
Abstract
Reavie ED. 2019. Paleolimnology supports aquatic management by providing early warnings of stressor impacts. Lake Reserv Manage. 36:210–217.
Recent, abrupt transitions to undesirable states have emerged as a common phenomenon in aquatic systems. Anthropogenic drivers such as climate change, species invasions, and pollution can push an aquatic system past a critical threshold for preservation of ecosystem services, resulting in a new condition. Recognizing these transitions is important for management because ecosystem function or services are typically altered or lost. A lack of aquatic monitoring data makes predictions about future conditions difficult. Evidence from sedimentary records can provide a replacement for these missing data. Paleolimnology is a valuable tool for management of aquatic systems because it can provide detailed records of recent changes that may serve as an early warning of future degradation if stressors remain unchecked. This review provides several examples of ways imminent environmental problems can be forecast using a variety of sedimentary indicators, including diatoms, pigments, aquatic insects, and the remains of invasive zooplankton. Using a case example of recent eutrophication, I recommend modeling approaches to make future projections of conditions to support aquatic management.
In studies of ecological changes during the Anthropocene, a proposed epoch representing the most recent centuries of notable human impact on Earth's ecosystems, the term “early warning” refers to some ecological signal of a less desirable future condition. These projected changes are generally detrimental shifts resulting in a loss of ecosystem services such as ecotourism, aesthetics, and fisheries (Dobson et al. Citation2006). There is a growing body of literature providing empirical evidence for early warning systems in environmental data (Carpenter and Brock Citation2006, Dakos et al. Citation2008, Drake and Griffen Citation2010, Carpenter et al. Citation2011). Ecosystem disruptions resulting from stressors can destabilize food webs, ultimately resulting in trophic cascades, a type of regime shift that alters food-web structure and ecosystem processes such as primary production, ecosystem respiration, and nutrient cycling. Detrimental results can include loss of fisheries, harmful algal blooms, and a wide range of socioeconomic impacts (Breffle et al. Citation2013). In efforts to detect early warning signals, research has focused on field experiments (e.g., Carpenter et al. Citation2011) and monitoring data (described in the following). Policies and practices intended to improve ecosystem services need to be based on verified information tracking loss to the system. Paleolimnology offers powerful tools to provide this early warning information. Traditionally, Anthropocene paleolimnological research has been undertaken to answer several questions for a given aquatic system, as clearly laid out by Smol (Citation2008): What stressors are causing impacts? When did it start? How bad has the situation become? While not necessarily unique from these prescribed goals, in this summary I encourage the use of sedimentary records as early warning systems for future environmental impacts.
Paleolimnology as a complement or replacement for monitoring
Sometimes monitored data are used to predict environmental changes. Early warning signals in environmental data may be discernible in advance of major regime shifts (Carpenter and Brock Citation2011). For example, the Canadian Ecological Monitoring and Assessment Network (Tegler et al. Citation2001) includes a broad suite of site-based sampling to track the presence of aquatic invasive species, among several other indicators. The detection of nonnative species through contemporary monitoring is probably one of the most obvious early warning systems, as it would detect invasions at a very early phase of establishment, potentially allowing for eradication. If the drivers of a detected regime shift are sufficiently gradual, such as the early detection of a nonnative species, then there may be time to take remedial action. Modeling studies have suggested that some regime shifts can be reversed by prompt intervention after early warnings are detected (Biggs et al. Citation2009). For example, zebra mussels were eradicated from a lacustrine quarry in Virginia following extensive remedial efforts following early detection (Fernald and Watson Citation2013). To date, such successes have been rare. If the ecosystem has been forced over some critical threshold, then remediation becomes less likely and instead the benefits of such a warning tend to manifest as ways to manage under the new regime. For instance, many Arctic pond systems have passed desiccation thresholds driven by recent climate change (Smol and Douglas Citation2007), so any ecosystem management considerations should assume these systems have been (or soon will be) lost.
While not frequently mentioned as a goal of monitoring programs, Bettinetti et al. (Citation2012) noted the suitability of zooplankton as an “early warning pollutant ring bell” in lakes using an example from Lake Iseo, northern Italy. They particularly observed that the accumulation of organic, hydrophobic pollutants increased in organisms lower in the food web, documenting that zooplankton make a superior indicator of impending problems well before they are recognized in fish, the organism that is, not surprisingly, the greater priority for protection.
