
Abstract
The ichnogenus Kouphichnium and associated ichnofossils attributed to xiphosuran activity are here re-examined from samples collected from the Pennsylvanian-age Steven C. Minkin Fossil Site at the Union Chapel Mine, in Walker County, Alabama, USA. The large sample size offers an unique opportunity to evaluate some Kouphichnium ichnospecies. Thus, the morphological variability resulting from the taphonomic, ethological, taxonomic variability and underprint fallout are evaluated using this large sample set. Three morphotypes have been segregated from the material previously assigned to K. aspodon discovered at this and adjacent sites. Ichnospecies of Kouphichnium identified at the Union Chapel Mine site include: K. lithographicum, K. aspodon and two new ichnospecies (K. atkinsoni and K. minkinensis). Additionally, Kouphichnium-like traces that are associated with “jumper” traces have been previously misinterpreted as Kouphichnium and Selenichnites, respectively, and are excluded from this study, leaving them in open nomenclature as they will be the subject of a subsequent publication. We here redescribe the holotype of K. aspodon and designate lectoparatypes to better define the ichnospecies. New trace makers for some Kouphichnium ichnospecies are hypothesized, in contrast to the traditional xiphosuran attribution.
Introduction
The Steven C. Minkin Paleozoic Footprint Site (Union Chapel Mine (UCM) site; Hunt et al. Citation2005) is located in Walker County, Alabama (33 48′33.10 N; 87 09′58.90 W; ), in a partially reclaimed open pit coal mine (Allen Citation2005; ). Since the site’s discovery, the UCM collection has become one of the world’s largest caches of Upper Carboniferous terrestrial ichnofossils, including thousands of vertebrate and invertebrate trackways as well as infaunal and surface burrows. The abundance of tetrapod tracks has been the focus of extensive recent studies at UCM, with as many as six tetrapod ichnogenera being recognized (Hunt et al. Citation2010; Haubold et al. Citation2005; Hunt and Lucas Citation2005; Hunt et al. Citation2005; Martin and Pyenson Citation2005); however, the invertebrate trackways and burrows have not received as much attention (Buta et al. Citation2005; Lucas and Lerner Citation2005). Thus, the invertebrate traces from the UCM have only been given a cursory study by Lucas and Lerner (Citation2005) and are in dire need of further investigation. According to Lucas and Lerner (Citation2005) and Buta et al. (Citation2005), Kouphichnium traces are very common at UCM but are represented by a range of morphological variants, all currently assigned to K. aspodon or K. isp. (Buta et al. Citation2005; Lucas and Lerner Citation2005).
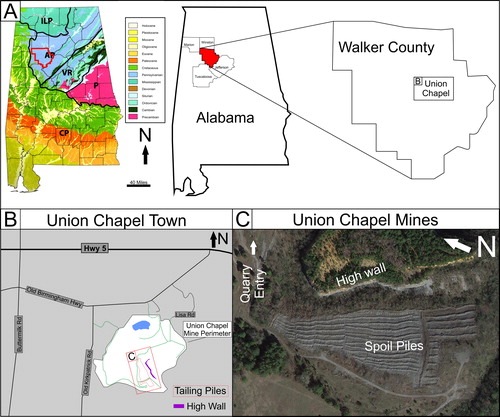
Figure 1. Location map; A) Geological map of Alabama and the site map for the Union Chapel Mine Fossil Site (modified from Buta and Kopaska-Merkel Citation2016) B) Local map for the town of Union Chapel and quarry map of the open pit site. C) Annotated Google image for the Union Chapel Mine locality (Modified from Google Earth images).
In addition to ichnofossils, the fossil record at UCM encompasses over 2000 other fossil specimens, including: allochthonous, but articulated and disarticulated plant fossils, and invertebrate body fossils (including both productid brachiopods, and rare arthropods; Wood Citation1963; Atkinson Citation2005; Dilcher et al. Citation2005; Buta and Kopaska-Merkel Citation2016). Plant fossils preserved within the UCM are thought to be detritus, transported from Pennsylvanian peat-accumulating swamps that were established inland, beyond the limits of the UCM site (Minkin Citation2005).
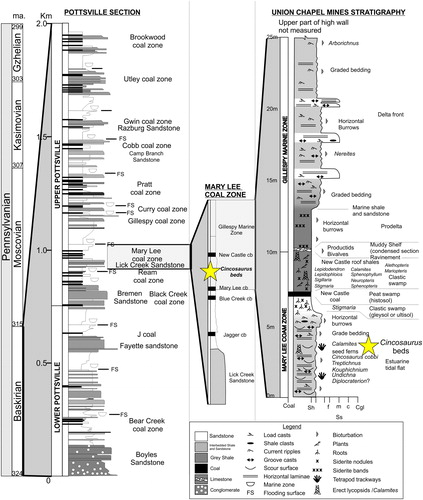
At the UCM, fossils have been collected from the spoil piles (Hooks Citation2005; ) that consist of the overburden strata of the Mary Lee Coal Bed of the Pottsville Formation (Hunt et al. Citation2005; ), making studying these traces in their precise stratigraphic context difficult (Rindsberg Citation2005). Pashin (Citation2005) has attempted to place the fossils from the spoil piles back into the strata in the high wall and has provided a preliminary interpretation of the biostratigraphy at UCM ().
Figure 2. Stratigraphic log for the Pottsville Formation and the exposed strata at the Union Chapel Mine site (modified from Pashin Citation2005). The Star indicates the stratigraphic location of the trace fossil beds.
In Alabama, Pottsville Formation trace fossils were first noted by Aldrich and Jones (Citation1930) from the Jagger Coal Seam strata in the Number 11 mine of the Galloway Coal Company near Carbon Hill, Walker County, Alabama. Aldrich and Jones (Citation1930) described a number of new ichnogenera and ichnospecies of vertebrate and invertebrate traces, including the type specimen of Bipedes aspodon that was originally, and erroneously, described as an amphibian trackway (Aldrich and Jones Citation1930). The type specimen of Bipedes aspodon was described from three individual bifid impressions on the outer edge of a broken rock slab (). The holotype of the ichnospecies (B. aspodon) conforms with the ichnogenus Kouphichnium (Minter and Braddy Citation2009) but has never been formally reassigned to that ichnogenus. Many subsequent and more complete specimens discovered from UCM are assignable to this ichnogenus but have only been informally assigned to “Kouphichnium aspodon” without a detailed ichnotaxonomic restudy and ascribed to xiphosurid trace makers (Buta et al. Citation2005). Body fossils of xiphosurids have not been found in the Pottsville Formation of Alabama, although some of its ichnotaxa have been interpreted to have been produced by xiphosurans, including: Kouphichnium, Selenichnites, Arborichnus and the “unassigned xiphosuran traces” (Buta et al. Citation2005; Lucas and Lerner Citation2005).
Figure 3. A) Kouphichnium aspodon, type specimen, AMNH 985.1.6 B) Interpretive sketch of complete trackway of type specimen, AMNH 985.1.6 C) Close-up of bifid impressions of the type specimen, AMNH 985.1.6, indicated by white arrows.

