
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Stager JC, Harvey L, Chimileski S. 2020. Long-term cultural eutrophication in White and Walden Ponds (Concord, Massachusetts, USA), Thoreau’s lakes of light. Lake Reserv Manage. XX:XXX–XXX.
Two historically important ponds in the vicinity of Boston, MA, were subjected to a comparative paleolimnological investigation of the timing and causes of eutrophication trends in each. The remarkable clarity of White Pond during the early 19th century led Henry David Thoreau to compare it favorably to nearby Walden Pond, but during the 20th century water quality in both ponds declined. Sediment core studies show that cultural eutrophication began at Walden during the 1930s, but no long-term sediment records have been available for White Pond, which makes it more difficult to determine the history and causes of eutrophication there. Here we use microfossil and geochemical analyses of sediment cores to show that major changes in the diatom community of White Pond began around 1900, when fish stocking commenced and soil erosion due to land use in the watershed increased, and that the trend intensified around 1960 and 1990. We also describe efforts to mitigate eutrophication at White Pond, highlight the ecological importance of benthic vegetation in nutrient cycling, and suggest that threats to water quality in both of these ponds will likely increase due to anticipated climatic changes in the region.
Walden Pond (Concord, MA) is a heavily used recreational destination for residents of the greater Boston area and an international symbol of wild nature due to the writings of Henry David Thoreau (Citation1852, Citation1854). Less widely known is White Pond, roughly half the size of Walden and located 4 km to the southwest. Both are deep glacial kettle ponds long recognized for their exceptional clarity, which led Thoreau (Citation1854) to write that "White Pond and Walden are great crystals, lakes of light." However, like many oligotrophic waters nationwide (Stoddard et al. Citation2016), both ponds have experienced water quality declines since the 19th century (Köster et al. Citation2005, ESS Group Citation2014, Citation2016–2017, Walker Citation2017, Stager et al. Citation2018, Gerzon Citation2019).
Recent efforts by the Massachusetts Department of Conservation and Recreation (DCR) to prevent further eutrophication in Walden Pond have been largely successful thus far (Maynard Citation2004, Stager et al. Citation2018). In contrast, water quality has continued to deteriorate at White Pond, where the watershed's mosaic of private residences, properties owned by the Town of Concord, and natural resources managed by the state make the situation more complicated. Unlike Walden, White Pond has been closed to swimming several times since 2015 due to toxic cyanobacterial blooms (WPAC Citation2015, Gerzon Citation2019).
Although field studies of limnological conditions in White Pond have been conducted since the 1980s (Walker and Ploetz Citation1988, Citation1989, Citation1990, ESS Group Citation2014, Citation2016–2017, Walker Citation2017), no longer term environmental records are yet available to inform its management, unlike the situation for Walden Pond, for which sediment core records of limnological conditions cover the last 1500 years (Winkler Citation1993, Köster et al. Citation2005, Stager et al. Citation2018). In this article, we summarize the current state of knowledge of ecological changes in White Pond and extend that record several centuries farther back in time with the aid of diatoms (single-celled algae with glassy shells or "frustules") and organic matter preserved in 2 sediment cores. Microscopic examination of diatom frustules in such cores allows the identification of species that reflect past ecological conditions, including water clarity, and fluctuations in sediment organic content often represent variable soil erosion regimes or changes in lake productivity. Together, these kinds of analyses can provide valuable insights into the environmental history of a lake, particularly for time periods from which direct historical observations are lacking. We also use our findings to address the following questions: (1) How has the diatom community of White Pond changed since Thoreau's time? (2) What are the most likely causes of those changes? (3) What can be done to protect water quality in White and Walden Ponds in the context of future climatic change?
Study site
White Pond (42°25'40" N; 71°23'27"W) is a flow-through seepage lake of ∼16 ha that is rimmed by steep slopes within a watershed of ∼46 ha (ESS Group Citation2014). Local bedrock is granitic and overlain by sandy glacial deposits (Walker and Ploetz Citation1989). The pond has a volume of 1.3 million m3, mean depth of 8 m, and maximum depth of ∼15 m () in 3 subbasins that develop very low concentrations of dissolved oxygen in summer (ESS Group Citation2014, Walker Citation2017). Surface pH has ranged between 6 and 7 since the 1970s (Walker and Ploetz, Citation1988, Citation1989, Citation1990), and Secchi disk transparency generally has ranged between 4 and 8 m since 1987 (Walker Citation2017). Aquatic mosses resembling Fontinalis or Drepanocladus () were retrieved in the tops of sediment cores collected from 10 to 15 m depth at multiple locations in 2017 and 2018, indicating light penetration sufficient to permit photosynthesis on the pond floor.
Figure 1. Site maps. (Top left) New England with Concord–Boston area highlighted, modified from Wikimedia Commons. (Top right) White and Walden Ponds in relation to Concord and Boston, modified from d-maps.com. (Bottom) Map of the White Pond watershed, with a contour map of the pond and land use classification of surrounding properties. Coring sites within the pond are indicated with stars. Land use map modified from ESS Group (Citation2016–2017), based on Town of Concord parcel data and USGS aerial imagery; contour map modified from MassWildlife (Citation2020).
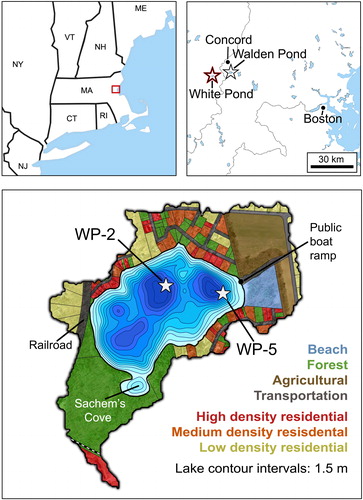
Figure 2. Benthic aquatic moss recovered from 15 m depth in White Pond. Millimeter scale shown above.

All hydrological inputs to White Pond derive from groundwater seepage, precipitation, and nearshore runoff. Outputs are by evaporation and groundwater flow-through with a flushing rate of ∼0.2 times/year (Walker and Ploetz Citation1988, Citation1989, ESS Group Citation2014). Groundwater pumping, including that from a town well southeast of the pond, may affect water levels slightly, but surface levels are most strongly correlated with precipitation (Walker and Ploetz Citation1988, Citation1989, ESS Group Citation2014, Walker Citation2017). Between 1996 and 2016, the level of the pond fluctuated by nearly 2 m (Walker, Citation2017).
More than 100 private lots with approximately 3 dozen residences, most of which are occupied year-round, occupy two-thirds of the watershed (; ESS Group Citation2014). Roughly 5 ha on the eastern rim of the watershed are cultivated and 20 ha of forested conservation and reservation lands in the southern sector are owned by the Town of Concord (ESS Group Citation2014). Public access for recreational activities is provided by a state-run boat ramp beside a formerly private beach on the eastern shore and footpaths to an undeveloped beach at Sachem's Cove on the southwestern shore ().
Historical background
Indigenous peoples lived in the region for thousands of years before Anglo-Europeans established the Town of Concord in 1635, and White Pond itself was purchased from native residents in 1684 (Gutteridge Citation1921, Marquet Citation1973, Gerzon Citation2019). However, the time frame most relevant to this study began during the early 19th century.
