
Abstract
Near-shore physical habitat is crucial for supporting biota and ecological processes in lakes. We used data from a statistical sample of 1101 lakes and reservoirs from the 2007 US Environmental Protection Agency National Lakes Assessment (NLA) to develop 4 indices of physical habitat condition: (1) Lakeshore Anthropogenic Disturbance, (2) Riparian Vegetation Cover Complexity, (3) Littoral Cover Complexity, and (4) Littoral–Riparian Habitat Complexity. We compared lake index values with distributions from least-disturbed lakes. Our results form the first US national assessment of lake physical habitat, inferring the condition (with known confidence) of the 49,500 lakes and reservoirs in the conterminous United States with surface areas >4 ha and depths >1 m. Among the physical and chemical characteristics examined by the NLA, near-shore physical habitat was the most extensively altered relative to least-disturbed condition. Riparian Vegetation Cover Complexity was good in 46% (±3%), fair in 18% (±2%), and poor in 36% (±2%) of lakes. Littoral–Riparian Habitat Complexity was good in 47% (±3%), fair in 20% (±2%), and poor in 32% (±2%) of lakes. In every ecoregion, habitat condition was negatively related to anthropogenic pressures. Gradients of increased anthropogenic disturbance were accompanied by progressive simplification of littoral and riparian physical habitat. Nationwide, the proportion of lakes with degraded near-shore physical habitat was equal to or greater than that for excess nutrients and much greater than that for acidic conditions. In addition to better management of lake basins, we suggest an increased focus on protecting littoral and riparian physical habitat through better management of near-shore anthropogenic disturbance.
Near-shore physical habitat structure has only recently been addressed in regional lake monitoring, even though anthropogenic activities and aquatic and riparian biota are concentrated in these areas. Monitoring and assessment of near-shore lake habitat conditions are needed because these areas are both ecologically important and especially vulnerable to anthropogenic perturbation (Schindler and Scheuerell Citation2002, Strayer and Findlay Citation2010, Hampton et al. Citation2011). Littoral and riparian zones are positioned at the land–water interface and tend to be more structurally complex and biologically diverse than either pelagic areas or upland terrestrial environments (Polis et al. Citation1997, Strayer and Findlay Citation2010). This complexity promotes interchange of water, nutrients, and biota between the aquatic and terrestrial compartments of lake ecosystems (Benson and Magnuson Citation1992, Polis et al. Citation1997, Palmer et al. Citation2000, Zohary and Ostrovsky Citation2011).
Riparian vegetation and wetlands surrounding lakes increase near-shore physical habitat complexity (e.g., Christensen et al. Citation1996, Francis and Schindler Citation2006) and buffer lakes from the influence of upland land use activities (Carpenter and Cottingham Citation1997, Strayer and Findlay Citation2010). Anthropogenic activities on or near lakeshores can directly or indirectly degrade littoral and riparian habitat (Francis and Schindler Citation2006). Increased sedimentation, loss of native plant growth, alteration of native plant communities, loss of physical habitat structure, and changes in littoral cover and substrate are all commonly associated with lakeshore human activities (Christensen et al. Citation1996, Engel and Pederson Citation1998, Whittier et al. Citation2002, Francis and Schindler Citation2006, Merrell et al. Citation2009). Such reductions in physical habitat structural complexity can deleteriously affect fish (Wagner et al. Citation2006, Taillon and Fox Citation2004, Whittier et al. Citation1997, 2002, Halliwell Citation2007, Jennings et al. Citation1999, Wagner et al. Citation2006), aquatic macroinvertebrates (Brauns et al. Citation2007), and birds (Kaufmann et al. Citation2014b).
Despite the growing recognition that protection of near-shore physical habitat structural complexity is essential to conserve biological condition in lakes, the lack of practical, standardized approaches has hindered its quantification and evaluation in large-scale regional and national lake assessments. We therefore developed rapid field methods to quantify physical habitat structure and near-shore anthropogenic disturbances (Kaufmann and Whittier Citation1997, USEPA Citation2007a) for application in national surveys. Based on data from these field methods, we calculated habitat indices that quantify the variety, structural complexity, and magnitude of areal cover from physical habitat elements within the near shore zones of lakes. Similar indices have been applied both in lakes (Kaufmann et al. Citation2014a) and streams (Hughes et al. Citation2010, Kaufmann and Faustini Citation2012). The structural complexity and variety of habitat quantified by these indices provide diverse opportunities for supporting assemblages of aquatic organisms (Strayer and Finlay Citation2010, Kovalenko et al. Citation2012) and foster the interchange of water, nutrients, and organisms between compartments within lake ecosystems (Schindler and Scheuerell Citation2002, Stayer and Findlay Citation2010).
We used the methods described herein to assess physical habitat condition, interpret biological information, and assess lake ecological condition in the US Environmental Protection Agency's National Lakes Assessment (NLA; USEPA Citation2009). This evaluation of near-shore physical habitat condition was an unprecedented effort at the national scale.
Our objectives in this article are to describe how we evaluated near-shore physical habitat data in the NLA and associated 4 key physical habitat condition indices with anthropogenic disturbances. We summarize key findings concerning the extent of lake physical habitat alteration across the United States and discuss the potential risks of physical habitat alteration to lake biota. This paper also describes how to calculate the NLA's physical habitat indices and derive physical habitat condition thresholds relative to least-disturbed conditions.
Methods
Analytical framework for assessing physical habitat condition
We took the following 7 steps to assess physical habitat condition in US lakes at national and regional levels:
surveyed littoral and riparian physical habitat structure and near-shore anthropogenic activities on a national probability sample of lakes and reservoirs;
defined ecoregions and classified lakes according to lake type and level of anthropogenic disturbance;
calculated a set of physical habitat metrics;
calculated multimetric indices of physical habitat condition;
estimated region-specific expected values for physical habitat condition indices;
set criteria for expected physical habitat condition based on least-disturbed lakes; and
estimated percentages and confidence bounds for the number of lakes with physical habitat in good, fair, and poor condition in the United States and its ecoregions.
National survey of lakes (Step 1)
Study area and site selection
The NLA field sampling effort targeted all lakes and reservoirs in the conterminous United States with surface areas >4 ha and depths >1 m. Field crews visited 1152 lakes and reservoirs () between May and October 2007. Of these, 1028 had been selected as a probability sample from the US Geological Survey (USGS)-USEPA National Hydrography Dataset (NHD) with a spatially balanced, randomized systematic design that excluded the Great Lakes and Great Salt Lake (Peck et al. Citation2013). The remaining 124 lakes were hand-selected to increase the number of lakes in least-disturbed condition, which were used to estimate potential condition and evaluate response of the indices to disturbance (following Stoddard et al. Citation2006).
Physical habitat data were collected from 1101 of the 1152 survey lakes that had surface areas <5000 ha (981 probability- and 120 hand-selected lakes; Peck et al. Citation2013). Both probability and hand-selected lakes were used to develop physical habitat condition indices and the distribution of expected values. A random subset of 90 probability lakes was visited twice during the sampling period to estimate the precision of the habitat measurements and indices (Kaufmann et al. Citation2014a.) Depending on the index, 965–973 of the 981 probability sample lakes had the requisite data for making statistical inferences concerning habitat condition in the national population of ∼49,500 lakes and reservoirs (Peck et al. Citation2013).
Field sampling design and methods
Our lake physical habitat field methods (USEPA Citation2007a, Kaufmann et al. Citation2014a) produced information concerning 7 dimensions of near-shore physical habitat: (1) water depth and surface characteristics, (2) substrate size and type, (3) aquatic macrophyte cover and structure, (4) littoral cover for biota, (5) riparian vegetation cover and structure, (6) near-shore anthropogenic disturbances, and (7) bank characteristics that indicate lake level fluctuations and terrestrial–aquatic interactions. At each lake, field crews characterized these 7 components of near-shore physical habitat at 10 equidistant stations along the shoreline. Each station included a littoral plot (10 × 15 m) and an adjoining riparian plot (15 × 15 m). Littoral depth was measured 10 m off-shore at each station. See Kaufmann et al. (Citation2014a) for a description of field methods, our approach for calculating whole-lake physical habitat metrics, and an assessment of habitat metric precision.
Classifications (Step 2)
Ecoregions
We report findings nationally and by 9 aggregated Omernik (Citation1987) level III ecoregions (Paulsen et al. Citation2008): the Northern Appalachians (NAP), Southern Appalachians (SAP), Coastal Plains (CPL), Upper Midwest (UMW), Temperate Plains (TPL), Northern Plains (NPL), Southern Plains (SPL), Western Mountains (WMT), and Xeric West (XER; ). We used ecoregions as a first-level classification for defining and evaluating near-shore riparian condition indicators (RVegQ and its variants) because ecoregions are useful predictors of landform, climate, and potential natural vegetation (Omernik Citation1987, Paulsen et al. Citation2008). For some aspects of habitat index development, we grouped ecoregions into broader ecoregions: the Eastern Highlands (EHIGH = NAP + SAP), the Plains and Lowlands (PLNLOW = CPL + UMW + TPL + NPL + SPL), Central Plains (CENPL = TPL + NPL + SPL), and the West (WMT + XER).
Lake types
Littoral habitat elements (e.g., aquatic macrophytes, snags, and substrate) are affected by lithology and water depth, clarity, temperature, and ionic strength, factors that were poorly associated with the 9 ecoregion classification. We therefore used a geographically constrained multivariate classification described by USEPA (appendix A in USEPA 2009) that included variables known to influence those littoral habitat elements to group lakes with similar expectations for littoral cover. Within each of the 3 broader ecoregions, lakes were grouped by their similarity in size, depth, morphology, elevation, temperature, precipitation, calcium concentration, latitude, and longitude.
