Abstract
New Zealand's Jurassic plant fossils are known from non-marine settings in two terranes: the Murihiku forearc basin and from a small area on the Rakaia Terrane, an accretionary prism. These terranes lay in relatively high latitudes along the Gondwana margin but their precise location, latitude and position with respect to each other is unclear. The flora was dominated by conifers, ferns, bennettitaleans, pentoxylaleans and locally equisetaleans, but it is relatively depauperate, perhaps reflecting a high latitude position. Most climate indicators suggest a warm temperate climate with rainfall that was not especially high and that might have been seasonal at times. Revision of the New Zealand Jurassic flora is overdue, and while new genera may be discovered, species numbers may well decrease.
Introduction
New Zealand may be the key source of information for the vegetation of high southern latitudes during the Jurassic, despite some uncertainty about its palaeogeographic position. Research began with the description of fossil wood by Unger (Citation1864), then came the now-classic work by Kidston & Gwynne-Vaughan (Citation1907-1910) on the structure of tree fern stems, and a taxonomic apogee was reached with the work of Arber (Citation1917) and Edwards (Citation1934). More than a century from the earliest study, Mildenhall (Citation1970) evaluated the taxonomic legitimacy of all New Zealand plant macrofossils listing many Jurassic species as invalid. Recent work on New Zealand's Jurassic floras has included an unpublished MSc thesis (Broekhuizen Citation1984), a field-trip guide (Grant-Mackie Citation1995), a review by Grant-Mackie et al. (Citation2000), the work of Pole (Citation1999, Citation2001) and Thorn (Citation2001, Citation2005) on fossil forests, including palynology and sedimentology, and a concerted effort to develop a palynological zonation (de Jersey & Raine Citation1990, Citation2002). Raine et al. (Citation2008) have also provided an atlas of palynomorphs for the New Zealand region encompassing Jurassic forms.
The purpose of this paper is to review the taxa currently known from New Zealand and to summarise climatic indications and recent advances in understanding palaeogeography.
Palaeogeography
Any discussion on the Jurassic vegetation of New Zealand must address the issue of where New Zealand was located. Or more specifically, where were the components of New Zealand? New Zealand is known to be a collage of tectonic terranes of diverse origins (Bishop et al. Citation1985; Mortimer & Campbell Citation1996), although Kamp (Citation2001) cautioned that the time has come to consider their potential inter-relationships more than their differences. By the mid-Cretaceous, the components had essentially amalgamated and New Zealand lay in high, polar latitudes, but in the Jurassic at least three units have been identified and their relative positions at that time are still not clear.
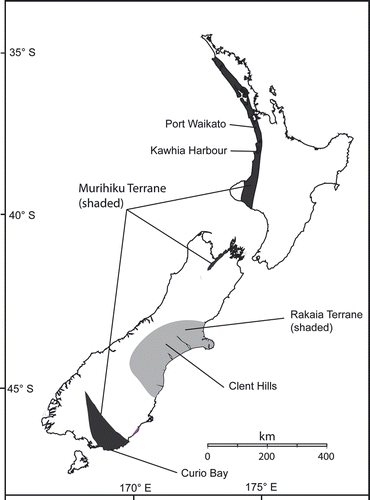
The Murihiku Terrane (MT) was a forearc basin that was active from the Triassic into the Cretaceous (Ballance & Campbell Citation1993; Landis et al. Citation1999 for a more recently recognised occurrence of Jurassic MT), and is the source of most of New Zealand's Jurassic plant fossils (Fig. ). The youngest Jurassic strata in New Zealand constitute the Huriwai Formation of Port Waikato, which is a part of the MT. It was initially thought to be Neocomian (Early Cretaceous) by Arber (Citation1917), but is now regarded as Tithonian (Edwards Citation1934; McQueen Citation1955; Norris Citation1968; Norris & Waterhouse Citation1970; Stevens & Speden Citation1978). In the broadest sense, the sediments of the MT accumulated somewhere along or off the margin of south-eastern Pangaea/Gondwana. The exact position has potential ecological implications, for instance, in determining whether the terrane and associated flora lay in a polar position or not. Pole (Citation1999) regarded the Middle Jurassic (Bathonian–Callovian, late Temaikan local stage; based on palynology, J.L. McKellar, J.I. Raine, and N. de Jersey, pers. comm. 1997; see also Speden (Citation1958), for marine faunal control of dating similar-aged leaf beds at Owaka Creek) Curio Bay fossil forests of the MT, southernmost New Zealand, as growing near to the southern polar circle, but had insufficient data to decide on which side. Thorn (Citation2001) claimed the Kawhia Harbour forests, of similar age to those at Curio Bay, grew at approximately 75–78°S, “well within the polar circle” and cited Stevens (Citation1980, Citation1985) in support of a warmer climate at the time, a poleward extension of the Tethyan faunal realm, and an absence of a distinct polar realm. However, the point that Stevens tried to make was that if New Zealand lay at high latitudes at the time, it would most likely have had a distinct cool-temperate “Palaeoaustral” faunal realm – to mirror the Boreal fauna that occurred in high northern latitudes. Stevens (Citation1980) concluded that New Zealand lay in mid-latitudes during the Middle Jurassic.
Fig. 1 Locality map of Jurassic sites in New Zealand, showing terranes (modified from Bishop et al. Citation1985) and localities mentioned in the text.
