
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.1. Introduction
During running, a major source of energy loss is the redirection of the center of mass (COM) motion from downward to upward during stance. This down-to-up redirection could, as a first approximation, be considered as a collision with the ground (Ruina et al., Citation2005). A collision occurs when a perpendicular relationship between force and velocity vector is not maintained; the angle of deviation from this relationship is called by Ruina et al. (Citation2005) the collision angle (ϕ).
These authors showed that the energy losses could be minimized via suitable path matching of the leg as it collides with the ground. During the first part of the stance phase (absorptive part), ϕ is negative and energy is absorbed by extending muscles and tendons as well as by deformation of other tissues (Fig. A). During the second part (generative part), ϕ is positive and energy is generated by muscles and tendon recoil (Fig. B). When the collision angle of the absorptive deceleration phase mirrors the subsequent collision angle of the generative acceleration phase, the collision is kinematically equivalent to a ‘pseudo-elastic collision’, which minimizes the collisional energy losses, reducing the muscular work.
Figure 1 Deviation of F from the vertical (θ), of V from the horizontal (λ) and collision angle (ϕ) over the absorptive (A) and generative (B) phases.
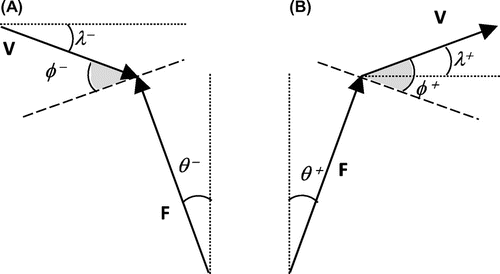
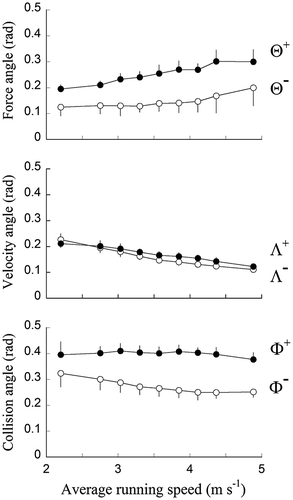
Figure 2 Average deviation of F from the vertical (Θ), average deviation of V from the horizontal (Λ) and average collision angle (ϕ), during the absorptive (white symbols) and generative (black symbols) phases.
Currently, collision-based analyses of human gait are done assuming that the COM within a step is symmetric around the mid-step (Lee et al., 2013). However, differences in mechanical events taking place during the descent and the lift of the center of mass occur during running (Cavagna, Citation2006). This deviation from a symmetrical pattern, defined by Cavagna as the ‘landing-takeoff asymmetry’, results from a distinctive response of the muscle-tendons units during the absorptive and generative phases because (i) according to the force-velocity relation, muscular force is greater during stretching than during shortening and (ii) a dissimilarity exists in the lever system (Cavagna et al., 2010). According to this author, the ‘landing-takeoff asymmetry, which would be nil in the elastic rebound of the symmetric spring-mass model for running, suggests a less efficient elastic energy storage and recovery during the bouncing step’.
We hypothesize that, due to landing-takeoff asymmetry, the average collision angle is different during the absorptive and generative part of stance, reflecting a deviation from a pseudo-elastic behavior. Therefore, we have measured the collision parameters during running on the level at different speeds.
2. Methods
Ten healthy recreational runners (3 ♀-7 ♂, age: 22–49 years; weight: 52–83 kg; height: 1.64–1.87 m) participated to the study. Informed consent was obtained. The experimental procedure was approved by the Ethic Committee of the Université catholique de Louvain.
Subjects ran on a treadmill (h/p/Comos-Stellar, Germany) at nine different speeds ranging from 8 to 18 km h−1. The treadmill was instrumented with four force transducers (Arsalis, Belgium) that measured the three components of force vector (F) exerted by the tread-belt under the foot. The velocity vector (V) of the COM was computed from F using a method similar to the one described in Dewolf et al. (Citation2016). The stance phase was defined as the period during which the vertical force was greater than 10 N.
