
Abstract
Sass GG, Shaw SL, Rooney TP, Rypel AL, Raabe JK, Smith QC, Hrabik TR, Toshner ST. 2019. Coarse woody habitat and glacial lake fisheries in the Midwestern United States: knowns, unknowns, and experiment to advance our knowledge. Lake Reserv Manage. 35:382–395.
Coarse woody habitat (CWH) additions have increased in popularity in glacial lakes (i.e. kettle lakes) of the Midwestern United States. However, most enhancements have not been treated as deliberate experiments to test for fish and aquatic ecosystem responses. Whole-lake CWH removal studies have shown reductions in fish growth rates, declines in forage fish abundance, and behavioral changes. Whole-lake CWH addition studies have shown improved reproductive output of certain fishes, increased availability and diversity of prey available to fishes, and influenced behavior and habitat use. Key uncertainties identified in previous CWH addition studies include: (1) Does CWH increase fish production? (2) Does CWH influence certain fish species differently? (3) Does CWH influence fish populations in larger lakes than previously studied? (4) Does CWH influence fish populations over longer periods of time? In 2015, we began a whole-lake CWH addition experiment on a northern Wisconsin lake aimed to address these uncertainties. Sanford Lake maintains a low productivity fish community and supports fishes not generally studied before in the context of CWH. Fish population dynamic/behavior and aquatic ecosystem response variables will be monitored, and tree drop CWH additions are slated for 3 phases over 20 yr. We introduce the Sanford Lake experiment and provide recommendations for expectations and the implementation of CWH additions in inland glacial lakes. Given the reliance of north-temperate inland glacial lake fisheries on allocthonous sources of energy and negative influences of lakeshore residential development on CWH, we hypothesize that CWH addition may contribute to maintaining or enhancing fish production.
Availability of suitable habitat is critical for the sustainability of fish and wildlife resources (Sass et al. Citation2017). Coarse woody habitat (CWH) (i.e., sticks, branches, and trees in the littoral zones of waterbodies) is a natural feature of many inland glacial lakes in the upper Midwestern United States. It is a critical source of nutrients, detritus, and fish habitat, particularly in aquatic ecosystems depauperate in alternative structural habitats (e.g., low availability of aquatic macrophytes, gravel, cobble, boulders; Sass, Kitchell, et al. Citation2006, Sass Citation2009, Gaeta et al. Citation2014, Czarnecka Citation2016, Sass et al. Citation2017). From an aquatic food web perspective, CWH degradation provides nutrients and a substrate for periphyton growth (Lebkuecher et al Citation1998, Keshavanath et al. Citation2001, Vadeboncoeur and Lodge Citation2000, Vadeboncoeur et al. Citation2006). This in turn attracts benthic invertebrates (Benke et al. Citation1985, Bowen et al. Citation1998, Smokorowski et al. Citation2006), and these provide a food source to fishes (Schindler and Scheuerell Citation2002, Sass et al. Citation2012). Coarse woody habitat has also been shown to provide refuge from predators (Everett and Ruiz Citation1993, Sass, Gille, et al. Citation2006, Sass et al. Citation2012), serve as a spawning substrate or attractant to spawning fishes (Becker Citation1983, Hunt and Annett Citation2002, Wills et al. Citation2004, Sass, Kitchell, et al. Citation2006, Lawson et al. Citation2011, Weis and Sass Citation2011), and influence fish growth rates (Schindler et al. Citation2000, Sass, Kitchell, et al. Citation2006, Gaeta et al. Citation2011, Citation2014). Presence/absence, abundance, and branching complexity of CWH strongly influence habitat partitioning in fishes (Newbrey et al. Citation2005, Sass et al. Citation2012) and fish behavior (Scheurell and Schindler Citation2004). For example, Ahrenstorff et al. (Citation2009) found that largemouth bass (Micropterus salmoides) change foraging behavior between sit-and-wait and actively foraging depending on the amount of littoral CWH. In oligotrophic, north-temperate inland glacial lakes, allocthonous sources of energy (including leaf litter and CWH) contributed to a substantial proportion of fish production (Pace et al. Citation2004). Because leaf litter provides annual terrestrial inputs of carbon and CWH can persist in aquatic ecosystems for hundreds of years (France Citation1997, Guyette and Cole Citation1999, Glaz et al. Citation2009, Gennaretti et al. Citation2014), reductions in terrestrial inputs of carbon over time have the potential to negatively influence fish production.
Recognition of CWH as potentially important for fish habitat started with Everett and Ruiz (Citation1993) in a Chesapeake Bay estuary and later by Christensen et al. (Citation1996), who found a strong, negative correlation between littoral CWH abundance and lakeshore residential development on northern Wisconsin and upper Michigan inland lakes. This pattern was independently identified for Pacific Northwest lakes, and in broader datasets from northern Wisconsin and upper Michigan (Jennings et al. Citation2003, Francis and Schindler Citation2006, Marburg et al. Citation2006). Subsequent studies identified physical removals of CWH by lakeshore residents and thinning of the riparian forest as primary mechanisms driving these relationships (Sass Citation2009). Forest fire suppression efforts, succession of the riparian forest itself (early successional vs. late successional trees), and beaver (Castor canadensis) control have also influenced littoral CWH abundances and new inputs (Roth et al. Citation2007, Sass Citation2009). Size-specific growth rates of largemouth bass and bluegill (Lepomis macrochirus) have been shown to be positively correlated with CWH abundances and can be life stage specific (Schindler et al. Citation2000, Sass, Kitchell, et al. Citation2006, Gaeta et al. Citation2011, Citation2014). Further, whole-lake experiments have simulated the lakeshore residential development process by physically removing CWH and testing for periphyton, benthic macroinvertebrate, and fish community responses (Sass, Gille, et al. Citation2006, Smokorowski et al. Citation2006). Reciprocal experiments of adding CWH have tested whether these additions can mitigate negative effects of CWH loss (Sass et al. Citation2012).
