
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Zastepa A, Pick FR, Blais JM. 2017. Distribution and flux of microcystin congeners in lake sediments. Lake Reserv Manage. 33:444–451.
Sediment concentrations of microcystin congeners and exchange across the sediment–water interface were determined in Lake of the Woods, a large water body between Canada and the United States experiencing cyanobacterial blooms. Dated sediment cores were used to examine historical occurrence of microcystins and showed that microcystins were below detection prior to the 2000s. In more recent sediments the most abundant congeners were MC-LA and -LR with -RR, -YR, -7dmLR, -WR, -LF, -LY, and -LW also present. MC-LA and -LR were also distributed in the pore waters whereas MC-RR and -YR were more strongly adsorbed to sediment particles. Sediment burial rates for MC-LA and -LR were determined from the product of the microcystin concentration on sediment particles (ng/g dw) and the burial rate (based on 210Pb radiochronology [g/m2/d]). Diffusion from sediments was estimated from the concentration gradient between pore water of surficial sediments and overlying water using Fick's first law. Overall, burial rates were low across sites (2.6 to 298.1 ng/m2/d) when compared to diffusion of microcystins from sediments to overlying water (303.1 to 1078.0 ng/m2/d) suggesting that sediments can be a source of microcystins to the water column. However, the relatively high diffusive flux may be short term and the result of a temporal disconnect between water column productivity and sediment processes. The higher diffusion fluxes and lower burial rates of MC-LA compared to MC-LR point to differences in environmental fate. Given that microcystin congeners vary in their toxicity, these results highlight the need for congener-specific measurements of environmental fate and persistence.
Toxins produced by cyanobacteria are a major problem for water quality around the world and the hepatotoxic microcystins have garnered the most attention due to their high toxicity and prevalence in freshwater ecosystems (Chorus and Bartram Citation1999). With over 100 different congeners described (Meriluoto et al. Citation2017), chemical analyses and risk assessment of these compounds represent significant challenges.
Recent investigations indicate that microcystins can accumulate in sediments of lakes prone to cyanobacterial blooms (Efting et al. Citation2011, Zastepa et al. Citation2015, Zastepa et al. Citation2017b). This microcystin reservoir in sediment raises concerns about the potential for microcystins to remobilize into the overlying water, thus posing human and ecological health risks. Furthermore, an assessment of the potential for microcystin diffusion through sediments is of critical importance in the accurate interpretation of their historical record in sediments.
Despite a high number of microcystin congeners with a wide range of chemical properties, measurements in sediments have been limited to a few congeners of similar chemistry, principally MC-LR, -YR, and -RR (Babica et al. Citation2006, Chen et al. Citation2006). Growing experimental and field evidence indicates significant differences in the environmental fate of microcystin congeners and implications for exposure. For example, MC-LA, with a mammalian toxicity as high as MC-LR (Chorus and Bartram Citation1999), appears to be more persistent in aquatic systems (e.g., Newcombe et al. Citation2003, Zastepa et al. Citation2014) and can lead to wildlife fatalities through food web bioaccumulation (Miller et al. Citation2010). Congener-specific estimates of post-depositional mobility in sediments would provide much-needed information for risk assessments of cyanotoxins to human and wildlife health. Furthermore, this information is required to interpret the historical records of microcystins reported in sediments from paleolimnological studies (Pawlik-Skowronska et al. Citation2010, Efting et al. Citation2011, Waters Citation2016).
The specific objectives of this study were to (1) determine the distribution of microcystin congeners between sediment particles and pore waters in lake sediment cores; (2) estimate sediment deposition rates of microcystin congeners; and (3) determine whether sediments are a significant sink or source of microcystins to the overlying water, and whether there are congener-specific differences in these sediment–water diffusion fluxes. To achieve these objectives, we analyzed microcystins in sediment and pore waters from dated lake sediment cores from Lake of the Woods, Ontario, where toxic cyanobacterial blooms are pervasive (Chen et al. Citation2007, Watson and Kling Citation2017, Zastepa et al. Citation2017a).