A nonhierarchical cluster analysis (FASTCLUS; SAS Institute Citation2008) of log10-transformed, standardized data within each of the 3 broad ecoregions yielded a useful grouping of 7 lake types (; ). Lake-type classification and type-specific index formulations reduced the among-region variation in index values in least-disturbed lakes and reduced ambiguity in the response to anthropogenic disturbances.
Table 1 Multivariate classification of lakes within 3 major ecoregions used in deriving expectations for littoral habitat characteristics in least-disturbed lakes of the NLA. The aggregated Omernik ecoregions are shown in A; multivariate cluster classification of sample lakes is shown in .
Anthropogenic disturbance and least-disturbed reference site screening
We examined water chemistry, near-shore anthropogenic influences, and evidence of anthropogenic lake drawdown in all 1101 survey lakes to classify them according to their level of anthropogenic disturbance (low, medium, high). Lakes meeting low-disturbance screening criteria served as least-disturbed reference sites for best-available condition. Low-disturbance stress (least-disturbed) lakes within each lake type (A–G in ) were identified on the basis of chemical variables (total phosphorus, total nitrogen, chloride, sulfate, acid neutralizing capacity, dissolved organic carbon, and dissolved oxygen in the epilimnion) and direct observations of anthropogenic disturbances along the lake margin (proportion of lakeshore with nonagricultural influences, proportion of lakeshore with agricultural influences, and the relative extent and intensity of anthropogenic influences of all types together).
For each lake type, a threshold value representing least-disturbed conditions was established as a “pass/fail” criterion for each parameter (Table A1 in USEPA Citation2010). Thresholds were values that would unlikely be found in least-disturbed lakes within each region, and the threshold values varied by lake type to account for regional variations in water chemistry and littoral–riparian anthropogenic activities (Herlihy et al. Citation2013). A lake was considered least-disturbed if it passed the screening test for all parameters; 160 potential least-disturbed reference lakes were identified. We subsequently excluded 11 lakes with large lake level fluctuations (>10 m horizontal) evident from field measurements (Kaufmann et al. Citation2014a), along with evidence of dams or lake level controls in field observations or aerial photos.
Lakes that were not classified as least-disturbed were provisionally considered intermediate in disturbance. The intermediate disturbance lakes were then screened with a set of high-disturbance thresholds applied to the same variables (Table A1 in USEPA Citation2010). Lakes that exceeded one or more of the high disturbance thresholds were considered highly disturbed. To avoid circularity in defining physical habitat alteration, we did not use any of the physical habitat cover complexity indices or their subcomponent metrics in defining lake disturbance classes.
Our screening process identified 149 least-disturbed lakes, 775 intermediate lakes, and 177 highly disturbed lakes. Of the 149 least-disturbed lakes, 107 were in the UMW, WMT, and NAP aggregated ecoregions (). Finding least-disturbed lakes that met our screening criteria was difficult in some other ecoregions; only 1, 6, 5, and 3 least-disturbed lakes, respectively, were indentified in the NPL, SPL, TPL, and XER ecoregions. To increase the useable sample size for estimating expected lake condition, we grouped least-disturbed lakes from the NPL, SPL, TPL into the Central Plains (CENPL), and the WMT and XER into the West. Due to insufficient numbers of least-disturbed lakes relative to the large amount of lake variability within ecoregions, we were unable to use totally independent subsets of lakes for developing and validating models of expected condition.
Table 2 Number of NLA sample lakes classified by anthropogenic disturbance stress as least-disturbed, intermediate, and most-disturbed, based on region-specific low- and high-disturbance screens. Parentheses show subtotals for the Central Plains and Western US regions formed by combining 2 or more of the 9 NLA aggregated ecoregions.
Calculation of lake physical habitat metrics (Step 3)
Near-shore disturbance metrics
Our variable names are from the publicly available NLA 2007 dataset released by the USEPA (http://water.epa.gov/type/lakes/NLA_data.cfm). We calculated extent of shoreline disturbance around the lakeshore (hifpAnyCirca) as the proportion of stations at which crews recorded the presence of at least one of the 12 anthropogenic disturbance types as described by Kaufmann et al. (Citation2014a). We calculated the disturbance intensity metric hiiAll as the sum of the 12 separate proximity-weighted means for all shoreline disturbance types observed at the 10 shoreline stations (Kaufmann et al. Citation2014a).
We also calculated subsets of total disturbance intensity by summing metrics for defined groups of disturbance types. For example, hiiAg sums the proximity-weighted presence metrics for row crop, orchard, and pasture; hiiNonAg sums the proximity-weighted presence metrics for the remaining 9 nonagricultural disturbance metrics: (1) buildings; (2) commercial developments; (3) parks or manmade beaches; (4) docks or boats; (5) seawalls, dikes, or revetments; (6) trash or landfill; (7) roads or railroads; (8) power lines; and (9) lawns.
Riparian vegetation metrics
To characterize riparian vegetation, we converted field observations of cover data to mean cover estimates for all the types and combinations of vegetation data (Kaufmann et al. Citation2014a). We assigned cover class arithmetic midpoint values (i.e., absent = 0%, sparse = 5%, moderate = 25%, heavy = 57.5%, and very heavy = 87.5%) to each plot's cover-class observations and then calculated lakeshore vegetation cover as the average of those cover values across all 10 plots. Metrics for combined cover types (e.g., sum of woody vegetation in 3 layers) were calculated by summing means for the single types (see Kaufmann et al. Citation1999, Citation2014a). Metrics describing the proportion of each lakeshore with presence (rather than cover) of particular features were calculated as the mean of presence (0 or 1) over the 10 riparian plots.
Littoral cover metrics
Metrics describing the mean cover (and mean presence) of littoral physical habitat features and aquatic macrophytes were calculated from observations at the 10 littoral plots as described earlier for riparian vegetation. Metrics for combined cover types (e.g., sum of natural types of fish cover) were calculated by summing means for single types. Cover-weighted geometric mean substrate diameter indices and their standard deviations were calculated for the littoral zone and separately for the 1 m band of exposed shoreline by first assigning the geometric mean between the upper and lower diameter bounds of each size class for each cover observation, weighting them by their mean cover across the 10 stations, and then averaging the weighted cover and computing its variance across size classes (Kaufmann et al. Citation2014a). This approach is an adaptation of that used for estimating geometric mean diameter from pebble count size-class percentages in streams (Faustini and Kaufmann Citation2007, Kaufmann et al. Citation2009).
Calculation of summary physical habitat condition indices (Step 4)
We calculated 4 multimetric indices of physical habitat condition and an integrated 5th index that combines the first 3:
RDis_IX: Lakeshore Anthropogenic Disturbance Index (intensity and extent);
RVegQ: Riparian Vegetation Cover Complexity Index;
LitCvrQ: Littoral Cover Complexity Index;
LitRipCvQ: Littoral–Riparian Habitat Complexity Index; and
LkShoreHQ: Lakeshore Physical Habitat Quality Index.
These indices were defined and calculated independent of their association with the lake classification by anthropogenic disturbance levels and the variables used to produce those classifications.
Lakeshore Anthropogenic Disturbance Index (RDis_IX)
This index was calculated as:
(1)
where disturbance intensity was represented by separate sums of the mean proximity-weighted tallies of near-shore agricultural and nonagricultural disturbance types, and extent was expressed as the proportion of the shore with presence of any type of disturbance. Specifically
(2) where:
hiiNonAg = proximity-weighted mean disturbance tally (mean among stations) of up to 9 types of nonagricultural activities;
hiiAg = proximity-weighted mean tally of up to 3 types of agriculture-related activities (mean among stations); and
hifpAnyCirca = proportion of the 10 shoreline stations with at least 1 of the 12 types of anthropogenic activities present within their 15 × 15 m riparian or 10 × 15 m littoral plots.
Table 3 Assignment of Riparian Vegetation Cover Complexity, Littoral Cover Complexity, and Littoral–Riparian Habitat Complexity index variants by aggregated ecoregion () and multivariate lake type (; ).
Field procedures classified only 3 types of agricultural disturbances, versus 9 types of nonagricultural disturbances, limiting the potential ranges to 0–3 for hiiAg and 0–9 for hiiNonAg. The observed ranges of these variables also differed; hiiAg ranged from 0 to 1.3, whereas hiiNonAg had an observed range of 0 to 6.4. To avoid under-representing agricultural disturbances and over-representing nonagricultural disturbances in the index, we weighted the disturbance intensity tallies for agricultural land use by a factor of 5 in Equationequation 2(2) . This weighting factor (ratio of observed ranges in nonagricultural to agricultural disturbance types) effectively scales agricultural land uses equal in disturbance potential to those for nonagricultural land uses. We scaled the final index from 0 to 1, where 0 indicates absence of any anthropogenic disturbances and 1 is the theoretical maximum approached as a limit at extremely high disturbance. We applied a single formulation of the disturbance index RDis_IX to all ecoregions and lake types in the NLA (; ).
Riparian Vegetation Cover Complexity Index (RVegQ)
This index is based on visual estimates of vegetation cover and structure in 3 vegetation layers at the 10 riparian plots along the lake shore. Because the potential vegetation cover differs among regions, we calculated 3 variants of the Riparian Vegetation Cover Complexity Index (RVegQ_2, RVegQ_7, or RVegQ_8) for application to different aggregated ecoregions (). The region-specific formulations reduce the among-region variation in index values in least-disturbed lakes and reduce ambiguity in their response to anthropogenic disturbances. If component metrics had potential maximum values >1, their ranges were scaled to range from 0 to 1 by dividing by their respective maximum values based on the 2007 NLA data ( in Kaufmann et al. Citation2014a).