The first recognition of the ichnogenus Kouphichnium isp. at UCM was made by Mángano and Buatois (Citation2004) and in Rindsberg and Kopaska-Merkel (Citation2003). Subsequent work by Lucas and Lerner (Citation2005), in a preliminary study of the ichnotaxonomy of invertebrate traces from the UCM, also used the name Kouphichnium isp. for traces that strongly resemble morphologies described later in the same volume by Buta et al. (Citation2005) as unassigned xiphosuran traces. Buta et al. (Citation2005) were the first to informally reassign the ichnospecies Bipedes aspodon to the ichnogenus Kouphichnium, but this was done without proper systematic ichnological work. Buta et al. (Citation2005) used the ichnospecies name liberally to encompass a wide array of morphologies for any walking traces they assumed to be made by xiphosurans. An exception in Buta et al. (Citation2005) are trackways that were left, and remain, in open nomenclature and that were only identified as “unassigned xiphosuran traces”. These unassigned xiphosuran traces are associated with cubichnia traces that have been erroneously assigned to the ichnogenus Selenichnites, although actual Selenichnites do exist in the UCM ichnofaunal assemblage (Lucas and Lerner Citation2005). We leave both the cubichnia and tracks assigned to unassigned xiphosuran traces in open nomenclature, as they do not conform to the Kouphichnium ichnogenus concept and are not interpreted as having been made by xiphosurans. Thus, they will only be mentioned briefly in this manuscript, as they will be the subject of a subsequent paper.
With the large sample size of invertebrate trackways now available for study from the UCM site, the ichnogenus Kouphichnium and its ichnospecies can now be re-examined to better characterize the morphology and distinguish the various Kouphichnium ichnospecies at this site with greater confidence. In particular, the purpose of this study is to re-examine the type of B. aspodon, and the multitude of specimens that have been assigned to K. aspodon from the UCM. We here segregate specimens assigned to this ichnospecies into four Kouphichnium ichnospecies, including a previously unidentified ichnospecies at UCM (K. lithographicum); we better define the ichnospecies K. aspodon, and name two new ichnospecies – K. minkinensis isp nov., and K. atkinsoni isp. nov. Many specimens from the UCM that previous authors erroneously identified as Kouphichnium are provisionally excluded from this ichnogenus and are left in open nomenclature pending further study. Additional limulid activity at UCM has been attributed to a number of ichnogenera, including, Selenichnites, Arborichnus and unassigned xiphosuran traces (Lucas and Lerner Citation2005), that will be examined here.
Geological background and stratigraphy
Regional setting
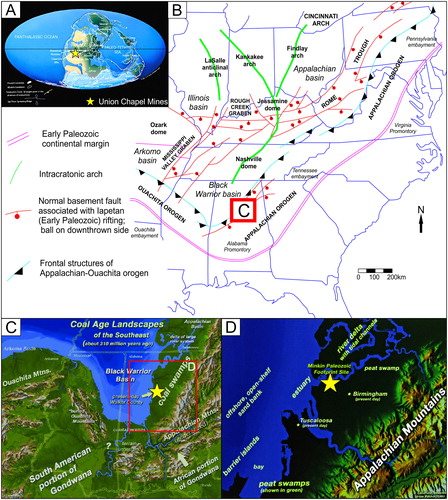
The Union Chapel Mine exposes strata of the Pottsville Formation () that have been dated to the Early Pennsylvanian (Morrowan Epoch; Langsettian Stage, Westphalian A; ) by both palynology and marine invertebrate correlations (Butts Citation1926; Eble and Gillespie Citation1989; Eble et al. Citation1991). The Pottsville Formation was deposited in the Warrior Coal Basin, which is part of the broader Black Warrior Basin, a foreland basin, that formed at the junction of the Appalachian and Ouachita orogenic belts (Pashin Citation2005; ). The siliclastic sediments of the Pottsville Formation were sourced from adjacent orogenic belts and accumulated within the Warrior Coal Basin from the Late Mississippian through the Pennsylvanian (Pashin Citation2005; ).
Figure 4. Paleogeographic location: A) Carboniferous global map (after Scotese Citation1997) B) Eastern USA Appalachian Orogen and Black Warrior Basin paleogeographical map (modified from Pashin Citation2005) C) Coal age landscape reconstruction for the Black Warrior Basin in Alabama and southeastern USA (after Buta and Kopaska-Merkel Citation2016) D) Coal age landscape reconstruction for the Black Warrior Basin in Walker County, Alabama (after Buta and Kopaska-Merkel Citation2016). Source: With permissions to reproduce by the original artist Jim Lacefield.
The sedimentology and paleontology of the Pottsville formation
The Pottsville Formation is a two-kilometer-thick heterogenous sedimentary succession composed mainly of shale, sandstone, bituminous coal and nodular limestone (Aldrich and Jones Citation1930; ). The formation is divided into upper and lower strata that are organized into “coal zones.” The 750-meter-thick, lower, sandstone-rich Pottsville Formation lacks economic coal beds, whereas the 1.25 km-thick, upper, shale-rich Pottsville Formation contains abundant economic bituminous coal that includes the Mary Lee Coal Zone exposed at the UCM. The Mary Lee Coal Zone is stratigraphically located 1 km above the base of the formation (250 meters above the base of the upper part of the formation) and is composed of four coal seams, two of which were mined at UCM; the Mary Lee and New Castle coal beds (Pashin Citation2005; ).
The upper Mary Lee Coal Zone strata, as exposed at the UCM, are 25 meters thick and are overlain by the Gillespy Marine Zone. The Mary Lee Coal Bed is directly overlain by the lower “Cincosaurus beds,” which contain all of the ichnofossils preserved at this site and other adjacent sites. The “Cincosaurus beds” preserve intervals dominated by tidal rhythmites of pinstripe-bedded mudstones, siltstone-shales and sandstone-shales that fine upwards. Individual beds range between 0.2 cm and 4 cm thick, with the trackways preserved on the tops of individual beds. These beds preserve an array of ichnofossils that include: tetrapod tracks (Haubold et al. Citation2005; Hunt et al. Citation2010), invertebrate traces (Buta et al. Citation2005; Lucas and Lerner Citation2005; Rindsberg and Kopaska-Merkel Citation2005a, Citation2005b; Lucas and Stimson Citation2013) and fish trails (Haubold et al. Citation2005; Martin and Pyenson Citation2005; ).
Tetrapod and invertebrate tracks are often associated with articulated allochthonous plant detritus as described by Dilcher and Lott (Citation2005) and Dilcher et al. (Citation2005). Pashin (Citation2005) noted the only autochthonous plants to the Cincosaurus beds were Calamites stems and a single fern stem. The paleoflora was studied by Dilcher et al. (Citation2005) and found to be similar to a previously studied, temporally equivalent paleoflora in Indiana (Wood Citation1963). The paleoflora is dominated by Calamites, lycopods and understory plants, including seed ferns and pteridosperms. Riparian plants are represented by isolated foliage, seeds and pith casts of cordaitaleans that likely occupied the hinterlands (Falcon-Lang Citation2006; Falcon-Lang and Bashforth Citation2004; Dilcher et al. Citation2005). The UCM flora has a fine preservation and a high degree of articulation that suggest the detritus was not transported far before burial (Dilcher et al. Citation2005).
The upper strata of the Cincosaurus Beds include a light gray, fine-grained sandstone that preserves wavy ripple bedforms with discoidal shale and siderite pebbles, often with the invertebrate resting trace Arborichnus repetitus. The uppermost meter of strata is a reddish-hued sandy underclay that preserves stigmarian roots below the New Castle coal beds (Pashin Citation2005) and is overlain by the Gillespy Marine Zone ().
Paleoenvironmental interpretations
During the Pennsylvanian, peat swamps accumulated in equatorial, low lying humid, tropical environments that extended from the American Midwest to Western Europe and Asia within the equatorial paleofloral belt (Gastaldo Citation1990; DiMichele and Phillips Citation1995; Dilcher et al. Citation2005; ). Paleogeographic reconstructions suggest that, during the Early Pennsylvanian, Alabama was located ∼10 degrees latitude south of the equator (Minkin Citation2005; Pashin Citation2005; ). River drainage from the Appalachian Orogen flowed northwest through peat-forming inter-distributary swamps, and through local, freshwater-dominated estuaries, preserved as mudflats, depositing heterogenous shales and sandstones of the Pottsville Formation (Ferm et al. Citation1967; Pashin Citation2005). This estuary eventually drained into the coastal shallow marine environments and offshore open waters of the Black Warrior Basin, interpreted to be normal marine as evidenced by the Gillepsy Marine Zone (Buta and Kopaska-Merkel Citation2016; ). Pashin (Citation1994a, Citation1994b) has demonstrated that marine and non-marine cycles within the Pottsville Formation are consistent with glacial eustasy and were primarily driven by polar ice sheet advance and retreat during the Pennsylvanian (Caputo and Crowell Citation1985; Frakes et al. Citation1992; Falcon-Lang Citation2003).
The Mary Lee Coal Bed represents a widespread rheotrophic peat swamp and forested wetlands dominated by arborescent lycopsids represented by the economic coal deposits of the upper Pottsville Formation. These wetlands are interpreted to have shed the articulated plants that have been transported to and preserved on the Cincosaurus beds mudflats that Pashin (Citation2005) interpreted as a freshwater, intertidal mudflat deposit, and that agrees with previous studies (Hobday Citation1974; Demko and Gastaldo Citation1996; Pashin Citation2005). The Cincosaurus beds are thought to have formed during the inundation of the Mary Lee Coal Bed during episodic tidal cycles in a mesotidal environment (Hobday Citation1974; Horne Citation1979). Evidenced by the low sulfur content, it is thought that the Mary Lee Coal Bed and the inundating waters were freshwater. However, the intercalated limestones preserve marine fauna, indicating an estuarine environment that was a lateral sedimentary facies expressed at the UCM during periods of transgressional, sea level high stand.
The tidally influenced deposits at UCM encompass a range of taphonomic conditions. The high degree of variability among the preservation of the trace fossils at UCM provides important insight into the paleoecology of coal-age Alabama (Lacefield and Relihan Citation2005) and the depositional environments that these animals were traversing (Pashin Citation2005). The exceedingly large collection also is an ideal opportunity to differentiate underprints from surface traces as well as to evaluate the extent of taphonomic variability such as extramorphological variants caused by sediment consistency, grain size and gait (Haubold et al. Citation2005; Davis et al. Citation2007), which all vary with paleoenvironmental conditions. This concept has to date only been applied at UCM to the vertebrate ichnofossils in a precursory way at the UCM.
Methods and materials
Initially examined in the field at the Union Chapel Mine site, this large collection of fossil material was collected by the Alabama Paleontology Society. These specimens were given UCM numbers before being distributed to the McWane Science Centre (MSC), Anniston Museum of Natural History (ANNMNH), the Alabama Museum of Natural History (AMNH), The Smithsonian Institution (USAM) and the New Mexico Museum of Natural History and Science (NMMNH). Museum specimens have been photographed under low angle light, using halogen work lamps. Through the use of photo-editing software and drafting software, photographs have been gray-scaled and the white balance changed to accentuate the impressions. Using freeware software (Image J), measurements of the specimens were taken from the photos. The photos from which the measurements were taken are assumed to have a 1:1 aspect ratio. Interpretive sketches were drafted using drafting software.
Supporting Information Appendices are included in this manuscript, but they are not intended to be a comprehensive list of all UCM specimens, as the dataset is too vast, and many specimens in museum collections are not yet catalogued, and many other specimens remain in private collections that are not discussed here. Those specimens listed include only those observed from photo records provided by the Alabama Paleontology Society and McWane Science Center, and those observed by the authors in museum collections. Additionally, we have included a list of those specimens that have appeared in previous publications, reassigning ichnotaxonomic affinities as needed (see Supporting Information Appendices 1–7).
Systematic ichnology
Kouphichnium Nopsca Citation19231862 Ichnites lithographicus; Oppel, (Type ichnospecies)
1923 Kouphichnium lithographus; Nopsca,
1930 Bipedes aspodon; Aldrich,
1935 Paramphibius didactylus; Willard,
1939 Kouphichnium didactylus; Caster,
1926 Micrichnus scotti; Abel,
1943 Limuludichnulus nagoldensis; Linck,
1944 Kouphichnium arizonae; Caster,
1949 Kouphichnium variabilis; Linck,
1949 Kouphichnium gracilis; Linck,
1964 Kouphichnium walchi; Malz,
1978 Kouphichnium minisculum; Actinolaza,
1985 Kouphichnium fernandez; Romano and Melindez,
2005 Kouphichnium pentapodus; Erickson,
Emended Diagnosis: Oppositely symmetrical, heteropodous and highly variable tracks that consist of up to five impressions in a track row series. Impressions consist of bifurcating, trifid or quadrifid, oval or punctate impressions together with variable secondary morphologies; with or without a medial drag impression (modified from Häntzschel Citation1975; Minter and Braddy Citation2009).
Remarks: The ichnogenus Kouphichnium is typically attributed to limulid walking activity characterized by the distinctive “pusher” impressions in addition to secondary impressions produced by the anterior walking appendages (pedipalps: Caster Citation1938, Citation1944; Linck Citation1949). Originally named by Nopsca (Citation1923) for trackways described by Oppel (Citation1862) as ichnites lithographicus, there are now 13 named ichnospecies of Kouphichnium, ranging from Ordovician (Fischer Citation1978) to Paleocene strata (Erickson Citation2005).
Ichnospecies of Kouphichnium that are currently recognized in the fossil record, include: K. arizonae (Caster Citation1944), K. didactylus (Willard Citation1935; Caster Citation1938), K. gracilis (Linck Citation1949), K. fernandez (Romano and Melendez Citation1985), K. aspodon (Aldrich and Jones Citation1930), K. walchi (Malz Citation1964), K. variabilis (Linck Citation1949), K. scotti (Caster Citation1938), K. pentapodus (Erickson Citation2005) and K. minisculum (Actenolaza Citation1978). Two ichnospecies represent cubichnia rather than walking traces: K. cordiformis (Fischer Citation1978), and K. rossendalensis (Hardy Citation1970). Although many of these ichnospecies are suspected to be junior synonyms of others, a comprehensive revision of the ichnogenus is beyond the scope of this article.
Kouphichnium lithographicum Nopsca
Figured Specimens: UCM 1094 (ANNMNH 2003.2.23), UCM 1368 (AMNH 6021), UCM 1377 (AMNH 13020), UCM 1378 (MSC 27958), UCM 4049 (MSC 25149), UCM 5015 (NMMNH P-79691) NMMNH P-69055, NMMNH P-78227 and NMMNH P-78266 (; ).
Referred Specimens: See Supporting Information Appendix 1.
Emended Diagnosis: Oppositely symmetrical, heteropodous and highly variable tracks that typically consist of a series of imprints, often arranged in an en echelon fashion of up to five imprints on each side. The imprints consist of a chevron-like series, each of four imprints that may vary in morphology, including punctate, oval or bifid “V” or “Y” shaped impressions that sometimes exhibit bifurcations (also trifid or quadrifid; occasionally on both sides of the impression), scratches or irregular impressions of similar size to one another. Impressions are often parallel to the mid-line, with the bifurcation opening posteriorly. One pair of larger and more deeply impressed bifid or trifid impressions is similar to the smaller, medial proximal impressions and is spaced at regular intervals. With or without medial (continuous or discontinuous) drag marks (Häntzschel Citation1975; Minter and Braddy Citation2009).
Description: Trackways are preserved in convex hyporelief (or concave epirelief). The trackway trajectory varies between straight to gently curved or sinuous. Outer widths vary between 9.3 mm and 23.8 mm, whereas inner widths vary from 2.4 mm to 3.4 mm. General foot morphology for the smaller imprints (pedipalps) is heteropodus, varying from punctate, to curvilinear, comma internal (external) or bifids. The pedipalp impressions are smaller and vary in size due to extramorphological distortion. The relatively larger pusher foot impressions are bifid and “Y” shaped, ranging from 1 mm to 1.9 mm in width, and 1.6 mm to 3.5 mm in length. The trackway is bilaterally symmetrical, with impressions arranged obliquely, and angled away from the midline. Pusher impressions can vary in position and distribution within a track series, depending on gait. Pusher impression width (new term: the distance between the middle of the large pusher impressions) consistently ranges from 7.6 mm to 18.5 mm. The repeat distance of the pusher footprint ranges from 3.6 mm to 12 mm. Trackways are often preserved with a telson drag that is continuous or discontinuous, however, it is not a defining feature (Caster Citation1938). Telson drags can be absent due to poor preservation, or if the specimen is likely an underprint (). Caster (Citation1938) discusses modern examples where the tail can be raised by the limulid, leaving no telson drag when the limulid traversed firm, muddy surfaces.
Figure 5. A) Kouphichnium lithographicum UCM 1377 (AMNH 13020) B) Close up of UCM 1377 (AMNH 13020) C) UCM 1378 (MSC 27958) D) UCM 5015 (NMMNH P-79691) E) UCM 1094 (ANNMNH 2003.2.23) F) UCM 1368 (MSC 27958) G) NMMNH P-78227 H) NMMNH P-78266.

Remarks: Originally named by Oppel (Citation1862) as Ichnites lithographicus, Nopsca (Citation1923) erected the ichnogenus Kouphichnium, reassigning the ichnospecies to K. lithographicum. Numerous interpretations of the ichnogenus were proposed, including footprints of pterodactyls, fish-like amphibian trackways, bipedal dinosaur tracks and the tracks of jumping mammals (e.g. Oppel Citation1862; Abel Citation1935; Willard Citation1935; Häntzschel Citation1975). Caster (Citation1938) convincingly made the connection that these complex tracks were produced by horseshoe crabs (xiphosurans). Anatomically, living horseshoe crabs have eight smaller pedipalp walking legs and a set of larger “pusher” feet, for a total of 10 appendages, with five on each side of their bilaterally symmetrical carapace (Shuster et al. Citation2003).
The Kouphichnium lithographicum specimens preserved at the UCM site are conspecific with the type (Oppel Citation1862; Minter and Braddy Citation2009). It is suspected that many of the named Kouphichnium ichnospecies are junior synonyms of the type ichnospecies; however, the revision of the ichnogenus is beyond the scope of this article. Minter and Braddy (Citation2009) commented on the ichnotaxonomic issues related to Kouphichnium. The UCM Kouphichnium specimens are assigned to K. lithographichium as they are most similar to the type ichno species in having larger bifid impressions (pusher foot) accompanied by smaller heteropodus impressions (pedipalps).
Kouphichnium aspodon Aldrich 1930 (; ; ; ; ; )
Type: AMNH 985.1.6 ()
Lectoparatypes: UCM 437 (ANNMNH 2003.2.76/74), UCM 544 (MSC 27749), UCM 662 (MSC uncatalogued), UCM 1072 (AMNH 13199), UCM 1505 (AMNH 13017), UCM 2517 (MSC uncatalogued), UCM 2716 (MSC 28058), UCM 2902 (MSC uncatalogued), UCM 5009 (NMMNH P-79689), UCM 5157(MSC 38454), UCM 5540 (MSC 38456), NMMNH P-68994.
Referred specimens: See Supporting Information Appendix 2.
Emended diagnosis: Individual impressions are small relative to the outer width of the trackway compared to other ichnospecies of Kouphichnium. All bifid impressions are of similar size, and bifurcate on one, or both ends, with no distinct, larger “pusher” impressions present. Medial telson impressions are rarely preserved, except in surface traces, or very shallow underprints. K. aspodon is distinguished from K. minkinensis based on morphology indicative of different behavior. Thus, K. aspodon represents continuous walking trackways, whereas K. minkinensis are characterized by clusters of impressions representing a jumping or hopping locomotion (see below).
Description: The type specimen was described from a partial specimen consisting of only three distinct, equal sized, well separated, distinct bifid tracks and with a depression for a pad (Aldrich 1930, in Aldrich and Jones Citation1930, 25 plate 5; ). Subsequent specimens collected from the UCM site are preserved in concave epirelief and convex hyporelief, and the trackways are linear to gently curved or sinuous. Track row series consist of up to five impressions on either side of the midline of individual heteropodus impressions, ranging from bifurcating to punctate. Specimens exhibit bifid and double bifid impressions (bifurcating on both sides, separated by a linear impression). All impressions are small relative to the outer width and are of similar size and morphology to each other, with no distinct, larger “pusher” impressions. Impressions may be oriented parallel to the midline or obliquely, often with at least one track oriented perpendicular to the midline. This impression is frequently the most distal impression (i.e. ).
Figure 6. Kouphichnium lithographicum: A) UCM 4049 (MSC 25149) B) Close up of UCM 4049 (MSC 25149) C) NMMNH P-69055.