Materials and methods
Study site
Lake of the Woods is a large (∼3850 km2), morphologically and hydrologically complex water body with variable depth (mean ∼8 m, max. ∼66 m), within a catchment area of approximately 70,030 km2 (excluding the lake area; Schupp and Macins Citation1977). It is made up of several interconnected basins occupying the Canadian provinces of Ontario and Manitoba as well as the US state of Minnesota. Water flows northward from the main inlet, the Rainy River, to the main outlet, the Winnipeg River. A strong gradient of total phosphorus generally tracks the flow of the Rainy River from the south, where the highest phosphorus loading occurs (Pla et al. Citation2005). Ice covers the lake from approximately November to April. The northern region of the lake is surrounded mostly by boreal forest and underlain by Precambrian Shield granitic bedrock, whereas the southern region is surrounded mostly by agricultural land and consists of old lake sediments deposited by glacial Lake Agassiz (Johnston Citation1915), resulting in shallower, more turbid, and more productive waters (Rusak and Mosindy Citation1997). Lake of the Woods and its catchment has a history of human impacts including land clearing by European settlers, logging and sawmill operations, and several dams constructed for water level management and hydroelectric power (Yang and Teller Citation2005, Serieyssol et al. Citation2009, Serieyssol-Bleser Citation2010). The lake has a history of cyanobacterial blooms but information on toxicity is limited (Chen et al. Citation2007, Edlund et al. Citation2017, Reavie et al. Citation2017). At the time of sediment core sampling, total phosphorus (TP) and pH across the 3 sites in Lake of the Woods were as follows: Rat Portage TP = 20 µg/L, pH = 7.66; Bigstone Bay TP = 18 µg/L, pH = 7.90; and Hay Island TP = 38 µg/L, pH = 7.64.
Sediment and water sampling
Lake of the Woods was sampled in July 2010 at 2 northern sites and 1 southern site. A 46 cm core was retrieved from polymictic Rat Portage Bay (49°43′20.7″ N, 94°33′12.7″ W) at a depth of 23 m. A 40 cm core was retrieved from dimictic Bigstone Bay (49°40′10.7″ N, 94°19′43.8″ W) at a depth of 23 m. Bigstone Bay thermally stratifies during the summer. A third core (40 cm) was retrieved from southern Hay Island (49°9′25.6″ N, 94°7′23.2″ W) at a depth of 13 m. Additionally, at each coring site, subsurface water (5 m depth) was collected using a Van Dorn sampler and stored in dark coolers with ice packs until filtering onto 47 mm GF/C filters on shore. Filters were extracted and analyzed for microcystins as described below.
Each sediment core was obtained using a modified gravity corer (Glew Citation1989). The overlying water from the top of the sediment core was also collected for diffusion calculations (500 mL). Cores were extruded on-site at 0.5 cm intervals, placed in pre-labeled Whirl-Pak bags, and immediately transported to the laboratory on ice in a dark cooler for pore water separation before being frozen in the dark at −20 C. Up to 22 lyophilized subsamples spanning the length of each core were prepared to determine age and sedimentation rate. This was achieved by measuring 210Pb and 226Ra activities with a high purity germanium detector in a gamma spectrometer (DSPec Spectrometer linked to Maestro II Software by Ortec, Tennessee, USA) according to methods of Appleby (Citation2001). Unsupported 210Pb was estimated by subtracting supported 210Pb (measured as 226Ra activity) from total 210Pb. As a result of nuclear arms testing circa 1954, 137Cs was used as an independent chronostratigraphic marker, peaking in 1963 prior to the Nuclear Test Ban Treaty. The constant rate of supply model (CRS), the constant initial concentration model (CIC), and the constant flux constant sedimentation model (CFCS) were evaluated to establish chronology in each core (Appleby and Oldfield Citation1978, Binford Citation1990, Blais et al. Citation1995). The CRS model was chosen because CRS and CIC dates were nearly equivalent to a depth of 25 cm and the CRS model has been shown to produce the best agreement with independent dating markers (Blais et al. Citation1995).
Extraction and analysis of microcystin congeners
Sediment core intervals were separated into a pore water fraction and a sediment particle fraction by centrifugation of 10 g aliquots of each 0.5 cm interval of sediment (1200 × g at 4 C for 25 min). Microcystins were extracted by accelerated solvent extraction using methanol and water (3:1), extracts cleaned by solid phase extraction and quantified by LC-MS/MS analysis according to Zastepa et al. (Citation2015). The analytical method was developed, optimized, and validated for the extraction and quantitation of microcystin-LR, -7dmLR, -RR, -YR, -WR, -LA, -LF, -LY, -LW with method detection limits ranging from 1.1 to 2.5 ng/g dw. Method detection limits for pore water, surface water, and water overlying sediment ranged from 0.06 to 0.17 µg/L, depending on the specific congener. Surface water and water overlying sediment were processed and measured according to Zastepa et al. (Citation2014).