Thoreau visited White Pond frequently during the early 1800s and produced a detailed bathymetric map of it, in addition to one of Walden Pond (Thoreau Citation1854, Deevey Citation1942). At that time the pond's beauty and clarity were already legendary. In Walden; or Life in the Woods (Citation1854), Thoreau wrote that "Since the woodcutters, and the railroad, and I myself have profaned Walden, perhaps the most attractive … of all our lakes, the gem of the woods, is White Pond." By the 1830s, however, roads bordering the northeastern corner of the watershed had encouraged more commerce and settlement near the pond (ESS Group Citation2014). Around 1875, the Framingham and Lowell Railroad opened a track near the western shore that continued to operate until the 1970s (Marquet Citation1973, Gerzon Citation2019).
During the early 20th century (and probably earlier), farming occurred on what are now residential lots along Powder Mill Road on the northern edge of the White Pond watershed (; Gerzon Citation2019). By the 1920s the pond had become a "very lively place" due to heavy recreational use of its beaches, and in 1930 the White Pond Association (WPA) was formed, in part, to help protect the shoreline from overuse (Marquet Citation1973, Gerzon Citation2019). In 1931, former farmland in the northern and southeastern sectors of the watershed was subdivided as the Pine Knoll Shores development and sold for summer homes (ESS Group Citation2014, Gerzon Citation2019). The WPA maintained a beach on the eastern shore for several hundred members from that time until 2019, when the Town of Concord acquired the property (Kaye D, Concord Department of Natural Resources, 2020, pers. comm.). Road access to the eastern side of the pond was improved around 1930 (ESS Group Citation2014) but anglers complained of lack of public right-of-way during the 1930s and 1940s (Kaye D, Concord Department of Natural Resources, 2020, pers. comm.). From this, it appears that the boat ramp () may not have been available to the public until after 1949. By the 1960s, the watershed contained a "very high density of residences" and many of the former summer homes were occupied year-round (ESS Group Citation2014).
Fish stocking commenced at White Pond in 1903 according to unpublished reports by the Massachusetts Department of Fish and Game (MADFG; Kaye D, Concord Department of Natural Resources, 2020, pers. comm.), and it continues today. An early record of fish community composition from 1911 (ESS Group Citation2014) listed golden shiners (Notemigonus crysoleucas) and pumpkinseeds (Lepomis gibbosus), but previous stocking had also introduced brown trout (Salmo trutta) and "pike perch" (possibly Sander sp.) by then. A seemingly random array of species was initially stocked for several decades, including yellow perch (Perca flavescens), smallmouth bass (Micropterus dolomieu), largemouth bass (Micropterus salmoides), bullheads (Ameiurus nebulosis), chain pickerel (Esox niger), crappie (Pomoxis nigromaculatus), brook trout (Salvelinus fontinalis), bluegills (Lepomis macrochirus), and pumpkinseeds. In 1949, only perch, smallmouth bass, and bluegills were considered well established, and in 1953 yellow perch were abundant and also stunted in size, presumably due to competition for food.
The pond was poisoned ("reclaimed") with rotenone in September 1953, prior to restocking with rainbow trout (Oncorhynchus mykiss), brown trout, and brook trout. Golden shiners, bullheads, largemouth bass, and pumpkinseeds were reported again in 1956, and the pond was reclaimed a second time in September 1958. By 2013–2014, bluegills, pumpkinseeds, brown and rainbow trout, golden shiners, and largemouth bass were reported yet again (ESS Group Citation2014).
In 1972, an unpublished fisheries report (MADFG, 1972, unpubl. data) concluded that White Pond was still "clean" but that woodlands near the southern shoreline should be protected from development. In response, the Town of Concord established 4 ha of forested White Pond Conservation Land in 1973 and 16 ha of White Pond Reservation Land in 1992 (ESS Group Citation2014). Swimming is discouraged on the town-owned shore of Sachem's Cove, but the rule has not been consistently enforced. In 1975, a White Pond Advisory Committee was formed to address pond-related issues, and Friends of White Pond was established in 1987 in response to continued concerns over the environmental condition of the pond (Gerzon Citation2019). In 2019, the town acquired the WPA beach, installed the first public sanitary facilities there, and constructed access stairways, fencing, and shore stabilization structures at Sachem's Cove (Kaye D, Concord Department of Natural Resources, 2020, pers. comm.).
Frequent limnological monitoring by Walker (Citation2017) since 1987 has shown that transparency in summer as measured by Secchi disk has declined and that it also became less variable during the past decade. Transparency formerly ranged between 3 and 10 m but it is now mostly within the 4–7 m range. The clarity of the pond as viewed from the surface is not necessarily indicative of its overall productivity, however. As early as the 1980s, a "bulge" of oxygen enrichment and high chlorophyll a concentrations indicated high phytoplankton productivity near the thermocline around 5–9 m depth (ESS Group Citation2016–2017, Walker Citation2017). The apparent absence of such a bulge in 1975 (Walker and Ploetz Citation1988) suggests that mid-depth productivity might have increased shortly thereafter.
The earliest report of cyanobacterial blooms in White Pond was in summer 1986, when Anabaena, Anacystis, and Chroococcus were predominant genera (Walker and Ploetz Citation1988). Summer blooms were also noted in 1987 and 1988 (ESS Group Citation2014), and no-swim advisories were issued due to cyanobacterial toxicity in 2015, 2016, 2017, and 2019 (ESS Group Citation2016–2017, Gerzon Citation2019).
It is not known how long deepwater anoxia has occurred in White Pond, but dissolved oxygen concentrations of <1 ppm were measured in the lowest 3 m of the pond during summer 1974 (MADFG, 1974, unpubl. data). Extreme oxygen depletion below 15 m depth was also documented in 1988 (Walker and Ploetz Citation1989), and the oxygen-depleted zone (<2 ppm) on the pond bottom increased in thickness from ∼2 m to 5 m between 2006 and 2016 (Walker Citation2017).
Materials and methods
Two sediment cores were collected for analysis from 2 locations within White Pond. Core WP-2 of 50 cm length was collected from 15 m water depth in the central basin in May 2017 (), using a UWITEC gravity corer. Gravity core WP-5 of 27 cm length was taken from 12.5 m depth in the eastern basin in July 2018. Both cores were extruded vertically in the field at 1 cm increments, bagged, and stored under refrigeration prior to analysis. The sediment–water interface in all cases appeared to be intact, and WP-5 also contained horizontal laminations within the uppermost centimeter.
Radiocarbon ages were determined for bulk sediment samples from the cores through accelerator mass spectrometry and were converted to calibrated ages with CALIB 7.1 (; Stuiver and Reimer Citation1993). Age–depth models representing the last 1–2 centuries () were constructed by measuring 210Pb activity with depth and applying the constant rate of supply model (Appleby and Oldfield Citation1978). Because the 210Pb profile of the shorter WP-5 core suggested that background activity levels might not have been reached at the base of the core (), the age-depth model of WP-2 was considered more reliable and only WP-2 was subjected to more detailed analyses. Organic contents of the sediments in core WP-2 were estimated by percent weight loss on ignition (%LOI) at 500 C (Sutherland Citation1998, Heiri et al. Citation2001).