Fish structural habitat enhancements using CWH (e.g., tree drops, “fish sticks”) in inland glacial lakes and reservoirs are becoming a common practice for many agencies, lake associations, and stakeholder groups. Although the outcomes of these practices are somewhat known (i.e., fish and angler attraction), goals of the enhancement projects are often not formally evaluated. Moreover, most fish structural habitat enhancements are not treated as deliberate experiments (i.e., pre- and postmanipulation monitoring, before-after-control-impact experimental designs, or reference system(s); Stewart-Oaten et al. Citation1986, Carpenter Citation1998). Therefore, uncertainty surrounds the utility of such fish structural habitat enhancement practices. This warrants a review of previous research and the identification of key uncertainties. Our objectives were to (1) review case studies of CWH removals and additions in inland glacial lakes of the upper Midwestern United States; (2) highlight key uncertainties derived from these case studies and introduce a long-term, whole-lake experiment aimed to address these uncertainties; and (3) discuss implementation considerations for CWH enhancements and associated expectations based on current knowledge. Although our review is focused on CWH and inland glacial lake fisheries in the upper Midwestern United States, our discussion is also pertinent to streams, rivers, reservoirs, and other natural lakes and their associated fisheries where similar uncertainties exist (Bassett Citation1994, Gregory et al. Citation2003, Sass Citation2009).
Whole-lake CWH removals
To our knowledge, inland glacial lake water quality and lower trophic level responses to CWH removals are limited to a few whole-lake experimental and modeling studies. Perhaps the most comprehensive study testing for lower trophic level responses, Smokorowski et al. (Citation2006) assessed water quality, periphyton, and macroinvertebrate responses to 40–70% reductions of CWH in 3 Ontario lakes. They found no change in water quality, and that chlorophyll a (Chl-a) concentration in periphyton was greater on highly decayed CWH compared to new CWH. Although macroinvertebrate biomass was greater on CWH, removal resulted in a low net loss of total macroinvertebrate productivity because CWH availability was low compared to available sediment (Smokorowski et al. Citation2006). A 70% reduction of CWH in a northern Wisconsin lake resulted in no difference in benthic macroinvertebrate community composition, diversity, and density between the treatment and reference basins (Helmus and Sass Citation2008). Although that study was conducted over a relatively short time frame, rapid effects of CWH removal on benthic macroinvertebrates were not evident and likely supported the findings of Smokorowski et al. (Citation2006). In a modeling exercise, Roth et al. (Citation2007) linked forage and piscivorous fish biomass to a strong positive correlation between CWH abundance and benthic macroinvertebrate biomass (Benke et al. Citation1985, Bowen et al. Citation1998) such that removal of CWH resulted in declines in fish biomass through this pathway. Although CWH has been shown to support benthic macroinvertebrate production (Benke et al. Citation1985, Bowen et al. Citation1998, Smokorowski et al. Citation2006), uncertainties still exist as to whether CWH loss negatively influences upper trophic level production significantly.
During the 2000s, whole-lake CWH removals were conducted on a northern Wisconsin lake to test for fish community responses under two different mechanisms of removal. Sass, Kitchell, et al. (Citation2006) tested for fish community responses to a rapid (∼8 weeks) physical removal of littoral CWH (∼70%) in the treatment basin of Little Rock Lake, Vilas County, Wisconsin. This lake was closed to public access and fishing and curtained into reference and treatment basins for experimental purposes. Gaeta et al. (Citation2014) tested for fish community responses to changes in CWH on the reference basin of Little Rock Lake. A prolonged drought reduced water levels by about 1 m, naturally removing available CWH by about 75%. Whole-lake experiments are often criticized for a lack of replication (Carpenter Citation1998). Interestingly, two of the primary fish community responses were identical between the experiments, suggesting the utility of whole-lake experiments for inference. Sass, Kitchell, et al. (Citation2006) and Gaeta et al. (Citation2014) observed functional collapses of the yellow perch (Perca flavescens) populations due to predation pressure from largemouth bass and the loss of refuge and spawning habitat for yellow perch. Similar findings were documented in a tethering study of fathead minnow (Pimephales promelas), Lepomis spp., and yellow perch across a gradient of lakes/lake basins differing in littoral zone aquatic macrophyte and CWH abundances in northern Wisconsin and upper Michigan (Sass, Gille, et al. Citation2006). Size-specific growth rates of largemouth bass declined in both studies after CWH removal. Largemouth bass diets were dominated by terrestrial sources of prey, and largemouth bass growth and attainment of harvestable lengths were positively correlated with CWH abundance (Sass, Kitchell, et al. Citation2006, Gaeta et al. Citation2014). Mechanisms causing the functional collapses of the yellow perch populations were through the direct loss of submerged CWH as refuge and spawning substrate in Sass, Kitchell, et al. (Citation2006), and through the desiccation of CWH and concentration of predator-prey interactions with reduced lake levels in Gaeta et al. (Citation2014).
Whole-lake CWH additions
Although agencies, lake associations, and stakeholder groups continue to use CWH additions in inland glacial lakes and reservoirs (e.g., tree drops, fish cribs, “fish sticks,” Christmas tree bundles) to “enhance” fish habitat, most projects have not been treated as deliberate learning experiments. Seldom is an unmanipulated reference system used. Baseline fish community attributes are measured neither before CWH additions, nor for a sufficient period after to test for responses (Carpenter Citation1998). Therefore, key uncertainties exist in the response of fish communities and anglers to these practices depending on the type of waterbody (inland glacial lake vs. reservoir), type of habitat enhancement, and fish species. It is well established that many fishes are attracted to CWH (Newbrey et al. Citation2005, Pratt et al. Citation2005, Sass, Gille, et al. Citation2006, Sass et al. Citation2012); however, structural habitats may also attract anglers and increase site-specific exploitation rates. It is uncertain whether CWH enhancements result in increased fish production to offset potential increases in angler exploitation.