Microcystin burial rates and flux calculations
Burial rates of microcystins in surficial sediments were calculated as the product of the concentration of microcystins on sediment particles (ng/g dw) and the particle sedimentation rate, which was obtained from 210Pb measurements (g/m2/d).
Fick's first law was used for estimating diffusion across the sediment–water interface, assuming steady state (Schwarzenbach et al. Citation2003). Diffusion of microcystin congeners from sediments was estimated based on the concentration gradient between the pore water of sediments (every 0.5 cm over the top 7.0 cm) and water overlying sediment. Diffusive flux was calculated at the 3 sites in Lake of the Woods.
(1)
(1)
where F is the flux of (ng/m2/d) of a solute with concentration C at depth x. φ is the porosity calculated using the following equation:
(2)
(2)
where a particle density of 2.65 g cm−3 was used (Brady and Weil Citation2002). θ is the tortuosity (dimensionless) and is related to porosity by the empirical relationship of Bourdeau (Citation1996):
(3)
(3)
Finally, Dw is the diffusion coefficient of the solute in water in the absence of sediment matrix. A diffusion coefficient of 1.4 × 10−6 cm2/s was calculated for both MC-LR and MC-LA using molecular volume according to Schwarzenbach et al. (Citation2003).
Results
Microcystin congener concentrations and distribution
At the time of sampling, surface water concentrations of microcystins ranged between 0.4 and 2.9 µg/L with MC-LA and MC-LR the dominant congeners. Within the sediment core from Hay Island, microcystins were detected down to 6.5 cm (). Sections from 6.5 cm to the end of the core (40 cm) had no detectable microcystins. The 6.5 cm depth corresponded to the early 2000s. The sediment core profile had 2 major peaks of total microcystins (sediment particles plus pore water) at a depth of 0.25 and 4.25 cm, which correspond to year 2010 and 2004, respectively. MC-LR and -LA were the dominant congeners in most intervals with the highest total concentration (sediment particles plus pore water) being MC-LR at almost 400 ng/g dw. MC-RR, -7dmLR, -YR, -LW, and -WR were generally present in relatively minor amounts with the exception of MC-RR at the 4.25 cm interval, where it was the dominant congener at a total concentration of 113 ng/g dw. MC-LA was detected in the pore waters of more than half the intervals where MC-LA was detected and typically more than half of MC-LA in each interval was distributed into the pore water (, ). In contrast, MC-LR and -RR, despite relatively higher concentrations, were only associated with sediment particles in all of the intervals where they were detected (, ) with the exception of the top core section (0–0.5 cm) where approximately 5% of pore water and sediment particle-bound MC-LR summed was present in the pore water.
Figure 1. Microcystin congener concentrations on sediment particles in the top 6.25 cm of a 40 cm sediment core from Hay Island, Lake of the Woods, Ontario. Below 6.25 cm, measurements were below the method detection limit. Interval midpoints are plotted (0.5 cm intervals from 0 to 7 cm). MC-LF and -LY were not detected in sediment. We analyzed a total of 80 half-cm intervals.
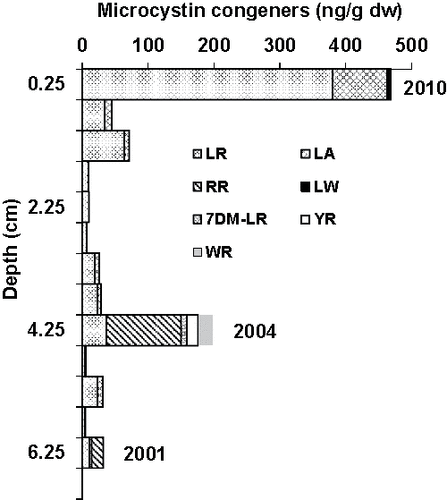
Figure 2. Individual microcystin congeners in the pore water of a 40 cm sediment core from Lake of the Woods (Hay Island) as a percentage of the congener in pore water and on sediment particles summed. Interval midpoints are plotted (0.5 cm intervals from 0 to 7 cm). Open circle (○) = LA, × = LR. RR, LW, 7dmLR, YR, and WR were not detected in pore water. MC-LF and -LY were not detected in the sediment core. We analyzed a total of 80 half-cm intervals.