Unfortunately, most aquatic systems are not monitored due to logistic and funding constraints, so our ability to detect early warnings is hampered by a lack of long-term observational data prior to transition. Lake sediment cores provide a wealth of information on past ecosystem states, yet a number of issues with the nature of the data need to be addressed before their potential is fully realized. While there are some examples of long-standing programs in marine (Wolfe et al. Citation1987; Nohe et al. Citation2016) and occasionally freshwater (Kovalenko et al. Citation2018) systems, more than 20 years of long-term ecological monitoring data are publicly available for fewer than 0.1% of the world’s 117 million lakes (Spears et al. Citation2016). Monitoring programs rarely provide sufficiently long time scales of water quality observations, so archived proxies from sedimentary records can be used to fill huge data gaps. The following are some examples of paleolimnological records that contain early warning signals.
Lake acidification in the Adirondacks
An early warning may manifest as the appearance or disappearance of key indicator species. Charles et al. (Citation1990) identified a previously undescribed species of diatom, Fragilaria acidobiontica Charles (among other taxa), distinctly increasing in the uppermost sediment intervals of several Adirondack (state of New York) lakes, a clear biological response to recent lake acidification. This taxon was limited to lakes with pH less than 5.5 and having aluminum concentrations higher than ∼200 µg/L. Such an “invasion” of a new taxon might have been detected had monitoring programs been in place, but the abundance in sediments confirmed that F. acidobiontica had become a new, dominant component of the primary producers in these lakes.
Climate, Cyclotella sensu lato, and changing food webs
The appearance of F. acidobiontica in so many Adirondack lakes suggests this was not a true invasion from a nonnative species, but instead the change in aquatic condition regionally favored a new clade of diatoms. Such changes in primary producers are especially important as early warning signals, as one would expect shifts so low on aquatic food webs to be the first detectable biological response to a change in water quality. Left unchecked, reorganization of primary producers driven by increasing stressors will eventually manifest as changes in upper trophic levels that rely on algae: invertebrates (e.g., zooplankton and aquatic insects) and vertebrates (e.g., fish). Such is the expectation in the Laurentian Great Lakes, where the rise in Cyclotella sensu lato in all of the lakes is predicted to be the early stages of widespread biological impacts on aquatic condition (Reavie et al. Citation2017). The relative rise in cyclotelloids in the Great Lakes is associated with multiple worldwide studies of similar trends (Rühland et al. Citation2008, Citation2015). Following on this early warning, it has been recommended that experimentation be explored to determine eventual food-web effects. In a multi-trophic-level study, evidence from fossil records in the Hudson Bay lowlands revealed that phytoplankton (typified by diatoms and the genus Cyclotella) exhibited the greatest modification due to recent climate change (Rühland et al. Citation2013), followed by invertebrates such as cladocerans and chironomids (Rühland et al. Citation2014). Given that the highest northern latitudes will experience the fastest and most extreme increases in temperature (IPCC Citation2007), it is not surprising that a larger functional portion of the food web has been impacted in these lakes.
Invasion ecology
Sediments may also hold records of species invasions. It is difficult to confirm whether recent establishment of small-celled primary producers is a result of human introduction, for instance, via ballast water discharge or interlake transport of bait buckets, because there are several other ways these microorganisms can be relocated, such as by wind currents, birds, and historical hydrological connectivity (Kristiansen Citation1996). In fact, paleolimnology has been used to provide evidence against purported algal invasives such as the prolific Didymosphenia geminata, which actually appears to have flourished in Quebec due to its tolerance to the modern climate regime (Lavery et al. Citation2014). Similarly, the nuisance alga Stephanodiscus binderanus was long considered to be a Eurasian introduction to North America (Round Citation1984), but fossil records from Lake Simcoe (Ontario, Canada) clearly indicated its presence in the lake well before foreign introductions to the system were possible (Hawryshyn et al. Citation2012). However, larger organisms such as zooplankton and fish leave remains that can be detected in sediment records and thus the timing of introduction, speed of establishment, future trajectories and ancillary impacts can be determined. A recent example is a fossil examination of the caudal spines of spiny waterflea, Bythotrephes longimanus. In a northern Minnesota reservoir, Branstrator et al. (Citation2017) noted that spines were being deposited in sediments as many as 12 years before the year of detection in the water column (late 1970s versus 1990; ), even before the first detection in any lake on the continent (i.e., 1989; Yan et al. Citation1992) with the exception of Lake Ontario. They were able to estimate first presence, early distribution, growth trajectory, and early impacts on daphnid prey species. Studies such as this clarify the dynamics of invading species by demonstrating their rate of establishment and food-web impacts, thereby enabling projections based on future management scenarios. Such historical studies of emerging threats related to nonnative species remain rare, but in the future will likely be supported by genetic information stored in sediments (Domaizon et al. Citation2017). Conceivably, molecular, micro- and macro-subfossil remains in sedimentary records could be used to detect the presence of nonnative species well before specimens are observed from the water column.