Adams et al. (Citation2007) examined the ages of detrital zircons in New Zealand's terranes; most MT zircons were contemporary and there was little in the way of an older or “continental” source. For this reason, Adams et al. (Citation2007) argued that the MT was an “isolated volcanic environment” and suggested that it existed in its present New Zealand position; they did acknowledge a source of older zircons in the Middle Jurassic, which might indicate a continental source. Their reconstruction of the Gondwana coastline shows the MT at around 70°S in the Late Triassic and lying at around 85°S by the Late Jurassic.
Outboard of the MT was a series of accretionary prisms including the Rakaia Terrane. This complex includes abundant Jurassic rocks, but most have either undergone significant metamorphism to schist (e.g. Mortimer Citation1993), or are marine. However, a small fluvial, plant fossil-containing, Late Jurassic (Tithonian) deposit in the Clent Hills (Canterbury region) accumulated on this terrane as it was being imbricated against the New Zealand margin and uplifted (Graham & Mortimer Citation1992; Kamp Citation2001).
Attempts to show the palaeolatitudinal positions of all New Zealand terranes in the Jurassic have been made by Grant-Mackie et al. (Citation2000) and Adams et al. (Citation2007). However, from a palaeoecological point of view (for instance, their position relative to the palaeopolar circle), it is important to realise that these are estimates.
The macrofossil taxa
New Zealand's Jurassic macrofossil taxa are summarised below. For convenience, they are grouped into likely or generally accepted higher taxa, but in the absence of cuticular and fertile material for most New Zealand specimens, these are provisional.
Clubmosses
Lycopodiaceae
Lycopodites arberi, based on an impression of a small, herbaceous shoot, is the only lycopod described from the New Zealand Jurassic (Edwards Citation1934).
Ferns
Osmundaceae
Arber (Citation1917, 31) regarded Cladophlebis australis as the “most abundant of all species, without exception” in the New Zealand Jurassic. Since then, Frenguelli (Citation1947) placed the New Zealand Jurassic forms into Cladophlebis patagonica and Retallack (Citation1983) argued that C. australis should be restricted to Triassic forms that have doubly forking veins. At least four other species of Cladophlebis occur in the New Zealand Jurassic (Arber Citation1917; Edwards Citation1934): Cladophlebis sp. cf. C. albertsi, C. antarctica, C. denticulate and C. reversa. Cladophlebis is generally accepted as the foliage associated with the fossil axes that were first described by Kidston & Gwynne-Vaughan (Citation1907-10) as Osmundites (see also Marshall Citation1926). The New Zealand species have since been placed into Osmundacaulis (Miller Citation1967), and Millerocaulis and Ashicaulis (Tidwell Citation1994). The size of these axes indicates that the plant was a small tree fern.
Dicksoniaceae
The foliage Coniopteris hymenophylloides recorded by Arber (Citation1917) was probably a ground fern. Edwards (Citation1934) also recorded C. lobata.
Dipteridaceae
Arber (Citation1917) listed two species of Dictyophyllum from New Zealand (D. acutilobum and D. obtusilobum), although Edwards (Citation1934) regarded them as specifically indeterminate and possibly the same species. Oliver (Citation1950) recorded a third species, D. rugosum. Following Herbst's (Citation1975) work on the family, Rees (Citation1993) reviewed the dipterid ferns of Antarctica and New Zealand and described a new species from the Clent Hills as Goeppertella cf. woodii, the first record of this genus from New Zealand. He pointed out that it would be difficult to differentiate Goeppertella from Dictyophyllum based on fragmentary material (i.e. if it was not clearly bipinnate). This suggests that Arber's records may need re-evaluation.
Schizaeaceae
Arber (Citation1917) recognised six species of Sphenopteris, including the cosmopolitan S. (Ruffordia) goepperti, S. currani previously known from Australia (Tenison-Woods Citation1883), three endemic and one undescribed species.
Equisetaceae
Two endemic species of Equisetum are validly described (as Equisetites) from the MT: E. hollowayei (Edwards Citation1934) and E. nicoli (Arber Citation1917). They are represented by small axes and leaf whorls and probably represent herbaceous plants.
Gymnosperms
Pteridosperms
Microphyllopteris was instituted by Arber (Citation1917) on New Zealand material for ferns that resemble Gleichenia and belong to the Gleicheniaceae, but Nagalingum & Cantrill (Citation2006) have proposed that it should be restricted to ferns that “cannot be ascribed conclusively to Gleicheniaceae”. Arber (Citation1917) described one species M. pectinata, and figured another. McLoughlin et al. (Citation2002) transferred M. pectinata to a new genus, Rintoulia, which they suggested was a seed fern based on its similarity to leaves with robust cuticle from the Victorian Cretaceous.
At least four species of Thinnfeldia are present according to Arber (Citation1917), all of them being widespread species. Barbacka (Citation1994) transferred T. indica (Feistmantel) (from India) and T. nordenskioeldii Nathorst (from Europe and possibly China) to a new genus, Komlopteris, and it may be that the New Zealand Thinnfeldia also belong to this new genus.
Bennettitales
The Bennettitales are probably represented by three species in three form genera. These include the widespread (though uncommon within assemblages) Ptilophyllum acutifolium and the endemic Pterophyllum matauriensis (Arber Citation1917), and Edwards (Citation1934) added Otozamites.