The type specimen is the largest known representative of the ichnospecies, with impressions that measure 11 mm by 5 mm and have a repeat distance of 53.2 mm and 56.7 mm in successive impressions. In more complete but smaller examples, outer widths vary between 39.5 mm and 84.4 mm, whereas inner widths vary from 11.8 mm to 21 mm. Bifid impressions measure between 4.9 mm and 5.9 mm in length and 1.6 mm and 2.8 mm in width. Bifids have a repeat distance between 12.5 mm and 19.5 mm. The preservation of these tracks is predominantly as underprints that lack a telson drag (). Rare examples with telson drags are suspected to be surface traces assignable to K. aspodon (). In these examples, and in shallow underprints, a telson drag is observed, alongside digit drags and appendage impressions that exhibit a high degree of extramorphological variability that implies that the surface in which these traces are preserved was water saturated (). In K. aspodon, the dominant morphological feature of K. lithographicum and many other ichnospecies, the impression of the pusher foot is not present or is not larger than the walking impressions of the pedipalps. This contrasts with the more typical larger size of the pusher foot in all other Kouphichnium ichnospecies. K. aspodon exhibits appendage impressions of equal size that are morphologically similar to one another, predominantly as bifids, although some morphological variants occur.
Figure 7. Kouphichnium aspodon: A) UCM 437 (ANNMNH 2003.2.76/74) B) UCM 5009 (NMMNH P-79689) C) NMMNH P-68994.

Figure 8. Kouphichnium aspodon underprints: A) UCM 1505 (AMNH 13017) B) UCM 544 (MSC 27749) C) UCM 2716 (MSC 28058) D) UCM 2517 (MSC uncatalogued).

Figure 9. Kouphichnium aspodon: A) UCM 5540 (MSC 38456) B) Close up of UCM 5540 (MSC 38456) C) UCM 5157 (MSC 38454) D) Close up of UCM 5157 (MSC 38454).

Remarks: Kouphichnium (Bipedes) aspodon was originally described by Aldrich, in Aldrich and Jones (Citation1930), as an amphibian track. Cotton et al. (Citation1995) also apparently considered it to be a tetrapod track, as it appeared in their taxonomic list for USA Paleozoic tetrapod localities at Carbon Hill Mine, Alabama. The ichnospecies was later used as an ichnospecies of Kouphichnium and an invertebrate trackway by Buta et al. (Citation2005), but they did not conduct any systematic ichnological work. Minter and Braddy (Citation2009) list the various ichnospecies of Kouphichnium and include K. aspodon, referencing Aldrich and Jones (Citation1930), but without formal reassignment of the ichnospecies Bipedes aspodon to the ichnogenus Kouphichnium.
Rarely found with a telson drag, Kouphichnium aspodon conforms to the ichnogenus concept in that impressions are heteropodous and, in most cases, bifurcate. However, due to the lack of a larger, prominent “pusher” impression, this trackway could be produced by a different tracemaker than the limulids that are normally identified as the trackmakers of Kouphichnium. Nevertheless, due to the bifurcations of the footprints, K. aspodon was likely made by an arthropod in the same family as xiphosurids (Merostomata). The trackway itself is orders of magnitude larger than those of Kouphichnium described from elsewhere in the Carboniferous (e.g. Goldring and Seilacher Citation1971; Chrisholm Citation1983; Romano and Melendez Citation1985; Archer et al. Citation1995; Buatois et al. Citation1998, Citation2005; Prescott et al. Citation2014) and is larger than all of the known xiphosurid body fossils from the late Paleozoic (Meek and Worthen Citation1865; Copeland Citation1957; Anderson et al. Citation1997; Shabica and Hay Citation1997; Babcock et al. Citation2000; Lerner et al. Citation2016).
Indeed, the type specimen of Kouphichnium aspodon is double the size of all other known limulid trackway specimens from the Pottsville Formation. Assuming allometric growth, if the smaller, more complete specimens are scaled up to match the holotype specimen, it would suggest that the type specimen would have an outer width of nearly 100 mm in width. This is, again, significantly large than any known Carboniferous xiphosuran.
On two specimens, UCM 5540 (MSC 38456; ) and UCM 5157 (MSC 38454; ), trackways begin abruptly as elongated foot impressions. Specimen UCM 5540 (MSC 38456; ) is a short trackway that terminates abruptly as well. Both examples are preserved on large slabs that contain other ichnofossils (i.e. Diplichnites gouldi; ). The abrupt beginning and end of these trackways, with no obvious continuation, suggest that the animal was suspended in the water column before landing on the sediment surface. The abrupt end of the trackway indicates that the animal re-entered the water column, suggesting it was likely a swimmer in addition to walking along the sediment surface.
Kouphichnium minkinensis isp. nov. (; )
Holotype: UCM 227 (ANNMNH 2003.5.24).
Paratypes: UCM 508 (ANNMNH 2003.20.10), UCM 2242 (MSC 9324), NMMNH P-69078.
Referred specimens: See Supporting Information Appendix 3.
Diagnosis: Similar to Kouphichnium aspodon, the distinctive, larger “pusher” impressions, characteristic of most Kouphichnium ichnospecies, are not present, and all bifid impressions are of equal size. Bifid impressions are closely clustered together and are made up of a maximum of five individual impressions that vary in orientation. Clusters of impressions are well spaced from each other and are the distinguishing characteristic of the ichnospecies when compared to K. aspodon.
Description: Preserved in concave epirelief (or convex hyperlief), these trackways are linear to gently curved. Outer widths vary between 55.8 mm and 80.3 mm, whereas inner widths vary from 20.7 mm to 37.4 mm. The preservation of these tracks is predominantly as underprints. Kouphichnium minkinensis exhibits appendage impressions of equal size and is morphologically similar to K. aspodon, and thus similarly lacks the defining “pusher” impression that is characteristic of other ichnospecies of Kouphichnium. The “pusher” foot appendage (if present) is the same size as the smaller pedipalp walking appendage impressions. Impression dimensions measure 7.4–6.4 mm (length) by 2.9 mm (width).
Remarks: Morphologically similar to Kouphichnium aspodon, K. minkinensis consists of bifurcating and double bifurcating impressions of equal size and lacks the larger “pusher” impression. This trace is within the same size range as K. aspodon. These two ichnospecies have the potential of having a similar tracemaker and may be found in future samples in taphoseries but are distinguished based on inferred ethological differences. Thus, K. aspodon represents a walking trackway, whereas K. minkinensis represents a semi-buoyant, “bouncing” or saltation trackway, perhaps due to locomotion within a water current. K. aspodon and K. minkinensis likely represent end members of an ichnological taphoseries with a gradation between them, so that transitional trackways may be difficult to classify into one end member taxon or the other.
The difference in locomotion represents different behaviors, so the traces are separated at the ichnospecific level, as ethology is considered a major source of morphological variation that provides a good ichnotaxobase for a new ichnospecies. Telson drags have not been observed in this ichnospecies of Kouphichnium. Telson drags have, however, been observed in K. aspodon surface and shallow underprints. The suspicion that K. aspodon and K. minkinensis are produced by the same trace maker suggests that the trace maker of K. minkinensis had a telson.
Etymology: This new ichnospecies is named for the late Steven Minkin, for his extensive contributions to the science of the UCM site.
Kouphichnium atkinsoni isp. nov. ()
Holotype: UCM 2737 (NMMNH P-69048).
Lectoparatypes: MSC 28063
Referred specimens: See Supporting Information Appendix 4.
Diagnosis: Trackways with well-spaced, bifurcating appendage impressions that are long, with multiple bifurcations and trifurcations forming a dendritic morphology that distinguishes K. atkinsoni from other ichnospecies of Kouphichnium.
Description: Preserved in either concave epirelief (or convex hyporelief), trackways are linear to gently curved and may be asymmetrical. Outer widths vary between 71.3 mm and 90.2 mm, whereas inner widths are 18.6 mm to 18.7 mm. The appendage impressions are heteropodus and arranged as well-spaced, complex impressions within each trackrow series with a repeat distance of 34.4 to 51.5 mm. Impressions are longer than wide and are oriented perpendicular to the midline. Proximally, impressions begin as circular to elliptical, punctate impressions and extend out distally as a thin, linear trace that bifurcates dendritically. Impressions are 24.1 mm to 24.5 mm long and 11.1 mm to 12.1 mm wide. Often, one bifurcating impression is short, spur-like, and tapers sharply at its distal end, whereas the second bifurcation continues and bifurcates a second or third time to the distal edges of the trackway. In asymmetric examples, one track row series has the appendage impressions curved proximally towards the midline and may be entirely overturned, likely representing extramorphological distortion. Known examples of K. atkinsoni lack a medial tail drag.
Remarks: Kouphichnium atkinsoni is consistent with the ichnogeneric concept of Kouphichnium in that it consists of heteropodous trackways that include bifurcating impressions, but these are unique in that they have dendritic, bifurcating impressions. The known examples of K. atkinsoni consist of shallow underprints, as compared to other full relief Kouphichnium specimens prints at UCM that are preserved in what was a soupy substrate and that often exhibits a wide range of extramorphological variation (). The bifurcation and trifurcation of the appendage impressions is a unique morphology, and it could be argued that that this represents a new ichnogenus, However, we take a conservative approach to the ichnotaxonomy pending further discoveries.
Etymology: This new ichnospecies is named for Dr. Prescott Atkinson for his continued contributions to scientific understanding of the UCM site. The unusual morphology of the type specimen was also first recognized by Dr. Atkinson.
Selenichnites Romano and Whyte Citation1987
Selenichnites rossendalensis Hardy Citation1970 ()
Figured Specimens: UCM 107 (AMNH 5920), UCM 1612 (MSC uncatalogued), UCM 3790 (MSC uncatalogued), NMMNH P-78237, MSC 27086.
Referred specimens: See Supporting Information Appendix 5.
Emended diagnosis: Shallow, sub-oval traces occurring as isolated or in a series. Traces are semi-lunate in morphology and are generally slightly wider than long, with a rounded anterior margin and are strongly convex (in hyporelief) or concave (in epirelief) lunate or are paired crescent-shaped lobes directly anterolaterally with or without scratch marks and an anterior ridge (Romano and Whyte Citation2015; Buhler and Grey Citation2017)
Description: Five specimens have been identified from the UCM. Three specimens of Selenichnites are compound traces of Selenichnites x Cruziana isp., and Selenichnites x Dendroidichnites isp. A small specimen (UCM 107 (AMNH 5920)) described provisionally by Lucas and Lerner (Citation2005) exhibits a number of semilunate impressions preserved in convex hyporelief (). Impressions are wider than long and taper on either side posteriorly, resembling the genal angles of a trilobite cephalon. The resting trace preserves a Cruziana trackway in taphoseries with the Selenichnites trace. The Selenichnites trace is 13.5 mm wide and 7.4 mm long, with a prosomal ridge 2 mm thick. The associated Cruziana is 6 mm wide and preserves two parallel trackrows of scratch marks, each 2.3 mm wide, with a medial furrow 1.24 mm wide. The trace is 19.7 mm long. Scratch marks are oriented oblique to the midline at 67°.
Figure 10. Kouphichnium aspodon with tail drag: A) UCM 1072 (NLMNH 13199); [Counterpart UCM 1070 (AMNH 6); not figured] B) Shallow underprint of Kouphichnium aspodon with tail drag, UCM 2902 (MSC uncatalogued) C) Shallow underprint of Kouphichnium aspodon with tail drag transitioning to Kouphichnium aspodon without a tail drag, UCM 662 (MSC uncatalogued).
![Figure 10. Kouphichnium aspodon with tail drag: A) UCM 1072 (NLMNH 13199); [Counterpart UCM 1070 (AMNH 6); not figured] B) Shallow underprint of Kouphichnium aspodon with tail drag, UCM 2902 (MSC uncatalogued) C) Shallow underprint of Kouphichnium aspodon with tail drag transitioning to Kouphichnium aspodon without a tail drag, UCM 662 (MSC uncatalogued).](/cms/asset/85d4699e-80ec-441d-8bca-605e815ef2a7/gich_a_1561447_f0010_c.jpg)
On the same surface a subsequent specimen (UCM 1612 [MSC uncatalogued]; ) preserves only the semilunate headshield impression with no posterior ridge or taphoseries trackway associated. Another specimen (NMMNH P-78237; ) is larger (17.2 mm long by 24.5 mm wide) and is more typical of the morphology of other known Selenichnites specimens. The semilunate impression has an 18.3 mm long posterior ridge (telson impression). The semilunate convex impression is directly associated with Dendroidichnites-like trace with elongate, curvilinear impressions that taper laterally from a medial drag and are similar to chevron tool marks. A similar trace (UCM 3790 [MSC uncatalogued]; ) was noted to be in taphoseries with a sinuous, ribbon-like trail terminating as an elliptical, semi-lunate convex impression.
Specimen MSC 27086 () strongly resembles specimens described by Chrisholm (1985) as Selenichnites bradfordensis, as it consists of a series of semilunate impressions that overlap one another consecutively. The Selenichnites on specimen MSC 27086 is in taphoseries with a Dendroidichnites-like trackway that is similar to specimen NMMNH P-78237.
Remarks: In examples of Selenichnites from other localities, traces are often associated with Kouphichnium or other arthropod traces (cf. Romano and Whyte Citation1987). Selenichnites has classically been ascribed to xiphosuran activity whereby the prosomal head shield is projected into the sediment in a burrowing fashion (Hardy Citation1970; Romano and Whyte Citation1987; Lerner and Lucas Citation2015; Buhler and Grey Citation2017). A recent revision of the ichnogenus Selenichnites was conducted by Romano and Whyte (Citation2015) that divided it into several ichnogenera. We do not accept the subdivision here, as most of the ichnospecies they recognize are suspected to be extramorphological variants and are thus in need of an ichnotaxonomic revision beyond our scope. We thus follow the ichnotaxonomy of Lerner and Lucas (Citation2015).
This is the first published example of a Selenichnites specimen that is preserved in taphoseries with Cruziana, and the first Selenichnites associated with Dendroidichnites. It has been suggested that other Cruziana specimens at the UCM were produced by trilobites (Crimes Citation1975; Fillion and Pickerill Citation1990; Buta et al. Citation2005). If a marine incursion were present at UCM, which could be indicated by the presence of productid brachiopods (NMMNH P-69097) in the Gillespy limestone that stratigraphically overlies maximum flooding surfaces () in the exposed strata at the UCM site, then a trilobite origin is possible, especially as trilobites are known from the marine bands. This would suggest that the cephalon impression of trilobites may be responsible for some Selenichnites specimens. This is not unlike interpretations of Gibb et al. (Citation2011), where the oldest known examples of Selenichnites are of Cambrian-age from Morocco where they have been interpreted to have been made by trilobites.
Specimens NMMNH P-78237 and MSC 27086 described here are associated with a Dendroidichnites-like trace that suggests the trace was made in a soupy substrate and likely represents a surface trace. Given the poor preservation of the associated trackway it is difficult to determine the trace maker. However, some of the Selenichnites at UCM match well to the size and morphology of Carboniferous limulids and match well to the size of the Kouphichnium lithographicum at the UCM.
Although xiphosurans are the classic interpretation for Selenichnites impressions, large examples at UCM may be attributed to any other chelicerate with a semilunate cephalon, including trilobites, eurypterids or synxiphosurans, given the association with Cruziana trackways.
At UCM, underprints of cubichnia () associated with trackways previously referred to as “unassigned xiphosuran traces” () were misinterpreted as Selenichnites resting traces and, by affiliation, many of the associated trackways were erroneously labeled as Kouphichnium (Buta et al Citation2005; Lucas and Lerner Citation2005; see Supporting Information Appendix 7). These resting or colloquially named “jumper” traces () and their associated trackways do not conform to the ichnogeneric concept of Kouphichnium or Selenichnites and are thus excluded from these ichnotaxa. The traces are here left in open nomenclature pending a subsequent publication.
Figure 11. Kouphichnium aspodon underprint fallout interpretation: A) UCM 2517 (MSC) B) UCM 544 (MSC 27749) C) UCM 437 (ANNMNH 2003.2.76/74) D) UCM 662 (MSC uncatalogued) E) UCM 1072 (AMNH 13199).
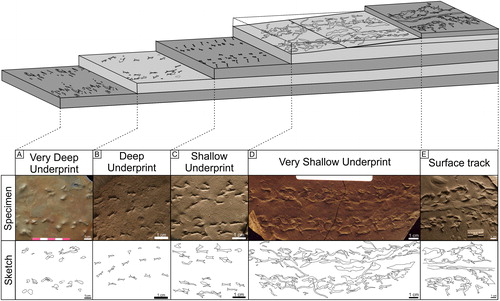
Figure 12. A) Kouphichnium minkinensis, type specimen UCM 227 (ANNMNH 2003.5.24) B) UCM 2242 (MSC 9324).