Figure 3. Lead-210 and sediment accumulation profiles with age–depth models for White Pond cores WP-2 (left) and WP-5 (right). Dotted lines indicate temporal uncertainty ranges for the age–depth models.
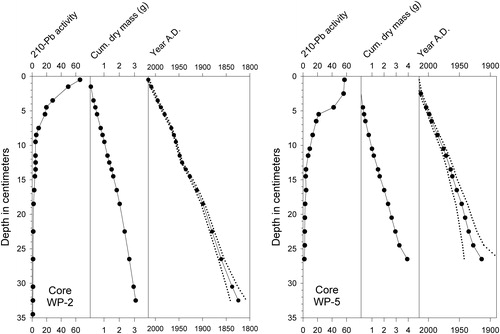
Table 1. Radiocarbon ages of sediment samples from White Pond cores WP-2 and WP-5. Age calibrations were conducted with CALIB 7.1 (Stuiver and Reimer Citation1993). Calibrated ages in parentheses are assumed to be anomalously old due to the Suess effect (see text).
Samples for diatom analysis were diluted in distilled water, dried on coverslips, and mounted on glass slides with Permount mounting medium. Microscopic examination showed no significant clumping or viewing interference from organic films or debris that would require removal by chemical treatments, so the subsamples were not subjected to further processing that could damage microfossils, an approach that has also been used successfully elsewhere (Stager et al. Citation2018, Citation2019).
A mean of 440 diatom valves per sample from the WP-2 core were identified at 1000 under oil immersion using standard references (Patrick and Reimer Citation1966, Citation1975, Krammer and Lange-Bertalot Citation1991, Diatoms of North America 2019). Siliceous scales of chrysophyte algae were also enumerated along with the diatoms. Diatoms in core WP-5 were enumerated at coarser temporal and taxonomic resolution for comparative purposes only, with a mean of 300 diatoms counted per sample.
Results
Lead-210 activity in core WP-2 decreased downward roughly exponentially to ∼16 cm depth, below which uniform background levels were reached (). In core WP-5, however, activity dropped more sharply below 5 cm depth and continued to decrease to levels at the base of the core that did not definitively represent background (Engstrom D, Science Museum of MN, 2018, pers. comm.). The 210Pb age models for cores WP-2 and WP-5 suggested that their basal sediments were deposited during the mid 18th century and circa AD 1900, respectively (). In contrast, the radiocarbon results () indicated basal ages closer to 1600 and 600 years, respectively ( and ).
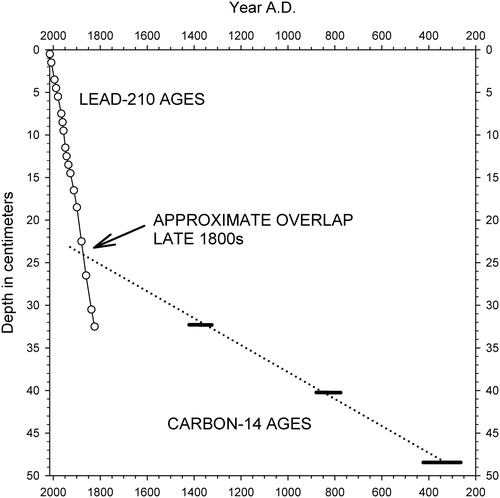
Figure 4. Composite of linear age–depth models for core WP-2 using 210Pb (solid line) and radiocarbon dating (dotted line). The approximated intersection of the age–depth lines at ∼23–24 cm depth suggests that sediment accumulation rates accelerated during the late 19th century.
The organic contents (%LOI) in WP-2 ranged close to 60% between 50 cm and 20 cm depths, with a minor decrease between 24 and 22 cm depth (). Above 20 cm, the %LOI values declined to the 40% range between 15 and 4 cm depth, then rose again to nearly 60% at the core top.
Figure 5. Stratigraphy of the most common diatom taxa in White Pond core WP-2. Ratios of chrysophyte scales to diatom valves indicated as "Scales:Diatoms." Organic content of sediments indicated by weight loss on ignition (%LOI). Horizontal dotted lines indicate qualitative assemblage zones A–E, as described in the text. STOCK indicates the onset of fish stocking in 1903. REC marks the time frame in which 2 reclamations occurred (see text). Ages of depth intervals based on 210Pb and radiocarbon dating are shown on right margin.
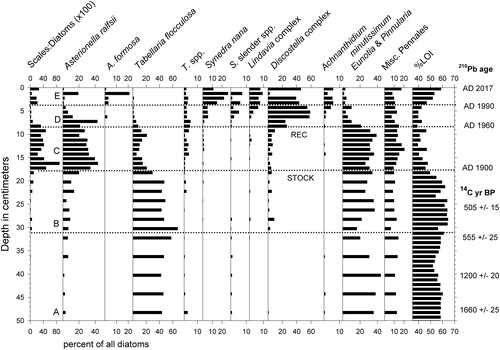
The most common diatom taxa in the lower half of WP-2 were Tabellaria flocculosa Roth (Kützing) and members of the genera Eunotia and Pinnularia (). Asterionella ralfsii var. americana Körner and members of the Discostella stelligera (Cleve & Grunow) Houk & Klee complex were most common in the upper half of the core, along with the Lindavia bodanica sensu lato group (mostly L. cf. affinis (Grunow) Nakov, Guillory, Julius, E.C. Ther., & Alverson), A. formosa Hassal, and Synedra nana F. Meister. The diatom assemblages and stratigraphy in core WP-5 were similar (). We visually grouped diatom assemblages in core WP-2 into stratigraphic zones for discussion purposes here (), based upon first appearances or major increases in ecologically informative taxa.
Figure 6. Stratigraphy of the most common diatom taxa in supplemental core WP-5. Ratios of chrysophyte scales to diatom valves indicated as "Scales:Diatoms." Horizontal dotted lines indicate major transitions in diatom assemblage composition of nature and timing similar to those in core WP-2. Ages of depth intervals based on 210Pb and radiocarbon dating are shown on right margin.
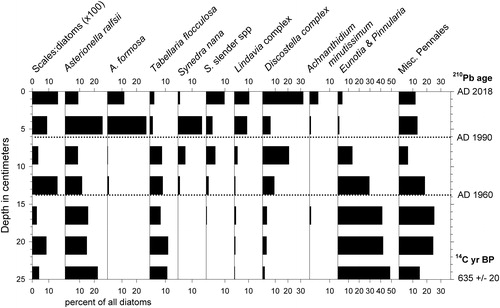
Zone A (50–31 cm; uncertain age to ca. 1840s)
Approximately half of the diatoms in this basal zone were benthic taxa belonging to the genera Eunotia and Pinnularia (mostly P. biceps W. Gregory), along with lesser abundances of other genera including Frustulia, Navicula, Cymbella, and Stauroneis. Also common were varieties of T. flocculosa, particularly the planktonic form IIIP sensu Koppen.
Zone B (31–18 cm; ca. 1840s to 1900)
The first appearance of small numbers of the D. stelligera complex at 31 cm depth characterized this zone. Chrysophyte algal scales also became slightly more numerous.
Zone C (18–9 cm; ca. 1900 to 1960)
Elongated A. ralfsii var. americana became abundant and percentages of T. flocculosa decreased. Discostella, Lindavia, S. nana, and Achnanthidium minutissimum (Kutzing) Czarnecki were present in small amounts, and ratios of chrysophyte scales (mainly in the genus Mallomonas) to diatom valves were the highest of the record.
