder, anxious, and healthy Vietnam veterans: Correlations with psychophysiologic responses. J Consult Clin Psychol 1990; 58: 329–335
- Orr SP, Lasko NB, Metzger LJ, Berry NJ, Ahern CE, Pitman RK. Psychophysiologic assessment of women with posttraumatic stress disorder resulting from childhood sexual abuse. J Consult Clin Psychol 1998; 66: 906–913
- Orr SP, Metzger LJ, Lasko NB, Macklin ML, Peri T, Pitman RK. De novo conditioning in trauma-exposed individuals with and without posttraumatic stress disorder. J Abnorm Psychol 2000; 109: 290–298
- Park CR, Campbell AM, Diamond DM. Chronic psychosocial stress impairs learning and memory and increases sensitivity to yohimbine in adult rats. Biol Psychiatry 2001; 50: 994–1004
- Park CR, Campbell AM, Woodson JC, Smith TP, Fleshner M, Diamond DM. Permissive influence of stress in the expression of U-shaped relationship between serum corticosterone levels and spatial memory errors in rats. Dose-Response 2006; 4: 55–74
- Parra L, Fuentes JA, Alsasua A. Vascular alterations in rats with high blood pressure induced by social deprivation stress. Life Sci 1994; 55: 669–675
- Paunovic N, Lundh LG, Ost L-G. Attentional memory bias for emotional information in crime victims with acute posttraumatic stress disorder. J Anxiety Disord 2002; 16: 675–692
- Peri T, Ben Shakhar G, Orr SP, Shalev AY. Psychophysiologic assessment of aversive conditioning in posttraumatic stress disorder. Biol Psychiatry 2000; 47: 512–519
- Pitman RK, Orr SP, Forgue DF, de Jong JB, Claiborn JM. Psychophysiologic assessment of posttraumatic stress disorder imagery in Vietnam combat veterans. Arch Gen Psychiatry 1987; 44: 970–975
- Pole N. The psychophysiology of posttraumatic stress disorder: A meta-analysis. Phychol Bull 2007; 133: 725–746
- Przybyslawski J, Sara SJ. Reconsolidation of memory after its reactivation. Behav Brain Res 1997; 84: 241–246
- Przybyslawski J, Roullet P, Sara SJ. Attenuation of emotional and nonemotional memories after their reactivation: Role of beta adrenergic receptors. J Neurosci 1999; 19: 6623–6628
- Pynoos RS, Goenjian A, Tashjian M, Karakashian M, Manjikian R, Manoukian G, Steinberg AM, Fairbanks LA. Post-traumatic stress reactions in children after the 1988 Armenian earthquake. Br J Psychiatry 1993; 163: 239–247
- Pynoos RS, Ritzmann RF, Steinberg AM, Goenjian A, Prisecaru I. A behavioral animal model of posttraumatic stress disorder featuring repeated exposure to situational reminders. Biol Psychiatry 1996; 39: 129–134
- Rabe S, Dorfel D, Zollner T, Maercker A, Karl A. Cardiovascular correlates of motor vehicle accident related posttraumatic stress disorder and its successful treatment. Appl Psychophysiol Biofeedback 2006; 31: 315–330
- Rasmusson AM, Hauger RL, Morgan CA, Bremner JD, Charney DS, Southwick SM. Low baseline and yohimbine-stimulated plasma neuropeptide Y (NPY) levels in combat-related PTSD. Biol Psychiatry 2000; 47: 526–539
- Rau V, DeCola JP, Fanselow MS. Stress-induced enhancement of fear learning: An animal model of posttraumatic stress disorder. Neurosci Biobehav Rev 2005; 29: 1207–1223
- Regehr C, Hill J, Glancy GD. Individual predictors of traumatic reactions in firefighters. J Nerv Ment Dis 2000; 188: 333–339
- Resnick HS, Yehuda R, Pitman RK, Foy DW. Effect of previous trauma on acute plasma cortisol level following rape. Am J Psychiatry 1995; 152: 1675–1677
- Reynolds M, Brewin CR. Intrusive cognitions, coping strategies and emotional responses in depression, post-traumatic stress disorder and a non-clinical population. Behav Res Ther 1998; 36: 135–147