Figure 13. Kouphichnium minkinensis: A) UCM 508 (ANNMNH 2003.20.10) B) NMMNH P-69078.

Lerner and Lucas (Citation2015) discuss Selenichnites bradfordensis from specimens recovered from New Mexico. In their paper, S. bradfordensis is considered a poorly preserved S. rossendalensis due to similarities to specimens preserved on the type specimen described by Hardy (Citation1970). We here agree with Lerner and Lucas (Citation2015) and consider this ichnospecies a minor morphological variation on the type ichnospecies and consider specimen MSC 27086 to be an extramorphological variation of Selenichnites rossendalensis.
Ichnogenus Arborichnus Romano and Meléndez, 1985
Arborichnus repetitus Romano and Meléndez, 1985()
1985 Arborichnus repetita: Romano and Meléndez,
fig. 1, pls.1, pls.2, fig. 3,
2005 Arborichnus repetita: Lucas and Lerner,
p. 150, fig. 2F.
2005 Arborichnus repetitus: Buta et al., pls. 103A-B,
104A-B, 105A-B, 106A-B.
2013 Arborichnus repetitus: Buta et al., pls. 3A-B.
2016 Arborichnus repetitus: Buta and
Kopaska-Merkel, fig. 16.11.
Figured Specimens: UCM 4646 (MSC 25968), UCM 3549 (MSC 27905).
Referred specimens: See Supporting Information Appendix 6.
Emended diagnosis: Sets of up to five pairs of symmetrical scratch marks. At the anterior end of the set, the scratch marks are short and directed inward and backward; the posterior scratches are aligned at about right angles to the mid-line of the set. Scratch marks are repeated as a track with the distance between the sets being approximately equal to the length of the set.
Description: Numerous UCM specimens are bilaterally symmetrical traces that consist of up to five sets of paired scratch marks (grooves in concave epirelief; or as paired ridges in convex hyporelief) that are oriented perpendicular to the midline. These traces flank a central, oval-shaped disturbed area. Impressions are straight to strongly curved and splay outwards from the oval-shaped center, with the posterior pairs of grooves being slightly longer in some specimens (Buta et al. Citation2013; Kopaska-Merkel and Buta Citation2013). Spacing between sets is approximately equal distance apart. These distinctive traces closely resemble the original diagnosis of Arborichnus repetita by Romano and Melendez (Citation1985).
Remarks: Arborichnus was not formally described until Romano and Melendez (Citation1985), although it had been known for a considerable time (see Caster Citation1938, fig. 1, plate 9, figs. 3 and 4). Other than Romano and Melendez’s (Citation1985) type descriptions from the Carboniferous of Spain, there are few recent reports of this ichnotaxon in the literature (Buta et al. Citation2005; Lucas et al. Citation2005; Lucas and Lerner Citation2005; Mansky and Lucas Citation2013; Buta et al. Citation2013; Buta and Kopaska-Merkel Citation2016). The large number of UCM A. repetita specimens, which number in the hundreds, provides an extensive sample with which to broaden our understanding of this trace and further clarify the taxonomy of this ichnogenus.
Specimens from the Crescent Valley Mine described by Buta et al. (Citation2013) in the Pottsville Formation were interpreted to have been made by xiphosurans and represented both swimming and grazing behavior in more distal facies in deep waters (Buta et al. Citation2013). A re-description of specimens from the UCM (Buta and Kopaska-Merkel Citation2016) has led them to suggest these traces were not made by xiphosurans, as was previously thought (Romano and Melendez Citation1985; Buta et al. Citation2005; Lucas and Lerner Citation2005). Buta et al. (Citation2013) and Buta and Kopaska-Merkel (Citation2016) have taken a conservative approach and interpreted Arborichnus as the resting traces of decapod crustaceans but admitted that the trace maker was probably not a horseshoe crab.
Discussion
Kouphichnium tracks were originally attributed to an array of possible trace makers, including: amphibians, bipedal dinosaurs, pterodactyls, reptiles and jumping mammals or other precursor tracemakers (Oppel Citation1862; Nopsca Citation1923; Aldrich and Jones Citation1930; Abel Citation1935; Häntzschel Citation1975). Caster (Citation1938) was the first to theorize and compare modern limulid footprints to Kouphichnium trackways preserved in the fossil record. It was then confirmed as the trace maker when a xiphosurid Mesolimulus was discovered at the end of a Kouphichnium trace in the famous Jurassic limestones of Solnhofen, Germany (K. walchii: Malz Citation1964). Thus, limulids as the tracemaker for Kouphichnium trackways has been widely accepted in the literature. The pusher foot is interpreted as the largest and deepest impression that occurs in a Kouphichnium trackway, whereas the smaller pedipalps produce a range of different and smaller morphologies, ranging from punctate to bifid (Caster Citation1938, Citation1944; Linck Citation1949).
Trace fossil interpretations can have profound implications for the understanding of Carboniferous paleoevironments (e.g. Calder Citation1998; Buatois et al. Citation2005; Falcon-Lang et al. Citation2006). Modern day limulids are marine organisms that periodically migrate into brackish and freshwater shorelines to mate and reproduce. Xiphosurids and their ancestors date back to the early Paleozoic (Rudkin et al. Citation2008). In the Pennsylvanian, only three genera of limulids are well documented: Euproops, Paleolimulus and Belinurus. All have been recorded from classic Carboniferous sections such as Mazon Creek, Illinois (Nitecki Citation2013), Joggins, Nova Scotia (Calder Citation1998; Falcon-Lang Citation2006; Stimson and MacRae Citation2010; Buhler and Grey Citation2017), West Bay, Nova Scotia (Copeland Citation1957) and Sydney, Cape Breton, Nova Scotia (Copeland Citation1957; Calder Citation1998). Historically, reconciling Carboniferous localities and marine xiphosurids has posed challenges where classic Carboniferous localities such as Joggins and Horton Bluff were interpreted as freshwater environments. As a result, many of the associated xiphosurids were interpreted to be fresh water organisms (Moore Citation1955). New data from historical localities (i.e. Joggins, Nova Scotia; Grey et al. Citation2011; Horton Bluff, Nova Scotia; Mansky and Lucas Citation2013) imply a distal marine connection, that may change our interpretation of Carboniferous xiphosurid ecologies.
Currently at the UCM, xiphosurid body fossils have not been found. However, the presence of Kouphichnium lithographicum implies xiphosurans were present at the UCM location. Unlike classic sections such as Joggins, the marine connection here is not ambiguous, as local limestones representing sea level high stands contain productid brachiopods (NMMNH P-69097) that are interpreted as being derived from marine environments (). The depositional environments of the trace-fossil-bearing strata are thought to be estuarine, freshwater-dominated mudflats with a marine connection. Thus, the trace makers for Kouphichnium at the UCM were likely xiphosurids or other similar, euryhaline chelicerates.
Taxonomy and taphonomy
The majority of traces found at the UCM of both vertebrates and invertebrates are underprints (Goldring and Seilacher Citation1971; Haubold et al. Citation2005). Traces on the walking surface are present, but they are much less common and are preserved with a high degree of extramorphological distortion due to the water-saturated substrate (; ; and ). In rare examples of surface tracks made by tetrapods, the shallow underprints are plantigrade, while the surface traces are highly distorted, with extramorphological digit extraction and elongated drag impressions, sometimes resembling Dendroidichnites (, P-43509). This variance is interpreted to be due to water saturation in the surface layer at the sediment-water interface that is influenced by inundating tidal waters, while the more cohesive subsurface sedimentary layers preserved a higher degree of detail in the trackways that penetrate the surface layers (commonly they preserve appendage impressions but no telson drag). The sediment was predominantly mud or very fine-grained sand or silt, periodically inundated by water, while the underlying layers were more compacted in the subsurface. Previous workers (Rindsberg Citation2005) have noted that there is no direct sign of prolonged subaerial exposure (i.e. mudcracks, rain pit impressions, ripple bedforms), suggesting that either the traces observed at the UCM are all underprints, and the walking surface is rarely encountered, or the sedimentary surface was at no point subaerially exposed or it was subaerially exposed for a very short time. However, there are no tetrapod swim tracks, so if the sediments were perpetually submerged, they were only submerged by a shallow film of water or not exposed long enough to desiccate.
Figure 14. A) Kouphichnium atkinsoni, type specimen NMMNH P-69048 B) MSC 28063.