Pentoxylales
The most common Taeniopteris species in New Zealand and Australia, T. daintreei, is generally accepted as the foliage of the pentoxylaleans (Drinnan & Chambers Citation1985), and this is supported by the presence in New Zealand of pentoxylalean cones, Carnoconites (Harris Citation1962) and permineralised stems known as Pentoxylon. Pole (Citation1999) illustrated in situ tree stumps in the fossil forest at Curio Bay, which show the distinctive polyxylic architecture of Pentoxylales. The group (or something closely related) is also clearly present in a private collection; R. Jackson, Christchurch, collected from the Slope Point area, south of Curio Bay; M. Pole, pers. obs. 2000. The widespread T. arctica, T. crassinervis, T. vittata and an endemic species T. thomsoniana are also present in New Zealand (Arber Citation1917), but it is not clear whether these were also pentoxylaleans since similar leaves were also produced by some cycads, bennettites and ferns. Taeniopteris is a very simple leaf morphogenus, leaving little to distinguish the species other than a range of dimensions, the approach used by Blaschke & Grant-Mackie (Citation1976) for delimiting Taeniopteris species at Port Waikato and the Clent Hills. A study of large collections would give a clearer idea of the species limits and could well reduce the number of taxa recognised from New Zealand. Edwards (Citation1934) adopted a broad species view and combined T. arctica, T. daintreei and T. vittata with T. spatulata (he regarded T. daintreei as a synonymy of T. spatulata).
Cycadales
The cycadales are possibly represented by the widespread Nilssonia compta and the endemic N. elegans as identified by Arber (Citation1917). However, in light of recent disagreement among authors on what constitutes Nilssonia or Taeniopteris (e.g. Gee Citation1989 versus Rees & Cleal Citation2004), these species may yet be placed in Taeniopteris. Arber (Citation1917) also recorded a Cycadites sp. from Curio Bay.
Conifers
The most common type of conifer foliage by far is a widespread form that has flattened and spreading single-veined leaves, and is commonly referred to Elatocladus conferta (Arber Citation1917). This happens to be the only Jurassic plant in New Zealand for which the cuticular detail is known from specimens in the Clent Hills locality (Townrow Citation1967); this information, together with associated reproductive organ data, allowed him to describe a new genus and species, Mataia podocarpoides (Townrow Citation1967). However, extreme caution should be exercised before automatically placing specimens described as E. conferta into M. podocarpoides because the former is a rather generalised morphospecies that may cover several species. An Elatocladus with longer leaves, E. plana, also occurs in the MT, in some cases in association with E. conferta (Bartrum Citation1921; pers. obs, Fig. .).
Fig. 2 Bedding surface of siltstone (False Island Formation, probably Bathonian–Callovian), with Elatocladus sp. and thin-leaved Taeniopteris from the coast north of Curio Bay, New Zealand. Rock hammer for scale.

Other conifers include forms with imbricate scale leaves identified as Brachyphyllum, Cryptomerites and Pagiophyllum (Arber Citation1917). The multiveined, endemic Podozamites gracilis is probably referable or related to the Araucariaceae, as are the similar Nageiopsis longifolia (Arber Citation1917) and Desmiophyllum (Edwards Citation1934). These latter taxa might be the foliage of the plants that produced Araucarites cutchensis and Araucarites grandis ovulate scales (Arber Citation1917).
Two form genera of conifer wood are known, Araucarioxylon or Dadoxylon and Mesembrioxylon (Crié Citation1889; Arber Citation1917; Edwards Citation1934), but the affiliation of the New Zealand woods needs to be updated given recent nomenclatural revisions (e.g. Bamford & Philippe Citation2001), and there is no doubt that more species are awaiting discovery and description. Certainly, palynological work indicates that the actual diversity of conifers in the region was much higher than that indicated by macrofossils (de Jersey & Raine Citation1990; Thorn Citation2001).
Palissyaceae
The distinctive but enigmatic cone Palissya bartrumi (Edwards Citation1934) is common in some localities in the MT. It was earlier assumed that these structures were produced by conifers, but a recent review that also documents Australian occurrences has suggested otherwise (Parris et al. Citation1995). On the basis of this work, Miller (Citation1999) ruled out a coniferous origin.
Palynology
Early palynological investigation of the Jurassic was carried out by Couper (Citation1953, Citation1960), Norris (Citation1968) and Norris & Waterhouse (Citation1970). Detailed work by de Jersey & Raine (Citation1990) documented a close relationship between the Early Jurassic palynoflora of New Zealand and that of Queensland, Australia. De Jersey & Grant-Mackie (Citation1989) and Grant-Mackie et al. (Citation2000) have also noted that the broader palynological sequence of the New Zealand Jurassic parallels that of Australia, New Caledonia, New Guinea, and perhaps more broadly, with Antarctica and Argentina. They noted that the New Zealand palynological record follows a similar sequence of a Cheirolepidiacean phase, followed by an Araucariacean phase, and finally by a Podocarpacean phase. Thorn (Citation2001) documented palynological assemblages from Kawhia Peninsula and Curio Bay.