Each variant of the final index was calculated as the mean of its component metric values. Index values range from 0 (indicating no vegetative cover at any station) to 1 (40–100% cover in multiple layers at all stations):
(3)
(4)
(5)
where:
rviWoody = sum of the mean areal cover of woody vegetation in 3 layers: canopy (large and small diameter trees), understory, and ground layers (rvfcCanBig + rvfcCanSmall + rvfcUndWoody + rvfcGndWoody);
rviLowWood = sum of mean areal cover of woody vegetation in the understory and ground cover layers (rvfcUndWoody + rvfcGndWoody);
rvfcGndInundated = mean areal cover of inundated terrestrial or wetland vegetation in the ground cover layer;
rvfpCanBig = proportion of stations with large diameter (>0.3 m dbh) trees present; and
ssiNATBedBld = sum of mean areal cover of naturally occurring bedrock and boulders (ssfcBedrock + sfcBoulders), and where the value of ssiNATBedBld was set to 0 in lakes that have a substantial extent of constructed seawalls and revetments (i.e., hipwWalls ≥ 0.10).
We used RVegQ_2 for mesic ecoregions with maximum elevations <2000 m (NAP, SAP, UMW, and CPL) where tree vegetation can be expected in relatively undisturbed locations (). RVegQ_2 sums the woody cover in 3 lakeside vegetation layers (rviWoody) and includes inundated groundcover vegetation (rvfcGndInundated) as a positive characteristic.
We used RVegQ_7 for Central Plains ecoregions (NPL, SPL, and TPL). Whereas perennial woody groundcover and shrubs can be expected on undisturbed lake shorelines throughout the Central Plains (West and Ruark Citation2004), the presence or absence of large trees (>5 m high) along lake margins in this region has ambiguous meaning without floristic information (Barker and Whitman Citation1988, Johnson Citation2002, Huddle et al. Citation2011). RVegQ_7 accommodates lack of tree canopy in least-disturbed lakes by summing only the lower 2 layers of woody vegetation (rviLowWood) and includes inundated ground cover vegetation as a positive characteristic.
We used RVegQ_8 for the West (WMT and XER), where climate ranges from wet to arid and where lakeshores can potentially grow large diameter riparian trees but may lack vegetated lake shorelines at high elevations, or where rock precludes vegetation (). RVegQ_8 sums the woody cover in 3 lakeside vegetation layers and includes inundated groundcover vegetation as a positive characteristic; it also includes the proportional presence of large diameter trees around the lakeshore as a positive characteristic. RVegQ_8 includes natural rock as an undisturbed riparian cover type to avoid penalizing relatively undisturbed lakes in arid areas
or at high elevations above timberline. For lakes with a substantial extent or abundance of constructed seawalls, dikes, or revetments along the shoreline, the substrate metric was set at 0.
Littoral Cover Complexity Index (LitCvrQ)
This index was based on the station averages for visual estimates of the areal cover of 10 types of littoral features, including aquatic macrophytes but excluding manmade structures, within each of the 10 littoral plots (see Kaufmann et al. Citation2014a). We calculated 3 variants but applied these to different lake types rather than to ecoregions (). Each variant of the index was calculated as the mean of its component metric scores so that index values range from 0 (no cover present at any station) to 1 (very heavy cover at all 10 stations). Component metrics with potential maximum values >1 within a given lake type were scaled from 0 to 1 by dividing by their respective maximum values in the NLA 2007 dataset:
(6)
(7)
(8) where:
fciNatural = summed areal cover of nonanthropogenic fish cover elements (fcfcBoulders + fcfcBrush + fcfcLedges + fcfcLivetrees + fcfcOverhang + fcfcSnag + fcfcAquatic);
SomeNatCvr = summed cover of natural fish cover elements excluding snags and aquatic macrophytes (fcfcBoulders + fcfcBrush + fcfcLedges + fcfcLivetrees + fcfcOverhang);
amfcFltEmg = summed cover of emergent plus floating aquatic macrophytes (amfcEmergent + amfcFloating); and
fcfcAquatic = total cover of aquatic macrophytes of any type.
All 3 variants of LitCvrQ include an expression of the summed cover of naturally occurring fish or macroinvertebrate cover elements. Snag cover is recognized as a particularly important element of littoral habitat complexity (Christensen et al. Citation1996, Francis and Schindler Citation2006, Miranda et al. Citation2010); therefore, we included snags as a separate contributing cover component in all 3 variants of the index and divided cover metrics by their maximum values in the 2007 NLA data to make the weightings of snag cover equal to those of the other 2 littoral cover sums. For LitCvrQ_c and LitCvrQ_d, we increased the emphasis on emergent and floating-leaf aquatic macrophytes relative to other littoral components in response to their reported importance as cover and their sensitivity to anthropogenic disturbances in many lake types and regions (Radomski and Geoman Citation2001, Jennings et al. Citation2003, Merrell et al. Citation2009, Beck et al. Citation2013).
We used LitCvrQ_b for lake type D (generally shallow, warm, low conductivity lakes that are almost exclusively in the CPL, but also found in the SPL and TPL; ; and ). We used LitCvrQ_c for lake type A, which consists primarily of reservoirs in the SAP ( and ), where disturbed sites commonly have substantial upland soil erosion, large water level fluctuations, and bare-soil shorelines. These conditions generate abiotic turbidity that suppresses submerged macrophytes, thereby diminishing their common association with anthropogenic nutrient inputs seen in other lake types. LitCvrQ_c emphasizes floating and emergent aquatic macrophytes in addition to snags but still includes submerged aquatic macrophytes along with other aquatic macrophytes and cover types in fciNatural. LitCvrQ_d excludes submerged aquatic macrophytes. We applied LitCvrQ_d in all lake types except A and D in the NLA (), where submerged aquatic macrophytes provide valuable cover but high submerged cover is frequently associated with anthropogenic eutrophication (Hatzenbeler et al. Citation2004, Merrell et al. Citation2009).
Littoral–Riparian Habitat Complexity Index (LitRipCvQ)
We averaged the lake values of the Littoral Cover Complexity and Riparian Vegetation Cover Complexity indices to calculate the Littoral–Riparian Habitat Complexity Index LitRipCvQ:
(9) where:
RVegQ_n = variant of the Riparian Vegetation Cover Complexity Index (n = 2, 7, or 8, depending on ecoregion; ); and
LitCvrQ_x = variant of Littoral Cover Complexity Index (x = b, c, or d, depending on lake type; ).
Lakeshore Physical Habitat Quality Index (LkShoreHQ)
We calculated an integrated Lakeshore Physical Habitat Quality Index (LkShoreHQ) by averaging the indices of Riparian Vegetation Cover Complexity, Littoral Cover Complexity, and [lack of] near-shore anthropogenic disturbance:
(10) where RDis_IX is calculated from Equationequation 2
(2) ; n denotes the appropriate variant of RVegQ by ecoregion (); and x denotes the appropriate variant of LitCvrQ by lake type ().
Deriving expected index values under least-disturbed conditions (Step 5)
We used 2 modeling approaches, null models and lake-specific predictive models, to estimate physical habitat expectations under least-disturbed condition. For the null models, we based expectations on the mean and distribution of log10-transformed physical habitat index scores in least-disturbed lakes from each ecoregion, an approach applied in previous bioassessments (e.g., Stoddard et al. Citation2005, Citation2006). We used the null model approach for all ecoregions except the West (WMT and XER).
For the West, we needed an alternative approach to reduce the variability in physical habitat condition indices among least-disturbed lakes. Air temperature, precipitation, soils, and lithology vary greatly across the West, resulting in wide variation in potential natural vegetation among least-disturbed lakes. In turn, that variation results in differences in the amount and complexity of littoral cover, especially for those elements derived from riparian vegetation. We derived lake-specific expected values in the West by modeling the influence of important nonanthropogenic environmental factors in relatively undisturbed lakes, an approach analogous to that used to predict least-disturbed conditions for multimetric fish assemblage indices (Pont et al. Citation2006, Citation2009, Esselman et al. Citation2013). We attempted unsuccessfully to derive similar regressions for the combined Central Plains ecoregion.
We used multiple linear regressions (MLR) to calculate lake-specific expected values of the Riparian Vegetation, Littoral Cover, and Littoral–Riparian Cover Complexity indices in the West, employing latitude, elevation, and ecoregion (WMT vs. XER) as surrogates for temperature, precipitation, soil, and other characteristics that control potential natural vegetation (). Ideally, expected cover and complexity would be calculated based only on least-disturbed lakes; however, the number of least-disturbed lakes in the West () was too small to adequately model expected lake physical habitat metrics values in relatively undisturbed lakes across this region. We therefore combined least-disturbed and intermediately disturbed lakes not greatly affected by drawdown (146 in WMT and 53 in XER) to avoid potential bias in modeled expectations by inadequately covering the range of the relevant natural controlling factors in our MLR. Excluding highly disturbed lakes from the MLRs minimized anthropogenic distortions of the influences of elevation, latitude, and ecoregion on lake physical habitat. Including many intermediately disturbed lakes in our regression predictions, however, necessitated a further adjustment of expected values, explained in the following section.
Table 4 Multiple linear regression models used for calculating expectations for indices of Riparian Vegetation Cover Complexity, Littoral Cover Complexity, and Littoral–Riparian Habitat Complexity for lakes in the Western Mountain and Xeric aggregated ecoregions. Highly disturbed lakes were excluded from calculations.
Condition criteria for near-shore lake physical habitat (Step 6)
For the Lakeshore Anthropogenic Disturbance Index RDis_IX, we used uniform criteria for all lakes. For RVegQ, LitCvrQ, and LitRipCvQ we set condition criteria based on the distribution of values of these indices observed in least-disturbed lakes. We have not yet set condition criteria for LkShoreHQ.
Lakeshore Anthropogenic Disturbance (intensity and extent) – condition criteria
Because RDis_IX is a direct measure of anthropogenic activities, we based criteria for high, medium, and low levels of disturbance on judgment:
Good (low disturbance): RDis_IX ≤ 0.20,
Fair (medium disturbance): RDis_IX > 0.20 but ≤ 0.75, and
Poor (high disturbance): RDis_IX > 0.75.