Figure 1. Bythotrephes longimanus spine accumulation rate as a function of estimated year of deposition in a sediment core from Island Lake, Minnesota. A best-fit line defined by the logistic equation is superimposed. B. longimanus image courtesy of the public domain Freshwater and Marine Image Bank. (Modified from Branstrator et al. [Citation2017].).
![Figure 1. Bythotrephes longimanus spine accumulation rate as a function of estimated year of deposition in a sediment core from Island Lake, Minnesota. A best-fit line defined by the logistic equation is superimposed. B. longimanus image courtesy of the public domain Freshwater and Marine Image Bank. (Modified from Branstrator et al. [Citation2017].).](/cms/asset/811bdb68-a4a4-4e83-b7fc-0a0d78ba6732/ulrm_a_1645249_f0001_b.jpg)
Early warnings can be more subtle in that a warning may manifest as evidence of a coming “tipping point” instead of a monotonic change in some informative sedimentary variable. It has been theorized that “flickering” and increased asymmetry of fluctuations in community records can indicate ecosystem stress and early warning of transition (Carpenter and Brock Citation2006), for instance, to a higher trophic state (Wang et al. Citation2012). Such early warning signals may be due to a system switching back and forth between alternative states after some impact. Wang et al. (Citation2012) largely theorized on this mechanism because high-resolution time series are required to discern flickering characteristics, a property that can be lacking in sedimentary archives because of the time integration of sediment intervals. Such flickering would be even harder to detect in lower intervals of cores that have greater compression and therefore greater time integration per unit depth. However, Wang et al. combined observed and mathematically modeled data for in-lake and catchment properties of Erhai Lake, Yunnan, China. They were able to show that rising variance in parameters (e.g., chironomid larval head capsules and calcium in the sediment, and catchment crop production and rainfall) started a few decades before the eventual transition to eutrophic lake conditions. Their findings strongly suggested that a flickering signal in an aquatic system provides a warning that the system has left the “safe operating space” (Rockström et al. Citation2009). They further suggested that paleolimnology had a distinct advantage in detecting such phenomena because of the provision of multidecadal time-series data.
Preferred terminology
The term “early warning” has been applied to historical events, such as a distinct shift in subfossil chironomid assemblages around 1975, which served as a precursory indicator of forthcoming eutrophication in Lake Tiiläänjärvi, Finland (Luoto and Ojala Citation2014). When used in a paleoecological context, I argue that the term “early warning” should demonstrate changes that have occurred recently, such as within the last few years or decades in a sedimentary record, and not for long-past incidents. As an anthropomorphic term, “warning” should compel human responses to these changes in anticipation of future impacts, as opposed to a long-past event where nobody was actually “warned.” Within Anthropocene time scales and for the sake of future management, early warnings in paleolimnology are probably best restricted to the most recent 5 decades, as that period can be used to estimate a trajectory for future change, especially if a permanent transition to a new (and possibly less desirable) state has not already occurred. Long-past sedimentary records that tracked early ecological transitions in response to some driver should be identified as “precursors,” while the “early warning” moniker can be reserved for contemporary changes.
The temporal resolution of a sedimentary record can vary widely due to post-depositional processes such as physical turbation or bioturbation and, given often low accumulation rates in sedimentary records, offer limited temporal resolution compared to well-funded contemporary monitoring. The best short-term resolution that can be expected would be months and more likely seasons (e.g., Köster and Pienitz Citation2006). Such a record would generally only be available at or near a core surface, where sediment compression has been minimal. While such limitations must be considered, they should rarely limit indications of early warning signals.
How can we use sedimentary early warning data to better support management?