The only comprehensive, whole-lake CWH addition study to our knowledge was conducted by Sass et al. (Citation2012). They added CWH to a northern Wisconsin lake and tested for fish community, population, and behavioral responses. Camp Lake, Vilas County, Wisconsin, is naturally separated into 2 basins by a small channel and fish movement between basins was low during premanipulation monitoring. Prior to ice-out in 2004, one piece of CWH (i.e., tree tops that varied by species, structural complexity, and size) was added per 10 m of shoreline on the treatment basin, while the reference basin remained unaltered. Additionally, two 100 m sections of littoral zone were also treated with one piece of CWH per 5 m of shoreline to test for fish behavioral and habitat use responses across a gradient of CWH abundance (0, 1, and 2 pieces of CWH per 10 m of shoreline). Fish population responses to the whole-lake addition of CWH were not as pronounced as fish behavioral responses. Largemouth bass and bluegill abundance and growth responses were coherent among basins suggesting no effect of the manipulation. However, proportional size distribution indices declined for both species, suggesting increased reproductive output. The terrestrial component of largemouth bass diets decreased in the treatment basin (reciprocal response to the Little Rock Lake experiment), and diet breadth and the percentage of empty stomachs declined, suggesting greater prey availability. Areas of higher CWH abundance and complexity showed greater habitat use by largemouth bass and bluegill. Addition of CWH resulted in a decline in home range size of individual largemouth bass, suggesting a switch from an active to a sit-and-wait foraging strategy (Ahrenstorff et al. Citation2009). A key uncertainty from the Sass et al. (Citation2012) study was that the fish community was only monitored for 4 yr post CWH addition, due to declining water levels on Camp Lake resulting from a prolonged drought in northern Wisconsin (Gaeta et al. Citation2014). In general, postmanipulation monitoring of fishes following perturbations should encompass the life span of at least one cohort. Bluegill and largemouth bass in northern Wisconsin may live up to 10 and 20 yr, respectively (Carlander Citation1977). Therefore, it is uncertain whether fish population responses might have become evident if Camp Lake were monitored for a longer period post CWH addition.
Uncertainties and an experiment to advance our knowledge
Previous CWH removal and addition experiments have been conducted on small inland glacial lakes (8–18 ha), in low-diversity fish communities (largemouth bass, yellow perch/bluegill), and monitored for relatively short periods post manipulation (Sass, Kitchell, et al. Citation2006, Sass et al. Citation2012). Key uncertainties identified in previous CWH addition experiments include: (1) Does CWH only attract fishes? (2) Does CWH increase fish production? (3) Does CWH influence certain fish species differently? (4) Does CWH influence fish populations in larger lakes than previously studied? (5) How does CWH influence fish populations over longer periods of time? (6) Does CWH addition offset the potential for greater angler exploitation by increasing fish production? Here, we introduce an ongoing long-term, whole-lake experiment aimed to address these key uncertainties. We also test for potential responses of terrestrial biota (e.g., riparian-zone web-building spiders) in the ecotone to the within-lake habitat additions.
Study site
Sanford Lake, Vilas County, Wisconsin (46°18'N, 89°69'W), is a mesotrophic, 37 ha inland glacial lake with a maximum depth of 16 m, mean depth of 4.8 m, and a mean Secchi disc transparency of 2.6 m (, ). The property surrounding Sanford Lake is owned by Dairymen’s Inc. The shoreline is undeveloped, and access to the lake is restricted to Dairymen’s Inc., members only. The lake is closed to public access and recreational fishing. The fish community of Sanford Lake includes naturally reproducing populations of walleye (Sander vitreus), muskellunge (Esox masquinongy), smallmouth bass (Micropterus dolomieu), yellow perch, bluegill, rock bass (Ambloplites rupestris), pumpkinseed (Lepomis gibbosus), largemouth bass (Micropterus salmoides), bluegill × pumpkinseed hybrids (Lepomis macrochirus × L. gibbosus), black bullhead (Ameiurus melas), grass pickerel (Esox americanus vermiculatus), golden shiner (Notemigonus crysoleucas), green sunfish (Lepomis cyanellus), creek chub (Semotilus atromaculatus), white sucker (Catostomus commersonii), black crappie (Pomoxis nigromaculatus), burbot (Lota lota), and common shiner (Luxilus cornutus) (). Dairymen’s Inc., member angling regulations for walleye and panfish on Sanford Lake follow Wisconsin statewide regulations for these species. Dairymen’s Inc., members are not allowed to harvest muskellunge or largemouth and smallmouth bass. They also maintain a compulsory creel census of all fish harvested from Sanford Lake.
Figure 1. Map of the locations of Sanford and Escanaba lakes (dot), Vilas County, Wisconsin (shaded), in relation to the United States and North America (A), along with satellite images of Sanford Lake (B) and Escanaba Lake (C).
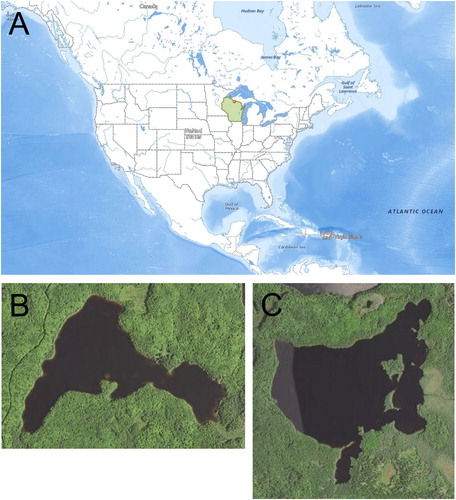
Table 1. Morphometric and chemical and physical limnological variables and fish species presence/absence in Sanford and Escanaba lakes, Vilas County, Wisconsin. An “X” associated with a fish species and lake denotes presence.