The collision of the leg with the ground was assessed at each instant from the following variables (Fig.): the instantaneous force angle θ was the deviation of F from vertical, the instantaneous velocity angle λ was the deviation of V from horizontal and the instantaneous collision angle ϕ was the deviation from an orthogonal relationship of the angle between F and V. The stance phase was divided in two parts: the absorptive part during which ϕ was negative and the generative part during which ϕ was positive.
Then, the absolute value of the force angle θ, the velocity angle λ and collision angle ϕ were averaged over the absorptive part (respectively ) and over the generative part of the ground contact (respectively
).
3. Results and discussion
During the absorptive part of stance, Θ– increases slightly with speed (F8,89 = 2.94, p = 0.006). More importantly, Λ– decreases when speed increases (F8,89 = 49.15, p < 0.001). As a result, Φ– slightly decreases with increasing speed (F8,89 = 5.44, p < 0.001). During the generative part of stance, Θ+ increases (F8,89 = 14.92, p < 0.001) while Λ+ decreases (F8,89 = 21.77, p < 0.001) when speed increases. As a result, Φ+ is about constant at all speeds (F8,89 = 1.07, p = 0.386).
At all speeds, Φ– < Φ+ showing that the angle between V and F is closer to 90° at the beginning than at the end of the stance. This difference is mainly due to a difference in θ during the absorptive and generative phase. As a consequence, the stance phase deviates from a pseudo-elastic collision. Considering that the energy changes are mostly accounted for by changes in the component of the velocity normal to the leg (Ruina et al. Citation2005), one can expect that the energy production by muscles during the generative phase exceeds the energy dissipation during the absorptive phase. This last result confirm those of DeVita et al. (Citation2008), showing that during running on the level at a constant average speed, the positive muscle work done is greater than the negative work absorbed, despite the fact that the average total mechanical energy remains constant. These observations suggest that muscles generate a net positive amount of work to overcome various energy losses by other tissues (Zelik & Kuo, 2012).
As speed increases, the difference between Φ– and Φ+ increases and the collision becomes more and more generative. As the collision wit the ground becomes greater at faster speeds, the work dissipated by the soft tissues increases and consequently a greater amount of net positive work is done by muscle-tendons units (Zelik & Kuo, 2012).
4. Conclusions
Analysis of collisional energy losses could be used as a complementary approach to capture mechanical dynamics and could offer new insight into the bouncing mechanism of running.
Acknowledgements
The authors want to thank Dr. L. Peñailillo for his help during the experiments. This study was funded by the Université catholique de Louvain.
References
- Cavagna GA. 2006. The landing-take-off assymetry in human running. J Exp Biol. 209:4051–4060.10.1242/jeb.02344
- Cavagna GA. 2010. Symmetry and asymmetry in boucing gaits. Symmetry. 2:1270–1321.10.3390/sym2031270
- DeVita P, Janshen L, Rider P, Solnik S, Hortobagyi T. 2008. Muscle work is biased toward energy generation over dissipation in non-level running. J Biomech. 41:3354–3359.10.1016/j.jbiomech.2008.09.024
- Dewolf AH, Penailillo LE, Willems PA. 2016. The rebound of the body during uphill and downhill running at different speeds. J Exp Biol. 219:2276–2288.
- Lee DV, Comanescu TN, Butcher MT, Bertram JEA. 2016. A comparative collision-based analysis of human gait. Proc Biol Sci. 280:20131779.
- Ruina A, Bertram JEA, Srinivasan M. 2005. A collisional model of the energetic cost of support work qualitatively explains leg sequencing in walking and galloping, pseudo-elastic leg behavior in running and the walk-to- run transition. J Theor Biol. 237:170.10.1016/j.jtbi.2005.04.004
- Zelik KE, Kuo AD. 2010. Human walking isn't all hard work: evidence of soft tissue contributions to energy dissipation and return. J Exp Biol. 213:4257–4264.10.1242/jeb.044297