The Forests
The Temaikan fossil forest at Curio Bay (actually one of several at that location and part of a broad sequence of plant fossil-rich sediments; Pole Citation2004) has long been known (Hector Citation1886; Park Citation1887; Arber Citation1917; Guy-Ohlson Citation1974; Raine & Pole Citation1988). The structure of the most completely exposed forest was documented by Pole (Citation1999) who regarded it as lying close to the palaeo-South Polar circle, but without sufficient data to determine on which side. The forest was probably of first generation, with a low canopy of about 10 m high, where most trees were about 70 years old, plus a few large emergents around 200 years old rising to about 30 m. There was a well-developed undergrowth of small gymnosperms (young conifers and Pentoxylon and osmundaceous “tree ferns”). There was nothing unique about the structure or density that might suggest that the forest grew in unusual polar conditions. The forest grew on a floodplain adjacent to a volcanic upland where intermittent, flashy and poorly confined floods overwhelmed at least 10 forests in about 40 m of section (Pole Citation2001).
In the North Island, Thorn (Citation2001) documented the Temaikan (Middle Jurassic MT) vegetation of Kawhia Harbour. She identified several plant communities that had grown in a braided river environment. Elatocladus was the only type of conifer foliage present. This taxon is generally assumed to belong to the Podocarpaceae, although interestingly, Thorn (Citation2001) did note that the local palynoflora was dominated by Araucariacites. Two types of coniferous wood were present: the Dadoxylon–Araucarioxylon type and Pentoxylon. Palynology suggested that several species of Araucariaceae or relatives were present, together with Podocarpaceae and possibly Taxodiaceae. Beyond this, only the most common Jurassic macrofossil genera in New Zealand were present: Taeniopteris and at least two species of Cladophlebis, although there were possible bryophytes, and some seeds and cones.
The Curio Bay and Kawhia forests both grew in relatively unstable environments and Ballance (Citation1988) envisaged a similar landscape for the Late Jurassic Huriwai braidplain. Lower energy, meandering fluvial systems did exist, for instance, along the Otara-Fortrose coastline in the southernmost MT (Pole Citation2004) but these await more complete documentation.
Climate
The Jurassic is recognised as a relatively warm period globally, with a low equatorial–polar temperature gradient. Although the evidence for any “notable” global climate change over its time span is inconsistent, variation certainly occurred (Parrish Citation1991; Hallam et al. Citation1993; Grant-Mackie et al. Citation2000).
Major advances have been made in recent years in techniques for quantitative estimates of the palaeoclimate of angiosperm-dominated macrofossil assemblages (e.g. Wolfe Citation1995). These techniques rely on the shapes and sizes of angiosperm leaves, which are very responsive to climate, or on the climatic requirement of nearest living relatives. Unfortunately, these techniques are not so applicable to the pre-angiosperm floras of the Jurassic. Non-angiosperm vegetation has a more conservative morphology with respect to climate, and at least until now, no one has devised a similar scheme for the Mesozoic or older. For this reason, most estimates of Jurassic climate have been qualitative. In broad terms, New Zealand lay in relatively high latitudes where growth rings in wood were prominent (Creber & Chaloner Citation1985).
Rees et al. (Citation2000) used multivariate analysis to illustrate a global floral gradient of genera (or form genera) for the Early Jurassic. They were able to show that microphyllous cycadophytes (in which they included bennettites) and conifers plotted together at one end of an ordination axis, while macrophyllous conifers and ginkgophytes plotted together at the other. They interpreted these two distinct floral associations in terms of hot and dry, and seasonally cool and/or dark (i.e. deciduous), respectively, and they went further to develop a “gradient score” for each Jurassic genus. This allowed them to “assign a value to any Jurassic plant locality, by averaging the scores of its constituent leaf genera” (Rees et al. Citation2000, 303), and thus place a locality on the ordination axis. For New Zealand, the relevant genera and their scores are: Equisetites, 76; Podozamites, 72; Coniopteris, 72; Cladophlebis, 70; Nilssonia, 55; Elatocladus, 52; Sphenopteris, 51; Taeniopteris, 50; Pterophyllum, 45; Pagiophyllum, 28; Ptilophyllum, 13; Brachyphyllum, 9 and Otozamites, 9. This gives an average of 46 and fits broadly with mid-latitudes (approximately 40–70°). The addition of the unpublished Hausmannia would raise the score slightly to 49 (meaning a move away from the pole).
But do microphyllous conifer leaves indicate a hot and dry climate? The conifer leaves of the New Zealand Jurassic are small and are of similar size to those living now in cool, ever-wet and evergreen New Zealand forests, particularly those living in the south, and in similar regions in Tasmania and Patagonia. Even the largest New Zealand Jurassic conifer leaves, those of Podozamites, are smaller than those of Agathis australis, which lives today in northernmost New Zealand. This suggests broadly similar temperatures to present-day mid- and southern New Zealand, and cooler than the far north. These figures generally agree with those determined for sea water from belemnite guards (Clayton & Stevens Citation1967; Stevens Citation1971; Stevens & Clayton Citation1971). With respect to T. daintreei, Drinnan & Chambers (Citation1985, 92) wrote “It is tempting to suggest that the abundance of leaf remains in some deposits may represent autumnal banks of leaves and that the species was deciduous.” Whereas this might suggest seasonally very cold conditions, a deciduous habit might also reflect a long polar night in high-latitude conditions.