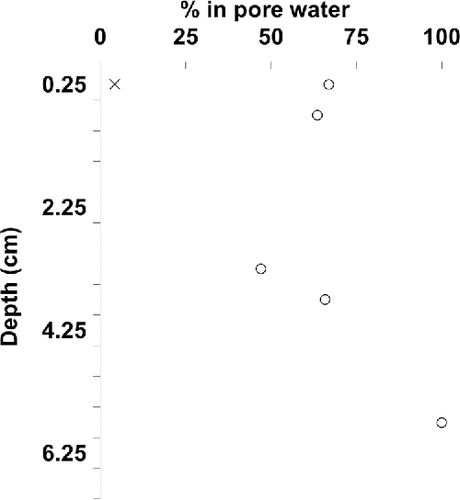
Table 1. The number of sediment intervals where microcystin congeners were detected in a 40 cm core from Hay Island, Lake of the Woods, Ontario. We analyzed a total of 80 half-cm intervals.
Microcystin congener deposition and diffusion
Microcystin concentrations and composition were measured in the water above the sediments at 3 sites in Lake of the Woods (). Averaged over the 3 sites, approximately 41% of total microcystins in the water overlying sediment was MC-LA while 59% was MC-LR. In the surficial sediment, about 11% of total microcystins was MC-LA while 89% was MC-LR. Concentrations in surficial sediments at the 3 sites in Lake of the Woods ranged from approximately 4 to 39 ng/g dw for MC-LA and 10 to 364 ng/g dw for MC-LR (). Particle sedimentation rates ranged from 0.62 to 1.64 g/m2/d. Using the surficial sediment concentrations and the particle sedimentation rates determined from 210Pb measurements, we estimated that the burial rates of 2 microcystin congeners at the 3 sites were relatively low (2.6 to 298.1 ng/m2/d), with MC-LR rates higher than MC-LA ().
Table 2. Concentrations, sedimentation rates, burial rates, and sediment–water diffusive fluxes of microcystin congeners (MC-LR and -LA) at 3 sites in Lake of the Woods: Hay Island (HI), Rat Portage Bay (RP), and Bigstone Bay (BB).
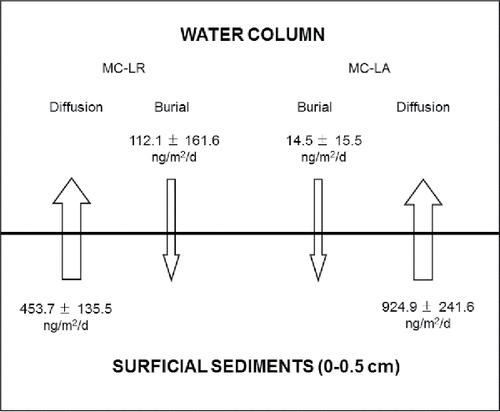
Microcystin pore water concentrations at the sediment surface (top 0.5 cm) of the 3 sites sampled were almost 3 times higher than the concentrations in the overlying water for MC-LA and almost 2 times higher for MC-LR (). Diffusive flux (from the sediment surface) at the 3 sites ranged from 646.3 to 1078.0 ng/m2/d for MC-LA and from 303.1 to 565.9 ng/m2/d for MC-LR (). Diffusive fluxes were lowest at the Hay Island site. A schematic summary of the burial rates and diffusive fluxes of microcystin congeners (MC-LR and -LA) in surficial sediments (0–0.5 cm) and at the sediment–water interface at 3 sites at the Lake of the Woods is presented ().
Figure 3. A schematic summary of the burial rates and diffusive fluxes of microcystin congeners (MC-LR and -LA; mean ± SD) in surficial sediments (0–0.5 cm) and at the sediment–water interface at 3 sites in Lake of the Woods (Hay Island, Rat Portage Bay, Bigstone Bay).
Discussion
Microcystin congener distribution in lake sediments
Sediment measurements indicated that microcystins were detectable in Lake of the Woods as early as the early 2000s (6.0–6.5 cm section in ), earlier than the first report of microcystins in surface waters in 2004 by Chen et al. (Citation2007). This is a relatively recent occurrence compared to findings for other lake studies (Latour et al. Citation2007, Pawlik-Skowronska Citation2010, Efting et al. Citation2011), including those for Lake Baptiste, Alberta, Canada, using the same methods (Zastepa et al. Citation2017b). In eutrophic Baptiste Lake, microcystins were detected at higher concentrations and throughout the entire core right to the bottom (46 cm), which corresponded to the early 1800s prior to European Settlement.