Figure 15. Selenichnites rossendalensis: A) UCM 107 (AMNH 5920) B) Close-up of UCM 107 (AMNH 5920) C) UCM 1612 (MSC uncatalogued) D) NMMNH P-78237) E) UCM 3790 (MSC uncatalogued) F) MSC 27086.

Biofilms have been interpreted at the UCM based on the presence of microbial bubbles associated with trackways (Rindsberg Citation2005). Biofilms allow for the preservation of fine details in fossil trackways and can stabilize the sediments by early cementation (Seilacher Citation2008; Marty et al. Citation2009). Generally, a clear print will occur after the appendage has passed the initial layer of soft, water saturated sediment and made contact with the first compacted layer in the subsurface, which, in turn, allows for a high preservation potential, while the unconsolidated sediments at the surface are mobile, with poor preservation potential (). At the UCM, the high-resolution traces preserved as shallow underprints decrease in detail with depth (Lucas and Lerner Citation2005; Buta and Kopaska-Merkel Citation2016; Stimson and MacRae Citation2010; Milàn and Bromley Citation2007; ). Evidence of this underprint fallout has been noted (Kopaska-Merkel and Buta Citation2013; Buta and Kopaska-Merkle Citation2016) before at the UCM site and can be seen in the preservation of “Cincosaurus” tracks (P-43509, ), as the walking surface and consecutive layers preserve such phenomena.
Trace makers
Kouphichnium tracks at the UCM are associated with both aquatic (Undichna) and terrestrial (tetrapod track) ichnofossils on different surfaces, and all ichnofossils occur in association with infaunal and surface burrows (). This suggests that the tracemaker of Kouphichnium was tolerant of both subaquatic and subaerial conditions.
Figure 16. Unassigned “jumper” tracks: A) “jumper trace” associated with unassigned “xiphosurid tracks”, UCM 5000b (MSC 38462) B) close up of UCM 5000b (MSC 38462).

It is likely that xiphosurids are the tracemaker of most Kouphichnium based on the behavior of modern day limulids. Modern day horseshoe crabs have a high salinity tolerance for both fresh and salt water environments (Chatterji and Abidi Citation1993). They live the majority of their lives in normal marine water, and return to the shoreline where they were born, to reproduce. Reproduction occurs under specific sedimentary conditions in freshwater or brackish, low energy environments, such as Delaware Bay (Shuster and Botton Citation1985) and is comparable to the freshwater-dominated environment at the UCM.
Given the high preservation of articulated plant fossils (organic matter) at the UCM, if xiphosurids inhabited the shorelines, bodies and molts should populate this environment as well. However, no evidence of limulid body fossils or molts have yet to be found at this site, suggesting that migration to this brackish environment could be interpreted as temporary, comparable to modern day limulid ecologies (Shuster and Botton Citation1985). Alternatively, taphonomic factors, such as sediment rates, could have precluded the burial and preservation of limulid body fossils.
Kouphichnium has traditionally been interpreted to be of limulid origin. However, four distinguishable ichnospecies have been identified at Union Chapel Mine, and likely represent at least three different trace makers discussed below.
Kouphichnium lithographicum
Classic Kouphichnium lithographicum morphologies are preserved at the UCM. However, this ichnospecies at the UCM is smaller than its Jurassic holotype specimen (Oppel Citation1862; Nopsca Citation1923; Abel Citation1935; Caster Citation1938, Citation1944; Malz Citation1964). Previous work has misidentified some small scale Kouphichnium traces at UCM as Paleohelcura and Stiaria (Buta et al. Citation2005; Buta and Kopaska-Merkel Citation2016; Supporting Information Appendix 7), which would have suggested arachnid trace makers such as spiders or scorpions. Larger examples of real Paleohelcura and Stiaria are known from the UCM (Buta and Kopaska-Merkel Citation2016) but are not discussed further here.
Specimens of Kouphichnium lithographicum from the Jurassic type locality at Solnhofen are found in association with the trace maker Mesolimulus. Limulids from the Mesozoic to present day are considerably larger than those from the Carboniferous and Permian (Copeland Citation1957; Haug et al. Citation2012; Lerner et al. Citation2016). The only known genera of horseshoe crabs in the Carboniferous are Paleolimulus and the poorly distinguished Euproops and Belinurus (Haug et al. Citation2012), known from various Carboniferous localities (Copeland Citation1957; Calder Citation1998; Falcon-Lang Citation2006; Mansky and Lucas Citation2013; Nitecki Citation2013; Lerner et al. Citation2016). They are considerably smaller than their younger limulid descendants (i.e. Mesolimulus: Malz Citation1964), and this accounts for the size difference between Mesozoic and Paleozoic Kouphichnium examples, as seen here. Large xiphosurans (> 5 cm) are known from older strata (Rudkin et al. Citation2008) but are not yet known from the Carboniferous. The smaller examples of K. lithographicum found at the UCM have a width similar to that of the known limulids of the Upper Carboniferous and exhibit the diagnostic larger “Y” shaped pusher impressions that distinguish them from Stiaria and Paleohelcura (Minter and Braddy Citation2009).
The examples at UCM are often preserved as light surficial tracks () that do not have deep underprints. If underprints occur they are relatively shallow (sub-mm) compared to the trackway surface, and generally lack a telson drag, with reduced resolution of the bifid-trifid pusher and pedipalp impressions ().
Figure 17. Unassigned “xiphosurid” tracks: A-B) UCM 1752 (AMNH 13108; counterpart to UCM 1740 (AMNH 6003) C) UCM 1119 (AMNH 5950) D) UCM 723 (AMNH 5959).

Kouphichnium aspodon
Kouphichnium aspodon has been identified from the UCM and other outcrops of the Pottsville Formation in Alabama, including its type locality at the Carbon Hill Mine, and the recently described Crescent Valley Mine (Buta et al. Citation2013). It is also known from the Mississippian (Visean) Mauch Chunk Formation of Pennsylvania (Fillmore et al. Citation2012). With re-examination and re-description of the ichnospecies at UCM, the Mauch Chunk examples will require re-examination, but this is beyond the scope of this manuscript.
Kouphichnium aspodon trackways are larger by an order of magnitude, with deeper impressions, than all other Paleozoic Kouphichnium specimens described previously (Buta et al. Citation2005; Lucas and Lerner Citation2005; Buta and Kopaska-Merkel Citation2016). K. aspodon differs from the K. lithographicum ichnospecies concept as it lacks a larger “pusher” impression. Additionally, the bifid impressions are double bifid impressions with a bifurcation on either end and are all equal in size but vary in length and orientation. In previously described underprint fallout studies of Kouphichnium, the pusher impressions are the deepest impressions, with the resolution of the smaller pedipalp impressions lessening with depth and the loss of a medial telson drag (Goldring and Seilacher Citation1971; Stimson and MacRae Citation2010). In the case of K. aspodon, if the pusher foot was anatomically present on the trace maker, it should be observed as the deepest impression in all K. aspodon examples (; ; Supporting Information Appendix 2). However, in the rare surface traces () and underprints (that comprise the majority of encountered specimens; ) the bifid impressions are the same size, and no pusher impression is observed, suggesting that they were produced by a tracemaker other than a limulid.
Also notable is that a shallow telson drag impression is only preserved on the rare surface specimens where the trackways exhibit a high degree of extramophological distortion, likely due to highly water-saturated sediments, but they still do not exhibit pusher feet morphologies. Telson drags do not appear to be present in any known examples of underprint fallout (; ). However, it was also noted by Caster (Citation1938; plate 13, fig. 7) that when modern Limulus polyphemous walked on stiff mud surfaces, the telson was raised off the sediment surface, leaving no impressions. The animal walked on the blade-edged pedipalps and pusher appendages, producing comma-like impressions. The pusher impression was still recognizable, so the trackway differs from Kouphichnium aspodon in that way.
The Carboniferous Period was inhabited by a number of arthropods that are known to have traversed both freshwater and marine environments and frequented brackish-euryhaline paleoenvironmental niches (Kjellesvig-Waering Citation1964; Buatois et al. Citation2005; Tetlie Citation2007). The oldest chelicerates known to have lived in euryhaline environments, at least habitually for mating and molting, are the eurypterids (Braddy Citation2001; Vrazo and Braddy Citation2011), though xiphosurans are now known from the Ordovician (Rudkin et al. Citation2008). A number of Carboniferous eurypterid groups are known in the Pennsylvanian, belonging to the Hibberopteridae and Adelophthalmidae (Tetlie Citation2007). These eurypterid families include both walking and swimming forms that encompass an array of morphologies that likely reflect different paleohabitats. Within these two groups, there is a wide diversity of eurypterids that are found in shallow marine, estuarine, fluvial or lacustrine environments, as well as representatives that were able to make short terrestrial excursions (Braddy Citation2001; Tetlie Citation2007; King et al. Citation2017). This would allow for a greater diversity of size and appendage morphology, so that Kouphichnium aspodon may have been made by eurypterids or synxiphosurids, though eurypterid walking traces are typically assigned to the ichnogenus Palmichnium (Briggs and Rolfe Citation1983; Minter and Braddy Citation2009).
At present, no eurypterid body fossils, molts or cuticle have been discovered at the UCM, and there is no direct evidence of paddle appendages associated with Kouphichnium aspodon specimens (Vrazo and Ciurca Citation2017). However, it was Buta et al. (Citation2013) who first suggested that Kouphichnium traces at both UCM and the Crescent Valley Mine may have been produced by a more diverse group of arthropods than simply xiphosurans. The broader group Merostomata was suggested and includes chelicerates such as eurypterids.
Close relatives to the xiphosurans are the synxiphosurans (Moore et al. Citation2007). They are morphologically similar to xiphosurans, but, whereas they lack the pusher foot, these animals could easily produce traces similar to Kouphichnium aspodon. Synxiphosurans are well known in rocks of Silurian through Devonian age, but they are rare in rocks of the Carboniferous (Moore et al. Citation2007). Only a single example of a Carboniferous synxiphosuran has been reported in the fossil record and is of Namurian (Serpukhovian) age (Moore et al. Citation2007). This rare example extended the fossil record of synxiphosurans into the Carboniferous, thus synxiphosurans are a plausible trace maker of K. aspodon.
Kouphichnium minkinensis
Morphologically, the individual impressions of Kouphichnium minkinensis (; ) resemble those assigned to K. aspodon, however, they differ in ethology. Thus, the trace exhibits well-spaced clusters of impressions that suggest a major change in gait and activity. It is possible that the animal was using the water currents to move, being suspended buoyantly in the water, touching down on the surface due to gravity and providing thrust and directionality for the next leap at regular intervals. Given that the specimens preserve underprints, sedimentary structures that would be at the sediment-water interface are not observed to confirm the movement of water. However, the “Cincosaurus beds” do exhibit tidal rhythmites, which suggests there were bidirectional currents. It is thus possible that the trace maker was moving with either an incoming or an outgoing tidal current aiding its locomotion.
Figure 18. Unassigned “xiphosurid” tracks: A) UCM 1740 (AMNH 6003) B) UCM 1738 (MSC uncatalogued) C) UCM 722 (MSC 27764) D) UCM 1748 (AMNH 6014) E) UCM 1356 (MSC 27781) F) UCM 1414 (MSC uncatalogued) G) UCM 1781 (AMNH 6012).

Figure 19. Arborichnus repetitus: A) UCM 4646 (MSC 25968) B) UCM 3549 (MSC 27905).