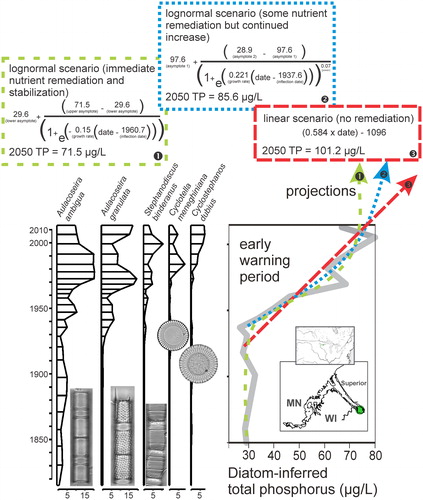
To identify an early warning of some growing threat to a particular lake is a useful step that may result in remedial action, assuming the results are clearly presented to policymakers. Such has been the case in a recent paleolimnology study of the St. Louis River Estuary, a major tributary to Lake Superior that has been subject to multiple stressors since settlement of the twin-ports cities of Duluth, MN, and Superior, WI. A paleolimnology study was undertaken to support delisting of the local area of concern (AOC), an impacted area comprising the estuary catchment, river, receiving bay, and a western portion of Lake Superior (Alexson et al. Citation2018). The AOC was defined in the late 1980s by the US Environmental Protection Agency (EPA) as requiring remediation based on a set of beneficial use impairments, including excess nutrients from anthropogenic sources (Minnesota Pollution Control Agency and Wisconsin Department of Natural Resources [MPCA and WDNR] Citation1992). The sedimentary analyses included diatoms, diatom-inferred phosphorus, algal pigments, metals, and pollen. As hypothesized, advanced sewage treatment facilities resulted in water quality improvements in the main stem of the river and western Lake Superior. These positive findings are being integrated into final reporting by local managing agencies in support of delisting the AOC. However, sediment cores from nearshore areas of the study system revealed unanticipated impacts that were not previously detected. In particular, nearshore bays exhibited rising cyanobacterial pigments that indicated potential for future harmful algal blooms. A suite of well-known, eutraphentic diatom indicators also increased in upper intervals (). Through the use of a diatom-based transfer function for total phosphorus (Reavie et al. Citation2006), quantitative data on recent nutrient increases were reconstructed, presenting a likely eutrophication trend (, right-hand panel). Eutrophication was attributed to possible factors (climate change, increasing precipitation and storms, stored sediment phosphorus) that were not considered during original listing of the AOC and so were not considered relevant to the delisting process, but it is clear that these unanticipated trends are an early warning of new ecosystem changes that are likely to continue.
Figure 2. Examples of five eutraphentic diatoms in a sedimentary record from Allouez Bay, Duluth-Superior Harbor (Alexson et al. Citation2018), and associated diatom-inferred lakewater total phosphorus concentrations. Examples of modeled future nutrient scenarios are based on holding conditions at current nutrient loads (1, green: 4-parameter model equation), some reduction in nutrients (2, blue: 5-parameter equation), and no remediation (3, red: linear equation).
From warnings to future predictions
The well-known and sometimes maligned “hockey-stick” plot of historical global temperatures, which is partly derived from paleoecological records, has received numerous treatments of future projections (Stocker et al. Citation2014). Unfortunately, such quantitative scenario applications are rarely applied to other paleorecords, which is surprising given frequent observations of the hockey-stick profile in paleolimnology studies. Here, I recommend a simple modeling approach that may be useful to convert early warnings into management recommendations.
The new eutrophication trend in the St. Louis River (Alexson et al. Citation2018) is reminiscent of the simultaneous re-eutrophication of western Lake Erie (Watson et al. Citation2016), a phenomenon that has received considerable attention and has resulted in significant threats to drinking-water supplies and other ecosystem services. The mechanisms driving this recent change are not yet understood. Nonetheless, to make meaningful management recommendations it is important to make quantitative projections of future scenarios, an approach that has received little attention in paleolimnology studies. By modeling the diatom-inferred phosphorus (DI-TP) data in , we derived 3 future scenarios: (1) a pessimistic, non-action and linear scenario that would result in hyper-eutrophication (prevailing P concentrations in excess of 100 µg/L); (2) an action scenario involving best nutrient management practices resulting in asymptotic nutrient concentrations at current levels; and (3) a reduction in the rate of nutrient increase, using a relatively complex rate coefficient that simulates a more gradual asymptotic nutrient response. While not shown, the latter scenario could be modified to a power function that includes reductions in nutrient concentration, if such a scenario is realistic. In this specific case, the current hypothesis is that increasing precipitation is supplying more nutrients from the watershed, so under an assumption of continuing climate change the second scenario (; blue) may be the most realistic.