Materials and methods
In 2015, Dairymen’s Inc., and the Wisconsin Department of Natural Resources (DNR) partnered to begin a long-term, whole-lake tree drop experiment on Sanford Lake to address key uncertainties related to CWH additions in inland glacial lakes (outlined in the preceding), with the primary goal of testing for fish production responses. Key considerations of whole-lake experiments include having a reference system and pre- and post-manipulation monitoring for a sufficient period to detect effects (Carpenter Citation1998). Escanaba Lake, Vilas County, Wisconsin (46°065'N, 89°58'W), was selected as the reference lake for the study (Carpenter Citation1998). Escanaba Lake (119 ha, maximum depth = 7.9 m, mean depth = 4.3 m) is undeveloped, has a similar fish community and water quality (mean Secchi disc transparency = 3.3 m), and maintains a compulsory creel census (). However, it is about 3 times greater in surface area than Sanford Lake (Kempinger and Carline Citation1977). Pre-manipulation monitoring of the fish communities and aquatic ecosystems of Sanford and Escanaba lakes began in spring 2015, with the intent to add tree drops to one quarter of Sanford Lake commensurate with CWH abundances observed in undeveloped lakes (Christensen et al. Citation1996, Marburg et al. Citation2006, ∼ 400 logs/km of shoreline). In June 2018, a professional logger dropped 140 trees into Sanford Lake along the north shore (). Tree species dropped into the lake were commensurate with riparian forest availability based on a previous survey (∼87% coniferous, ∼13% deciduous) (Rooney TP, Wright State University, July 2017, unpublished data). Tree drops were not cabled to the stump or shoreline, and two 100 m stretches of shoreline were left unaltered because they were known walleye spawning habitat. Post-manipulation monitoring of Sanford and Escanaba lakes will continue for 5 yr (2018–2022). This will be followed by a second round of tree drops on the west shore of Sanford Lake (2023), another 5 yr of monitoring (2023–2027), and then a third round of tree drops on the remainder of Sanford Lake (2028), with the final monitoring period continuing until 2032. Tree drop manipulations were staggered for two reasons: (1) to test for fish production saturation, such that further addition of tree drops does not increase fish production, and (2) because many inland glacial lakes may only have a limited amount of riparian area where trees may be dropped due to lakeshore residential development. Note, however, that “fish sticks” are trees brought into waterbodies from upland sources and are the preferred CWH fish structural habitat enhancement method used by the Wisconsin DNR because this maintains riparian forest integrity and this source of natural CWH input to inland lakes. Tree drops in lieu of “fish sticks” were used on Sanford Lake at the request of Dairymen’s Inc. Although tree drops and “fish sticks” come from different forest sources, they are functionally equivalent as fish habitat.
Figure 2. Photographs of a white pine (Pinus strobus) being dropped into Sanford Lake, Wisconsin (upper), and tree drops along the northern shore of the lake (lower) in June, 2018.

Fish community response variables measured annually (Apr–Oct) in Sanford Lake (and similarly for Escanaba Lake) include mark-recapture population estimates for the major fish species (Ricker Citation1975), species-specific demographic information (length, weight, sex, age, growth, body condition, proportional size distribution) (Zale et al. Citation2012), assimilated diets based on stable isotope analyses (Boecklen et al. Citation2011, Layman et al. Citation2012, Glibert et al. Citation2018), fish production (Rypel et al. Citation2015, Citation2018), and fish behavior. Annual mark-recapture population estimates are conducted in spring (or fall, age-0 and age-1 walleye, age-0 muskellunge) immediately after ice-out using fyke nets as a marking and recapture gear and AC electrofishing as an additional recapture gear. The Chapman-modified, continuous Schnabel estimator is used for all population estimates (Ricker Citation1975). All captured fishes are measured for total length (mm), weighed (g), sexed (by expression of gametes), have an aging structure removed (scale or dorsal fin spine on walleye ≥ 508 mm), and are given a unique mark. Small fishes are marked with a year-specific permanent fin clip, and those in a subsample are implanted with a 12 mm passive integrated transponder (PIT) tag inserted near the pelvic girdle. Larger fishes are marked with individually numbered Floy tags and implanted with a 32 mm PIT tag inserted near the pelvic girdle. Pre- and post-manipulation fish production estimates (biomass, production [kg/ha/yr], and production:biomass ratios [P/B]) will be calculated for several fish species in Sanford and Escanaba lakes using methods described in Rypel et al. (Citation2015, Citation2018). Sanford Lake currently supports a low-productivity fish community like other northern Wisconsin lakes. We hypothesize that an increase in fish production with CWH addition will manifest through bottom-up effects of increases in lower trophic level production (periphyton, benthic macroinvertebrates), transferring energy to mid-trophic-level forage fishes (yellow perch, bluegill, rock bass) and then to upper trophic level (walleye, muskellunge, smallmouth bass) production.
Fish behavior and habitat use in Sanford Lake are monitored using 2 methods. First, standard radio telemetry is being used to monitor lake-wide behavior of muskellunge, walleye, and smallmouth bass. Our radio-telemetry monitoring is restricted to larger fishes due to the size of the tags and associated 2–3 yr life of the battery. Due to tag loss, extinction of battery life, and mortality, new radio tags have been implanted in additional fish every year. Second, we have adapted PIT tag receivers that are generally used to monitor fish movements in lotic environments to passively monitor littoral zone habitat use of fishes in Sanford Lake. Ten PIT tag receivers are currently deployed around the littoral zone of Sanford Lake at relatively equal distances apart. Each PIT tag receiver consists of a receiver, two tuner boxes, and two 1 × 10 m rectangular antennae that rest on the bottom of the lake. When a PIT-tagged fish passes within the detection distance of an antenna (up to 0.3 m for 12 mm tags and up to 1.0 m for 32 mm tags), the PIT tag number and time are recorded by the receiver, and direction of travel can be established if the fish passes over both antennae. Each receiver is downloaded twice per week. Passive monitoring with PIT tag receivers will allow us to test whether littoral zone fish habitat use changes with the addition of CWH.