Of the 9 congeners studied, the most frequently found and in the highest concentrations in Lake of the Woods (Hay Island; ) were MC-LR and -LA. These 2 congeners were also the dominant ones found in the sediment record of Baptiste Lake (Zastepa et al Citation2017b). These 2 congeners have the highest mammalian toxicity reported for the congeners tested to date (Chorus and Bartram Citation1999, Miller et al. Citation2010). MC-LA had a tendency to distribute more into the pore water phase in comparison to MC-LR; other congeners (MC-LW, -RR, -YR, -WR, and -dmLR) were not detected in this phase in any section despite their presence on sediment particles (, ). This lower affinity of MC-LA for sediment particles has been reported in surficial sediments of other temperate lakes (Zastepa et al. Citation2015). The low affinity of MC-LA for sediment particles may be caused by electrostatic repulsion and/or high solvation resulting from the ionization (2−) of MC-LA in sediment pore waters. Newcombe et al. (Citation2003) concluded that electrostatic interactions likely play a role in sediment adsorption of MC-LA based on laboratory experiments measuring the effect of increasing ionic strength. In contrast, MC-RR and -YR had a clear tendency to adsorb to sediment particles: neither congener was detected in pore waters despite their relatively high concentrations on sediment particles. This is consistent with laboratory results indicating strong affinity of both MC-RR and -YR for sediment particles (Harada and Tsuji Citation1998, Zastepa et al. Citation2015).
Generally, a greater diversity of congeners was observed in the upper sections of Lake of the Woods (Hay Island) sediment cores. This changing congener composition may be reflective of changing toxigenic cyanobacterial communities, possibly due to recent changes in environmental conditions (Rühland et al. Citation2010, Reavie et al. Citation2017) or, alternatively, may be due to differential preservation throughout the sediment core. To more accurately interpret sedimentary archives of microcystins, quantitative studies on the natural, post-depositional decomposition of different microcystin congeners in sediments are required.
Microcystin congener deposition and diffusion in Lake of the Woods
The surficial sediment concentrations of microcystins (ng/g dw) in Lake of the Woods cores () were within the range of concentrations recently reported for other lake sediments (Babica et al. Citation2006, Chen et al. Citation2006, Zastepa et al. Citation2015). Hay Island, the shallowest of the 3 sites, had relatively high concentrations of microcystins on sediment particles, possibly due to greater exchange between surface water productivity and bottom waters at this site. In general, the microcystin sediment burial rates estimated in this study (2.6 to 298.1 ng/m2/d, based on 210Pb measurements) were low. However, they were close to MC-LR burial rates calculated by Efting et al. (Citation2011) in Nebraska using 210Pb as well as by Kankaanpaa et al. (Citation2009) in the northern Baltic Sea using sediment traps (maximum of ∼1095.2 ng/m2/d). In contrast, rates were much lower than those estimated by Wörmer et al. (Citation2011), also using sediment traps (10–2500 µg/m2/d or 365–91,250 ng/cm2/yr), for 3 Spanish reservoirs. Wörmer et al. (Citation2011) cautioned that the use of sediment traps may overestimate sedimentation rates, as being close to the lake bottom they likely also trapped resuspended microcystins. This was supported by observations in another reservoir where microcystins were found in sediment traps even though they were not detected in the water column (Wörmer et al. Citation2011). The higher sedimentation rates reported by Wörmer et al. (Citation2011) may be further explained by the widespread presence of a toxic bloom that produced high surface water concentrations of microcystins (up to 65 µg/L) during the course of their sediment trap deployment.