For the Curio Bay forest environment, Pole (Citation2001) suggested that the climate was less humid, although not arid, with seasonal rainfall. Coal is virtually absent from the system. By contrast, Thorn (Citation2001) noted fossil forest horizons at Kawhia Harbour that were rooted in coal, and she interpreted conditions as comparable to the modern mesothermal zone – with humid, warm conditions, high average rainfall and with large storms.
Biodiversity
Based on foliage macrofossils, New Zealand as a whole has a relatively low diversity of Jurassic plants (with a total of about 43 species); also the per-site diversity is strikingly low. Many assemblages contain little more than Cladophlebis and Taeniopteris, with the Elatocladus group of conifers the next most abundant (Fig. ). Most other taxa are much less common. For instance, in more than 25 years working in the MT Jurassic, the author has collected only one Ptilophyllum. The New Zealand flora shared species with surrounding Gondwanan areas, such as Australia, Antarctica and Patagonia (e.g. Grant-Mackie et al. Citation2000), but there is no evidence yet of stronger ties with any particular one of those areas. The New Zealand Jurassic seems to have been essentially a depauperate version of the Australian flora. If the identification of Hausmannia is correct, then an assemblage from the Otara coast correlates with that described from Queensland by McLoughlin & Drinnan (Citation1995).
New Zealand's low-diversity Jurassic flora might be partly a function of a very unstable landscape, but is more likely a reflection of a suboptimal climate at high latitudes. Some low-diversity communities are likely to reflect specific habitats rather than climate per se, and such associations would match similar low-diversity communities elsewhere on the globe. For instance, in the southernmost MT the author has found exclusively Equisetum-dominated zones (M. Pole, pers. obs.), which might be similar to the extensive Equisetum stands envisaged by Batten (Citation1974) for the English Wealden in areas of shallow standing water, such as delta tops.
As noted above, Rees et al. (Citation2000) assigned Jurassic leaf genera globally to 10 morphological categories and found that maximum morphological diversity was at mid-latitudes in both hemispheres, peaking at 9–10 “morphocategories”. The New Zealand Jurassic had at least four morphocategories (sphenophytes, ferns, microphyllous cycadophytes, and unassigned and microphyllous conifers) and lacked ginkgophytes and macrophyllous conifers. This was, therefore, somewhat below the maximum achievable morphodiversity.
The total macrofossil diversity of New Zealand is comparable with that found in just the single Mid-Jurassic (cf. Hunter et al. Citation2005 for discussion of the age) locality of Hope Bay, on the Antarctic Peninsula, where 43 species were accepted by Gee (Citation1989) and 37 by Rees & Cleal (Citation2004). Continuing research may add a few genera to New Zealand's Jurassic flora, but it may also reduce the species list. In any case, the diversity of most localities is likely to remain low.
Conclusions
There is an abundant record of New Zealand's Jurassic vegetation. The combined flora was relatively low in diversity, dominated by conifers, ferns, bennettitaleans, pentoxylaleans and locally, equisetaleans. Other groups, such as ginkgophytes, which are important elsewhere, were absent. Some of the species documented from New Zealand, such as the cosmopolitan Coniopteris hymenophylloides, were already known from elsewhere and some described from New Zealand have since been discovered elsewhere, as was P. bartrumi in Australia, but many of the species from New Zealand remain endemic taxa. However, since these species were described, the Jurassic floras in surrounding lands, such as Antarctica, have been revised and in some cases re-revised. Revision of New Zealand's Jurassic taxa is long overdue and it might be found that there is less endemicity and stronger links with other regions. However, New Zealand's palaeolocation remains unclear. All of its component pieces lay in relatively high southern latitudes, but the question of whether they were supra- or sub-polar might benefit from further geophysical evidence. With revision of the flora and a clearer picture of where it grew, New Zealand provides a key piece of the global Jurassic picture.
Acknowledgements
The author gratefully acknowledges the helpful comments of S. McLoughlin, S. Turner and V. Vajda. This is a contribution to IGCP 506: Marine/Non-marine Jurassic correlation and was performed within the framework of the Lund University Centre for Studies of Carbon Cycle and Climate Interactions (LUCCI).
Related Research Data
References
- Adams , C.J. , Campbell , H.J. and Griffin , W.L. 2007 . Provenance comparisons of Permian to Jurassic tectonostratigraphic terranes in New Zealand: Perspectives from detrital zircon age patterns . Geological Magazine , 144 : 701 – 729 .
- Arber , E.A.N. 1917 . The earlier Mesozoic floras of New Zealand . New Zealand Geological Survey Palaeontological Bulletin , 6 : 1 – 80 .
- Ballance , P.F. 1988 . “ The Huriwai braidplain delta of New Zealand: A Late Jurassic, coarse-grained, volcanic-fed depositional system in a Gondwana forearc basin ” . In Fan deltas: Sedimentology and Tectonic Settings , Edited by: Nemec , W. and Steel , R.J. 430 – 444 . Glasgow : Blackie & Son .
- Ballance , P.F. and Campbell , J.D. 1993 . “ The Murihiku arc-related basin of New Zealand (Triassic–Jurassic) ” . In Sedimentary Basins of the World , Edited by: Ballance , P.F. Volume 2 , 21 – 33 . Amsterdam : Elsevier . 413 pp
- Bamford , M.K. and Philippe , M. 2001 . Jurassic–Early Cretaceous Gondwanan homoxylous wood: a nomenclatural revision of the genera with taxonomic notes . Review of Palaeobotany and Palynology , 113 : 287 – 297 .