In addition to the fish response variables, numerous aquatic ecosystem and ecotone variables are being monitored in Sanford Lake because they have the potential to be influenced by CWH addition, with identical variables also being monitored on Escanaba Lake as the reference system. A standard suite of limnological variables (Secchi disk transparency, water temperature/dissolved oxygen profile, surface Chl-a) are measured at the 2 deep holes of Sanford Lake biweekly. In association with limnological sampling, entire-water-column zooplankton tows (80 µm) are also collected. In 2015, and biweekly in 2018, fast limnology and measurement (FLAMe) has been used to measure real-time surface limnological attributes of Sanford Lake (Crawford et al. Citation2015). Deuterium (2H) stable isotope analysis is being used to assess allochthonous energy inputs that may change because of CWH addition (Batt et al. Citation2012, Citation2015). Littoral-zone benthic macroinvertebrates are collected monthly at various fixed sites using a kick net. Sonar surveys are used to create bathymetric maps, measure bottom hardness, and estimate submersed aquatic vegetation biovolume annually at the peak of the growing season (August) (). Given our hypothesis that CWH addition will increase carbon input to Sanford Lake, we are also measuring several terrestrial ecotone response variables to test whether aquatic energy input also results in energy output to riparian zone organisms through emergent insects. Therefore, we are measuring pre- versus post-CWH-addition relative abundances of web-building spiders, and quantifying diets through stable isotope analysis of 13C, 15N, and 2H. Lastly, angler effort dynamics and exploitation in relation to the CWH addition are being monitored by the compulsory creel census maintained by Dairymen’s Inc.
Figure 3. Bathymetric maps of Sanford (A) and Escanaba (B) lakes, Vilas County, Wisconsin with darker shading denoting greater bottom depth in meters.

Remaining key uncertainties regarding inland glacial lake fish structural habitat enhancements, and a ripe area for future research, relate to tests of similarities or differences in functions and outcomes of CWH additions versus fish cribs (i.e., wooden-log cabin structures submerged in deeper water) versus artificial habitat structures. The current preference of the Wisconsin DNR is to use “fish sticks” for inland glacial lake fish structural enhancements. Although more popular in the past, fish cribs have generally fallen out of favor as a structural enhancement tool in Wisconsin inland glacial lakes. Still, questions remain as to whether fish cribs serve a similar or different function to “fish sticks” or tree drops. Littoral zone structural habitat enhancements are generally preferred because many fishes are co-adapted with the availability of CWH in north-temperate inland glacial lake littoral zones. In contrast, fish cribs provide an unnatural structural element in deeper portions of the lake. It is well established that isolated fish cribs attract fish and anglers; however, it is unclear whether fish cribs benefit fish populations, and if there are benefits, whether they offset potential increases in directed effort and exploitation by anglers. Future studies should test whether isolated fish cribs only attract adult fishes, influence mortality rates on juvenile fishes seeking pelagic refuge, and/or increase angler effort and exploitation rates. Given these uncertainties, multiple fish cribs should be aggregated when used to simulate the complexity of natural rock bars or submersed aquatic macrophyte beds, which may in turn diffuse predator–prey interactions and angler exploitation. State statute prohibits the introduction of artificial structural habitats into Wisconsin waterbodies. The use of commercial and human-made artificial habitat structures is increasing in popularity, particularly in private waterbodies, in aging reservoirs, and in areas where trees are limited (Bolding et al. Citation2004, Baumann et al. Citation2016). Although arguments for using artificial habitat structures include fish attraction, observed growth of periphyton, lack of degradation, potential to remove them, and the lower probability of snagging and losing fishing lures (Bolding et al. Citation2004, Baumann et al. Citation2016), they are not natural. Fishes have not coevolved with them, and they may not provide the same natural benefits as CWH tree drops or “fish sticks” (i.e., slow nutrient release through decomposition). Because artificial habitat structures are non-biodegradable, may only attract fishes and anglers (with the potential to increase directed effort and exploitation rates) with limited potential benefit to the overall aquatic ecosystem, and are not allowed in public waters such as those found in Wisconsin, we recommend that natural CWH additions be used and promoted for fish structural habitat enhancements.
Expectations
Because few CWH addition experiments have been evaluated under the guidance provided for ecosystem studies (Carpenter Citation1998), expectations of such practices are still unclear. Our Sanford Lake experiment is designed to address many of these key uncertainties (e.g., fish production responses). It is well established and highly likely that numerous inland glacial lake fish species will be attracted to new CWH, particularly in areas of greater CWH abundance and complexity (Newbrey et al. Citation2005, Ahrenstorff et al. Citation2009, Sass et al. Citation2012). However, species-specific, life-stage-dependent, and seasonal habitat use may vary, and littoral zone depth and slope, degree of shading, and preexisting habitat conditions may influence behavioral responses (Newbrey et al. Citation2005). Along with fish behavioral responses and habitat use, we would also expect that directed effort of anglers, and potentially exploitation, would increase in areas where CWH is added.
Species-specific reproductive output may also increase, particularly for parental-guarding species such as centrarchids (Sass et al. Citation2012). Largemouth and smallmouth bass are known to construct nests in association with CWH (Becker Citation1983, Hunt and Annett Citation2002, Wills et al. Citation2004, Lawson et al. Citation2011, Weis and Sass Citation2011), while black and white crappie (Pomoxis annularis) and bluegill often nest in association with CWH and stumps (Becker Citation1983, Spotte Citation2007). Yellow perch are known to lay their egg ribbons on CWH (Becker Citation1983). The influence of CWH addition on reproductive output has not been examined in broadcast and non-parental-guarding species such as muskellunge, northern pike (Esox lucius), and walleye. Although northern pike prefer shallow, muck/detrital areas for spawning (Becker Citation1983), to which decomposition of CWH contributes, walleye and muskellunge spawning habitat usually consists of sand, gravel, cobble, and boulder structure devoid of CWH (Becker Citation1983, Bozek et al. Citation2011). Questions remain regarding the effects of naturally occurring or placed CWH in walleye and muskellunge spawning habitats. Mechanisms of CWH transport along shorelines have been found to cluster CWH along lakeshores, with developed and wind-exposed (north and east) shorelines having the least CWH (Marburg et al. Citation2009). Marburg et al. (Citation2009) found CWH loss rates of 1 to 2 decades from littoral areas, highlighting the mobility of CWH and potential short-lived effects of wood on walleye and muskellunge spawning habitat. Potential effects of CWH on walleye and muskellunge spawning substrate and subsequent spawning success are unknown and comprise an area that warrants further study. The question of effects of CWH is of further interest because some fisheries managers have disallowed CWH additions by lakeshore residents and stakeholder groups on known walleye spawning areas citing negative effects on walleye reproduction. Since Dairymen’s Inc., is our partner on the Sanford Lake experiment and specifically requested that walleye spawning areas do not receive CWH placement, our study is not designed to test for effects of CWH additions in walleye spawning areas.