Concentration gradient measurements between the pore water of surficial sediments (top 0.5 cm) and the overlying water enabled the calculation of the diffusive fluxes of microcystins (). Microcystin concentrations in the surficial pore water of Lake of the Woods sediments were within the range of concentrations reported in a survey of 7 Canadian lakes (500–130,000 ng/L; Zastepa et al. Citation2015). The diffusive flux estimates () suggest that a return of microcystins to the water column may be important in lakes, as the rates observed exceed diffusive flux estimates for some contaminants (e.g., mercury) at polluted sites (Delongchamp et al. Citation2010). Furthermore, diffusion of microcystins from sediments exceeded microcystin sediment burial rates. The ratio of diffusive flux to sediment burial rate indicates the proportion of deposited microcystins that is remobilized from the sediments. The observation that microcystins are remobilized at a higher rate than they are buried in sediments suggests sediments can act as a source of microcystins, although the duration of this is unknown since only 1 sampling was undertaken in this study. The release of microcystin may be the result of dissolution or the active production of microcystins within sediments. Several studies have found microcystins in samples of benthic cyanobacteria; however, active production within sediments has not been confirmed (Mez et al. Citation1997, Hitzfeld et al. Citation2000, Izaguirre et al. Citation2007). The most compelling evidence of active production within sediment was reported by Misson et al. (Citation2012) who recorded mcyB RNA in every benthic Microcystis population investigated, even in those buried for 6 yr in sediments. The presence of mcyB RNA indicates the ability to initiate microcystin production and is consistent with the high remobilization observed in this study. However, differences between burial rates and diffusive flux could also be the result of a temporal disconnect between water column productivity and sediment processes (timing of surface blooms, lysis of bloom deposits, pore water mobility of microcystins) and the relatively high diffusion could be limited to only part of the summer following bloom events. These are the first estimates of diffusive flux of microcystins from sediments and additional measurements over time, among lakes, and using different methods (e.g., peepers) should be obtained. Furthermore, it is important to note that these flux estimates are diffusive fluxes and do not consider additional fluxes caused by bioturbation, bioirrigation, and resuspension of sediments. As such, these flux estimates based solely on molecular diffusion may be underestimated.
MC-LR had a higher sediment burial rate than MC-LA and this was reflected in the higher proportion of MC-LR (of total microcystins) in the surficial sediments compared to the water above the sediments. In contrast, a higher proportion of MC-LA (of total microcystins) was found in the water above the sediments compared to the surficial sediments. Evidence from laboratory experiments similarly suggests that different congeners may have different sedimentary fate especially due to differential adsorption to sedimentary particles (Harada and Tsuji Citation1998, Newcombe et al. Citation2003, Zastepa et al. Citation2015). In contrast, field observations thus far have found no differences in congener composition between the water column and sediments (Welker et al. Citation2007). However, this could be because microcystins preferentially deposit into sediments within intact cells (Wörmer et al. Citation2011) and, therefore, the adsorption of dissolved microcystins to settling particles may be relatively minor. When high levels of dissolved microcystins are present during toxic bloom senescence, differential sedimentary fate can be observed: in a study of the fate of microcystins in a small eutrophic lake with toxic Microcystis blooms, MC-RR showed a greater tendency to deposit into sediments in comparison to MC-LA which persisted in the water column (Zastepa et al. Citation2014).
Diffusion of microcystins from sediments represents a potential risk for human exposure in the absence of visible surface blooms. Diffusion of microcystins from sediments may affect drinking water supplies that withdraw from deep waters. Particularly in shallow systems, sediments may represent a previously unrecognized route of exposure to dissolved microcystins in the absence of blooms. Risk management strategies, therefore, cannot simply rely on macroscopic (bloom) or microscopic identification of potentially toxigenic species of cyanobacteria; testing for microcystins may be necessary even in the absence of bloom episodes particularly in lakes with a history of toxigenic blooms. The higher diffusive flux and lower sediment burial rate of MC-LA compared to MC-LR suggest a higher remobilization of MC-LA into the waters overlying sediments. These findings underscore the importance of congeners-specific assessment of environmental fate and exposure risk of microcystins.
Acknowledgments
We would like to thank Todd Sellers of the Lake of the Woods Water Sustainability Foundation and Andrew Paterson of the Ontario Ministry of the Environment and Climate Change for facilitating this research. A special thank you to Linda E. Kimpe and Ammar Saleem for their guidance and analytical support as well as Crystal Hyatt, Ron Ingram, and Gavin Olson for their tireless assistance and guidance in the field.
References
- Appleby PG. 2001. Chronostratigraphic techniques in recent sediments. pp. 171–203. In: Last WM, Smol JP (eds.) Tracking environmental change using lake sediments, volume 1. Basin analysis, coring, and chronological techniques. Kluwer Academic Publishers: Dordrecht (Netherlands).
- Appleby PG, Oldfield F. 1978. The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210Pb to the sediment. Catena 5:1–8.