Zone D (9–4 cm; ca. 1960 to 1990)
The D. stelligera complex, including both D. stelligera and D. pseudostelligera (Hustedt) Houk & Klee, represented more than half of the diatom assemblage. This zone saw declines in relative abundances of Eunotia, Pinnularia, A. ralfsii, and T. flocculosa, accompanied by increases in S. nana and other elongated, lightly silicified taxa listed here as "Synedra slender species," including one resembling unicellular Fragilaria crotonensis Kitton. The earliest appearance of A. formosa occurred here, as well. Ratios of chrysophyte scales to diatom valves were low.
Zone E (4–0 cm; ca. 1990 to 2017)
Members of the D. stelligera complex were numerous, and Eunotia, Pinnularia, and T. flocculosa remained uncommon. Widely fluctuating percentages of A. ralfsii, A. formosa, and S. nana each represented up to ∼20% of the assemblage, and Lindavia, Synedra slender species, and A. minutissimum ranged up to 10% of the assemblage. Ratios of chrysophyte scales to diatom valves were nearly as high as in Zone C.
Discussion
The 210Pb profiles of the 2 cores indicated that WP-2 represented a longer time period than WP-5 (). That result was expected because WP-5 was shorter and was also collected from a site closer to shore that was more directly exposed to runoff from the boat ramp (), a setting in which sediment accumulation rates were likely to be higher than offshore.
Radiocarbon ages of sediments in the lower portion of core WP-2 were much older than expected from the 210Pb-derived ages in the upper 30 cm ( and and ), and they were also significantly older than in a core that was collected from similar depth in Walden Pond (). Because the 210Pb-derived date for the 25.5 cm interval in the WP-2 core was circa 1870, the much older radiocarbon age obtained for sediment from the same interval () can reasonably be attributed to the Suess effect, by which global contamination with the ancient carbon in fossil fuel emissions since the 18th century has skewed the apparent ages of organic matter to older dates (Tans et al. Citation1979). A radiocarbon date from 25.5 cm depth in core WP-5 () was also considered anomalous due to the Suess effect.
Figure 7. Diatom stratigraphy of Walden Pond core WAL-3, after Stager et al. (Citation2018). Ratios of chrysophyte scales to diatom valves indicated as "Scales:Diatoms." Organic content of sediments indicated by weight loss on ignition (%LOI). Ages of depth intervals based on 210Pb and radiocarbon dating are shown on right margin.
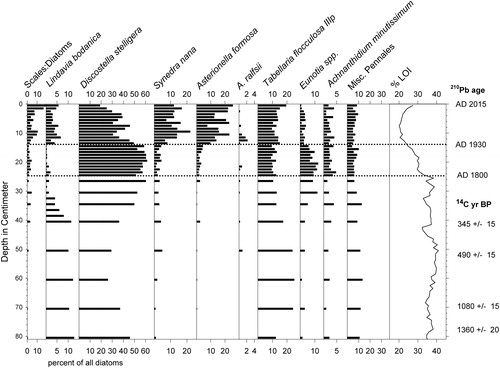
In contrast, the 3 oldest radiocarbon ages from WP-2 appeared to be accurate. They represented time periods that likely preceded the Suess effect, and contamination of the sediments with ancient carbon from local mineral deposits is unlikely to cause age discrepancies at White Pond because of the granitic bedrock and siliceous sands of the watershed. We find no evidence of stratigraphic disturbance because the ages increased systematically with depth (), and the radiocarbon ages of sediments in WP-2 and WP-5 were generally comparable at equivalent depths (). In addition, the diatom records of the 2 cores were similar ( and ) despite having been collected from different sites.
We therefore conclude that (1) the WP-2 core represents ∼1600 years of lake history, and (2) the steeper slope of the age–depth model for core WP-2 that was based on 210Pb dating reflected a major acceleration of sediment inputs to White Pond since the late 19th century ().
Ecological changes in the 19th century
The earliest noteworthy change in the WP-2 diatom record was the first appearance of the D. stelligera complex that defined the boundary between Zone A and Zone B in the core (). This taxon is common in the plankton of oligotrophic to mesotrophic lakes and has been associated with land clearance and soil erosion elsewhere (Beck et al. Citation2016, Wengrat et al. Citation2019). Unfortunately, the age–depth model for the 35–25 cm interval remains unresolved due to conflicting dates where the radiocarbon and 210Pb series overlap (), and the inferred radiocarbon age of the transition was several centuries older than the age derived from the 210Pb series. If the former age is correct, then the change occurred close to the time when Concord was first settled by non-Indigenous people. If the latter is correct, then the change occurred during the early to mid 1800s, around the time when Thoreau (Citation1854) described the pond as being extremely clear ("pellucid") but also green in color. The green coloration is consistent with moderate nutrient enrichment from forest fires, land clearance, shoreline disturbance, and/or other human impacts on the watershed, which is also consistent with the WP-2 record.
More definitively dated ecological changes occurred at White Pond during the late 19th century as local land use intensified. A brief decline of %LOI values in core WP-2 that began during the 1870s ( and ) probably reflected inorganic inputs from soil erosion during construction of the railroad near the western edge of the lake. A more sustained decline in %LOI above the 20 cm level in WP-2 was most likely due to soil erosion after circa 1890 that resulted from increased foot traffic around Sachem's Cove and development of the woodlands surrounding the pond. Changes in the diatom community accompanied the %LOI decline, including 4-fold increases in percentages of A. ralfsii and ratios of chrysophyte scales to diatoms ( and ). Similar sediment inputs have also been shown to influence diatom assemblage composition and productivity elsewhere (Köster et al. Citation2005, Maier et al. Citation2018, Citation2019) and could account for faster sediment accumulation rates since the late 19th century ().
Figure 8. Summary of major changes in diatom assemblages and organic content in White Pond vs. time, in comparison to Walden Pond and weather records from Bedford, Massachusetts (National Centers for Environmental Information [NOAA] Citation2017). Percentages of diatoms in core WP-2 (A–D): (A) Eutrophication indicators A. formosa and Synedra species. (B) Centric Discostella and Lindavia species groups. (C) Asterionella ralfsii. (D) Benthic taxa including Eunotia, Pinnularia, and Achnanthidium. (E) Organic content of sediments indicated by weight loss on ignition (%LOI). (F) Percentages of A. formosa and S. nana indicating eutrophication in Walden Pond core WAL-3. Vertical dotted lines highlight approximate dates of major stratigraphic transitions in core WP-2: (1) Onset of the rise in percentages of A. ralsfii ca. 1900. (2) Onset of the rise in centric taxa and decline in benthic diatoms ca. 1960. (3) Onset of the latest rise in diatoms indicative of eutrophication during the early 1990s.