Invertebrate ichnotaxonomy is based on the ethology (the behavior) of a trace fossil as opposed to the taxonomy of the trace makers and extramorphological variation within a single ichnotaxon. Minter and Braddy (Citation2009) discussed the difference between major and minor morphological variation. Major morphological variation includes ethological differences that are considered to be useful as an ichnotaxobase, whereas minor morphological variants such as gait variants are not. The morphological differences between Kouphichnium minkinensis and K. aspodon are likely changes in the locomotion style rather than gait, but no taphoseries has yet been noted. Despite morphological similarities in the individual appendage impressions, the mode of locomotion is different in K. minkinensis compared to K. aspodon, suggesting an ethological variant. K. aspodon represents a walking mode of locomotion as opposed to K. minkinensis, which represents a saltation mode of locomotion whereby the tracemaker undertook semi-buoyant locomotion while being carried in a current, periodically touching the bottom with all of its appendages. This behavioral difference dictates the morphological difference and what distinguishes the two ichnospecies. Thus, eurypterids, synxiphosurans or other chelicerates, as noted above for K. aspodon, are trace maker candidates.
Kouphichnium atkinsoni
Kouphichnium atkinsoni follows the ichnogenus concept in that the impressions are heteropodous and bifurcating. However, its dendritic morphology with multiple bifurcations per imprint is not similar to any other known ichnospecies of Kouphichnium, thus justifying a new ichnospecies. Two specimens of this ichnospecies have been found at the UCM and are among the largest invertebrate trackways currently known at this site (; Supporting Information Appendix 4). K. atkinsoni has an outer width that is approximately 9 cm, which is larger than other Carboniferous Kouphichnium specimens and approximately 30% larger than most K. aspodon and K. minkinensis, with no growth series between them. The size is comparable to the type specimen of K. aspodon and likely represents a similar walking behavior.
Figure 20. Paleoenvironmental reconstruction and interpretation of the ichnofaunal assemblages: A) paleoenvironmental reconstruction (modified from Pashin Citation2005) B) Low tide ichnofossil assemblage C) Rising and falling tide ichnofossil assemblage D) High tide ichnofossil assemblage (based on original work by Minkin Citation2005).
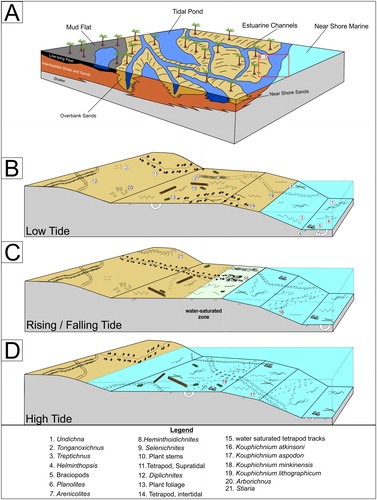
Kouphichnium atkinsoni is a shallow underprint that preserves a series of dendritic, elongate appendage impressions with repeated bifurcations. These appendage impressions were likely flexible, as the trackway is often asymmetrical, with one side preserving appendage impressions that are medially curved backwards. Similar to the other large arthropod trackways of K. aspodon and K. minkinensis, these tracks are interpreted to represent underprint fallout due to the detailed and clear impressions seen in other examples of both invertebrate and vertebrate trackways where both surface and underprints are preserved in taphoseries. By comparison, these trackways are interpreted to be made below the water-surface interface and not in the water-saturated surface sediment. This ichnospecies currently has not been discovered with a medial telson impression, which suggests either that this is an underprint and a medial telson was not preserved in the underprint fallout, or that the trace maker did not have a telson.
Kouphichnium atkinsoni is not likely produced by a limulid due to its size being orders of magnitude larger than that of the known limulids of the Upper Carboniferous. The bifurcating impressions suggests a chelicerate arthropod, but some of the bifurcations may have been produced by other segmentations or spines along the jointed appendages. Similar to both K. aspodon and K. minkinensis, this trace may have been produced by a eurypterid or some other unknown chelicerate arthropod. The brackish conditions limit possible trace maker candidates for K. atkinsoni, and limulids are apparently excluded based on trackway size. Eurypterids and synxiphosurans remain the most likely candidates.
The unique morphology of Kouphichnium atkinsoni adheres to the broad ichnogenus concept of Kouphichnium, justifying a new ichnospecies. However, the dendritic nature of this trace has not been seen before in any known arthropod trackways and could be segregated in the future with more discoveries and novel specimens, perhaps in full relief, to justify a new ichnogenus. We take a conservative ichnotaxonomic approach here, given that only two specimens of K. atkinsoni have been found at the UCM.
Other xiphosuran traces
Although Kouphichnium is typically associated with xiphosuran activity, other examples of xiphosurid ichnofossils are also present at the UCM. The ichnogenus Selenichnites was first described by Hardy (Citation1970) as semi-lunate impressions associated with Kouphichnium walking traces. These traces were interpreted as being the result of the prosoma (head shield) of the horseshoe crab plowing into the sediment in a digging fashion. Similar traces (Limulicubichnus rossendalensis) were also noted by Miller (Citation1982), although more tear-drop shaped rather than semi-lunate, that were also interpreted as the digging activity of limulids. These cubichnia resemble traces noted in the Solnhofen limestones associated with Kouphichnium tracks and Mesolimulous body fossils at the ends of the trackways as mortichnia. Selenichnites traces at the UCM are rare and are only described from five known samples (; Supporting Information Appendix 5). Specimen UCM 007(2005.7.32) exhibits a unique co-occurrence of Cruziana terminating in a Selenichnites resting trace. Cruziana is typically attributed to the walking and plowing traces of trilobites, as mentioned in previous work at the UCM (Buta and Kopaska-Merkel Citation2016; Lucas and Lerner Citation2005) but are here associated with a resting trace typically attributed to limulids. This suggests that with varying taphonomic conditions, Cruziana may be produced by various arthropods that include trilobites but may be produced by other invertebrates, apparently including xiphosurans, or that Selenichnites may be produced by other arthropods in addition to limulids, such as trilobites or eurypterids.
Additional specimens of Selenichnites from the UCM are larger and associated with Dendroidichnites-like trackways. Although typically preserved at other localities as either isolated impressions (underprints) or associated with Kouphichnium traces (Lerner and Lucas Citation2015; Buhler and Grey Citation2017), this is the first association of Selenichnites with Dendroidichnites. Dendroidichnites has been discussed by Davis et al. (Citation2007) as being the result of a water-saturated substrate where details of trackways are not preserved and can thus be the product of a range of animals rather than attributed to a single biotaxon. It is thus not beyond possibility that these Dendroidichnites traces, where they are associated with Selenichnites, may be the soupy substrate product of xiphosuran, trilobite, synxiphosuran or eurypterid activity.
Arborichnus is a common trace fossil in the upper strata of the UCM mine and is typical of the sandy facies (). First described by Romano and Melendez (Citation1985), Arborichnus was interpreted as being the product of limulid activity. Arborichnus typically has four to five linear impressions interpreted as leg impressions on either side of the bilaterally symmetrical medial impression. Previous workers at the UCM have attributed these to limulids. Buta et al. (Citation2013) suggested that Arborichnus is a resting or feeding trace produced by an invertebrate. Recent work by Buta et al. (Citation2013) and Buta and Kopaska-Merkel (Citation2016) has suggested this was not likely made by a limulid, as previously thought. They suggested that the Arborichnus specimens at the Crescent Valley Mine were likely produced by a decapod crustacean, although they also concluded that the trace maker is unknown. Regardless, the traces were imprinted on the tops of a sandy substrate that was deposited under high energy conditions, such as storm deposits (Buta and Kopaska-Merkel Citation2016). Given the reinterpretation of Kouphichnium aspodon as potentially being made by eurypterids, and Selenichnites associated with Cruziana traces that may be of trilobite origin, any of the decapods may be the trace maker of Arborichnus. Additional body fossils of a spider-like insect have been found at the UCM (Buta and Kopaska-Merkel 2005) that are also plausible candidates.
Figure 21. Underprint fallout of a Cincosaurus trackway, UCM 1708 (NMMNH P-43509).