- Babica P, Kohoutek J, Bláha L, Adamovský O, Marsalek B. 2006. Evaluation of extraction approaches linked to ELISA and HPLC for analyses of microcystin-LR, -RR and -YR in freshwater sediments with different organic material contents. Anal Bioanal Chem. 385:1545–1551.
- Binford MW. 1990. Calculation and uncertainty analysis of 210Pb dates for PIRLA project lake sediment cores. J Paleolimnol. 3:253–267.
- Blais JM, Kalff J, Cornett RJ, Evans RD. 1995. Evaluation of 210Pb dating in lake sediments using stable Pb, Ambrosia pollen, and 137Cs. J Paleolimnol. 13:169–178.
- Boudreau BP. 1996. The diffusive tortuosity of fine-grained unlithified sediments. Geochim Cosmochim Ac. 60:3139–3142.
- Brady NC, Weil RR. 2002. The nature and properties of soils, 13th ed. New York (NY): Prentice Hall.
- Chen H, Burke JM, Dinsmore WP, Prepas EE, Fedorak PM. 2007. First assessment of cyanobacterial blooms and microcystin-LR in the Canadian portion of Lake of the Woods. Lake Reserv Manage. 23:169–178.
- Chen W, Li L, Gan N, Song L. 2006. Optimization of an effective extraction procedure for the analysis of microcystins in soils and lake sediments. Environ Pollut. 143:241–246.
- Chorus I, Bartram J. 1999. Toxic cyanobacteria in water: a guide to their public health consequences, monitoring and management. London and New York: World Health Organization, E & FN Spon.
- Delongchamp T, Ridal J, Lean D, Poissant L, Blais JM. 2010. Mercury transport between sediments and the overlying water of the St. Lawrence River area of concern near Cornwall, Ontario. Environ Pollut. 158:1487–1493.
- Edlund MB, Schottler SP, Reavie ED, Engstrom DR, Baratono NG, Leavitt PR, Heathcote AJ, Wilson B, Paterson AM. 2017. Historical phosphorus dynamics in Lake of the Woods (USA-Canada) – does legacy phosphorus still affect the southern basin? Lake Reserv Manage. 33:386–402.
- Efting A, Snow D, Fritz S. 2011. Cyanobacteria and microcystin in the Nebraska (USA) Sand Hills Lakes before and after modern agriculture. J Paleolimnol. 46:17–27.
- Glew JR. 1989. A new trigger mechanism for sediment samplers. J Paleolimnol. 2:241–243.
- Harada KI, Tsuji K. 1998. Persistence and decomposition of hepatotoxic microcystins produced by cyanobacteria in natural environment. Toxin Rev. 17:385–403.
- Hitzfeld BC, Lampert C, Spaeth N, Mountfort D, Kaspar K, Dietrich D. 2000. Toxin production in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Toxicon. 38:1731–1748.
- Izaguirre G, Jungblut A, Neilan BA. 2007. Benthic cyanobacteria (Oscillatoriaceae) that produce microcystin-LR, isolated from four reservoirs in southern California. Water Res. 41:492–498.
- Johnston WA. 1915. Rainy River District, Ontario: surficial geology and soils. Survey of Canada, Geological Series Report No. 68.
- Kankaanpaa HT, Sjovall O, Huttunen M, Olin M, Karlsson K, Hyvarinen K, Sneitz L, Harkonen J, Sipia VO, Meriluoto JA. 2009. Production and sedimentation of peptide toxins nodularin-R and microcystin-LR in the northern Baltic Sea. Environ Pollut. 157:1301–1309.
- Latour D, Salencon M, Reyss J, Giraudet H. 2007. Sedimentary imprint of Microcystis aeruginosa (cyanobacteria) blooms in Grangent reservoir (Loire, France). J Phycol. 43:417–425.
- Meriluoto J, Spoof L, Codd GA, (eds.). 2017. Handbook of cyanobacterial monitoring and cyanotoxin analysis. John Wiley & Sons: West Sussex, UK.
- Mez K, Beattie KA, Codd GA, Hanselmann K, Hauser B, Naegeli H, Preisig H. 1997. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur J Phycol. 32:111–117.
- Miller MA, Kudela RM, Mekebri A, Crane D, Oates SC, Tinker MT, Staedler M, Miller WA, Toy-Choutka S, Dominik C, Hardin D, Langlois G, Murray M, Ward K, Jessup DA. 2010. Evidence for a novel marine harmful algal bloom: cyanotoxin (microcystin) transfer from land to sea otters. PLOS ONE. 5(9):e12576.