- Barbacka , M. 1994 . Komlopteris Barbacka, gen. nov., a segregate from Pachypteris Brongniart . Review of Palaeobotany and Palynology , 83 : 339 – 349 .
- Batten , D. 1974 . Wealden palaeoecology from the distribution of plant fossils . Proceedings of the Geological Association , 85 : 29 – 458 .
- Bartrum , J.A. 1921 . Note on the Port Waikato Mesozoic flora . New Zealand Journal of Science and Technology , 4 : 1 – 258 .
- Bishop , D.G. , Bradshaw , J.D. and Landis , C.A. 1985 . “ Provisional Terrane map of South Island, New Zealand ” . In Tectonostratigraphic terranes of the circum–Pacific Region , Edited by: Howell , D.G. 515 – 521 . Houston : Circum-Pacific Council for Minerals and Energy .
- Blaschke , P.M. and Grant-Mackie , J.A. 1976 . Mesozoic leaf genus Taeniopteris at Port Waikato and Clent Hills, New Zealand . New Zealand Journal of Geology & Geophysics , 19 : 933 – 941 .
- Broekhuizen, P., 1984: Studies on the Huriwai Group flora, Port Waikato: Taeniopteris Brongniart and Cladophlebis Brongniart. M.Sc. Thesis (Botany), University of Auckland, Auckland, 1–86
- Clayton , R.N. and Stevens , G.R. 1967 . Palaeotemperatures of New Zealand Jurassic and Cretaceous . Tuatara , 16 : 323 – 326 .
- Couper , R.A. 1953 . Upper Mesozoic and Cainozoic spores and pollen grains from New Zealand . New Zealand Geological Survey Paleontological Bulletin , 22 : 1 – 77 .
- Couper , R.A. 1960 . New Zealand Mesozoic and Cainozoic plant microfossils . New Zealand Geological Survey Paleontological Bulletin , 32 : 1 – 87 .
- Creber , G.T. and Chaloner , W.G. 1985 . Tree growth in the Mesozoic and early Tertiary and the reconstruction of palaeoclimates . Palaeogeography, Palaeoclimatology, Palaeoecology , 52 : 35 – 60 .
- Crié , L. 1889 . Beitrage zur Kenntniss der fossilen Flora einiger Inseln des Südpacifischen und Indischen Ocean . Palaeontologia , 5 : 77 – 91 .
- de Jersey , N.J. and Grant Mackie , J.A. 1989 . Palynofloras from the Permian, Triassic and Jurassic of New Caledonia (South Pacific Ocean) . New Zealand Journal of Geology and Geophysics , 32 : 463 – 476 .
- de Jersey , N.J. and Raine , J.I. 1990 . Triassic and earliest Jurassic miospores from the Murihiku Supergroup, New Zealand . New Zealand Geological Survey Paleontological Bulletin , 62 : 1 – 164 .
- de Jersey , N.J. and Raine , J.I. 2002 . Early to Middle Jurassic miospore zonation . IPC 2002, Geological Society of Australia, Abstracts , 68 : 41 – 42 .
- Drinnan , A.N. and Chambers , T.C. 1985 . A reassessment of Taeniopteris daintreei from the Victorian Early Cretaceous: a member of the Pentoxylales and a significant Gondwanaland plant . Australian Journal of Botany , 33 : 89 – 100 .
- Edwards , W.N. 1934 . Jurassic plants from New Zealand . Annals and Magazine of Natural History , 10 : 81 – 109 .
- Frenguelli , J. 1947 . El género Cladophlebis y sus representantes en la Argentina . Anales del Museo de La Plata (Nueva Serie) , 2 : 1 – 74 .
- Gee , C.T. 1989 . Revision of the Late Jurassic/Early Cretaceous flora from Hope Bay, Antarctica . Palaeontographica , 213B : 149 – 214 .
- Graham , I.J. and Mortimer , N. 1992 . Terrane characterisation and timing of metamorphism in the Otago Schist, New Zealand, using Rb–Sr and K–Ar geochronology . New Zealand Journal of Geology and Geophysics , 35 : 391 – 401 .
- Grant-Mackie , J.A. 1995 . “ Aspects of the geology of the Port Waikato region: a field guide ” . In Field Trip Guides, Geological Society of New Zealand 1995 Annual Conference Edited by: O'Connor , B.M. and Ballance , P.F. 52 – 67 .
- Grant-Mackie , J.A. , Aita , Y. , Balme , B.E. , Campbell , H.J. , Crallinor , A.B. , MacFarlane , D.A.B. , Molnar , R.E. , Stevens , G.R. and Thulborn , R.A. 2000 . Jurassic palaeobiogeography of Australasia . Association of Australasian Palaeontologists Memoir , 23 : 311 – 353 .
- Guy-Ohlson , D. 1974 . Paleobotanisk exkursion i Nya Zeelands jura: Curio Bay – den förstenade skogen . Fauna och Flora , 6 : 271 – 282 .