![Figure 8. Summary of major changes in diatom assemblages and organic content in White Pond vs. time, in comparison to Walden Pond and weather records from Bedford, Massachusetts (National Centers for Environmental Information [NOAA] Citation2017). Percentages of diatoms in core WP-2 (A–D): (A) Eutrophication indicators A. formosa and Synedra species. (B) Centric Discostella and Lindavia species groups. (C) Asterionella ralfsii. (D) Benthic taxa including Eunotia, Pinnularia, and Achnanthidium. (E) Organic content of sediments indicated by weight loss on ignition (%LOI). (F) Percentages of A. formosa and S. nana indicating eutrophication in Walden Pond core WAL-3. Vertical dotted lines highlight approximate dates of major stratigraphic transitions in core WP-2: (1) Onset of the rise in percentages of A. ralsfii ca. 1900. (2) Onset of the rise in centric taxa and decline in benthic diatoms ca. 1960. (3) Onset of the latest rise in diatoms indicative of eutrophication during the early 1990s.](/cms/asset/1a4f100a-2ade-46b3-aa06-47d378d96781/ulrm_a_1839606_f0008_b.jpg)
Thoreau (Citation1854) claimed that Walden Pond was less pristine than White Pond during the early to mid 19th century, an observation that is consistent with our sediment core records. Organic contents in Walden sediments began to decrease around 1800 (), presumably due to forest disturbance and recreational activity that increased soil erosion and inputs of wastes from humans and livestock (Köster et al. Citation2005, Stager et al. Citation2018). Railroad construction near Walden Pond's western shore in 1844 would have further increased erosional inputs, as would subsequent forest fires set by sparks from locomotives. After sediment organic contents began to decline, planktonic Asterionella formosa became more common while benthic taxa declined slightly, indicating moderate cultural eutrophication ().
Ecological changes in the 20th century
In 1911, a fisheries assessment report (MADFG, 1911, unpubl. data) described White Pond as "perfectly clear, like Walden" with the "clearest water I have seen." However, the microfossil record of core WP-2 shows that the diatom community was already beginning to change significantly by then.
Percentages of A. ralfsii in White Pond rose sharply around 1900. This planktonic taxon is considered to be an indicator of acidification in many oligotrophic lakes (Sivarajah et al. Citation2017, Stager et al. Citation2019), and its brief appearance at Walden Pond during the mid 20th century () led Winkler (Citation1993) to interpret it as a sign of mild acidification there. However, circumneutral pH measurements at White Pond during the last half-century revealed no evidence of significant acidification (Walker and Ploetz Citation1988, Citation1989, Citation1990). The increase of A. ralfsii in this case more likely resulted from nutrient enrichment due to land use and soil erosion, as has been documented elsewhere (Turkia et al. Citation1998).
The onset of fish stocking in 1903 might also have influenced the diatom community through top-down trophic effects, waste inputs from hatchery fish, mortality during stocking and angling, and/or increased shoreline degradation caused by anglers in pursuit of the stocked fish (Carpenter et al. Citation1985, Lyons et al. Citation2016, Sienkiewicz and Gạsiorowski 2016). Diatom communities of oligotrophic lakes in Poland have undergone similar changes after fish stocking (Sienkiewicz and Gạsiorowski 2016), but the composition of the fish community and degree of fishing pressure at White Pond prior to stocking are unknown. It is therefore not possible to rigorously evaluate possible impacts of stocking apart from noting the nature and timing of such activities in relation to the shift in the diatom record.
Scales of chrysophyte algae also became more abundant in the sediments circa 1900. Increases of chrysophyte populations in other temperate zone lakes have been variously attributed to climate warming (Paterson et al Citation2001, Ginn et al. Citation2010), pH changes (Paterson et al. Citation2001, Dixit et al. Citation2002), and land use (Lott et al. Citation1994). The early onset of the change in White Pond makes local nutrient enrichment the most likely cause rather than large-scale external factors, particularly in light of enhanced sediment inputs (reduced %LOI) and fisheries activities then, as well as the lack of a contemporaneous rise in chrysophyte abundances in Walden Pond until much later ().
More pronounced eutrophication began at White Pond around 1960, as indicated by the first appearance of A. formosa and increased percentages of D. stelligera and S. nana ( and ), which are common in Walden Pond () and other moderately productive lakes (Turkia et al. Citation1998, Saros et al. Citation2005, Stager et al. Citation2018, Wengrat et al. Citation2019). Decreasing percentages of benthic Eunotia and Pinnularia in WP-2 ( and ) are consistent with shading by increasingly dense phytoplankton populations.
The change in diatoms circa 1960 occurred shortly after the reclamations of the lake, which indicates a possible causal link. The ecological impacts of nutrients released over short time periods by reclamations can be amplified by reduced concentrations of dissolved oxygen due to bacterial decay of the carcasses (Hupfer and Lewandowski Citation2008, Nürnberg Citation2009, Stager Citation2018). Once in place, a resultant cycle of anoxia, internal loading of nutrients, and phytoplankton productivity can become self-sustaining (Ekdahl et al. Citation2004, Citation2007).
The mass of fish killed during the 1953 reclamation was said by the MADFG to be 98 kg/ha (MADFG, 1956, unpubl. data), representing ∼1570 kg in total. If the water content of yellow perch is taken to be 80% and the phosphorus (P) content is 0.2% of dry weight (González et al. Citation2006), then the reclamation could potentially have rapidly mobilized 0.6 kg of biologically active P in the water and bottom sediments. That amount of P is less than the amount (22 kg) that the pond has been said to be capable of absorbing annually without increasing productivity (ESS Group Citation2014). In 1958, the second reclamation reportedly killed half as many fish per hectare (MADFG, 1961, unpubl. data), potentially representing half as much P-mobilization as in 1953. However, MADFG fisheries staff typically removed as many fish carcasses as possible following reclamations in other ponds in Massachusetts (Richards T, MassWildlife, Division of Fisheries and Wildlife, 2019, pers. comm.). If the same practice was applied to White Pond, then the nutrient releases from reclamation should have been negligible.
Although effects of fisheries management practices on the phytoplankton community of White Pond cannot be ruled out, the pond's primary source of long-term nutrient enrichment is the surrounding watershed. The eastern boat ramp lacked public sanitary facilities until recently, and erosion of slopes and footpaths at Sachem's Cove have long added nutrients to the pond, as has the absence of sanitary facilities there. The largest, most continuous sources of P to White Pond are residential septic systems, lawn and garden fertilizer, and surface runoff and soil erosion from the shoreline, trails, boat ramp, and roads (Walker and Ploetz Citation1988, Citation1989, ESS Group Citation2014).
Climatic changes are also likely to have contributed to the shift in the diatom community of White Pond circa 1990 ( and ) as temperatures and precipitation increased in Massachusetts, most notably since the 1980s. We are unable to make definitive inferences linking specific climatic parameters to short-term changes in the abundances of particular taxa, as has been done with seasonally resolved lake monitoring studies in Scandinavia (Maier et al. Citation2018, Citation2019). However, we do know that thickening of the epilimnion of White Pond in response to summer warming from 2006 to 2016 widened the zone of temperatures unsuitable for coldwater fish (>20 C) from ∼5 to 8 m in depth (Walker Citation2017). Surface warming that increases the stability and duration of thermal stratification in lakes in this manner often contributes to oxygen depletion and internal loading of P from bottom sediments (ESS Group Citation2014, Citation2016–2017) that can affect the composition and abundance of diatom communities. Increased precipitation in the region () along with warming is also likely to have stimulated more nutrient release from soils in the watershed by microbial decay, and heavier storm events would have washed more nutrients in from surrounding roads and slopes.