Implementation of “fish sticks” or tree drops
Key considerations in the implementation of CWH additions such as “fish sticks” and tree drops to inland glacial lakes include permitting, the source of CWH, tree species to be used, and location and configuration of placement. Most fish structural habitat enhancements will require appropriate permitting from state or federal agencies. For Wisconsin, the permitting process has been fast-tracked in a general permit to allow lakeshore residents, lake associations, and stakeholder groups to conduct habitat enhancement projects with relative ease (https://dnr.wi.gov/topic/fishing/documents/outreach/FishSticksBestPractices.pdf). Permitting is generally required for structural enhancement projects because CWH can become a navigational hazard in public waterbodies under certain circumstances. “Fish sticks” are generally preferred because they do not influence the riparian forest, which is a source of natural inputs of CWH to inland glacial lakes through senescence, beaver activity, windstorms, and fire (Roth et al. Citation2007, Sass Citation2009). In general, “fish sticks” and tree drops should be attached to shore or the remaining stump and to other trees in complex bundles to prevent them from becoming navigational hazards. If available, fully crowned hardwood (e.g., oak, maple) and/or coniferous tree species should be used for “fish sticks” and tree drops because they degrade less rapidly than more labile species like aspen and birch (Hodkinson Citation1975, Guyette et al. Citation2002, Marburg Citation2006, Sass Citation2009). Because structural complexity and abundance of CWH are positively correlated with fish use, relative abundances, and fish species diversity (Newbrey et al. Citation2005, Sass et al. Citation2012), “fish sticks” and tree drops should be configured in complex bundles. This may not only attract more fishes, but also diffuse angler effort to an extent. Further, placement within a lake is another factor to consider due to water depth, and ice, wind, and wave action. “Fish sticks” or tree drops placed over deeper water may attract a greater diversity of fish species (Newbrey et al. Citation2005), and site selection should generally avoid littoral areas of high ice, wind, and wave action because these factors tend to reduce complexity rapidly and accelerate decomposition.
Management implications
Like aging US reservoirs (Juracek Citation2015), low-productivity north-temperate inland glacial lakes may be “aging” as a consequence of lakeshore residential development, thinning of the riparian forest, beaver control, fire suppression, and physical removals of CWH (Sass Citation2009). Further, permitting is generally required to add fish structural habitat, whereas there is no permit required to remove fish structural habitat. Because many north-temperate inland glacial lakes, fisheries, and aquatic food webs are supported by allochthonous sources of energy (Pace et al. Citation2004), it is likely that changes to the riparian and littoral zones of lakes have led to a regional decline in lake and fish productivity over time (Rypel et al. Citation2018). Therefore, CWH additions may be a critical management tool for supporting and enhancing fish community production. For inland glacial lakes with known CWH additions (i.e., permitted projects), collecting data on the additions (e.g., quantity, CWH characteristics) and fish community (e.g., relative abundance, size structure) may allow for evaluations of spatial and temporal trends from multiple systems, including larger lakes. However, the influence of CWH additions may be difficult to infer due to multiple confounding factors (e.g., stocking, exploitation, regulations). Therefore, deliberate whole-lake experiments, such as our Sanford Lake study, may prove essential for addressing key uncertainties regarding inland glacial lake fish structural habitat enhancements.
Acknowledgments
Special thanks to Tim Hanson, Jerry Geiger, and Dairymen’s Inc., for allowing us access to Sanford Lake and for their logistical and field support. We also thank Joe Nohner and the Midwest Glacial Lakes Partnership for sponsoring a symposium at the Midwest Fish and Wildlife Conference, Milwaukee, Wisconsin, 2018, where this work was first presented and for organizing this special issue in Lake and Reservoir Management. We also acknowledge the editor-in chief, associate editor, and two anonymous reviewers for providing constructive comments on an earlier draft of this manuscript. This research was supported by the US Fish and Wildlife Service, Federal Aid in Sportfish Restoration funding to the Wisconsin Department of Natural Resources, project F-95-P, FHCW.
References
- Ahrenstorff TD, Sass GG, Helmus MR. 2009. The influence of littoral zone coarse woody habitat on home range size, spatial distribution, and feeding ecology of largemouth bass (Micropterus salmoides). Hydrobiologia. 623(1):223–233.
- Bassett CE. 1994. Use and evaluation of fish habitat structures in lakes of the eastern United States by the USDA forest service. Bull Marine Sci. 55:1137–1148.
- Batt RD, Carpenter SR, Cole JJ, Pace ML, Cline TJ, Johnson RA, Seekell DA. 2012. Resources supporting the food web of a naturally productive lake. Limnol Oceanogr. 57(5):1443–1452.
- Batt RD, Carpenter SR, Cole JJ, Pace ML, Johnson RA, Kurtzweil JT, Wilkinson GM. 2015. Altered energy flow in the food web of an experimentally darkened lake. Ecosphere. 6(3):33.