- Reynolds M, Brewin CR. Intrusive memories in depression and posttraumatic stress disorder. Behav Res Ther 1999; 37: 201–215
- Richter-Levin G. Acute and long-term behavioral correlates of underwater trauma—potential relevance to stress and post-stress syndromes. Psychiatry Res 1998; 79: 73–83
- Rostrup M, Westheim A, Kjeldsen SE, Eide I. Cardiovascular reactivity, coronary risk factors, and sympathetic activity in young men. Hypertension 1993; 22: 891–899
- Sack M, Hopper JW, Lamprecht F. Low respiratory sinus arrhythmia and prolonged psychophysiological arousal in posttraumatic stress disorder: Heart rate dynamics and individual differences in arousal regulation. Biol Psychiatry 2004; 55: 284–290
- Sahar T, Shalev AY, Porges SW. Vagal modulation of responses to mental challenge in posttraumatic stress disorder. Biol Psychiatry 2001; 49: 637–643
- Sandi C, Loscertales M, Guaza C. Experience-dependent facilitating effect of corticosterone on spatial memory formation in the water maze. Eur J Neurosci 1997; 9: 637–642
- Sandi C, Merino JJ, Cordero MI, Touyarot K, Venero C. Effects of chronic stress on contextual fear conditioning and the hippocampal expression of the neural cell adhesion molecule, its polysialylation, and L1. Neuroscience 2001; 102: 329–339
- Sandi C, Woodson JC, Haynes VF, Park CR, Touyarot K, Lopez-Fernandez MA, Venero C, Diamond DM. Acute stress-induced impairment of spatial memory is associated with decreased expression of neural cell adhesion molecule in the hippocampus and prefrontal cortex. Biol Psychiatry 2005; 57: 856–864
- Sara SJ. Retrieval and reconsolidation: Toward a neurobiology of remembering. Learn Mem 2000; 7: 73–84
- Sawamura T, Shimizu K, Nibuya M, Wakizono T, Suzuki G, Tsunoda T, Takahashi Y, Nomura S. Effect of paroxetine on a model of posttraumatic stress disorder in rats. Neurosci Lett 2004; 357: 37–40
- Sawchuk CN, Roy-Byrne P, Goldberg J, Manson S, Noonan C, Beals J, Buchwald D. The relationship between post-traumatic stress disorder, depression and cardiovascular disease in an American Indian tribe. Psychol Med 2005; 35: 1785–1794
- Schmahl CG, Elzinga BM, Ebner UW, Simms T, Sanislow C, Vermetten E, McGlashan TH, Bremner JD. Psychophysiological reactivity to traumatic and abandonment scripts in borderline personality and posttraumatic stress disorders: A preliminary report. Psychiatry Res 2004; 126: 33–42
- Servatius RJ, Ottenweller JE, Natelson BH. Delayed startle sensitization distinguishes rats exposed to one or three stress sessions: Further evidence toward an animal model of PTSD. Biol Psychiatry 1995; 38: 539–546
- Shalev AY, Orr SP, Pitman RK. Psychophysiologic assessment of traumatic imagery in Israeli civilian patients with posttraumatic stress disorder. Am J Psychiatry 1993; 150: 620–624
- Shimizu K, Sawamura T, Nibuya M, Nakai K, Takahashi Y, Nomura S. An animal model of posttraumatic stress disorder and its validity: Effect of paroxetine on a PTSD model in rats. Nihon Shinkei Seishin Yakurigaku Zasshi 2004; 24: 283–290
- Shimizu K, Kikuchi A, Wakizono T, Suzuki G, Toda H, Sawamura T, Nibuya M, Takahashi Y, Soichiro N. An animal model of posttraumatic stress disorder in rats using a shuttle box. Nihon Shinkei Seishin Yakurigaku Zasshi 2006; 26: 93–99
- Shors TJ, Seib TB, Levine S, Thompson RF. Inescapable versus escapable shock modulates long-term potentiation in the rat hippocampus. Science 1989; 244: 224–226