In contrast to White Pond, Walden Pond experienced no major changes in its diatom or chrysophyte communities when routine stocking of multiple fish species began in 1905 or when reclamation occurred in 1968 ( and ). The lack of responses to those practices at Walden suggests that similar fisheries management activities were not the primary cause of changes in the diatom community of White Pond around 1900 (stocking) and 1960 (reclamations). However, the volume of White Pond is much less than that of Walden (1.3 million and 3.2 million m3, respectively; Colman and Friesz Citation2001, ESS Group Citation2014), which could make its ecosystem more sensitive to such perturbations.
Conversely, a lack of contemporaneous changes at White Pond during the 1930s ( and ) when the first major increases of A. formosa and S. nana occurred at Walden ( and ) supports previous conclusions that the shift at Walden was due to local cultural eutrophication rather than to a regional driver such as climate change or acid deposition (Maynard Citation2004, Köster et al. Citation2005, Stager et al. Citation2018). Shoreline stabilization, closure of a nearby dump that reduced droppings from seagulls, and regulation of the number of visitors were among the effective strategies employed at Walden to mitigate eutrophication since the DCR took over management of the watershed in 1975 (Maynard Citation2004).
Lakes of light in the 21st century
As lakes nationwide lose their transparency to eutrophication and also to "browning" due to the release of dissolved organic carbon from soils by warmer, wetter climates (Seekel et al. Citation2015a,b, Solomon et al. Citation2015, Williamson et al. Citation2015, Stager et al. Citation2019), the clarity that waterbodies such as White and Walden Ponds displayed in the past has become increasingly rare (Stoddard et al. Citation2016). Although both ponds remain clear in comparison to many New England lakes, they rarely if ever display the 9–10 m transparencies observed by Thoreau at Walden (Deevey Citation1942).
During the last 2 decades, White Pond's phytoplankton community was more taxonomically altered than at any time in recent centuries, and it has also become much more similar to that of Walden Pond (). Planktonic A. formosa and S. nana, indicating more productive conditions (Köster et al. Citation2005, Sienkiewicz and Gạsiorowski Citation2016), are now common and benthic diatoms have declined as the water has become less transparent. The rising organic content of sediments since the mid 1990s ( and ) is also consistent with increasing productivity, and laminations at the top of core WP-5 indicated recent hypoxia sufficient to prevent sediment mixing by benthic organisms. Similar conditions caused sediment laminations to develop due to cultural eutrophication at Crawford Lake, Ontario, as well (Ekdahl et al. Citation2004, Citation2007), and they can also be exacerbated by warmer climatic regimes such as those that have developed in New England in recent decades ().
Although the eutrophication trend at Walden has more or less leveled off since the 1970s ( and ; Stager et al. Citation2018), water quality in White Pond has continued to decline. Deep-water oxygen depletion is also more severe at White Pond than at Walden (Walker and Ploetz Citation1988, ESS Group Citation2016–2017). Fortunately, local residents and the Town of Concord have recently launched aggressive efforts to mitigate the trend, despite having fewer economic and organizational resources readily available than the state-run management structure at Walden. Those efforts include shoreline stabilization, installation of public sanitary facilities, and upgrading of residential waste disposal systems. Nonetheless, the presence of several dozen residences atop the porous sandy soils surrounding such a small waterbody means that chronic nutrient inputs from former and current leach fields, lawns, gardens, and roadways are now a permanent feature of the ecosystem.
Anticipated warming in New England during this century (USGCRP Citation2017) is likely to further amplify the risk of eutrophication in both ponds by increasing the intensity and duration of summer stratification and internal loading of P (Winder and Sommer Citation2012). Future warming will also tend to draw more visitors to the ponds. Wastes released by bathers currently represent only a small fraction of the annual phosphorus budget for White Pond (ESS Group Citation2014), but much larger numbers of swimmers in Walden Pond represent as much as half of the summer phosphorus budget there (Colman and Friesz Citation2001). If storm events also become increasingly severe (USGCRP Citation2017), more abundant runoff and groundwater seepage will further increase external nutrient inputs to both ponds (Sinha et al. Citation2017). Monitoring and management of trophic status and water clarity will therefore become even more important as climates continue to evolve in future (Jeppesen et al. Citation2017).
Benthic vegetation can help to maintain water clarity by sequestering nutrients on the bottom beyond the reach of phytoplankton (Wagner et al. Citation2000). Benthic photosynthesis is also a source of dissolved oxygen that helps to mitigate internal loading of P, and it probably contributes to bulges in dissolved oxygen concentrations at depth in both ponds (Colman and Waldron Citation1998, Colman and Friesz Citation2001, ESS Group Citation2014, Citation2016-2017, Walker Citation2017). Eutrophication and associated clouding of the water column can inhibit benthic vegetation through shading and thereby lead to a self-amplifying cycle of increasing oxygen depletion, internal loading, and greater uptake of a lake's nutrient budget by phytoplankton (Wagner et al. Citation2000, Arnold et al. Citation2019). At Walden, benthic meadows represent far more biomass than the phytoplankton, meaning that a large reservoir of nutrients that is currently trapped on the bottom could become available to plankton if the macrophytes are lost due to shading (Colman and Waldron Citation1998, Colman and Friesz Citation2001). The same appears to be true for White Pond, as well.
Like a canary in a coal mine, benthic vegetation can warn of impending shifts in trophic states. However, only limited information on the composition, extent, and health of macrophyte communities in the ponds is currently available. To our knowledge, no comprehensive ecological study of the benthic community of White Pond has yet been conducted, but Thoreau collected macrophytes from up to 12 m depth there (Thoreau Citation1852), and our core samplers retrieved aquatic mosses at several offshore locations. At Walden Pond, Thoreau (Citation1854) mentioned finding a "fibrous green weed," possibly Nitella, on boat anchors during the early 19th century, Deevey (Citation1942) dredged living Fontinalis from 15.7 m depth in 1939, and Winkler (Citation1993) reported declining abundances of Isoetes due to shading during the 20th century. Greater scrutiny of the benthic vegetation in both ponds is therefore warranted.
Summary and conclusions
The diatom community of White Pond has changed more significantly since the late 1800s than at any other time in recent centuries. Formerly dominated by Tabellaria flocculosa, the planktonic component of the community has become more diverse and characterized by taxa that likewise reflect cultural eutrophication in Walden Pond. The timing of major shifts in diatom assemblages circa 1900 and 1960 indicates that fisheries management practices might have contributed somewhat to those changes. However, chronic nutrient inputs from recreational activities and settlement in the watershed were probably more important in causing the overall eutrophication trend during the 20th century. Together, the combined effects of fisheries management, nutrient enrichment, and climate change have contributed to a stepwise evolution of phytoplankton assemblages that has pushed White Pond into successively higher trophic states.
Degradation of water quality at Walden Pond has been largely mitigated in recent decades by shoreline stabilization, closure of a local landfill, provision of sanitary facilities, and other efforts by the DCR. In contrast, eutrophication has continued at White Pond to the point that cyanobacterial blooms have become a frequent health hazard, and benthic anoxia has become more extreme than at Walden. The town and shoreline residents have recently increased the availability of sanitary facilities for visitors and stabilized some of the slopes and footpaths around White Pond, as was done previously at Walden (Maynard Citation2004). Nonetheless, anticipated long-term warming and increased runoff are likely to amplify the risk of water quality problems in both ponds. Monitoring the benthic macrophyte communities could represent an early warning system for such ecological changes in the future.