Paleoecological interpretation
The paleoenvironment at the UCM was interpreted by Pashin (Citation2005) to be an estuarine tidal flat, thought to have been deposited within a deltaic system at the mouth of a large river that was tidally dominated (). The paleoecology of the animals that inhabited this tidal flat are inferred almost entirely from ichnofossils, as invertebrate body fossils are rare at UCM (Atkinson Citation2005; Minkin Citation2005; Buta and Kopaska-Merkel Citation2016). All of the known ichnofossils are preserved within the finely interbedded tidal rhythmites and are interpreted to represent alternating subaquatic and subaerial ichnofossil assemblages (Pashin Citation2005; ). A lack of rain impressions and desiccation features suggest that the substrate was permanently wet or submerged (Rindsberg Citation2005). Estuary conditions are typically hostile environments for most life, as the mix of fresh and salt water excludes many groups of animals except those euryhaline taxa tolerant of both salinity extremes. As discussed above, a number of invertebrate taxa are tolerant of these conditions, in addition to terrestrial taxa that may occupy this niche during subaerial exposure.
Observations in this study suggest that at the UCM many invertebrate and vertebrate traces were being made contemporaneously. Terrestrial trace fossils (tetrapod trackways: Haubold et al. Citation2005; and terrestrial arthropod trackways: Lucas and Lerner Citation2005) were produced under subaerial conditions on emergent muddy freshwater-dominated estuarine tidal flats during falling or low tides (Minkin Citation2005; Dilcher et al. Citation2005). Previous workers inferred that tetrapods were feeding on intertidal invertebrate faunas that may have sustained populations of juvenile amphibians and amniotes (Rindsburg et al. Citation2004; Martin and Pyenson Citation2005), as invertebrate traces are common within the Cincosaurus beds (Martin and Pyenson Citation2005). Abundant infaunal burrows (Treptichnus, Planolites, Cochlichnus, Helminthoidichnites, Heminthopsis and Gordia) are present in both ichnofossil assemblages, and represent an intertidal invertebrate fauna (Rindsberg and Kopaska-Merkel Citation2005a,Citationb; ). Subaquatic traces such as Undichna were made during high tides (). Subaquatic and subaerial traces are separated stratigraphically at millimeter scale, with the exception of underprint fallout of larger, often tetrapod trackways and infaunal burrows that penetrate through multiple strata. The tetrapod tracks do not show evidence of swimming, suggesting that they were deposited during low tides under subaerial or near subaerial conditions. Undichna (fish trails) are found on bedding planes that lack tetrapod footprints, suggesting that fish and tetrapod traces were made under different depositional conditions (i.e. subaerial vs subaqueous; Pashin Citation2005). The Undichna traces were deposited during high tide as well as during rising or falling tidal cycles (Martin and Pyenson Citation2005; Minkin Citation2005). It has been interpreted that fish traces (Undichna) were made during falling tides, while tetrapod and some invertebrate trackways were made on emergent mudflats (Rindsburg et al. Citation2004; Haubold et al. Citation2005; Lucas and Lerner Citation2005; Martin and Pyenson Citation2005; Pashin Citation2005). The Undichna represent distal faunal elements that ephemerally occupied the tidal flat niche during subaquatic times. Similarly, tetrapod and terrestrial invertebrate traces represent proximal faunal elements that only occupied the tidal flat niche during times of subaerial exposure. The infaunal burrows are present in all strata and represent interstitial dwelling animals, while many euryhaline invertebrates interpreted from Kouphichnium trackways are also present under all tidal conditions when present.
Conclusions
The importance of paleoichnology to the understanding of paleoecology and paleo-communities is reinforced by studies at the UCM within transitional marine-freshwater environments. The UCM habitat was a biologically diverse ecosystem, and continued research will shed more light on the intricate interplay of paleoecologies at a marginal terrestrial-aquatic estuarine paleoenvironment. The paleoecology of the animals that inhabited this paleoenvironment is inferred almost entirely from ichnofossils. Sites preserving both body fossils and tracks are rare, likely due to taphonomic differences, making understanding the tracemakers at fossil sites dominated by ichnofossils, such as the UCM, important.
Although a comprehensive revision of the ichnogenus Kouphichnium is in dire need (Buatois et al. Citation1998), this article assesses the ichnospecies assigned to Kouphichnium at the UCM. Previous to this study, only Kouphichnium isp. and Kouphichnium aspodon were noted, and the ichnogenus was used liberally, encompassing a wide range of morphologies and a number of specimens that were erroneously assigned to this ichnogenus (See Supporting Information Appendix 7). These previously unassigned xiphosurid traces are here noted and are excluded from the ichnogenus Kouphichnium, leaving them in open nomenclature, pending further investigation.
In part, the ambiguity surrounding Kouphichnium aspodon has been derived from an incomplete holotype specimen. Subsequently collected examples from the UCM have shed new light on and allowed for a better ichnological diagnosis of the ichnospecies here assessed to be valid ichnotaxa. K. lithographicum has been assigned to specimens previously misinterpreted as Paleohelcura trackways. Consequently, from this detailed study of K. aspodon, two new ichnospecies of Kouphichnium are described (K. atkinsoni and K. minkinensis). K. atkinsoni loosely fits the ichnogeneric concept of Kouphichnium, but additional specimens may reveal that it is unique enough, with its dendritic bifurcating tracks, that it should be segregated as a separate ichnogenus. With only two specimens encountered in this study, we conservatively assign the new ichnospecies to Kouphichnium ().
The wide range of morphological variation preserved at the UCM allows a better understanding of the underprint fallout. Sediment water-saturation affects the overall morphology and taphonomy of some Kouphichnium ichnospecies and has wider implications for the taphonomy of the ichnology at UCM (). In this study, K. aspodon, K. minkinensis and K. atkinsoni are interpreted to not have been made by xiphosurans based on the lack of pusher appendage impressions. Eurypterids appear to be the most likely candidates given the paleoecological conditions and similarities in both size and morphology of the imprints to the chelicerate anatomy of known Carboniferous eurypterid groups. Furthermore, they follow an ethology similar to known eurypterid ecologies from older strata (Braddy Citation2001; Tetlie Citation2007; Vrazo and Braddy Citation2011). Through this study, we have suggested that some Kouphichnium ichnospecies may be produced by more than just limulids, and that other chelicerates should be considered as potential trace makers. This has broader implications for paleoenvironmental studies when ichnofossils are heavily relied on for paleoenvironmental and paleoecological interpretations.
Supplemental Material
Download MS Word (42.4 KB)Acknowledgments
The authors thank the Alabama Paleontology Society for their hospitality and support throughout the duration of this project. We thank Dr. Ron Buta and Dr. Prescott Atkinson for their continued assistance with and the use of photographs. We thank the McWane Science Centre (Jun Embersol), the Anniston Museum of Natural History (Dan Spaulding), and The Alabama Museum of Natural History (Dana Ehret) for access to their collections, cooperation and curatorial information on curated UCM specimens. OK and MS thank the continual support of Christopher Taylor and Marion Homer. MS thanks Nancy Jacque and David Wagner for their moral and financial support during the writing of this manuscript. We thank David Rudkin and Graham Young for providing information on Early Paleozoic xiphosurids. Michael Young at Dalhousie University is thanked for his support throughout the project and allowing an early version of this manuscript to be used for academic credit. Steven Hinds, Dr. R. Andrew MacRae and Dr. John Calder are thanked for insightful discussions. We would also like to thank the anonymous reviewers for their thorough reviews. We thank Ron Buta and David Kopaska-Merkel for permission to reproduce figures from their popular book “Footprints in Stone, Fossil Traces of Coal-Age Tetrapods” as well as Jim Lacefield for permissions to reproduce his paleogeographic artwork within that book.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Abel, O. 1935. Vorzeitliche Lebensspuren (644 pp). Jena, Germany: Gustav Fischer.
- Actenolaza, F. G. 1978. Trazas fosiles de la formacion patquia en el bordo atravesado, Sierra de Famatina, La Rioja. Acta Geologica Lilloana 15:19–29.
- Aldrich, T. H. Sr., and Jones, W. B. 1930. Footprints from the coal measures of Alabama (64 p). Museum Paper 9, Geological Survey of Alabama, Tuscaloosa, Alabama.
- Allen, A. 2005. Discovery of the Union Chapel Mine site. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 15–17. Birmingham, AL: Alabama Paleontological Society Monograph.
- Anderson, L., J. A. Dunlop, C. A. Horrocks, H. M. Winkelmann, and R. M. C. Eagar. 1997. Exceptionally preserved fossils from Bickershaw, Lancashire UK (Upper Carboniferous, Westphalian A (Langsettian)). Geological Journal 32(3):197–210.
- Archer, A. W., J. H. Calder, M. R. Gibling, R. D. Naylor, D. R. Reid, and W. G. Wightman. 1995. Invertebrate trace fossils and agglutinated foraminifera as indicators of marine influence within the classic Carboniferous section at Joggins, Nova Scotia, Canada. Canadian Journal of Earth Sciences 32(12):2027–2039.
- Atkinson, T. P. 2005. Arthropod body fossils from the Union Chapel Mine. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 169–176. Birmingham, AL: Alabama Paleontological Society Monograph.
- Babcock, L. E., D. F. Merriam, and R. R. West. 2000. Paleolimulus, an early limuline (Xiphosurida), from Pennsylvanian-Permian Lagerstaetten of Kansas and taphonomic comparison with modern limulus. Lethaia 33:129–141.
- Braddy, S. J. 2001. Eurypterid palaeoecology: palaeobiological, ichnological and comparative evidence for a ‘mass-moult-mate’ hypothesis. Palaeogeography, Palaeoecology, Palaeoclimatology 172(1–2):115–132.
- Briggs, D. E., and Rolfe, W. I. 1983. A giant arthropod trackway from the Lower Mississippian of Pennsylvania. Journal of Paleontology 57(2):377–390.
- Buatois, L. A., M. G. Mangano, C. G. Maples, and W. P. Lanier. 1998. Ichnology of an Upper Carboniferous fluvio-estuarine paleovalley: the Tonganoxie Sandstone, Buildex Quarry, Eastern Kansas, USA. Journal of Paleontology 72(01):152–180.
- Buatois, L. A., M. K. Gingras, J. MacEachern., M. G. Mangano, J. P. Zonneveld, S. G. Pemberton, R. G. Netto, and A. Martin. 2005. Colonisation of brackish-water systems through time: evidence from the trace-fossil record. Palaios 20(4):321–347.
- Buhler, P. B., and Grey, M. 2017. Xiphosuran digging traces at the Late Carboniferous Joggins Fossil Cliffs UNESCO World Heritage Site, Nova Scotia, Canada. Ichnos 24(3):179–190.
- Buta, R. J., and Kopaska-Merkel, D. C. 2016. Footprints in Stone, Fossil Traces of Coal-Age Tetrapods. Tuscaloosa, Alabama: University of Alabama Press.
- Buta, R. J., D. C. Kopaska-Merkel, A. K. Rindsberg, and A. J. Martin. 2005. Atlas of Union Chapel Mine invertebrate trackways and other traces. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 277–337. Birmingham, AL: Alabama Paleontological Society Monograph.
- Buta, R. J., J. C. Pashin, N. J. Minter, and D. C. Kopaska-Merkel. 2013. Ichnology and stratigraphy of the Crescent Valley Mine: evidence for a Carboniferous megatracksite in Walker County, Alabama. In The Carboniferous-Permian transition, ed. S. G. Lucas, vol. 60, 42–56. Albuquerque, NM: New Mexico Museum of Natural History & Science Bulletin.
- Butts, C. 1926. The Paleozoic rocks. In Geology of Alabama, ed. G. I. Adams, C. Butts, L. W. Stephenson, and C. W. Cooke, vol. 14, 40–230. Tuscaloosa, AL: Alabama Geological Survey Special Reports.
- Calder, J. H. 1998. The Carboniferous evolution of Nova Scotia. In Lyell: The past is the key to the present, ed. D. J. Blundell and A. C. Scott, vol. 143, 261–302. London, UK: Geological Society of London Special Publication.
- Caputo, M. V., and Crowell, J. C. 1985. Migration of glacial centers across Gondwana during Paleozoic Era. Geological Society of America Bulletin 96(8):1020–1036.
- Caster, K. E. 1938. A restudy of the tracks of Paramphibius. Journal of Paleontology 12:3–60.
- Caster, K. E. 1944. Limuloid trails from the Upper Triassic (Chinle) of the Petrified Forest National Monument, Arizona. American Journal of Science 242(2):74–84.
- Chatterji, A., and Abidi, S. A. H. 1993. The Indian horseshoe crab: a living fossil. Journal of Indian Ocean Studies 1(1):43–48.
- Chrisholm, J. I. 1983. Xiphosurid trace Kouphichnium aff. variabilis (Linck) from the Namurian Upper Haslingden Flags of Whitworth, Lancashire, rep. Institute of Geological Sciences 83(10):37–44.
- Copeland, M. J. 1957. The arthropod fauna of the Upper Carboniferous rocks of the Maritime Provinces. Geological Survey of Canada 286:W110.
- Cotton, W. D., A. P. Hunt, and J. E. Cotton. 1995. Paleozoic tracksites in Eastern North America. Bulletin of New Mexico Museum of Natural History and Science 6:189–211.
- Crimes, T. P. 1975. Trilobite traces from the Lower Tremadoc of Tortworth. Geological Magazine 112(01):33.
- Davis, R. B., N. J. Minter, and S. J. Braddy. 2007. The neoichnology of terrestrial arthropods. Palaeogeography, Palaeoclimatology, Palaeoecology 255(3–4):284–307.
- Demko, T. M., and Gastaldo, R. A. 1996. Eustatic and autocyclic influences on deposition of the Lower Pennsylvanian Mary Lee coal zone, Warrior Basin, Alabama. International Journal of Coal Geology 31(1-4):3–19.
- Dilcher, D. L., and T. A. Lott. 2005. Atlas of Union Chapel Mine Fossil Plants. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 339–365. Birmingham, AL: Alabama Paleontological Society Monograph.
- Dilcher, D. L. T. A. Lott, and B. J. Axsmith. 2005. Fossil plants from the Union Chapel Mine, Alabama. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 153–168. Birmingham, AL: Alabama Paleontological Society Monograph.
- DiMichele, W. A., and T. L. Phillips. 1995. The response of hierarchially-structured ecosystems to long-term climatic change: a case study using tropical peat swamps of Pennsylvanian age. In Effect of past global change on life, ed. S. M. Stanley, 134–155. Washington, DC: National Research Council Studies on Geophysics.
- Eble, C. F., and Gillespie, W. H. 1989. Palynology of selected coal beds from the central and southern Appalachian basin: correlation and stratigraphic implications, Guidebook T352, 61–66. Paper presented at the 28th International Geological Congress, Washington, DC.