- Siegmund A, Wotjak CT. A mouse model of posttraumatic stress disorder that distinguishes between conditioned and sensitised fear. J Psychiatr Res 2007a; 41: 848–860
- Siegmund A, Wotjak CT. Hyperarousal does not depend on trauma-related contextual memory in an animal model of posttraumatic stress disorder. Physiol Behav 2007b; 90: 103–107
- Solomon Z, Mikulincer M, Avitzur E. Coping, locus of control, social support, and combat-related posttraumatic stress disorder: A prospective study. J Pers Soc Psychol 1988; 55: 279–285
- Solomon Z, Mikulincer M, Benbenishty R. Locus of control and combat-related post-traumatic stress disorder: The intervening role of battle intensity, threat appraisal and coping. Br J Clin Psychol 1989; 28: 131–144
- Southwick SM, Krystal JH, Morgan CA, Johnson D, Nagy LM, Nicolaou A, Heninger GR, Charney DS. Abnormal noradrenergic function in posttraumatic stress disorder. Arch Gen Psychiatry 1993; 50: 266–274
- Southwick SM, Bremner JD, Rasmusson A, Morgan CA, III, Arnsten A, Charney DS. Role of norepinephrine in the pathophysiology and treatment of posttraumatic stress disorder. Biol Psychiatry 1999a; 46: 1192–1204
- Southwic SM, Morgan CA, III, Charney DS, High JR. Yohimbine use in a natural setting: Effects on posttraumatic stress disorder. Biol Psychiatry 1999b; 46: 442–444
- Southwick SM, Paige S, Morgan CA, III, Bremner JD, Krystal JH, Charney DS. Neurotransmitter alterations in PTSD: Catecholamines and serotonin. Semin Clin Neuropsychiatry 1999c; 4: 242–248
- Speckens AE, Ehlers A, Hackmann A, Clark DM. Changes in intrusive memories associated with imaginal reliving in posttraumatic stress disorder. J Anxiety Disord 2006; 20: 328–341
- Speckens AE, Ehlers A, Hackmann A, Ruths FA, Clark DM. Intrusive memories and rumination in patients with post-traumatic stress disorder: A phenomenological comparison. Memory 2007; 15: 249–257
- Spence JD. Cerebral consequences of hypertension: Where do they lead?. J Hypertens Suppl 1996; 14: S139–S145
- Stam R. PTSD and stress sensitisation: A tale of brain and body Part 1: Human studies. Neurosci Biobehav Rev 2007a; 31: 530–557
- Stam R. PTSD and stress sensitisation: A tale of brain and body Part 2: Animal models. Neurosci Biobehav Rev 2007b; 31: 558–584
- Strawn JR, Geracioti TD. Noradrenergic dysfunction and the psychopharmacology of posttraumatic stress disorder. Depress Anxiety 2007, In Press
- Sutker PB, Allain AN, Jr., Winstead DK. Psychopathology and psychiatric diagnoses of World War II Pacific theater prisoner of war survivors and combat veterans. Am J Psychiatry 1993; 150: 240–245
- Suzuki A, Josselyn SA, Frankland PW, Masushige S, Silva AJ, Kida S. Memory reconsolidation and extinction have distinct temporal and biochemical signatures. J Neurosci 2004; 24: 4787–4795
- Takahashi T, Morinobu S, Iwamoto Y, Yamawaki S. Effect of paroxetine on enhanced contextual fear induced by single prolonged stress in rats. Psychopharmacology (Berl) 2006; 189: 165–173
- Taylor F, Cahill L. Propranolol for reemergent posttraumatic stress disorder following an event of retraumatization: A case study. J Trauma Stress 2002; 15: 433–437
- Trofimiuk E, Walesiuk A, Braszko JJ. St John's wort (Hypericum perforatum) diminishes cognitive impairment caused by the chronic restraint stress in rats. Pharmacol Res 2005; 51: 239–246
- Ullman SE, Filipas HH. Predictors of PTSD symptom severity and social reactions in sexual assault victims. J Trauma Stress 2001; 14: 369–389