Lakes with RDis_IX ≤ 0.20 have very low levels of lake and near-lake disturbance, typically having anthropogenic disturbance on <8% of their shorelines. Those with RDis_IX > 0.75 have very high levels of disturbance, typically having anthropogenic activities evident on 100% of their shorelines.
Condition criteria for RVegQ, LitCvrQ, and LitRipCvQ
No model perfectly predicts expected values in least-disturbed lakes; rather, the models predict distributions of values. Furthermore, because the screening process for least-disturbed lakes was not perfect, disturbance was another source of variation in physical habitat index values in these lakes. Consequently, we set condition criteria for RVegQ, LitCvrQ, and LitRipCvQ with reference to the distributions of these indices in least-disturbed lakes within each of the 6 merged ecoregions (). Specifically, for ecoregions where we applied the null model, we compared log-transformed index values at each lake with the distribution of those values among least-disturbed lakes in their respective ecoregion.
Table 5 Mean and standard deviation of habitat index values in regional sets of least-disturbed reference lakes in the aggregated ecoregions of the NLA. Geometric mean and standard deviation are antilogs of mean and SD of log10-transformed index values (LogMean and LogSD). Parentheses enclose the minimum values (i.e., the most precise models for each index). Bold text marks the least precise models. SDs calculated from log-transformed variables are expressions of the proportional variance of these distributions and so are directly comparable among regions with different geometric means. A range of ±1SD is calculated by multiplying and dividing the geomMean by the geomSD. For example, the LogMean ± 1LogSD for the Riparian Vegetation Cover Complexity Index in least-disturbed lakes of the NAP (−0.571 ± 0.128) translates to a range of habitat index values from 0.200 to 0.359: the geometric mean habitat index value of 0.268 multiplied and divided by 1.34, the antilog of 0.128. The multiple 1.34 in this case is equivalent to a CV of 134%.
For the West, we calculated physical habitat index observed/expected (O/E) values for each lake by dividing the observed index value at each lake by the lake-specific expected value derived from regressions (). In most O/E applications, the calculated O/E value directly expresses the degree of alteration of each lake from values expected under least-disturbed conditions. Because we calculated expected values in the West using intermediately disturbed lakes in addition to least-disturbed lakes, we then compared the lake-specific O/E values to the distribution of O/E values in the 41 least-disturbed lakes in that region. This approach carries 2 assumptions: (1) the distribution of expected values in the least-disturbed lakes are accurately predicted from our MLR based on the set of lakes we used, and (2) the deviations of observed values from expected values are responses to anthropogenic disturbances and are not affected by the variables used to predict expected values.
We based our definitions of physical habitat alteration on percentiles of the distribution of physical habitat indices in least-disturbed lakes (or for the West, the distribution of O/E values in least-disturbed lakes); however, the small number of lakes meeting our low-disturbance criteria in most regions precluded obtaining reliable percentiles directly from the least-disturbed lake distributions. Consequently, we used the central tendency and variance of index values in least-disturbed lakes values to model their distributions and to estimate percentiles from their z-scores (Snedecor and Cochran Citation1980). The log10-transformed index values (and the log10-transformed O/E values for the West) in the least-disturbed lakes had symmetrical, approximately normal distributions. We calculated mean and standard deviations of log10-transformed data, converted them to z-scores, and estimated the 5th and 25th percentiles based on the log-normal approximation of the index distributions in least-disturbed lakes within each ecoregion.
Where null models were used to estimate least-disturbed condition, the index geometric mean and geometric standard deviation for each regional set of least-disturbed lakes represent the central tendency and dispersion of expected values for sample lakes within each region (, columns 5 and 6). Because means and standard deviations (SD) are all log values, a range of ±1 SD would be calculated by multiplying and dividing the geometric mean by the geometric SD.
For the MLR O/E models, the geometric mean and SD of O/E values in least-disturbed lakes describe the central tendency and distribution of O/E values expected under least-disturbed conditions (, column 5). The index geometric SD for each regional set of least-disturbed lakes describes the variation of expected values for sample lakes within their respective regions (, column 6). The geometric SDs for MLR-based O/E values in the least-disturbed lakes within regions are also directly comparable to those based on the null model.
Lakes with index values (null model) or O/E values (MLR model) that are ≥25th percentile for least-disturbed lakes within their regions were considered to have habitat in good condition (i.e., similar to that in the population of least-disturbed lakes of the region). Similarly, lakes with index or O/E values <5th percentile of least-disturbed lakes were considered to have poor habitat quality (i.e., they have significantly lower cover and complexity than observed within the subpopulation of least-disturbed lakes of the region). Those with index or O/E values between the 5th and 25th percentiles of least-disturbed lakes were scored as fair.
We emphasize that our designations of good, fair, and poor are relative to the least-disturbed sites available in each ecoregion. We define good condition as habitat quality that is well-within the distribution of habitat in least-disturbed sites of the ecoregion, and poor condition as habitat quality not likely to be found within that distribution. Our designations of poor condition do not indicate impaired water body status. Conversely, our designations of good condition mean that habitat is similar to the least-disturbed sites available in a region, which does not mean pristine, only the best available, and could be relatively disturbed in extensively and highly disturbed regions.
Population estimation procedures (Step 7)
Following Peck et al. (Citation2013), we estimated percentages and confidence bounds for the number of lakes with near-shore physical habitat in good, fair, and poor condition in the conterminous 48 states of the United States and its eco-regions. These lake population estimates employed sample site weighting factors inversely proportional to each lake's probability of being selected for sampling. We report findings nationally by the 9 ecoregions and separately for national populations of natural lakes versus reservoirs.
Table 6 Condition criteria for habitat index values and scaled habitat index values based on the distribution of index values in least-disturbed lakes within each region. The 5th and 25th percentiles, respectively, were set as the upper bounds for poor and fair condition, respectively. These percentiles were estimated, respectively, as the mean of log-transformed index values minus 1.65 and 0.67 times the SD of log-transformed habitat index values (see for the least-disturbed means and SDs, and see for the variant of each index used). All percentiles are expressed as antilogs of log-transformed values. For the null models, percentiles are in the units of the physical habitat indices (the O/E percentiles are not). Scaled percentiles in the last 2 columns are directly comparable between null and O/E models. Parentheses denote null model estimates that were replaced by lake-specific regression O/E model estimates in the West.
Results
Least-disturbed reference distributions and regressions
Geometric means for RVegQ, LitCvrQ, and LitRipCvQ in least-disturbed lakes differed among regions (), but these unscaled values are not directly comparable because the habitat index formulations also differed among regions. The unscaled mean values () served as denominators by which observed index values at each lake were scaled. In the West, the mean modeled O/E values RVegQ, LitCvrQ, and LitRipCvQ among least-disturbed lakes were divided into the modeled O/E values observed at each site, whether least-, intermediate-, or most-disturbed. Mean O/E values for least-disturbed lakes in the West (, column 5) were >1.0 for the 3 physical habitat indices (1.149, 1.007, and 1.130) because the regression analyses for expected condition included both least- and intermediate-disturbance lakes. This resulted because Littoral Cover Complexity in the least-disturbed lakes tended to be greater than for intermediate-disturbance lakes.
The geometric SDs for least-disturbed lakes of each region (, column 6) were calculated from log-transformed variables and therefore are expressions of the proportional variance of these distributions. As such, they are directly comparable as measures of model precision among regions with different geometric means, or between null and MLR modeling approaches. The precision in modeling least-disturbed condition using null models was best in the NAP (lowest geometric SD) and worst in the West (highest geometric SD). Null model geometric SDs were generally higher (less precise) for LitCvrQ (1.58–2.21) than for RVegQ (1.34–1.58) or LitRipCvQ (1.28–1.55).
The smaller the SD of index values (or O/E values) among least-disturbed lakes, the easier it is to confidently distinguish disturbed lakes from least-disturbed lakes. The MLR models for the West were marginally better than null models, but they improved the precision of models for expected condition in the West to within the range of those based on null models in the other regions. The proportions of total variance explained by the regression equations derived for the West were generally low (R2 < 0.25), but the models were highly significant (p < 0.0001) for RVegQ and LitRipCvQ (), with logSDs 8% lower than those for the null model. The equation for LitCvrQ, which used only elevation as a dependent variable, explained 5% of the total variance in LitCvrQ but was significant at p = 0.0075. Although the logSD of the model for estimating expected LitCvrQ in least-disturbed lakes of the West was only slightly lower than that for the null model, we chose to use it because it set realistically lower expectations in high elevation lakes where there is no natural source for littoral cover features derived from riparian trees.
Physical habitat index expectations and condition criteria
The (unscaled) 5th and 25th percentiles for least-disturbed lakes (, columns 3 and 4) are the habitat index values we used as condition criteria for the national lake population estimates. For the null model, these percentiles are in the units of the physical habitat indices described in equations 3–9; for the MLR models, they are O/E values. Although differences in habitat index criteria values for good, fair, or poor condition are evident by comparing these values, direct comparisons of the values among regions and indices are not appropriate because the index formulations and the central tendencies of their expected values in least-disturbed lakes differ among regions.
The scaled 5th and 25th percentiles of index values among least-disturbed lakes (, columns 5 and 6) are analogous to traditional O/E values. In our case they are calculated by dividing observed values by the mean in least-disturbed lakes and express the degree of deviation in habitat index values from the central tendency in least-disturbed lakes. Comparisons among scaled percentiles show the relative degree of alteration from least-disturbed conditions necessary for poor or fair condition classification in various regions or among indicators that have different ranges and means.