- Eble, C., C. F. Eble, W. H. Gillespie, and T. W. Henry. 1991. Palynology, paleobotany, and invertebrate paleontology of Pennsylvanian coal beds and associated strata in the Warrior and Cahaba coal fields. In Mississippian-Pennsylvanian tectonic history of the Cahaba synclinorium, ed. W. A. Thomas and W. E. Osborne, 28th Annual Field Trip, 119–132. Tuscaloosa, AL: Alabama Geological Society.
- Erickson, B. R. 2005. Crocodile and arthropod tracks from the Late Paleocene Wannagan Creek Fauna of North Dakota, USA. Ichnos 12(4):303–308.
- Falcon-Lang, H. J. 2003. Response to Late Carboniferous tropical vegetation to transgressive-regressive rhythms at Joggins, Nova Scotia. Journal of the Geological Society of London 160(4):643–648.
- Falcon-Lang, H. J. 2006. Vegetation ecology of Early Pennsylvanian alluvial fan and piedmont environments in Southern New Brunswick, Canada. Palaeogeography, Palaeoclimatology, Palaeoecology 233(1–2):34–50.
- Falcon-Lang, H. J., and Bashforth, A. R. 2004. Pennsylvanian uplands were forested by giant cordaitalean trees. Geology 32(5):417–420.
- Falcon-Lang, H. J., M. J. Benton, S. J. Braddy, and S. J. Davies. 2006. The Pennsylvanian tropical biome reconstructed from the Joggins Formation of Nova Scotia, Canada. Journal of the Geological Society, London 163(3):561–576.
- Ferm, J. C., R. Ehrlich, and T. L. Neathery. 1967. A field guide to Carboniferous detrital rocks in northern Alabama. Coal division field trip guidebook, W101. New Orleans, LA: Geological Society of America.
- Fillion, D., and Pickerill, R. K. 1990. Ichnology of the Lower Ordovician Bell Island and Wabana Groups of Eastern Newfoundland. Palaeontographica Canadiana 7:1–119.
- Fillmore, D. L., S. G. Lucas, and E. L. Simpson. 2012. Ichnology of the Mauch Chunk Formation, eastern Pennsylvania. Albuquerque: New Mexico Museum of Natural History and Science. Bulletin 54:132.
- Fischer, W. A. 1978. The habitat of the early vertebrates: trace and body fossil evidence from the Harding Formation (Middle Ordovician), Colorado. The Mountain Geologist 15:1–26.
- Frakes, L. A., J. E. Francis, and J. I. Syktus. 1992. Climate modes of the Phanerozoic, W274. Cambridge, UK: Cambridge Press.
- Gastaldo, R. A. 1990. Early Pennsylvanian swamp forests in the Mary Lee Coal Zone, Warrior Basin, Alabama. In Carboniferous coastal environments and paleocommunities of the Mary Lee Coal Zone, Marion and Walker Counties, Alabama, ed. R. A. Gastaldo, 1–54. Tuscaloosa, AL: Geological Survey of Alabama.
- Gibb, S., B. D. Chatterton, and S. G. Pemberton. 2011. Prod traces (Selenichnites) from the Middle Cambrian of Morocco, with hypotheses on the ethology of the tracemaker(s). Ichnos 18(3):156–165.
- Goldring, R., and Seilacher, A. 1971. Limulid undertracks and their sedimentological implications. Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen 137:422–442.
- Grey, M., P. K. Pufahl, and A. A. Aziz. 2011. Using multiple environmental proxies to determine degree of marine influence and paleogeographical position of the Joggins Fossil Cliffs, UNESCO World Heritage Site. Palaios 26(5): 256–263.
- Häntzschel, W. 1975. Treatise on invertebrate paleontology: Part W, Miscellanea, Supplement 1, W269. Lawrence, KS: Geological Society of America and University of Kansas Press.
- Hardy, P. G. 1970. New xiphosurid trails from the upper carboniferous of Northern England. Palaeontology 13:188–190.
- Haubold, H. A. Allen, T. P. Atkinson, R. J. Buta, J. A. Lacefield, S. C. Minkin, and B. A. Relihan. 2005. Interpretation of the tetrapod footprints from the Early Pennsylvanian of Alabama. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 75–111. Birmingham, AL: Alabama Paleontological Society Monograph.
- Haug, C., P. V. Roy, A. Leipner, P. Funch, D. M. Rudkin, L. Schöllmann, and J. T. Haug. 2012. A holomorph approach to xiphosuran evolution—a case study on the ontogeny of Euproops. Development Genes and Evolution 222(5):253–268.
- Hobday, D. K. 1974. Beach and barrier island facies in the upper Carboniferous of Northern Alabama. Geological Society of America Special Paper 148:209–224.
- Hooks, G. E. III. 2005. The importance of the amateur paleontologist in buta. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J., Rindsberg, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 187–188. Birmingham, AL: Alabama Paleontological Society Monograph.
- Horne, J. C. 1979. The effects of Carboniferous shoreline geometry on paleocurrent distribution. In Carboniferous depositional environments in the Appalachian region, ed. J. C. Ferm and J. C. Horne, 509–516. Columbia, SC: Carolina Coal Group, University of South Carolina.
- Hunt, A. P., and S. G. Lucas. 2005. Tetrapod ichnofacies and their utility in the Paleozoic. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 113–119. Birmingham, AL: Alabama Paleontological Society Monograph.
- Hunt, A. P., S. G. Lucas, and N. D. Pyenson. 2005. The significance of the Union Chapel Mine Site: A Lower Pennsylvanian (Westphalian A) Ichnological Konzentratlagerstätte, Alabama, USA. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 3–14. Birmingham, AL: Alabama Paleontological Society Monograph.
- Hunt, A. P., S. G. Lucas, and M. G. Lockley. 2010. Large pelycosaur footprints from the Lower Pennsylvanian of Alabama, USA. Ichnos 11(1–2):39–45.
- King, O. A., R. F. Miller, and M. R. Stimson. 2017. Ichnology of the Devonian (Emsian) Campbellton Formation, New Brunswick, Canada. Atlantic Geology 53:1–8.
- Kjellesvig-Waering, E. N. 1964. A synopsis of the family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida). Journal of Paleontology 38:331–361.
- Kopaska-Merkel, D. C., and Buta, R. J. 2013. Field trip guidebook to the Steven C. Minkin Paleozoic Footprint site, Walker County, Alabama. In The Carboniferous-Permian Transition, 60, 178–198.
- Lacefield, J. A., and B. A. Relihan. 2005. The significance of the Union Chapel Mine site to Alabama paleontology. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 201–204. Birmingham, AL: Alabama Paleontological Society Monograph.
- Lerner, A. J., and Lucas, S. G. 2015. A Selenichnites ichnoassociation from early Permian tidal flats of the Prehistoric Trackways National Monument of South-central New Mexico. New Mexico Museum of Natural History and Science Bulletin 65:141–152.
- Lerner, A. J., S. G. Lucas, and C. F. Mansky. 2016. The earliest paleolimulid and its attributed ichnofossils from the Lower Mississippian (Tournaisian) Horton Bluff Formation of Blue Beach, Nova Scotia, Canada. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 280(2):193–214.
- Linck, O. 1949. Lebens-Spuren aus dem Schilfsandstein (Mittl. Keuper km 2) N Wurttembergs und ihre Bedeutung fur die Bildungsgeschichte der Stufe. Jahreshefte Des Vereins Fur Vaterlandische Naturkunde in Wurttemberg 97–101:1–100.
- Lucas, S. G., and A. J. Lerner. 2005. Lower Pennsylvanian invertebrate ichnofossils from the Union Chapel Mine, Alabama: a preliminary assessment. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 147–152. Birmingham, AL: Alabama Paleontological Society Monograph.
- Lucas, S. G., and M. R. Stimson. 2013. Re-evaluation of Arenicolites and Treptichnus ichnospecies from the Pennsylvanian Pottsville Formation, Union Chapel Mine, Alabama. In The Carboniferous - Permian transition, ed. S. G. Lucas, vol. 60, 240–243. Albuquerque, NM: New Mexico Museum of Natural History, Bulletin.
- Lucas, S. G., N. J. Minter, J. A. Spielmann, A. P. Hunt, S. J. Braddy, and K. E. Zeigler. 2005. Early Permian ichnofossil assemblage from the Fra Cristobal Mountains, southern New Mexico. New Mexico Museum of Natural History and Science Bulletin 31: 140–150.
- Malz, H. 1964. Kouphichnium walchi, die Geschichte einer Fährte und ihres Tieres. Natur und Museum 94:81–97.
- Mángano, M. G., and L. A. Buatois. 2004. Reconstructing early Phanerozoic intertidal ecosystems: Ichnology of the Cambrian Campanario Formation in northwest Argentina. In Trace fossils in evolutionary palaeoecology: Fossils and strata, ed. B. Webby, M. G. Mángano, and L. A. Buatois, vol. 51, 17–38. Abingdon, UK: Taylors & Francis.
- Mansky, C. F., and S. G. Lucas. 2013. Romer's Gap revisited: continental assemblages and ichno-assemblages from the basal Carboniferous of Blue Beach, Nova Scotia, Canada. In The Carboniferous-Permian transition. Albuquerque: Bulletin 60, ed. S. G. Lucas, W. A. DiMichele, J. E. Barrick, J. W. Schneider, and J. A. Speilmann, 244–273. Albuquerque, NM: New Mexico Museum of Natural History and Science.
- Martin, A. J., and N. D. Pyenson. 2005. Behavioral significance of vertebrate trace fossils from the Union Chapel site. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, no. 1, 59–74. Birmingham, AL: Alabama Paleontological Society Monograph.
- Marty, D., A. Strasser, and C. A. Meyer. 2009. Formation and taphonomy of human footprints in microbial mats of present-day tidal-flat environments: Implications for the study of fossil footprints. Ichnos 16(1–2):127–142.
- Meek, F., and Worthen, A. 1865. Notice of some new types of organic remains, from the coal measures of Illinois. Proceedings of the Academy of Natural Sciences of Philadelphia 17(1):41–48.
- Milàn, J., and Bromley, R. G. 2007. The impact of sediment consistency on track and undertrack morphology: Experiments with emu tracks in layered cement. Ichnos 15(1):19–27.
- Miller, M. F. 1982. Limulicubichnus: a new ichnogenus of limulid resting traces. Journal of Paleontology 56:429–433.
- Minkin, S. C. 2005. Paleoenvironment of the Cincosaurus beds, Walker County, Alabama. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 31–38. Birmingham, AL: Alabama Paleontological Society Monograph.
- Minter, N. J., and Braddy, S. J. 2009. Ichnology of an Early Permian intertidal flat: The Robledo Mountains Formation of Southern New Mexico, USA. Special Papers in Palaeontology 82:1–107.
- Moore, R. 1955. Treatise on invertebrate paleontology: Part P, Arthropoda, W181. Lawrence, KS: Geological Society of America.
- Moore, R., S. C. McKenzie, and B. S. Lieberm. 2007. A Carboniferous synxiphosurine (Xiphosura) from the Bear Gulch Limestone, Montana, USA. Palaeontology 50(4):1013–1019.
- Nitecki, M. H. ed. 2013. Mazon Creek fossils. New York, NY: Elsevier.
- Nopsca, F. B. 1923. Die Familien der Reptilien. Fortschritte in Der Geologie Und Paläontologie 2:1–210.
- Oppel, A. 1862. Uber Fahrten im lithographischen Schiefer (Ichnites lithographicus). Museum Des Koniglich Bayerischen Staates, Palaontologische Mittheilungen 1:121–125.
- Pashin, J. C. 1994a. Cycles and stacking patterns in Carboniferous rocks of the Black Warrior foreland basin. Gulf Coast Association of Geological Societies, Transactions 44:555–563.
- Pashin, J. C. 1994b. Flexurally influenced eustatic cycles in the Pottsville Formation (Lower Pennsylvanian), Black Warrior basin, Alabama. In Tectonic and eustatic controls on sedimentary cycles, ed. J. M. Dennison and F. R. Ettensohn, vol. 4: 89–105. Chapel Hill, NC: Society for Sedimentology, Concepts in Sedimentology and Paleontology.
- Pashin, J. C. 2005. Pottsville stratigraphy and the Union Chapel Lagerstätte. In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1: 39–58. Birmingham, Alabama: Alabama Paleontological Society Monograph.
- Prescott, Z. M., M. R. Stimson, L. T. Dafoe, M. R. Gibling, R. A. Macrae, J. H. Calder, and B. L. Hebert. 2014. Microbial mats and ichnofauna of a fluvial-tidal channel in the Lower Pennsylvanian Joggins Formation, Canada. PALAIOS 29(12):624–645.
- Rindsberg, A. K. 2005. Gas-escape structures and their paleoenvironmental significance at the Steven C. Minkin Paleozoic Footprint Site (Early Pennsylvanian, Alabama). In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 177–183. Birmingham, AL: Alabama Paleontological Society Monograph.
- Rindsberg, A. K., A. Uchman, and D. C. Kopaska-Merkel. 2004. Treptichnus made by insect larvae in the Pennsylvanian of Alabama (USA): First International Congress on Ichnology, vol. 70. Trelew, Patagonia Argentina: Museo Paleontológico Egidio Feruglio, April 19–23.
- Rindsberg, A. K., and Kopaska-Merkel, D. C. 2003. Treptichnus and related trace fossils from the Union Chapel Mine (Langsettian, Alabama, USA). In Work-shop on Permo-Carboniferous Ichnology, Program and Abstracts, ed. A. J. Martin, 27–28. Tuscaloosa: University of Alabama.
- Rindsberg, A. K., and D. C. Kopaska-Merkel. 2005a. Treptichnus and Arenicolites from the Steven C. Minkin Paleozoic Footprint Site (Langsettian, Alabama, USA). In Pennsylvanian footprints in the Black Warrior Basin of Alabama, ed. R. J. Buta, A. K. Rindsberg, and D. C. Kopaska-Merkel, vol. 1, 121–141. Birmingham, AL: Alabama Paleontological Society Monograph.
- Rindsberg, A. K., and Kopaska-Merkel, D. C., eds., 2005b. Pennsylvanian footprints in the Black Warrior Basin of Alabama, vol. 1, 59–73. Birmingham, AL: Alabama Paleontological Society Monograph.
- Romano, M., and Melendez, B. 1985. A arthropod (merostome) ichnocoenosis from the Carboniferous of northwest Spain. Compte Rendu Neuvieme Congres International de Stratigraphie et de Geologie du Carbonifere 5: 317–325.
- Romano, M., and Whyte, M. A. 1987. A limulid trace fossil from the Scarborough Formation (Jurassic) of Yorkshire; its occurrence, taxonomy and interpretation. Proceedings of the Yorkshire Geological Society 46(2): 85–95.
- Romano, M., and Whyte, M. A. 2015. A review of the trace fossil Selenichnites. Proceedings of the Yorkshire Geological Society 60(4): 275–288.
- Rudkin, D. M., G. A. Young, and G. S. Nowlan. 2008. The oldest horseshoe crab: a new xiphosurid from Late Ordovician Konservat-Lagerstätten deposits, Manitoba, Canada. Palaeontology 51(1): 1–9.
- Scotese, C. R. 1997. Continental Drift, 7th ed., 79. PALEOMAP Project, Arlington, Texas.
- Seilacher, A. 2008. Biomats, biofilms, and bioglue as preservational agents for arthropod trackways. Palaeogeography, Palaeoclimatology, Palaeoecology 270(3–4):252–257.
- Shabica, C. W., and A. Hay, ed. 1997. Richardson's guide to the fossil fauna of Mazon Creek, 308. Chicago, IL: Northeastern Illinois University.
- Shuster, C. N., and Botton, M. L. 1985. A contribution to the population biology of horseshoe crabs, Limulus polyphemus (L.), in Delaware Bay. Estuaries 8(4):363–372.
- Shuster, C. N., R. B. Barlow, and H. J. Brockmann. 2003. The American horseshoe crab, 472. Cambridge, UK: Harvard University Press.
- Stimson, M. R., and MacRae, R. A. 2010. Evidence of fossil horseshoe crabs from Joggins, Nova Scotia: Paleoichnology and paleonvironmental implications. Atlantic Geology 46(1):69.
- Tetlie, O. E. 2007. Distribution and dispersal history of Eurypterida (Chelicerata). Palaeogeography, Palaeoclimatology, Palaeoecology 252(3–4):557–574.
- Vrazo, M. B., and Braddy, S. J. 2011. Testing the ‘mass-moult-mate’ hypothesis of eurypterid palaeoecology. Palaeogeography, Palaeoclimatology, Palaeoecology 311(1–2):63–73.
- Vrazo, M. B., and Ciurca, S. J. Jr. 2017. New trace fossil evidence for eurypterid swimming behaviour. Palaeontology 61:1–18.
- Willard, B. 1935. Chemung tracks and trails from Pennnsylvania. Journal of Paleontology 9:43–56.
- Wood, J. M. 1963. The Stanley Cemetery Flora (Early Pennsylvanian) of Greene County, Indiana. Indiana Geological Survey Bulletin 29:1–73.