- Baumann JR, Oakley NC, McRae BJ. 2016. Evaluating the effectiveness of artificial habitat designs in turbid reservoirs using sonar imagery. N Am J Fish Manage. 36(6):1437–1444.
- Becker GC. 1983. Fishes of Wisconsin. Madison (WI): The University of Wisconsin Press.
- Benke AC, Henry RM, III, Gillespie DM, Hunter RJ. 1985. Importance of snag habitat for animal production in southeastern streams. Fisheries. 10(5):8–13.
- Boecklen WJ, Yarnes CT, Cook BA, James AC. 2011. On the use of stable isotopes in trophic ecology. Annual Rev Ecol Evol Syst. 42(1):411–440.
- Bolding B, Bonar S, Divens M. 2004. Use of artificial structures to enhance angler benefits in lakes, ponds, and reservoirs: a literature review. Rev Fish Sci. 12(1):75–96.
- Bowen KL, Kaushik NK, Gordon AM. 1998. Macroinvertebrate communities and biofilm chlorophyll on woody debris in two Canadian oligotrophic lakes. FAL. 141(3):257–281.
- Bozek MA, Haxton TJ, Raabe JK. 2011. Walleye and sauger habitat. In Barton BA, editor. Biology, management, and culture of walleye and sauger. Bethesda (MD): American Fisheries Society.
- Carlander KD. 1977. Handbook of freshwater fishery biology: volume 2. Ames (IA): The Iowa State University Press.
- Carpenter SR. 1998. The need for large-scale experiments to assess and predict the response of ecosystems to perturbation. In Pace ML, Groffman P.M., editors. Successes, limitations, and frontiers in ecosystem science. New York (NY): Springer.
- Christensen DL, Herwig BR, Schindler DE, Carpenter SR. 1996. Impacts of lakeshore development on coarse woody debris in north temperate lakes. Ecol App. 6(4):1143–1149.
- Crawford JT, Loken LC, Casson NJ, Smith C, Stone AG, Winslow LA. 2015. High-speed limnology: using advanced sensors to investigate spatial variability in biogeochemistry and hydrology. Environ. Sci. Technol. 49(1):442–450.
- Czarnecka M. 2016. Coarse woody debris in temperate littoral zones: implications for biodiversity, food webs and lake management. Hydrobiologia. 767(1):13–25.
- Everett RA, Ruiz GM. 1993. Coarse woody debris as a refuge from predation in aquatic communities. Oecologia. 95:475–486.
- France RI. 1997. Macroinvertebrate colonization of woody debris in Canadian shield lakes following riparian clearcutting. Cons Biol. 11(2):513–521.
- Francis TB, Schindler DE. 2006. Degradation of littoral habitats by residential development: woody debris in lakes of the Pacific Northwest and Midwest, United States. Ambio. 35(6):274–280.
- Gaeta JW, Guarascio MJ, Sass GG, Carpenter SR. 2011. Lakeshore residential development and growth of largemouth bass (Micropterus salmoides): a cross-lakes comparison. Ecol Freshw Fish. 20(1):92–101.
- Gaeta JW, Sass GG, Carpenter SR. 2014. Drought-driven lake level loss: effects on coarse woody habitat and fish. Can J Fish Aquat Sci. 71(2):315–325.
- Gennaretti F, Arseneault D, Begin Y. 2014. Millennial stocks and fluxes of large woody debris in lakes of the North American taiga. J Ecol. 102(2):367–380.
- Glaz PN, Nozais C, Arseneault D. 2009. Macroinvertebrates on coarse woody debris in the littoral zone of a boreal lake. Mar Freshwater Res. 60(9):960–970.
- Glibert PM, Middelburg JJ, McClelland JW, Vander Zanden MJ. 2018. Stable isotope tracers: enriching our perspectives and questions on sources, fates, rates, and pathways of major elements in aquatic systems. Limnol Oceanogr. 64(3):950–981.
- Gregory SV, Boyer KL, Gurnell AM. 2003. The ecology and management of wood in world rivers. Bethesda (MD): American Fisheries Society.
- Guyette RP, Cole WG. 1999. Age characteristics of coarse woody debris (Pinus strobus) in a lake littoral zone. Can J Fish Aquat Sci. 56(3):496–505.
- Guyette RP, Cole WG, Dey DC, Muzika R-M. 2002. Perspectives on the age and distribution of large wood in riparian carbon pools. Can J Fish Aquat Sci. 59(3):578–585.
- Helmus MR, Sass GG. 2008. The rapid effects of a whole-lake reduction of coarse woody debris on fish and benthic macroinvertebrates. Freshwater Biol. 53(7):1423–1433.
- Hodkinson ID. 1975. Dry weight loss and chemical changes in vascular plant litter of terrestrial origin in a beaver pond ecosystem. J Ecol. 63(1):131–142.
- Hunt J, Annett CA. 2002. Effects of habitat manipulations on reproductive success of individual largemouth bass in an Ozark reservoir. N Am J Fish Manage. 22(4):1201–1208.
- Jennings MJ, Emmons EE, Hatzenbeler GR, Edwards C, Bozek MA. 2003. Is littoral habitat affected by residential development and land use in watersheds of Wisconsin lakes? Lake Res Manage. 19(3):272–279.
- Juracek KE. 2015. The aging of America’s reservoirs: in-reservoir and downstream physical changes and habitat implications. J Am Water Res Assoc. 51(1):168–184.
- Kempinger JJ, Carline RF. 1977. Dynamics of the walleye (Stizostedion vitreum vitreum) population in Escanaba Lake, Wisconsin, 1955–72. J Fish Res Board Can. 34(10):1800–1811.
- Keshavanath P, Gangadhar B, Ramesh TJ, van Rooij JM, Beveridge MCM, Baird DJ, Verdegem MCJ, van Dam AA. 2001. Use of artificial substrates to enhance production of freshwater herbivorous fish in pond culture. Aquac Res. 32(3):189–197.