This case example illustrates a nearly monotonic historic change reflecting an early warning, which allows for relatively simple statistical modeling techniques for future projections. However, occurrences of recent flickering in sedimentary records would be more difficult to model. Such complex warning systems still need to be identified in modern records, a research area that is complicated because sediment time series can contain irregular sampling intervals and compaction of sediments. Annually laminated sequences would be ideal for characterization of flickering because seasonal and annual conditions would be observable. Unfortunately, such records are rare and often restricted to highly stable sedimentary regimes (e.g., meromictic and some Arctic systems; Anderson et al. Citation1985, Besonen et al. Citation2008).
Conclusions
Paleolimnology should be a part of any aquatic environmental program that requires retrospective data for management of stressor-driven future scenarios. Discovery of unanticipated impacts may result from these records, as was the case in the St. Louis River, so such applications may have value in any environmental sampling program where limnological characteristics have been sufficiently stable to preserve sedimentary records. These records may be used in lieu of monitoring data or in combination with them because of the integrative nature of sedimentary archives. While this review cannot summarize the multitude of indicators and types of impacts that can be characterized, all sedimentary records have the potential to show recent trends leading to not-yet-realized losses of ecosystem services. Further, the types of recent trends may range from monotonic (easy to interpret) to erratically flickering (difficult to interpret), but there is potential to use modeling to make quantitative projections of future conditions.
References
- Alexson EE, Reavie ED, Axler RP, Yemets SV, Krasutsky PA, Edlund MB, Pillsbury RW, Desotelle D. 2018. Paleolimnology of a freshwater estuary to inform area of concern nutrient delisting efforts. J Paleolimnol. 59(3):373–95. doi:10.1007/s10933-017-0014-8.
- Anderson RY, Dean WE, Bradbury JP, Love D. 1985. Meromictic lakes and varved lake sediments in North America. US Geological Survey Bulletin 1607. Washington (DC): US Government Printing Office.
- Besonen MR, Patridge W, Bradley RS, Francus P, Stoner JS, Abbott MB. 2008. A record of climate over the last millennium based on varved lake sediments from the Canadian High Arctic. Holocene. 18(1):169–80. doi:10.1177/0959683607085607.
- Bettinetti R, Garibaldi L, Leoni B, Quadroni S, Galassi S. 2012. Zooplankton as an early warning system of persistent organic pollutants contamination in a deep lake (Lake Iseo, northern Italy). J Limnol. 71(2):e36. doi:10.4081/jlimol.2012.e36.
- Biggs R, Carpenter SR, Brock WA. 2009. Turning back from the brink: detecting an impending regime shift in time to avert it. Proc Natl Acad Sci USA. 106(3):826–31. doi:10.1073/pnas.0811729106.
- Branstrator DK, Beranek AE, Brown ME, Hembre LK, Engstrom DR. 2017. Colonization dynamics of the invasive predatory cladoceran, Bythotrephes longimanus, inferred from sediment records. Limnol Oceanogr. 62(3):1096–110. doi:10.1002/lno.10488.
- Breffle WS, Muralidharan D, Donovan RP, Liu F, Mukherjee A, Jin Y. 2013. Socioeconomic evaluation of the impact of natural resource stressors on human-use services in the Great Lakes environment: a Lake Michigan case study. Resour Policy. 38(2):152–61. doi:10.1016/j.resourpol.2012.10.004.
- Carpenter SR, Brock WA. 2006. Rising variance: a leading indicator of ecological transition. Ecol Lett. 9(3):311–8. doi:10.1111/j.1461-0248.2005.00877.x.
- Carpenter SR, Brock WA. 2011. Early warnings of unknown nonlinear shifts: a nonparametric approach. Ecology. 92(12):2196–201.
- Carpenter SR, Cole JJ, Pace ML, Batt R, Brock WA, Cline T, Coloso J, Hodgson JR, Kitchell JF, Seekell DA, et al. 2011. Early warnings of regime shifts: a whole-ecosystem experiment. Science. 332(6033):1079–82. doi:10.1126/science.1203672.