The scaled percentile values (condition criteria) also allow direct comparison between condition criteria derived from null and O/E models. Excluding those for the null model in the West, scaled 25th percentiles for the RVegQ in least-disturbed lakes showed little variation, ranging from 0.75 in the CPL to 0.82 in the NAP (, column 6). Scaled 5th percentiles of RVegQ in least-disturbed lakes also showed little variation, ranging from 0.50 in the West (O/E model) to 0.62 in the NAP (, column 5). For LitCvrQ, the scaled 25th percentile values for least-disturbed lakes were >0.7 in the NAP, CPL, and UMW, and lower (∼0.6) in the SAP, plains, and western regions. Scaled 5th percentiles of LitCvrQ in least-disturbed lakes were greatest (∼0.47) in the NAP and CPL and lowest (∼0.3) in the CENPL and West. For LitRipCvQ, scaled 25th percentile values were >0.8 for least-disturbed lakes in all aggregated ecoregions and habitat regions except the CENPL and the West (∼0.76). The lowest scaled 5th percentile values for LitRipCvQ were for least-disturbed lakes in the CENPL and the West (∼0.51), with all other regions having values ∼0.59–0.67.
Regional distributions of index scores and scaled index scores for all lakes
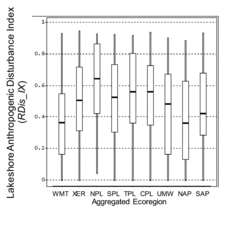
Distributions of RDis_IX are directly comparable within and among regions because its formulation and the criteria for defining low, medium, and high levels were identical everywhere. Interquartile ranges of RDis_IX in sample lakes varied widely among the 9 ecoregions, but extreme values spanned nearly the full potential range of the index in every region (). Lakeshore Anthropogenic Disturbance was distinctly highest for sample lakes in the NPL and lowest in the WMT and NAP ( unweighted sample statistics).
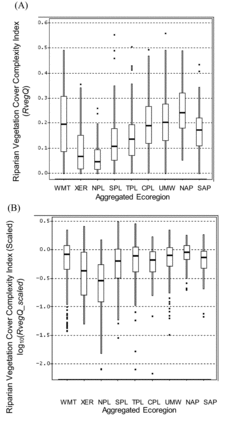
The sample distributions of unscaled values among regions are not directly comparable for the other physical habitat indices (RVegQ, LitCvrQ, and LitRipCvrQ; , , and ) because formulations differed among regions. The scaled index values allow interregional comparisons of the degree of alteration relative to least-disturbed conditions (, , and ). For example, the unscaled index RVegQ was highest in the WMT and NAP and lowest in the NPL and XER (), but the inclusion of natural rock as a riparian cover category in WMT and XER complicates interregional comparisons. Similarly, the NPL, SPL, and TPL regions excluded the top layer (height >5 m) in RVegQ formulation. After RVegQ was scaled, Riparian Vegetation Cover Complexity was relatively less altered in the WMT, NAP, UMW, and TPL than in the NPL or XER ().
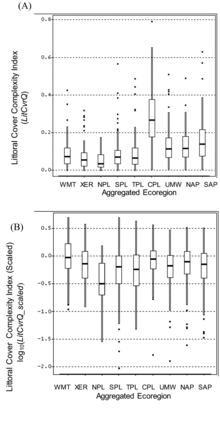
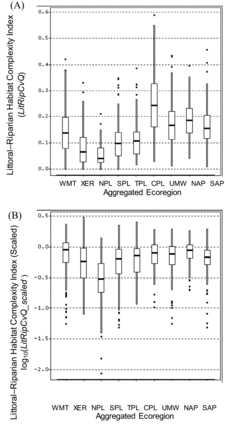
The notably high unscaled values of LitCvrQ in the CPL () are not directly comparable to those in many of the other ecoregions where the index formulation decreased the weighting of submerged forms and increased that for emergent and floating macrophytes. After scaling LitCvrQ to facilitate regional comparisons, several ecoregions (e.g., WMT, CPL, and NAP) had many lakes similar to least-disturbed condition, with sample median LitCvrQ_scaled values near 1.0 (log10-transformed values near 0 in ). As with the unscaled values, the NPL had the lowest sample median.
As was the case for LitCvrQ, the high unscaled LitRipCvQ values in the CPL () are magnified by its different treatment of submerged aquatic macrophytes, and more direct regional comparisons are made by comparing LitRipCvrQ_scaled (). Median values of LitRipCvrQ_scaled approached 1 (log10 = 0) in the WMT, NAP, CPL and UMW. As was the case for the scaled Littoral Cover Complexity Index, the lowest median LitRipCvrQ_scaled was for lakes in the NPL. The NPL median log10(LitRipCvrQ_scaled) value <−0.5 shows that most sample lakes in that region had Littoral-Riparian Habitat Complexity Index index values less than one-third of the mean value in least-disturbed lakes of this ecoregion ().
Physical habitat index responses to anthropogenic disturbance
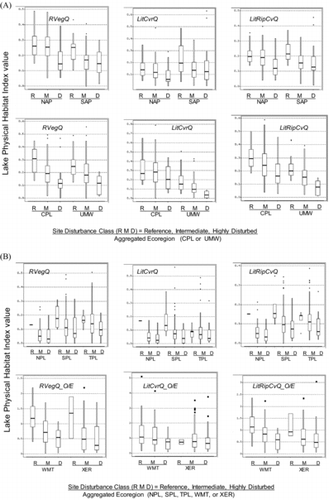
There were substantial declines in RVegQ, LitCvrQ, and LitRipCvQ from lakes with least to highest anthropogenic disturbance in nearly all 9 ecoregions (). Strong contrasts in RVegQ were evident between high and low disturbance lakes in all ecoregions, especially in the CPL, UMW, and WMT (). Among the 3 indices, LitCvrQ was least strongly related to anthropogenic disturbance. It was moderately related to disturbance in all regions except for the CPL, WMT, and XER, where the relationship was weak. LitRipCvQ showed steep declines with anthropogenic disturbance in all ecoregions, especially the UMW, SPL, and WMT ().
Except for the O/E indices in the West, we presented the physical habitat indices in without scaling to highlight the relative differences in potential cover and complexity among the disturbance classes in the original units of these indices. Within these regions, unscaled index values for different anthropogenic disturbance classes are directly comparable; however, scaling the indices was necessary for the 2 ecoregions of the West because the expected values under least-disturbed conditions differ for individual lakes. Plots of RVegQ, LitCvrQ, and LitRipCvQ for the West (not shown) look similar to the O/E plots (), but they do not distinguish anthropogenic disturbance categories as clearly as do the O/E plots, which model part of the natural variability of habitat among lakes throughout the West.
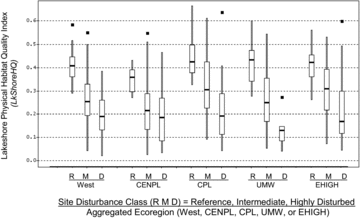
The LkShoreHQ index clearly discriminated reference lakes from disturbed lakes in all regions of the United States, showing progressive decreases with increasing anthropogenic stress from reference, through intermediate, to highly disturbed lakes. LkShoreHQ is an integrative measure of habitat complexity and disturbance, and one of its subcomponents (RDist_IX) was used in screening reference and disturbed sites (). These variables were used with 7 water chemistry screens and evidence of anthropogenic drawdown to classify lake anthropogenic pressures (R, M, and D in ). The distinct separation of disturbance levels (Fig. 7) is therefore not only a response of habitat to near-site disturbance. As for the other physical habitat indices, the declining levels of LkShoreHQ with disturbance classes are a reflection of the covariance of littoral and riparian habitat complexity with near-shore anthropogenic disturbance, eutrophication, chemical perturbation, and basin land-uses and disturbances associated with the lake water quality screening variables.
NLA population estimates for near-shore physical habitat
Population estimates () were based on data from 1033 probability-selected lakes representing ∼49,500 lakes in the conterminous United States that have depths >1 m and surface areas >4 ha. To interpret these populations (), we estimate, for example, that 34.8% (SE = 2.74%) of the total number of lakes have low lakeshore disturbance (, top left). This means that an estimated 17,226 lakes (±1356) had low values of lakeshore disturbance (RDis_IX < 0.20).
Table 7 Population estimates of the percentage of lakes (standard errors in parentheses) in good, fair, or poor condition with respect to NLA indices of near-shore anthropogenic disturbance and habitat cover complexity. Estimates are provided for the conterminous 48 US states for natural lakes, for manmade lakes (reservoirs), and for 9 aggregated Omernik ecoregions (adapted from USEPA Citation2009). Highest and lowest percentages of the 2 polar condition classes are shown in bold font. Underlined percentages indicate highest disturbance or poorest condition. National and regional lake sample and population numbers are shown on the bottom 2 rows. Only probability sample lakes are included in the sample size for population estimates. Physical habitat field measurements and observations were not made on lakes with surface areas >5000 ha (number of lakes shown in last row) resulting in a percentage of the lake population for which habitat condition was not assessed. “No data” represent lakes that were sampled, but habitat data were not collected or were determined to be invalid after review.
We lacked physical habitat data for some of the 1033 probability-selected lakes, leading to sets of lakes for which condition could not be determined. Physical habitat data were not collected at the 44 sample lakes that had surface areas >5000 ha, and these represent 190 (±77) lakes in the total population. Depending on the particular habitat index, missing or incomplete physical habitat data precluded calculating habitat condition at an additional 9 to 15 sample lakes, and these represent 131 (±126) to 189 (±39) lakes in the national population. These unassessed lakes represent <0.5% of the national population and ≤1% of lakes in any ecoregion ().