- Valles A, Marti O, Garcia A, Armario A. Single exposure to stressors causes long-lasting, stress-dependent reduction of food intake in rats. Am J Physiol Regul Integr Comp Physiol 2000; 279: R1138–R1144
- Veazey CH, Blanchard EB, Hickling EJ, Buckley TC. Physiological responsiveness of motor vehicle accident survivors with chronic posttraumatic stress disorder. Appl Psychophysiol Biofeedback 2004; 29: 51–62
- Vouimba RM, Yaniv D, Diamond D, Richter-Levin G. Effects of inescapable stress on LTP in the amygdala versus the dentate gyrus of freely behaving rats. Eur J Neurosci 2004; 19: 1887–1894
- Vouimba RM, Munoz C, Diamond DM. Differential effects of predator stress and the antidepressant tianeptine on physiological plasticity in the hippocampus and basolateral amygdala. Stress 2006; 9: 29–40
- Vyas A, Mitra R, Shankaranarayana Rao BS, Chattarji S. Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons. J Neurosci 2002; 22: 6810–6818
- Vyas A, Bernal S, Chattarji S. Effects of chronic stress on dendritic arborization in the central and extended amygdala. Brain Res 2003; 965: 290–294
- Vyas A, Jadhav S, Chattarji S. Prolonged behavioral stress enhances synaptic connectivity in the basolateral amygdala. Neuroscience 2006; 143: 387–393
- Vythilingam M, Anderson GM, Owens MJ, Halaszynski TM, Bremner JD, Carpenter LL, Heninger GR, Nemeroff CB, Charney DS. Cerebrospinal fluid corticotropin-releasing hormone in healthy humans: Effects of yohimbine and naloxone. J Clin Endocrinol Metab 2000; 85: 4138–4145
- Wakizono T, Sawamura T, Shimizu K, Nibuya M, Suzuki G, Toda H, Hirano J, Kikuchi A, Takahashi Y, Nomura S. Stress vulnerabilities in an animal model of post-traumatic stress disorder. Physiol Behav 2007; 90: 687–695
- Walesiuk A, Trofimiuk E, Braszko JJ. Gingko biloba extract diminishes stress-induced memory deficits in rats. Pharmacol Rep 2005; 57: 176–187
- Watanabe Y, Gould E, Cameron HA, Daniels DC, McEwen BS. Phenytoin prevents stress- and corticosterone-induced atrophy of CA3 pyramidal neurons. Hippocampus 1992a; 2: 431–435
- Watanabe Y, Gould E, Daniels DC, Cameron H, McEwen BS. Tianeptine attenuates stress-induced morphological changes in the hippocampus. Eur J Pharm 1992b; 222: 157–162
- Watanabe Y, Gould E, McEwen BS. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res 1992c; 588: 341–345
- Woodson JC, Macintosh D, Fleshner M, Diamond DM. Emotion-induced amnesia in rats: Working memory-specific impairment, corticosterone–memory correlation, and fear versus arousal effects on memory. Learn Mem 2003; 10: 326–336
- Wright RL, Conrad CD. Chronic stress leaves novelty-seeking behavior intact while impairing spatial recognition memory in the Y-maze. Stress 2005; 8: 151–154
- Wright RL, Conrad CD. Enriched environment prevents chronic stress-induced spatial learning and memory deficits. Behav Brain Res 2007, in press
- Yehuda R. Neuroendocrine aspects of PTSD. Handbook Exp Pharmacol 2005; 371–403
- Yehuda R, Golier JA, Tischler L, Stavitsky K, Harvey PD. Learning and memory in aging combat veterans with PTSD. J Clin Exp Neuropsychol 2005; 27: 504–515
- Yehuda R, Tischler L, Golier JA, Grossman R, Brand SR, Kaufman S, Harvey PD. Longitudinal assessment of cognitive performance in Holocaust survivors with and without PTSD. Biol Psychiatry 2006; 60: 714–721
- Zoladz PR, Campbell AM, Park CR, Schaefer D, Danysz W, Diamond DM. Enhancement of long-term spatial memory in adult rats by the noncompetitive NMDA receptor antagonists, memantine and neramexane. Pharmacol Biochem Behav 2006; 85: 298–306