- Charles DF, Binford MW, Furlong ET, Hites RA, Mitchell MJ, Norton SA, Oldfield F, Paterson MJ, Smol JP, Uutala AJ, et al. 1990. Paleoecological investigation of recent lake acidification in the Adirondack Mountains. J Paleolimnol. 3(3):195–241. doi:10.1007/BF00219459.
- Dakos V, Scheffer M, van Nes EH, Brovkin V, Petoukhov V, Held H. 2008. Slowing down as an early warning signal for abrupt climate change. Proc Natl Acad Sci USA. 105(38):14308–12. doi:10.1073/pnas.0802430105.
- Dobson A, Lodge D, Alder J, Cumming GS, Keymer J, McGlade J, Mooney H, Rusak JA, Sala O, Wolters V, et al. 2006. Habitat loss, trophic collapse, and the decline of ecosystem services. Ecology. 87(8):1915–24.
- Domaizon I, Winegardner A, Capo E, Gauthier J, Gregory-Eaves I. 2017. DNA-based methods in paleolimnology: new opportunities for investigating long-term dynamics of lacustrine biodiversity. J Paleolimnol. 58(1):1–21. doi:10.1007/s10933-017-9958-y.
- Drake JM, Griffen BD. 2010. Early warning signals of extinction in deteriorating environments. Nature. 467(7314):456–9. doi:10.1038/nature09389.
- Fernald RT, Watson BT. 2013. Eradication of zebra mussels (Dreissena polymorpha) from Millbrook Quarry, VA. In: Quagga and zebra mussels: biology, impacts and control. Boca Raton (FL): Lewis Publishers. p. 195–214.
- Hawryshyn J, Rühland KM, Julius M, Smol JP. 2012. Absence of evidence is not evidence of absence: is Stephanodiscus binderanus (Bacillariophyceae) an exotic species in the Great Lakes region? J Phycol. 48(2):270–4. doi:10.1111/j.1529-8817.2012.01123.x.
- IPCC. 2007. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL, editors. Climate change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press. p. 996.
- Köster D, Pienitz R. 2006. Seasonal diatom variability and paleolimnological inferences–a case study. J Paleolimnol. 35(2):395–416. doi:10.1007/s10933-005-1334-7.
- Kovalenko KE, Reavie ED, Barbiero RP, Burlakova LE, Karatayev AY, Rudstam LG, Warren GJ, Watkins J. 2018. Patterns of long-term dynamics of aquatic communities and water quality parameters in the Great Lakes: are they synchronized? J Great Lakes Res. 44(4):660–9. doi:10.1016/j.jglr.2018.05.018.
- Kristiansen J. 1996. Dispersal of freshwater algae—a review. Hydrobiologia. 336(1-3):151–7. doi:10.1007/BF00010829.
- Lavery JM, Kurek J, Rühland KM, Gillis CA, Pisaric MF, Smol JP. 2014. Exploring the environmental context of recent Didymosphenia geminata proliferation in Gaspésie, Quebec, using paleolimnology. Can J Fish Aquat Sci. 71(4):616–26. doi:10.1139/cjfas-2013-0442.
- Luoto TP, Ojala AE. 2014. Paleolimnological assessment of ecological integrity and eutrophication history for Lake Tiiläänjärvi (Askola, Finland). J Paleolimnol. 51(4):455–68. doi:10.1007/s10933-014-9766-6.
- Minnesota Pollution Control Agency and Wisconsin Department of Natural Resources (MPCA and WDNR). 1992. The St. Louis River system remedial action plan stage one [accessed 2019 Apr 25]. https://stlouisriver.org/wp-content/uploads/2015/08/SLRRAP1992.pdf.
- Nohe A, Lagring R, de Cauwer K, Goffin A, Tyberghein L, Deneudt K, Vyverman W, Sabbe K. 2016. Long-term phytoplankton monitoring data (1970-2010) from the Belgian North Sea reveal shifts in community composition and seasonal dynamics. In: Mees J, Seys J, editors. Book of abstracts—VLIZ Marine Scientist Day. Brugge, Belgium, 12 February 2016. VLIZ Special Publication, 75. Oostende (Belgium): Vlaams Instituut voor de Zee - Flanders Marine Institute (VLIZ). p. xii. [accessed 2019 May 1] http://www.vliz.be/imisdocs/publications/285381.pdf.