Population estimates for RDis_IX and the 3 physical habitat indices show a substantial number and percentage of lakes in the conterminous United States with high lakeshore anthropogenic disturbance and near-shore physical habitat having low cover complexity (poor condition), but the results differ by region and lake type (). Although 35% of the target lake population exhibited RDis_IX ≤ 0.20 and 17% had RDis_IX > 0.75, natural lakes had less lakeshore disturbance (46% low, 12% high) than did manmade lakes (18% low, 24% high). Nationally, 46% of lakes had high RVegQ whereas 36% had very low RVegQ (). With respect to LitCvrQ, 59% of lakes had high and 20% had low values. For LitRipCvQ, 47% of lakes were in good condition (high complexity) and 32% were in poor condition (low complexity). Natural lakes tended to have higher values of all 3 physical habitat complexity indices than did manmade lakes.
Population estimates of RDis_IX were lowest in the WMT and UMW and greatest in the NPL and XER ecoregions (). The NAP, TPL, and UMW ecoregions had the highest percentages of lakes with RVegQ in good condition. Despite having the highest percentage of lakes with RVegQ comparable to least-disturbed condition (66%), the NAP had the lowest percentage (46%) with LitCvrQ comparable to least-disturbed condition. Although the WMT had the second lowest percentage of lakes with high RDis_IX, it ranked below the NAP, TPL, and UMW in terms of the percentage of lakes with RVegQ in good condition. Among all 9 ecoregions, the NPL had the highest percentages of lakes in poor condition for all 4 physical habitat indices and the lowest percentage of lakes in good condition for all indices except LitCvrQ, for which it had the second lowest percentage.
Discussion
Implications of uncertainty in modeling condition in least-disturbed lakes
The power to discriminate altered habitat from that in least-disturbed lakes depends on the magnitude of the alteration relative to the central tendency and dispersion in the distribution of index scores among least-disturbed lakes, measured by the reference means and SDs (). The effect of this interaction can be illustrated by box-and-whisker plots (). Reductions in RipVegQ and LitRipCvQ in disturbed lakes are large relative to the median and the magnitude of their dispersion among least-disturbed (reference) lakes in nearly all the ecoregions, giving us confidence in the condition classifications for these indices. Conversely, reductions in LitCvrQ among disturbed lakes are small relative to the median and the magnitude of dispersion among least-disturbed (reference) lakes in nearly all the ecoregions, particularly the West (WMT and XER) and CENPL (NPL, SPL, and TPL) ecoregions. LitCvrQ index alterations are therefore, more difficult to discern, and the net effect is to underestimate habitat alteration.
Variation in natural conditions not captured by the expected condition regressions () contributes to the uncertainty in the LitCvrQ condition classifications in the West. Consequently, population estimates of the percentages of lakes in poor condition with respect to LitCvrQ may be underestimated in the West. The CENPL ecoregions have both a large SD and a substantial amount of anthropogenic disturbance in the combined set of least-disturbed lakes used to set condition criteria. Inclusion of substantial amounts of anthropogenic disturbance in the set of least-disturbed CENPL lakes reduces the central tendency of LitCvrQ among those lakes. Compared with other ecoregions, larger reductions in LitCvrQ must be observed in sample lakes of the CENPL before we would classify them in poor condition with respect to that index. Again, the result of these limitations is that our population estimates of the percentages of lakes having poor LitCvrQ are likely underestimates for ecoregions of the CENPL.
Lake physical habitat condition in the US
Among all the chemical and physical condition indicators measured in the NLA, degradation of RVegQ and LitRipCvQ were the most widespread (USEPA Citation2009). Physical habitat condition in lakes of every ecoregion was negatively associated with the gradient of anthropogenic disturbance of basins and near-shore zones expressed by the classification of least-disturbed reference, intermediate, and highly disturbed lakes (). The numbers of US lakes with poor littoral or riparian habitat condition exceeded those with excessive nutrients (nitrogen and phosphorus), turbidity, low dissolved oxygen, or acidity (USEPA Citation2009). Although physical and chemical degradation were more widespread in manmade lakes, the relative ranking of the types of degradation by their extent was similar for both lake types.
The ecological impact of physical habitat degradation on lakes can be quantified by evaluating the relative risk to lake biota when physical habitat degradation occurs (Van Sickle Citation2013). In the relative risk context, we refer to physical and chemical habitat degradation as stressors on biota, recognizing that these proximal stressors are in-turn influenced by more distal pressures such as near-shore, basin, and landscape-scale anthropogenic disturbances (Allen et al. Citation1999), as they are in streams (Sály et al. Citation2011, Marzin et al. Citation2013, CitationMacedo et al. forthcoming).
Certainly basin- and landscape-scale pressures affect lake physical, chemical, and biological habitat, but the relative and attributable risk analyses reported by USEPA (Citation2009) and Van Sickle (Citation2013) did not focus on those distal pressures. USEPA (Citation2009) reported that the highest risk to zooplankton and phytoplankton assemblages nationally was associated with degradation of RVegQ around lakes. RVegQ had a relative risk of 3 for biological plankton assemblages, meaning that lakes with poor riparian vegetation cover complexity were 3 times more likely to exhibit poor biological conditions than those that did not have poor riparian cover complexity (USEPA Citation2009). Degradation of LitCvrQ, LitRipCvQ, excess nitrogen and phosphorus, and turbidity had relative risks between ∼2 and 2.5 (USEPA Citation2009), showing the potential importance of both littoral–riparian cover complexity and water quality for protecting or improving biotic condition. Van Sickle (Citation2013) bundled similar stressors in the NLA and found the relative risk to lake biotic condition from degradation of combined lake physical habitat stressors (∼3) was greater than that for chemical stressors, including excess nutrients (∼2).
Although the apparent importance of lakeshore and littoral habitat to planktonic assemblages may be unexpected, most US lakes are small and relatively shallow; 80% of the lakes in the NLA population have surface areas <0.5 km2 (USEPA Citation2009). Especially in small or shallow lakes, pelagic water quality and plankton can be directly or indirectly affected by lakeshore disturbance and littoral habitat conditions. Riparian vegetation may buffer lakes from diffuse inputs of nutrients, sediment, and toxins (Allen et al. Citation1999, Wetzel Citation2001). Even in lakes of moderate size, primary production may be dominated by littoral processes, including nutrient uptake by aquatic macrophytes (Wetzel Citation1975). Littoral aquatic macrophytes can also affect dissolved nutrient concentrations and provide cover and structure that influences phytoplankton, zooplankton, macrobenthos, and fish (Wetzel Citation1975, Citation2001, Strayer and Findlay Citation2010). Thus, littoral cover complexity can directly influence lake plankton by providing cover and structure for planktonic organisms and by moderating water quality. Riparian vegetation cover complexity can positively influence plankton through inputs of structural elements that provide littoral structure (e.g., snags) and by buffering the lake from upland and lakeside anthropogenic activities, disturbances, and contaminant inputs.
The ecological importance of physical or chemical habitat degradation as stressors to lake biota is dependent on both the extent of the stressors and the severity of biological impacts when the stressors are present (Van Sickle et al. Citation2006). Attributable risk is derived by combining the relative extent and the relative risk into a single number. Conceptually, attributable risk is an estimate of the proportion of poor biological conditions that could be reduced if a particular stressor were restored from poor condition to fair or better condition, assuming that the relative risk association is causative, stressors are independent, and the stressor effects are reversible (Van Sickle Citation2013). Other important stressors (e.g., excess nutrients, fine sediment, or toxins) can be moderated by riparian buffers, and the recovery of biota after riparian rehabilitation will also depend on the extent to which associated stressors are reduced.
The attributable risk value represents the importance of a potential stressor, and ranking such risks can be useful for policy makers and managers. The NLA estimated the attributable risk of various lake stressors to biological condition expressed by the planktonic O/E index (USEPA Citation2009, Van Sickle Citation2013). Reduction in RVegQ had the highest attributable risk for phytoplankton and zooplankton taxa loss (43%) among the stressors examined. Thus, we expect lake planktonic taxa richness would be most improved by improving conditions of lakeshore vegetation cover and complexity, including inundated wetland vegetation. Removing riparian degradation as a stressor could potentially restore 43% of ∼11,000 (4700) lakes in poor biological condition nationally to intermediate or good condition, or approximately 9.5% of lakes nationwide.
Related research
Many studies at smaller spatial scales corroborate our national findings associating littoral and riparian habitat simplification with anthropogenic development around lakes (Kaufmann et al. Citation2014b). To date, however, no results from other national probability surveys of lake physical habitat are available for direct comparison with the NLA results. Interestingly, a recent assessment of the ecological condition of wadeable streams in the United States (Paulsen et al. 2008) showed results similar to those we report for lakes. Both assessments found the greatest percentages of near-shore (or near bank) disturbance, poor riparian vegetation cover complexity, and poor littoral or instream cover complexity in the NPL. They both identified the WMT as the region with the least near-bank disturbance and lowest percentage of waters with poor physical habitat condition. The relative ranking of stressors by extent and severity was also similar between the national lake and stream surveys (Paulsen et al. Citation2008, Van Sickle Citation2013). Both showed that physical habitat simplification was a pervasive stressor with high relative risk to biological condition.
The results of Miranda et al. (Citation2010) are similar to ours, although their survey concerned the condition of 494 randomly selected US public reservoirs ranging from 200 to 131,000 ha selected from the National Inventory on Dams database (NID Citation2008). In addition, they based their assessment on the expert opinion of 221 lake managers responding to an online survey. They reported that well-established riparian zones and wetlands were absent around many reservoirs, and the littoral zones of reservoirs were generally barren, lacking woody debris and native aquatic plants. In some cases, reservoirs contained excessive growth of nonnative aquatic vegetation. Miranda et al. (Citation2010) also cited degradation of reservoir physical habitat by fluctuating water levels and drawdown that limited access to backwaters and reduced shoreline–riparian interaction.
We can also compare a smaller-scale probability survey of lakes in the Northeastern United States with the NLA results. The physical habitat field methods were similar in the 2 surveys (Kaufmann et al. Citation2014a), but the Northeastern lakes survey included smaller lakes than surveyed by NLA in 2007. In addition, results were based on slightly different formulations of lakeshore disturbance indices and different condition criteria than used for the NLA (Kaufmann et al. Citation2014a, Citation2014b). Despite these differences, major results and conclusions concerning the importance of habitat simplification and its association with anthropogenic disturbances were similar between the 2 surveys.