- Lawson ZJ, Gaeta JW, Carpenter SR. 2011. Coarse woody habitat, lakeshore residential development, and largemouth bass nesting behavior. N Am J Fish Manage. 31(4):666–670.
- Layman CA, Araujo MS, Boucek R, Hammerschlag-Peyer CM, Harrison E, Jud ZR, Matich P, Rosenblatt AE, Vaudo JJ, Yeager LA, et al. 2012. Applying stable isotopes to examine food web structure: an overview of analytical tools. Biol. Rev. 87(3):545–562.
- Lebkuecher JG, Neville TD, Wallace KL, Barber LF. 1998. Primary production in sandy-bottom streams of the West Sandy Creek watershed of Tennessee. Castanea. 63:130–137.
- Marburg AE. 2006. Spatial and temporal patterns of riparian landcover, forests, and littoral coarse wood in the Northern Highland Lakes District, Wisconsin, USA. PhD dissertation. Madison (WI). University of Wisconsin-Madison.
- Marburg AE, Turner MG, Kratz TK. 2006. Natural and anthropogenic variation in coarse wood among and within lakes. J Ecol. 94(3):558–568.
- Marburg AE, Bassak SB, Kratz TK, Turner MG. 2009. The demography of coarse wood in north temperate lakes. Freshw Biol. 54(5):1110–1119.
- Newbrey MG, Bozek MA, Jennings MJ, Cook JE. 2005. Branching complexity andmorphological characteristics of coarse woody structure as lacustrine fish habitat. Can J Fish Aquat. Sci. 62(9):2110–2133.
- Pace ML, Cole JJ, Carpenter SR, Kitchell JF, Hodgson JR, Van de Bogert MC, Bade DL, Kritzberg ES, Bastviken D. 2004. Whole-lake carbon-13 additions reveal terrestrial support of aquatic food webs. Nature. 427(6971):240–243.
- Pratt TC, Smokorowski KE, Muirhead JR. 2005. Development and experimental assessment of an underwater video technique for assessing fish-habitat relationships. Arch. Hydrobiol. 164(4):547–571.
- Ricker WE. 1975. Computation and interpretation of biological statistics of fish populations. Fish Res Board Can Bulletin 191. Anchorage: ARLIS.
- Roth BM, Kaplan IC, Sass GG, Johnson PT, Marburg AE, Yannarell AC, Havlicek TD, Willis TV, Turner MG, Carpenter SR. 2007. Linking terrestrial and aquatic ecosystems: the role of woody habitat in lake food webs. Ecol Model. 203(3–4):439–452.
- Rypel AL, Goto D, Sass GG, Vander Zanden MJ. 2015. Production rates of walleye and their relationship to exploitation in Escanaba Lake, Wisconsin, 1965–2009. Can J Fish Aquat Sci. 72(6):834–844.
- Rypel AL, Goto D, Sass GG, Vander Zanden MJ. 2018. Eroding productivity of walleye populations in northern Wisconsin lakes. Can J Fish Aquat Sci. 75(12):2291–2301.
- Sass GG, Kitchell JF, Carpenter SR, Hrabik TR, Marburg AE, Turner MG. 2006. Fish community and food web responses to a whole-lake removal of coarse woody habitat. Fisheries. 31(7):321–330.
- Sass GG, Gille CM, Hinke JT, Kitchell JF. 2006. Whole-lake influences of littoral structural complexity and prey body morphology on fish predator-prey interactions. Ecol Freshwater Fish. 15(3):301–308.
- Sass GG. 2009. Coarse woody debris in lakes and streams. In: Likens GE, editor. Encyclopedia of inland waters. volume 1. Oxford (UK): Elsevier.
- Sass GG, Carpenter SR, Gaeta JW, Kitchell JF, Ahrenstorff TD. 2012. Whole lake addition of coarse woody habitat: response of fish populations. Aquat Sci. 74(2):255–266.
- Sass GG, Rypel AL, Stafford JD. 2017. Fisheries habitat management: lessons learned from wildlife ecology and a proposal for change. Fisheries. 42(4):197–209.
- Scheurell MD, Schindler DE. 2004. Changes in the spatial distribution of fishes in lakes along a residential development gradient. Ecosystems. 7:98–106.
- Schindler DE, Geib SI, Williams MR. 2000. Patterns of fish growth along a residential development gradient in north temperate lakes. Ecosystems. 3(3):229–237.
- Schindler DE, Scheuerell MD. 2002. Habitat coupling in lake ecosystems. Oikos. 98(2):177–189.
- Smokorowski KE, Pratt TC, Cole WG, McEachern LJ, Mallory EC. 2006. Effects of periphyton and macroinvertebrates from removal of submerged wood in three Ontario lakes. Can J Fish Aquat Sci. 63(9):2038–2049.
- Spotte S. 2007. Bluegills: biology and behavior. Bethesda (MD): American Fisheries Society.
- Stewart-Oaten A, Murdoch WW, Parker KR. 1986. Environmental impact assessment: “pseudoreplication” in time? Ecology. 67(4):929–940.
- Vadeboncoeur Y, Lodge DM. 2000. Periphyton production on wood and sediment: substratum-specific response to laboratory and whole-lake nutrient manipulations. J N Am Benthol Soc. 19(1):68–81.
- Vadeboncoeur Y, Kalff J, Christoffersen K, Jeppesen E. 2006. Substratum as a driver of variation in periphyton chlorophyll and productivity in lakes. J N Am Benthol Soc. 25(2):379–392.
- Weis JJ, Sass GG. 2011. Largemouth bass nest site selection in small, north temperate lakes varying in littoral coarse woody habitat abundances. N Am J Fish Manage. 31(5):943–951.
- Wills TC, Bremigan MT, Hayes DB. 2004. Variable effects of habitat enhancementstructures across species and habitats in Michigan reservoirs. Trans Am Fish Soc. 133(2):399–411.
- Zale AV, Parrish DL, Sutton TM. 2012. Fisheries techniques. 3rd ed. Bethesda (MD): American Fisheries Society.