- Reavie ED, Axler RP, Sgro GV, Danz NP, Kingston JC, Kireta AR, Brown TN, Hollenhorst TP, Ferguson MJ. 2006. Diatom-based weighted-averaging transfer functions for Great Lakes coastal water quality: relationships to watershed characteristics. J Great Lakes Res. 32(2):321–47. doi:10.3394/0380-1330(2006)32[321:DWTFFG.2.0.CO;2]
- Reavie ED, Sgro GV, Estepp LR, Bramburger AJ, Chraïbi VLS, Pillsbury RW, Cai M, Stow CA, Dove A. 2017. Climate warming and changes in Cyclotella sensu lato in the Laurentian Great Lakes. Limnol Oceanogr. 62(2):768–83. doi:10.1002/lno.10459.
- Rockström J, Steffen W, Noone K, Persson A, Chapin FS, Lambin EF, Lenton TM, Scheffer M, Folke C, Schellnhuber HJ, et al. 2009. A safe operating space for humanity. Nature. 461(7263):472–5. doi:10.1038/461472a.
- Round FE. 1984. The ecology of the algae. Cambridge (UK): Cambridge University Press.
- Rühland KM, Hargan KE, Jeziorski A, Paterson AM, Keller WB, Smol JP. 2014. A multi-trophic exploratory survey of recent environmental changes using lake sediments in the Hudson Bay Lowlands, Ontario, Canada. Arct Antarct Alp Res. 46(1):139–158. doi:10.1657/1938-4246-46.1.139.
- Rühland KM, Paterson AM, Keller W, Michelutti N, Smol JP. 2013. Global warming triggers the loss of a key Arctic refugium. Proc Biol Sci. 280(1772):20131887. doi:10.1098/rspb.2013.1887.
- Rühland KM, Paterson AM, Smol JP. 2008. Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European lakes. Glob Chang Biol. 14:2740–5.
- Rühland KM, Paterson AM, Smol JP. 2015. Lake diatom responses to warming: reviewing the evidence. J Paleolimnol. 54(1):1–35. doi:10.1007/s10933-015-9837-3.
- Smol JP. 2008. Pollution of lakes and rivers: a paleoenvironmental perspective. 2nd ed. Malden (MA): Blackwell Publishing.
- Smol JP, Douglas MS. 2007. Crossing the final ecological threshold in high Arctic ponds. Proc Natl Acad Sci USA. 104(30):12395–7. doi:10.1073/pnas.0702777104.
- Spears BM, Carvalho L, Futter MN, May L, Thackeray SJ, Adrian R, Angeler DG, Burthe SJ, Davidson TA, Daunt F, et al. 2016. Ecological instability in lakes: a predictable condition? Environ Sci Technol. 50(7):3285–6. doi:10.1021/acs.est.6b00865.
- Stocker TF, Qin D, Plattner G-K, Tignor MMB, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM. 2014. Climate change 2013: the physical science basis: Working Group I contribution to the fifth assessment report of the Intergovernmental Panel on Climate Change. New York (NY): Cambridge University Press.
- Tegler B, Sharp M, Johnson MA. 2001. Ecological monitoring and assessment network’s proposed core monitoring variables: an early warning of environmental change. Environ Monit Assess. 67(1/2):29–55.
- Wang R, Dearing JA, Langdon PG, Zhang E, Yang X, Dakos V, Scheffer M. 2012. Flickering gives early warning signals of a critical transition to a eutrophic lake state. Nature. 492(7429):419–22. doi:10.1038/nature11655.
- Watson SB, Miller C, Arhonditsis G, Boyer GL, Carmichael W, Charlton MN, Confesor R, Depew DC, Höök TO, Ludsin SA, et al. 2016. The re-eutrophication of Lake Erie: harmful algal blooms and hypoxia. Harmful Algae. 56:44–66. doi:10.1016/j.hal.2016.04.010.
- Wolfe DA, Champ MA, Flemer DA, Mearns AJ. 1987. Long-term biological data sets: their role in research, monitoring, and management of estuarine and coastal marine systems. Estuaries. 10(3):181–93. doi:10.2307/1351847.
- Yan ND, Dunlop WI, Pawson TW, MacKay LE. 1992. Bythotrephes cederstroemi (Schoedler) in Muskoka Lakes: first records of the European invader in inland lakes in Canada. Can J Fish Aquat Sci. 49(2):422–6. doi:10.1139/f92-048.