Thoreau (Citation1854) said of White and Walden Ponds, "If they were permanently congealed, and small enough to be clutched, they would, perchance, be carried off … like precious stones, … but being liquid, and ample, and secured to us and our successors forever, we disregard them." As successors in the 21st century, we see both truth and error in that passage. These iconic waters may still be gem-like, but they are not secured forever and are highly vulnerable to our presence. Fortunately, the ponds are not disregarded by those who know and care for them, and we hope that this study contributes to their protection on behalf of successors yet to come.
Acknowledgments
Financial and technical support for this project was provided by the Draper-Lussi Endowment and Paul Smith's College. We thank Delia Kaye, Christine Gerzon, and Todd Richards for providing background information and literature regarding the histories of White Pond and Walden Pond, and William Walker kindly provided feedback on the manuscript. Paul Smith's College students Matt Spadoni and David Prosser assisted with sediment coring. Lead-210 dating was performed by Dan Engstrom and colleagues, and radiocarbon dating was performed by NOSAMS staff at Woods Hole Oceanographic Institute.
References
- Appleby PG, Oldfield F. 1978. The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210Pb to the sediment. CATENA 5(1):1–8.
- Arnold TE, Brenner M, Kenney WF, Bianchi TS. 2019. Recent trophic state changes of Florida lakes inferred from bulk sediment geochemical variables and biomarkers. J Paleolimnol. 62(4):409–423.
- Beck KK, Medeiros AS, Finkelstein SA. 2016. Drivers of change in a 7300-year Holocene diatom record from the hemi-boreal region of Ontario, Canada. PLOS One. 11(8):e0159937. doi:10.1371/journal.pone.0159937.
- Carpenter SR, Kitchell JF, Hodgson JR. 1985. Cascading trophic interactions and lake productivity. BioScience. 35(10):634–639.
- Colman JA, Waldron MC. 1998. Walden Pond, Massachusetts: environmental setting and current investigations. USGS Fact Sheet FS-064-98
- Colman JA, Friesz PJ. 2001. Geohydrology and limnology of Walden Pond, Concord, Massachusetts U.S. Geological Survey. Water-Resources Investigations Report 01–4137. Northborough (MA).
- Deevey ES. 1942. A re-examination of Thoreau's "Walden." Quart Rev Biol. 17:1–11.
- Diatoms of North America. 2019; [cited 2 Sep 2019]. Available from https://diatoms.org/.
- Dixit SS, Dixit AS, Smol JP. 2002. Diatom and chrysophyte functions and inferences of post-industrial acidification and recent recovery trends in Killarney lakes (Ontario, Canada). J Paleolimnol. 27(1):79–96. doi:10.1023/A:1013571821476.
- Ekdahl EJ, Teranes JL, Guilderson TP, Turton CL, McAndrews JH, Wittkop CA, Stoermer EF. 2004. Prehistorical record of cultural eutrophication from Crawford Lake. Geology. 32(9):745–748.
- Ekdahl EJ, Teranes JL, Wittkop CA, Stoermer EF, Reavie ED, Smol JP. 2007. Diatom assemblage response to Iroquoian and Euro-Canadian eturophication of Crawford Lake, Ontario, Canada. J Paleolimnol. 37(2):233–246.
- ESS Group. 2014. White Pond watershed management plan; [cited 10 Jan 2020]. Available from http://www.wwwalker.net/whitepond/reports/ESS_White_Pond_Report_Oct_2014.pdf.
- ESS Group. 2016-2017. White Pond final monitoring report; [cited 10 Jan 2020]. Available from https://concordma.gov/DocumentCenter/View/10231/2016—2017-White-Pond-Monitoring-Report?bidId=.
- Gerzon R. 2019. White Pond and Nine Acre Corner history; [cited 21 Dec 2019]. Available from http://www.preservewhitepond.org/white-pond-history.html.
- Ginn GK, Rate M, Cumming BF, Smol JP. 2010. Ecological distribution of scaled chrysophyte assemblages from the sediments of 54 lakes in Nova Scotia and southern New Brunswick. J Paleolimnol. 43(2):293–308.
- González S, Flick GJ, O’Keefe SF, Duncan SE, McLean E, Craig SR. 2006. Composition of farmed and wild yellow perch (Perca flavescens). J Food Comp Anal. 19(6-7):720–726.
- Gutteridge WH. 1921. A brief history of the Town of Maynard, Massachusetts. Boston (MA): Hudson Printing Co.
- Heiri O, Lotter AF, Lemcke G. 2001. Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. J Paleolimnol. 25(1):101–110. doi:10.1023/A:1008119611481.
- Hupfer M, Lewandowski J. 2008. Oxygen controls the phosphorus release from lake sediments - a long-lasting paradigm in limnology. Internat Rev Hydrobiol. 93(4-5):415–432.
- Jeppesen E, Søndergaard M, Zhengwen L. 2017. Lake restoration and management in a climate change perspective: an introduction. Water 9(2):122. doi:10.3390/w9020
- Köster D, Pienitz P, Wolfe BB, Barry S, Foster DR, Dixit SS. 2005. Paleolimnological assessment of human-induced impacts on Walden Pond (Massachusetts, USA) using diatoms and stable isotopes. Aquat Ecosys Health Mgt. 8(2):117–131.
- Krammer K, Lange-Bertalot H. 1991. Süßwasserflora von Mitteleuropa, 3 Teil: Centrales Fragilariaceae, Eunotiaceae. Stuttgart: Gustav Fischer Verlag.
- Lott AM, Siver PA, Marsicano LJ, Kodama KP, Moeller RE. 1994. The paleolimnology of a small waterbody in the Pocono Mountains of Pennsylvania, USA: reconstructing 19th–20th century specific conductivity trends in relation to changing land use. J Paleolimnol Paleolimnol. 12(2):75–86. doi:10.1007/BF00678088.
- Lyons RA, Johnson LK, McIntyre BM. 2016. Phosphorus loading rates in lakes with development and stocked fish in the Sierra Nevada Mountains, California, USA. Ecosphere. 7(11):e01554. 10.1002/ecs2.1554.
- Maier DB, Gälman V, Renberg I, Bigler C. 2018. Using a decadal diatom sediment trap record to unravel seasonal processes important for the formation of sedimentary diatom signal. J Paleolimnol. 60(2):133–152. doi:10.1007/s10933-018-0020-5.
- Maier DB, Diehl S, Bigler C. 2019. Inter-annual variation of seasonal diatom sedimentation reveals the importance of late winter processes and their timing for sediment signal formation. Limnol Oceanogr. 64:1186–1199.
- Marquet C. 1973. The history and legend of the White Pond and Nine Acre Corner area, Concord, MA; [cited 25 Jan 2020]. Available from http://www.preservewhitepond.org/marquets-history-and-legend-of-white-pond-and-nine-acre-corner-area.html.
- MassWildlife. Massachusetts pond maps; [cited 20 Mar 2020]. Available from https://www.mass.gov/info-details/massachusetts-pond-maps.
- Maynard WB. 2004. Walden Pond: a history. New York (NY): Oxford University Press.