- Misson B, Sabarta M, Amblard C, Latour D. 2012. Benthic survival of Microcystis: long-term viability and ability to transcribe microcystin genes. Harmful Algae. 13:20–25.
- Newcombe G, Cook D, Brooke S, Ho L, Sylman N. 2003. Treatment options for microcystin toxins: similarities and differences between variants. Environ Technol. 24:299–308.
- Pawlik-Skowronska B, Kornijow R, Pirszel J. 2010. Sedimentary imprint of cyanobacterial blooms – a new tool for insight into recent history of lakes. Pol J Ecol. 58:663–670.
- Pla S, Paterson AM, Smol JP, Clark BJ, Ingram R. 2005. Spatial variability in water quality and surface sediment diatom assemblages in a complex lake basin: Lake of the Woods, Ontario, Canada, J Great Lakes Res. 31:253–266.
- Reavie E, Edlund M, Andresen N, Engstrom D, Leavitt P, Schottler S, Cai M. 2017. Paleolimnology of the Lake of the Woods southern basin: continued water quality degradation despite lower nutrient influx. Lake Reserv Manage. 33:369–385.
- Rühland K, Paterson AM, Hargan K, Jenkin A, Clark BJ, Smol JP. 2010. Reorganization of algal communities in the Lake of the Woods (Ontario, Canada) in response to turn-of-the-century damming and recent warming. Limnol Oceanogr. 55:2433–2451.
- Rusak JA, Mosindy T. 1997. Seasonal movements of lake sturgeon in Lake of the Woods and the Rainy River, Ontario. Can J Zool. 74:383–395.
- Schupp DH, Macins V. 1977. Trends in percid yields from Lake of the Woods, 1888–1973. J Fish Res Board Can. 34:1784–1791.
- Schwarzenbach RP, Gschwend PM, Imboden DM. 2003. Environmental organic chemistry, 2nd ed. New York (NY): John Wiley & Sons.
- Serieyssol C, Edlund M, Kallemeyn L. 2009. Impacts of settlement, damming, and hydromanagement in two boreal lakes: a comparative paleolimnological study. J Paleolimnol. 42:497–513.
- Serieyssol-Bleser C. 2010. Determining the impacts of damming, water-level fluctuations, climate, and landscape changes in Voyageurs National Park and vicinity. [dissertation]. [Minneapolis (MN)]: University of Minnesota.
- Waters MN. 2016. A 4700-year history of cyanobacteria toxin production in a shallow subtropical lake. Ecosystems. 19:426–436.
- Watson SB, Kling H. 2017. Lake of the Woods phyto- and picoplankton: spatiotemporal patterns in blooms, community composition and nutrient deficiency. Lake Reserv Manage. 33:415–432.
- Welker M, Sejnohova L, Nemethova D, Dohren H, Jarkovsky J, Marsalek B. 2007. Seasonal shifts in chemotype composition of Microcystis sp. communities in the pelagial and the sediment of a shallow reservoir. Limnol Oceanogr. 52:609–619.
- Wörmer L, Cirés S, Quesada A. 2011. Importance of natural sedimentation in the fate of microcystins. Chemosphere. 82:1141–1146.
- Yang Z, Teller J. 2005. Modeling the history of Lake of the Woods since 11,000 cal yr B.P. using GIS. J Paleolimnol. 33:483–497.
- Zastepa A, Pick FR, Blais JM. 2014. Fate and persistence of particulate and dissolved microcystin-LA from Microcystis blooms. Hum Ecol Risk Assess. 20:1670–1686.
- Zastepa A, Pick FR, Blais JM, Saleem A. 2015. Analysis of intracellular and extracellular microcystin variants in sediments and pore waters by accelerated solvent extraction and high performance liquid chromatography-tandem mass spectrometry. Anal Chim Acta. 872:26–34.
- Zastepa A, Watson SB, Kling H, Kotak B. 2017a. Spatial and temporal patterns in microcystin toxins in Lake of the Woods surface waters. Lake Reserv Manage. 33:433–443.
- Zastepa A, Taranu ZE, Kimpe LE, Blais JM, Gregory-Eaves I, Zurawell RW, Pick FR. 2017b. Reconstructing a long-term record of microcystins from the analysis of lake sediments. Sci Total Environ. 579:893–901.