Whittier et al. (Citation2002) reported substantially greater riparian and littoral habitat structural complexity in the 27% (±9%) of the lake population in the Northeast having no direct evidence of shore zone anthropogenic activities compared with those having more shoreline anthropogenic disturbance. Conversely, the 23% (±10%) of Northeast lakes with large amounts of lakeshore anthropogenic disturbance had half the riparian woody cover and lower littoral habitat complexity. Whittier et al. (Citation2002) concluded that although stressors such as nonnative fish introductions, mercury contamination, and lakeshore physical habitat alteration were not a focus for lake management at that time, they were as widespread as eutrophication and more extensive than acidification in Northeastern lakes. Brauns et al. (Citation2007) and USEPA (Citation2009), respectively, emphasized the need to recognize physical habitat alteration as an important stressor meriting education, management, and regulatory attention for European and US lakes.
Kaufmann et al. (Citation2014b) found that the strongest relationships with fish and bird assemblages in the Northeastern lake survey were with LkShoreHQ. Compared with its subcomponents and with basin road density, LkShoreHQ was the best predictor of intolerant fish species richness, tolerant fish species richness, the percentage of neotropical migrant birds, and the percentage of tolerant bird taxa in the near-shore lake environment of lakes in the Northeastern lake survey (Kaufmann et al. Citation2014b).
Kaufmann et al. (2014a) reported good precision and signal:noise ratio for LkShoreHQ in the NLA, with ratio of repeat visit standard deviation:potential range ratio (SDrep/Rgpot) = 0.05 and S/N = 4.2. We believe that LkShoreHQ has great potential in regional and national lake assessments as an integrated index that combines the dimensions of anthropogenic disturbance, cover, and structural complexity in the terrestrial and aquatic compartments of the near-shore lake environment. The strong association of LkShoreHQ with the disturbance gradients we used () may be somewhat accentuated because field measures of riparian anthropogenic disturbance contributing to LkShoreHQ were also among the many variables used to screen least- and most-disturbed lakes. To avoid circularity in assessments, condition criteria for LkShoreHQ could be set based on biotic tolerances (e.g., Bryce et al. 2010, Whittier and Van Sickle Citation2010) or in the manner we described for the other lake habitat complexity indices, but by employing reference sites that do not use lakeshore disturbance in their screening.
Summary and recommendations
Our NLA findings show widespread alteration and degradation of near-shore riparian and littoral habitat in lakes of the conterminous 48 US states. Lake physical habitat simplification was associated with basin and near-shore anthropogenic disturbances in all regions. Many smaller-scale studies have associated lakeshore anthropogenic disturbances with habitat simplification and reduction in biological condition (Kaufmann et al. Citation2014b). Our results validate the generality of responses to known stressors reported in those studies. In addition, the more intensive, smaller-scale research suggests plausible mechanisms underlying the associations we observed. The weight of evidence from combining this knowledge increases the potential to make reasonable decisions concerning conservation and rehabilitation of physical habitat quality and biological condition in lakes and watersheds that have not been extensively studied.
Of the 8 types of in-lake or near-shore physical and chemical degradation examined by the NLA (USEPA Citation2009), poor quality (low) lake Riparian Vegetation Cover Complexity was identified as the most widespread. Attributable risk analyses suggest that eliminating the effects of poor lakeshore habitat cover could improve the biological condition in many lakes nationwide (USEPA Citation2009). Therefore, in addition to better managing basin land use, we suggest that lake managers at local, state, and national levels should better protect and rehabilitate littoral and riparian physical habitat through better management of anthropogenic disturbance in near-shore areas. Anthropogenic disturbance along lakeshores (such as tree removal and residential construction) impairs lakeshore and shallow water habitats, affecting terrestrial and aquatic biota alike (Hughes et al. Citation2014a). Many states and the USEPA are promoting low impact development and maintenance or restoration of green infrastructure to protect and rehabilitate more natural hydrology and to improve biological and physical and chemical habitat conditions in surface waters (USEPA Citation2007b, Hughes et al. Citation2014b).
Acknowledgments
The field data used in our study were collected by Federal and State agency collaborators, USEPA regional staff, and contract crews from TetraTech, GLEC, and Dynamac too numerous to mention here, but their conscientious field work made our paper possible. We thank C. Burch-Johnson for preparing the maps; John Van Sickle, Tony Olsen, and Tom Kincaid for technical advice concerning design, statistics and analysis; and Susan Holdsworth, Sarah Lehmann, and Amina Pollard for NLA project management and funding. Our manuscript was improved as a result of recommendations from Bob Ozretich, Ekatrina Kovalenko, Kellie Merrell, Chuck Hawkins, and 5 anonymous reviewers. The manuscript was reviewed by the National Health and Environmental Effects Research Laboratory's Western Ecology Division and approved for publication. Approval does not signify that the contents reflect the views of the Agency, nor does mention of trade names or commercial products constitute endorsement or recommendation for use.
Funding
Funding for data analysis and manuscript preparation was largely from the USEPA Office of Research and Development in support of the EPA Office of Water's National Lakes Assessment through cooperative agreement #CR-818606 with Oregon State University and contract GSFU594G with Raytheon Information Services. Bob Hughes was supported in manuscript preparation during sabbatical in Brazil by Fulbright Brasil, Companhia Energetica de Minas Gerais (CEMIG), and Fundacao de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) grant 00011/09.
References
- Allen AP, Whittier TR, Kaufmann PR, Larsen DP, O’Connor RJ, Hughes RM, Stemberger RS, Dixit SS, Brinkhurst RO, Herlihy AT, Paulsen SG. 1999. Concordance of taxonomic composition patterns across multiple lake assemblages: Effects of scale, body size, and land use. Can J Fish Aquat Sci. 56:2029–2040.
- Barker WT, Whitman WC. 1988. Vegetation of the Northern Great Plains. Rangelands. 10:266–272.
- Beck MW, Vondracek B, Hatch LK. 2013. Between- and within-lake responses of macrophyte richness metrics to shoreline development. Lake Reserv Manage. 29:179–193.
- Benson BJ, Magnuson JJ. 1992. Spatial heterogeneity of littoral fish assemblages in lakes: Relation to species diversity and habitat structure. Can J Fish Aquat Sci. 49:1493–1500.
- Brauns M, Garcia X-F, Walz N, Pusch MT. 2007. Effects of human shoreline development on littoral macroinvertebrates in lowland lakes. J Appl Ecol. 44:1138–1144.
- Bryce SA, Lomnicky SG, Kaufmann PR. 2010. Protecting sediment-sensitive aquatic species in mountain streams through the application of biologically-based streambed sediment criteria. J N Am Benthol Soc. 29:657–672.
- Carpenter SR, Cottingham KL. 1997. Resilience and restoration of lakes. Conserv Ecol 1(1):. w. . Available from: http://www.consecol.org/vol1/iss1/art2
- Christensen DL, Herwig BR, Schindler DE, Carpenter SR. 1996. Impacts of lakeshore residential development on coarse woody debris in north temperate lakes. Ecol Appl. 6:1143–1149.
- Engel S, Pederson J. 1998. The construction, aesthetics, and effects of lakeshore development: A literature review. Madison (WI) : Wisconsin Department of Natural Resources. . Report 177. Available from: http://digital.library.wisc.edu/1711.dl/EcoNatRes.DNRRep177
- Esselman PC, Infante DM, Wang L, Cooper AR, Wieferich, Tsang Y-P, Thornbrugh DJ, Taylor WW. 2013. Regional fish community indicators of landscape disturbance to catchments of the conterminous United States. Ecol Indic. 26:163–173.
- Faustini JM, Kaufmann PR. 2007. Adequacy of visually classified particle count statistics from regional stream habitat surveys. J Am Water Resour As. 43(5):1293–1315.
- Francis TB, Schindler DE. 2006. Degradation of littoral habitats by residential development: Woody debris in lakes of the Pacific Northwest and Midwest, United States. Ambio. 35:274–280.
- Halliwell D. 2007. Lake habitat measures. New England Lake News. New England Chapter of the North American Lake Management Society. 2(2):1–4.
- Hampton SE, Fradkin SC, Leavitt PR, Rosenberger EE. 2011. Disproportionate importance of nearshore habitat for the food web of a deep oligotrophic lake. Mar Freshw Res. 62:350–358.
- Hatzenbeler GR, Kampa JM, Jennings MJ, Emmons EE. 2004. A comparison of fish and aquatic plant assemblages to assess ecological health of small Wisconsin lakes. Lake Reserv Manage. 20:211–218.
- Herlihy AT, Kamman NC, Sifneos JC, Charles D, Enache MD, Stevenson RJ. 2013. Using multiple approaches to develop nutrient criteria for lakes in the conterminous USA. Freshwater Sci. 32(2):361–384.
- Huddle JA, Awada T, Martin DL, Zhou X, Pegg SE, Josiah SJ. 2011. Do invasive riparian woody plants affect hydrology and ecosystem processes? Digital Commons at University of Nebraska – Lincoln. Papers in Natural Resources. Paper 298. . Available from: http://digitalcommons.unl.edu/natrespapers/298
- Hughes RM, Dunham S, Maas-Hebner KG, Yeakley JA, Schreck CB, Harte M, Molina N, Shock CC, Kaczynski VW, Schaeffer J. 2014a. A review of urban water body challenges and approaches: (1) Rehabilitation and remediation. Fisheries. 39:18–29.
- Hughes RM, Dunham S, Maas-Hebner KG, Yeakley JA, Schreck CB, Harte M, Molina N, Shock CC, Kaczynski VW. 2014b. A review of urban water body challenges and approaches: (2) Mitigation and research needs. Fisheries. 39:30–40.