- Hallam , A. , Crame , J.A. , Mancenido , M.O. , Francis , J. and Parrish , J.T. 1993 . Jurassic climates as inferred from the sedimentary and fossil record [and Discussion] . Philosophical Transactions: Biological Sciences , 341 : 287 – 296 .
- Harris , T.M. 1962 . The occurrence of the fructification Carnoconites in New Zealand . Transactions of the Royal Society of New Zealand, Geology , 1 : 17 – 27 .
- Hector , J. 1886 . Detailed catalogue and guide to the geological exhibits, New Zealand Court, Indian and Colonial Exhibition, London , Wellington : Government Printer .
- Herbst , R. 1975 . Consideraciones sobre las Dipteridaceae (Filices) del Gondwana . Actas I Congreso Argentino de Paleontologia y Biostratigrafia, Tucumán, Argentina , 1 : 525 – 535 .
- Hunter , M.A. , Cantrill , D.J. , Flowerdew , M. and Millar , I.L. 2005 . Middle Jurassic age for the Botany Bay Group implications for Weddell Sea creation and southern hemisphere biostratigraphy . Journal of the Geological Society of London , 162 : 745 – 748 .
- Kamp , P.J.J. 2001 . Possible Jurassic age for part of Rakaia Terrane: Implications for tectonic development of the Torlesse accretionary prism . New Zealand Journal of Geology and Geophysics , 44 : 185 – 203 .
- Kidston , R. and Gwynne-Vaughan , D.T. 1907 . On the fossil Osmundaceae. Part I . Transactions of the Royal Society of Edinburgh , 45 : 759 – 780 .
- Kidston , R. and Gwynne-Vaughan , D.T. 1908 . On the fossil Osmundaceae. Part II . Transactions of the Royal Society of Edinburgh , 46 : 213 – 232 .
- Kidston , R. and Gwynne-Vaughan , D.T. 1909 . On the fossil Osmundaceae. Part III . Transactions of the Royal Society of Edinburgh , 46 : 651 – 667 .
- Kidston , R. and Gwynne-Vaughan , D.T. 1910 . On the fossil Osmundaceae. Part IV . Transactions of the Royal Society of Edinburgh , 47 : 455 – 477 .
- Landis , C.A. , Campbell , H.J. , Aslund , T. , Cawood , P.A. , Douglas , A. , Kimbrough , D.L. , Pillai , D.D.L. , Raine , J.I. and Willsman , A. 1999 . Permian–Jurassic strata at Productus Creek, Southland, New Zealand: Implications for terrane dynamics of the eastern Gondwanaland margin . New Zealand Journal of Geology and Geophysics , 42 : 255 – 278 .
- Marshall , P. 1926 . A new species of Osmundites from Kawhia, New Zealand . Transactions of the New Zealand Institute , 56 : 210 – 203 .
- McLoughlin , S. and Drinnan , A.N. 1995 . A Middle Jurassic flora from the Walloon Coal Measures, Mutdapilly, Queensland, Australia . Memoirs of the Queensland Museum , 38 : 257 – 272 .
- McLoughlin , S. , Tosolini , A.-M. , Nagalingum , N. and Drinnan , A.N. 2002 . The Early Cretaceous (Neocomian) flora and fauna of the lower Strzelecki Group, Gippsland Basin, Victoria, Australia . Association of Australasian Palaeontologists Memoirs , 26 : 1 – 144 .
- McQueen , D.R. 1955 . Revision of supposed Jurassic angiosperms from New Zealand . Nature , 175 : 177 – 178 .
- Mildenhall , D.C. 1970 . Checklist of valid and invalid plant macrofossils from New Zealand . Transactions of the Royal Society of New Zealand , 8 : 77 – 89 .
- Miller , C.N. 1967 . Evolution in the fern genus Osmunda . Contributions Museum of Paleontology of the University of Michigan , 21 : 139 – 203 .
- Miller , C.N. Jr. 1999 . Implications of fossil conifers for the phylogenetic relationships of living families . Botanical Review , 65 : 239 – 277 .
- Mortimer , N. 1993 . Jurassic tectonic history of the Otago Schist, New Zealand . Tectonics , 12 : 237 – 244 .
- Mortimer , N. and Campbell , H.J. 1996 . “ Devonian to Jurassic rocks in New Zealand: Classification, content and Gondwana context ” . In Ninth International Gondwana Symposium, Hyderabad, India, January 1994 , vol. 2 , 788 – 790 . Rotterdam : Balkema .
- Nagalingum , N.S. and Cantrill , D.C. 2006 . Early Cretaceous Gleicheniaceae and Matoniaceae (Gleicheniales) from Alexander Island, Antarctica . Review of Palaeobotany and Palynology , 138 : 73 – 93 .
- Norris , G. 1968 . Plant microfossils from the Hawks Crag Breccia, south-west Nelson, New Zealand . New Zealand Journal of Geology and Geophysics , 11 : 312 – 344 .
- Norris , G. and Waterhouse , J.B. 1970 . Age of the Hawks Crag Breccia . Transactions of the Royal Society of New Zealand. Earth Sciences , 7 : 241 – 250 .
- Oliver , W.R.B. 1950 . The fossil flora of New Zealand . Tuatara , 3 : 1 – 11 .
- Park , J. 1887 . On the Jurassic rocks of the Hokonui Hills, Mataura, and Waikawa . New Zealand Geological Survey Report of Geological Explorations 1886–1887 , : 141 – 153 .