- [NOAA] National Centers for Environmental Information. 2017. United States Historical Climatology Network (USHCN) [cited 9 Sep 2020]. Available from https://www.ncdc.noaa.gov/data-access/land-based-station-data/land-baseddatasets/us-historical-climatology-network-ushcn.
- Nürnberg GK. 2009. Assessing internal phosphorus load - problems to be solved. Lake Reserv Manage. 25(4):419–432.
- Paterson AM, Cumming BF, Smol JP, Hall R. 2001. Scaled chrysophytes as indicators of water quality changes since pre-industrial times in the Muskoka-Haliburton region, Ontario, Canada. Can J Fish Aquat Sci. 58:2468–2481.
- Patrick R, Reimer CW. 1966. The diatoms of the United States. Monographs of the Academy of Natural Sciences of Philadelphia Number, 131 Volume. Philadelphia (PA).
- Patrick R, Reimer CW. 1975. The diatoms of the United States. Monographs of the Academy of Natural Sciences of Philadelphia Number, 132 Volume. Philadelphia (PA).
- Saros J, Michel TJ, Interlandi SJ, Wolfe AP. 2005. Resource requirements of Asterionella formosa and Fragilaria crotonensis in oligotrophic alpine lakes: implications for recent phytoplankton community reorganizations. Can J Fish Aquat Sci. 62(7):1681–1689.
- Seekel DA, Lapierre J-F, Karlsson J. 2015a. Trade-offs between light and nutrient availability cross gradients of dissolved organic carbon concentration in Swedish lakes: implications for patterns in primary production. Can J Fish Aquat Sci. 72(11):1663–1671.
- Seekel DA, Lapierre J-F, Ask J, Bergström A-K, Deininger A, Rodríguez P, Karlsson J. 2015b. The influence of dissolved organic carbon on primary production in northern lakes. Limnol Oceanogr. 60(4):1276–1285.
- Sienkiewicz E, Gąsiorowski M. 2016. The effects of fish stocking on mountain lake plankton communities identified using palaeobiological analyses of bottom sediment cores. J Paleolimnol. 55(2):129–150.
- Sinha E, Michalak AM, Balaji V. 2017. Eutrophication will increase during the 21st century as a result of precipitation changes. Science. 357(6349):405–408. doi:10.1126/science.aan2409.
- Sivarajah B, Rühland KM, Smol JP. 2017. Are diatoms recovering to pre-acidification assemblages in a warming world? Revisiting Killarney Provincial Park lakes (Ontario, Canada). Fund App Lim. 190(1):13–28. ).
- Solomon CT, Jones SE, Weidel BC, Buffam I, Fork ML, Karlsson J, Larsen S, Lennon JT, Read JS, Sadro S, et al. 2015. Ecosystem consequences of changing inputs of terrestrial dissolved organic matter to lakes: current knowledge and future challenges. Ecosystems. 18(3):376–389., doi:10.1007/s10021-015-9848-y.
- Stager JC. 2018. Still waters: the secret world of lakes. New York (NY): W.W. Norton.
- Stager JC, Wiltse B, Hubeny JB, Yankowsky E, Nardelli D, Primack R. 2018. Climate variability and cultural eutrophication at Walden Pond (Massachusetts, USA) during the last 1800 years. PLOS One. 13(4):e0191755. doi:10.1371/journal.pone.0191755.
- Stager JC, Wiltse B, Cumming B, Holsen T, Stetler J, Laxson C, Marcillo C, Charles D. 2019. A novel ecological state at Bear Pond (Adirondack Mountains, NY, USA) following acidification and partial recovery. Lake Reserv Manage. 35(2):208–223.
- Stoddard JL, Van Sickle J, Herlihy AT, Brahney J, Paulsen S, Peck DV, Mitchell R, Pollard AI. 2016. Continental-scale increase in lake and stream phosphorus: are oligotrophic systems disappearing in the United States? Environ Sci Technol. 50(7):3409–3415. doi:10.1021/acs.est.5b05950.
- Stuiver M, Reimer PJ. 1993. Extended 14C database and revised CALIB radiocarbon calibration program. Radiocarbon. 35(1):215–230.
- Sutherland RA. 1998. Loss-on-ignition estimates of organic matter and relationships to organic carbon in fluvial bed sediments. Hydrobiologia. 389(1/3):153–167. doi:10.1023/A:1003570219018.
- Tans PP, de Jong AFM, Mook WG. 1979. Natural atmospheric 14C variation and the Suess effect. Nature. 280(5725):826–828.
- Thoreau HD. 1852. Journal 3, Chapter 6 (Mar 11, 1852); [cited 10 Jan 2020]. Available from https://www.walden.org/collection/journals/
- Thoreau HD. 1854. Walden; or life in the woods. Boston (MA): Ticknor and Fields.
- Turkia J, Sandman O, Huttunen P. 1998. Paleolimnological evidence of forestry practices disturbing small lakes in Finland. Boreal Env Res. 3:45–61.
- [USGCRP] United States Global Change Research Program. 2017. Third national climate assessment; [cited 5 Mar 2020]. Available from http://nca2014.globalchange.gov/report.
- Wagner DH, Christy JA, Larson DW. 2000. Deep-water bryophytes from Waldo Lake. Oregon Lake Reserv Manage. 16(1-2):91–99.
- Walker WW. 2017. Summary of long-term water quality monitoring data from White Pond; [cited 20 Feb 2020]. Available from http://www.wwwalker.net/whitepond/.
- Walker WW, Ploetz GP. 1988. White Pond preliminary diagnostic study. Prepared for White Pond Advisory Committee. Concord, MA. 46. pp.
- Walker WW, Ploetz GP. 1989. White Pond water quality studies 1988. Prepared for White Pond Advisory Committee, Concord, MA; [cited 15 Jan 2020]. Available from http://www.wwwalker.net/whitepond/reports/wp_wqs_jul_1989.pdf.
- Walker WW, Ploetz GP. 1990. White Pond water quality studies 1989. Prepared for White Pond Advisory Committee, Concord, MA; [cited 15 Jan 2020]. Available from http://www.wwwalker.net/whitepond/reports/wp_wqd_mar_1990.pdf.
- Wengrat S, Bennion H, Ferreira PA, de L, Figueira RCL, Bicudo DC. 2019. Assessing the degree of ecological change and baselines for reservoirs: challenges and implications for management. J Paleolimnol. 62(4):337–357.
- [WPAC] White Pond Advisory Committee. 2015. A shared future: a comprehensive vision for White Pond, its watershed, and its neighborhoods; [cited 15 Jan 2020]. Available from https://www.concordma.gov/DocumentCenter/View/4018/A-Vision-for-White-Pond-PDF?bidId=.
- Williamson CE, Overholt EP, Pilla RM, Leach TH, Brentrup JA, Knoll LB, Mette EM, Moeller RE. 2015. Ecological consequences of long-term browning in lakes. Sci Reports. 5:18666 doi:10.1038/srep18666.
- Winder M, Sommer U. 2012. Phytoplankton response to a changing climate. Hydrobiologia. 698(1):5–16.
- Winkler M. 1993. Changes at Walden Pond during the last 600 years In: Schofield EA and Baron RC, editors. Thoreau's world and ours: a natural legacy. Golden (CO): North American Press; p. 199–211.