- Hughes RM, Herlihy AT, Kaufmann PR. 2010. An evaluation of qualitative indexes of physical habitat applied to agricultural streams in ten U.S. states. J Am Water Resour As. 46(4):792–806.
- Jennings MJ, Bozek MA, Hatzenbeler GR, Emmons EE, Staggs MD. 1999. Cumulative effects of incremental shoreline habitat modification on fish assemblages in north temperate lakes. N Am J Fish Manage. 19:18–27.
- Jennings MJ, Emmons EE, Hatzenbeler GR, Edwards C, Bozek MA. 2003. Is littoral habitat affected by residential development and land use in watersheds of Wisconsin lakes?Lake Reserv Manage. 19:272–279.
- Johnson WC. 2002. Riparian vegetation diversity along regulated rivers: Contribution of novel and relict habitats. Freshwater Biol. 47:749–759.
- Kaufmann PR, Faustini JM. 2012. Simple measures of channel habitat complexity predict transient hydraulic storage in streams. In: The importance of habitat complexity in waterscapes. Hydrobiologia (Special Issue). 685:69–95.
- Kaufmann PR, Hughes RM, Van Sickle J, Whittier TR, Seeliger CW, Paulsen SG. 2014a. Lake shore and littoral habitat structure: A field survey method and its precision. Lake Reserv Manage. . 30:157–176.
- Kaufmann PR, Hughes RM, Whittier TR, Bryce SA, Paulsen SG. 2014b. Relevance of lake physical habitat assessment indices to fish and riparian birds. Lake Reserv Manage. . 30: 177–191.
- Kaufmann PR, Larsen DP, Faustini JM. 2009. Bed stability and sedimentation associated with human disturbances in Pacific Northwest streams. J Am Water Resour As. 45:434–459.
- Kaufmann PR, Levine P, Robison EG, Seeliger C, Peck DV. 1999. Quantifying physical habitat in wadeable streams. Corvallis (OR) : US Environmental Protection Agency, Environmental Monitoring and Assessment Program. . EPA 620/R-99/003.
- Kaufmann PR, Whittier TR. 1997. Habitat assessment. In: Baker JR, Peck DV, Sutton DW, editors. Environmental Monitoring and Assessment Program; Surface waters: Field operations manual for lakes. Washington (DC) : US Environmental Protection Agency. . EPA/620/R-97/001. p. 5-1 to 5–26.
- Kovalenko KE, Thomaz SM, Warfe DM. 2012. Habitat complexity: approaches and future directions. Hydrobiologia. 685:1–17.
- Macedo DR, Hughes RM, Ligeiro R, Ferreira WR, Castro M, Junqueira NT, Silva DRO, Firmiano KR, Kaufmann PR, Pompeu PS, Callisto M. Forthcoming. The relative influence of multiple spatial scale environmental predictors on fish and macroinvertebrate assemblage richness in cerrado ecoregion streams, Brazil. Landscape Ecology.
- Marzin A, Verdonschot PFM, Pont D. 2013. The relative influence of catchment, riparian corridor, and reach-scale anthropogenic pressures on fish and macroinvertebrate assemblages in French rivers. Hydrobiologia. 704:375–388.
- Merrell K, Howe E, Warren S. 2009. Examining shorelines, littorally. Lakeline. 29:1.
- Miranda LR, Spickard M, Dunn T, Webb KM, Aycock JN, Hunt K. 2010. Fish habitat degradation in U.S. reservoirs. Fisheries. 35(4):175–184.
- [NID] National Inventory on Dams. 2008. Alexandria (VA) : US Army Corps of Engineers; . [cited 23 Feb 2014]. Available from: http://crunch.tec.army.mil/nidpublic/wepages/nid.cfm
- Omernik JM. 1987. Ecoregions of the conterminous United States. Ann Assoc Am Geogr. 77:118–125.
- Palmer MA, Covich AP, Lake S, Biro P, Brooks JJ, Cole J, Dahm C, Gibert J, Goedkoop W, Martens K, et al. 2000. Linkages between aquatic sediment biota and life above sediments as potential drivers of biodiversity and ecological processes. BioScience. 50:1062–1075.
- Paulsen SG, Mayio A, Peck DV, Stoddard JL, Tarquinio E, Holdsworth SM, Van Sickle J, Yuan LL, Hawkins CP, Herlihy AT, et al. 2008. Condition of stream ecosystems in the US: An overview of the first national assessment. J N Am Benthol Soc. 27:812–821.
- Peck DV, Olsen AR, Weber MH, Paulsen SG, Peterson C, Holdsworth SM. 2013. Survey design and extent estimates for the National Lakes Assessment. Freshwater Sci. 32:1231–1245.
- Polis GA, Anderson WB, Holt RD. 1997. Toward an integration of landscape and food web ecology: The dynamics of spatially subsidized food webs. Ann Rev Ecol System. 28:289–316.
- Pont D, Hughes RM, Whittier TR, Schmutz S. 2009. A predictive index of biotic integrity model for aquatic-vertebrate assemblages of western U.S. streams. T Am Fish Soc. 138:292–305.
- Pont D, Hugueny B, Beier U, Goffaux D, Melcher A, Noble R, Rogers C, Roset N, Schmutz S, 2006. Assessing river biotic condition at a continental scale: A European approach using functional metrics and fish assemblages. J Appl Ecol. 43:70–80.
- Radomski P, Geoman TJ. 2001. Consequences of human lakeshore development on emergent and floating-leaf vegetation abundance. N Am J Fish Manage. 21:41–46.
- Sály P, Takács P, Kiss I, Bíró P, Eros T. 2011. The relative influence of catchment- and site-scale environmental factors on stream fish assemblages in a human-modified landscape. Ecol Freshw Fish. 20:251–262.
- SAS Institute. 2008. SAS/STAT 9.2 User's Guide. Cary (NC) : SAS Institute Inc.
- Schindler DE, Scheuerell MD. 2002. Habitat coupling in lake ecosystems. Oikos. 98:177–189.
- Snedecor GW, Cochran WG. 1980. Statistical methods, . 7th ed. Ames (IA) : Iowa State University Press. 507 p.
- Stoddard JL, Larsen DP, Hawkins CP, Johnson RK, Norris RH. 2006. Setting expectations for the ecological condition of streams: The concept of reference condition. Ecol Appl. 16(4): 1267–1276.
- Stoddard JL, Peck DV, Paulsen SG, Van Sickle J, Hawkins CP, Herlihy AT, Hughes RM, Kaufmann PR, Larsen DP, Lomnicky G, et al. 2005. An ecological assessment of western streams and Rrivers. Washington (DC) : US Environmental Protection Agency. . EPA 620/R-05/005.
- Strayer DL, Findlay SEG. 2010. Ecology of freshwater shore zones. Aquat Sci. 72:127–163.
- Taillon D, Fox M. 2004. The influence of residential and cottage development on littoral zone fish communities in a mesotrophic north temperate lake. Environ Biol Fish. 71:275–285.
- [USEPA] US Environmental Protection Agency. 2007a. Survey of the nation's lakes. Field operations manual. Washington (DC) : US Environmental Protection Agency. . EPA 841-B-07-004.
- [USEPA] US Environmental Protection Agency. 2007b. Reducing stormwater costs through low impact development (LID) strategies and practices. Washington (DC) : US Environmental Protection Agency. . EPA 841-F-07-006.
- [USEPA] US Environmental Protection Agency. 2009. National lakes assessment: A collaborative survey of the nation's lakes. Washington (DC) : US Environmental Protection Agency, Office of Water and Office of Research and Development. . EPA 841-R-09-001.
- [USEPA] US Environmental Protection Agency. 2010. National lakes assessment: Technical appendix. Washington (DC) : US Environmental Protection Agency, Office of Water and Office of Research and Development. . EPA 841-R-09-001a. 63 p.
- Van Sickle J. 2013. Estimating the risks of multiple, covarying stressors in the National Lakes Assessment. Freshwater Sci. 32:204–216.
- Van Sickle J, Stoddard JL, Paulsen SG, Olsen AR. 2006. Using relative risk to compare the effects of aquatic stressors at a regional scale. Environ Manage. 38:1020–1030.
- Wagner T, Jubar AK, Bremigan MT. 2006. Can habitat alteration and spring angling explain largemouth bass nest success? T Am Fish Soc. 135:843–852.
- West E, Ruark G. 2004. Historical evidence of riparian forests in the Great Plains and how that knowledge can aid with restoration and management. . US Department of Agriculture, US Forest Service/University of Nebraska, Lincoln Faculty Publications. Paper 7. Digital Commons at University of Nebraska – Lincoln. Available from: http://digitalcommons.unl.edu/usdafsfacpub/7
- Wetzel RG. 1975. Limnology. Philadephia (PA) : W.B. Saunders.
- Wetzel RG. 2001. Limnology: Lake and river ecosystems. Waltham (MA) : Elsevier.
- Whittier TR, Halliwell DB, Paulsen SG. 1997. Cyprinid distributions in northeast U.S.A. lakes: Evidence of regional-scale minnow biodiversity losses. Can J Fish Aquat Sci. 54:1593–1607.
- Whittier TR, Paulsen SG, Larsen DP, Peterson SA, Herlihy AT, Kaufmann PR. 2002. Indicators of ecological stress and their extent in the population of Northeastern lakes: A regional-scale assessment. Bioscience. 52(3):235–247.
- Whittier TR, Van Sickle J. 2010. Macroinvertebrate tolerance values and an assemblage tolerance index (ATI) for western USA streams and rivers. J North Am Benthol Soc. 29:852–866.
- Zohary T, Ostrovsky I. 2011. Ecological impacts of excessive water level fluctuations in stratified freshwater lakes. Inland Waters. 1:47–59.