- Parris , K.M. , Drinnan , A.N. and Cantrill , D.J. 1995 . Palissya cones from the Mesozoic of Australia and New Zealand . Alcheringa , 19 : 87 – 111 .
- Parrish , J.T. 1991 . “ Jurassic climate and oceanography of the Pacific region ” . In The Jurassic of the circum-Pacific , Edited by: Westermann , G.E.G. 365 – 379 . Cambridge : Cambridge University Press .
- Pole , M.S. 1999 . Structure of a near-polar latitude forest from the New Zealand Jurassic . Palaeogeography, Palaeoclimatology, Palaeoecology , 147 : 121 – 139 .
- Pole , M. 2001 . Repeated flood events and fossil forests at Curio Bay (Middle Jurassic), New Zealand . Sedimentary Geology , 144 : 223 – 242 .
- Pole , M. 2004 . Early–Middle Jurassic stratigraphy of the Fortrose–Chaslands region, southernmost South Island, New Zealand . New Zealand Journal of Geology and Geophysics , 47 : 129 – 139 .
- Raine , J.I. and Pole , M.S. 1988 . Middle Jurassic forest beds, Curio Bay . New Zealand Geological Survey Record , 33 : 47 – 57 .
- Raine , J.I. , Mildenhall , D.C. and Kennedy , E.M. 2008 . New Zealand fossil spores and pollen: An illustrated catalogue , 3rd ed. , GNS Science miscellaneous series no. 4 http://www.gns.cri.nz/what/earthhist/fossils/spore_pollen/catalog/index.htm
- Rees , P.M. 1993 . Dipterid ferns from the Mesozoic of Antarctica and New Zealand and their stratigraphical significance . Palaeontology , 36 : 637 – 656 .
- Rees , P.M. and Cleal , C.J. 2004 . Lower Jurassic floras from Hope Bay and Botany Bay, Antarctica . Special Papers in Palaeontology , 72 : 1 – 90 .
- Rees , P.M. , Ziegler , A.M. and Valdes , P.J. 2000 . “ Jurassic phytogeography and climates: New data and model comparisons ” . In Warm Climates in Earth History , Edited by: Huber , B.T. , Macleod , K.G. and Wing , S.L. 297 – 318 . Cambridge : Cambridge University Press .
- Retallack , G.J. 1983 . Middle Triassic megafossil marine algae and land plants from near Benmore Dam, southern Canterbury, New Zealand . Journal of the Royal Society of New Zealand , 13 : 129 – 154 .
- Speden , I.G. 1958 . A note on the age of the Jurassic flora of Owaka Creek, south-east Otago, New Zealand . New Zealand Journal of Geology and Geophysics , 1 : 530 – 532 .
- Stevens , G.R. 1971 . Relationship of isotopic temperatures and faunal realms to Jurassic–Cretaceous Paleogeography, particularly of the south-west Pacific . Journal of the Royal Society of New Zealand , 1 : 145 – 158 .
- Stevens , G.R. 1980 . Southwest Pacific faunal palaeobiogeography in Mesozoic and Cenozoic times: a review . Palaeogeography, Palaeoclimatology, Palaeoecology , 31 : 153 – 196 .
- Stevens , G.R. 1985 . Lands in collision: Discovering New Zealand's past geography , Wellington : Science Information Publishing Centre . 128 pp
- Stevens , G.R. and Clayton , R.N. 1971 . Oxygen isotope studies on Jurassic and Cretaceous belemnites from New Zealand and their biogeographic significance . New Zealand Journal of Geology and Geophysics , 14 : 829 – 879 .
- Stevens , G.R. and Speden , I.G. 1978 . “ New Zealand ” . In The Phanerozoic Geology of the World II , Edited by: Moullade , M. and Nairn , A.E.M. 251 – 328 . Amsterdam : Elsevier . The Mesozoic
- Tenison-Woods , J.E. 1883 . On the fossil flora of the coal deposits of Australia . Proceedings of the Linnean Society of New South Wales , 8 : 1 – 131 .
- Thorn , V. 2001 . Vegetation communities of a high paleolatitude Middle Jurassic forest in New Zealand . Palaeogeography, Palaeoclimatology, Palaeoecology , 168 : 273 – 289 .
- Thorn , V. 2005 . A Middle Jurassic fossil forest from New Zealand . Palaeontology , 48 : 1021 – 1039 .
- Tidwell , W.D. 1994 . Ashicaulis, a new genus for some species of Millerocaulis (Osmundaceae) . Sida , 16 : 253 – 261 .
- Townrow , J.A. 1967 . On Rissikia and Mataia, podocarpaceous conifers from the lower Mesozoic of southern lands . Papers and Proceedings of the Royal Society of Tasmania , 101 : 103 – 136 .
- Unger , F. 1864 . Fossile Pflanzenreste aus Neu-Seeland. Paläontologie von Neu–Seeland, Novara–Expedition . Th. 1: Bd. 2, 1–13. Der kaiserlich–koniglichen Hof-und Staatsdruckerei in Commission bei Karl Gerold's Sohn, Wien
- Wolfe , J.A. 1995 . Paleoclimatic estimates from Tertiary leaf assemblages . Annual Review Earth and Planetary Sciences , 23 : 119 – 142 .