Abstract
Ammonite biostratigraphy and its correlation potential are evaluated for Ataxioceras-bearing deposits in the eastern Iberian Range, E–NE Spain. The Chronostratigraphic Standard Hypselocyclum Zone of sub-Mediterranean areas is analysed. Correlation of the new Ataxioceras lothari Biozone is proposed utilising first appearance datum (FAD)-based biostratigraphy, resulting from a review of ammonite (Ataxioceratinae) systematics. The lower boundary of the A. lothari Biozone and its chronostratigraphic equivalent Lothari Chronozone are defined by the FAD of Ataxioceras Fontannes s.s., and their upper boundaries by the FAD of Crussoliceras Enay. Distinction of the sub-biozones (subzones) A. hippolytense and A. lothari is confirmed, but these units are slightly reinterpreted. No ammonite subdivisions are defined for the A. hippolytense subzone, although two informal “intra-subzone biodivisions” (A. lothari and Geyericeras aragoniense) are recognised within the A. lothari subzone, the latter defined for the first time. The correlation potential of the proposed biostratigraphic scheme (zone and intra-zone levels using Ataxioceratinae) is high for marine successions from both epicontinental and oceanic-fringe environments under Tethyan influence (southern Europe and north-western Africa). The potential for correlation at the intra-zone level is higher for epicontinental than for oceanic-fringe areas; yet the precise correlation of some biostratigraphic boundaries is difficult at the “intra-subzone biodivision” level between the eastern Iberian Range and SE France. Updated identification and subdivision of the equivalent Metahaploceras strombecki Biozone in oceanic-fringe sections are presented, together with this unit's correlation potential with respect to the proposed A. lothari Biozone.
Introduction
The record of ataxioceratin ammonites in the epicontinental, sub-Mediterranean Jurassic ammonite province offers good opportunities for biostratigraphic zonation and correlation of lower Kimmeridgian deposits, thereby potentially improving chronostratigraphic resolution. However, early Kimmeridgian Ataxioceratinae systematics is still poorly resolved as a consequence of the ammonite assemblages available to, and the taxonomic approaches applied by, previous workers (e.g. Atrops Citation1982; Hantzpergue Citation1989). In this paper, biostratigraphic zonation and correlations are based on a recent revision of the lower Kimmeridgian Ataxioceratinae, including Ataxioceras Fontannes (L. Moliner, PhD thesis submitted). These ammonites are assumed to range throughout the Ataxioceras hypselocyclum Biozone (e.g. Hantzpergue et al. Citation1997), which corresponds to the chronostratigraphic Hypselocyclum Zone within the sub-Mediterranean lower Kimmeridgian (a secondary standard of Callomon Citation1984). Although significant progress in ammonite biostratigraphy has led to better resolution of the lower Kimmeridgian in Tethyan epicontinental and oceanic-fringe settings (i.e. the epioceanic environment in Olóriz Citation1997, Citation2000) (cf. Hantzpergue et al. Citation1997 for a valuable revision), the A. hypselocyclum Biozone is still open to alternative categorisation and subdivision (Geyer Citation1961; Enay et al. Citation1971; Atrops Citation1982).
Three major alternatives exist for interpreting the temporal range of the Standard Hypselocyclum Zone in the sub-Mediterranean province, according to alternative interpretations of the A. hypselocyclum Biozone: (i) to use the stratigraphic range of the registered acme of Ataxioceras and Parataxioceras, which embraces the first appearance datum (FAD) of Garnierisphinctes below the first record of “Katroliceras” (reinterpreted at present as belonging to genus Crussoliceras) and Crussoliceras sensu Geyer (Citation1961, see Table 107); (ii) to use the acme interval for the same genera in combination with the FAD of “katroliceratins”, occurring in the uppermost part of the acme interval (Enay et al. Citation1971). The “katroliceratins” were reinterpreted as Crussoliceras by Enay et al. (Citation1971), a reinterpretation that remains valid; or (iii) to use the acme range of Ataxioceras (i.e. in the form of an assemblage zone) as reinterpreted by Atrops (Citation1982), i.e. incorporating the total range for Ammonites lothari Oppel, Ataxioceras (Parataxioceras) oppeli Geyer and the Perisphinctes (Ataxioceras) hypselocyclum Fontannes complex, including the earliest records of Crussoliceras and Garnierisphinctes within the upper part (Atrops Citation1982, p. 327; Hantzpergue et al. Citation1997).
Thus, whatever the option favoured, the identification of the Standard Hypselocyclum Zone derives from the application of acme zones (biostratigraphic units), i.e. the acme of Ataxioceras and Parataxioceras. Based on these hypotheses, the lower boundary is indirectly marked by the last appearance datum (LAD) of Sutneria platynota (Reinecke) (Moliner & Olóriz Citation2009a, this volume). In contrast, the upper boundary has been variably placed below or above the FADs of Crussoliceras Enay and Garnierisphinctes Enay. This fact is significant for correlation, since it invokes stratigraphic-range differences in the A. hypselocyclum Biozone and the overlying Crussoliceras divisum Biozone in different areas.
The studied area
The study area within the eastern Iberian Range of E–NE Spain was part of a widespread, epicontinental shelf system during the Late Jurassic (Fig. ). During the Kimmeridgian, the epicontinental sea that covered a large part of the Iberian Plate was confined in this region: by the Iberian Massif to the west, the Ebro and Armorican massifs to the north, and the Catalonian Massif and the Tarragona High to the northeast. The Montalban-Ejulve High constituted a raised bottom tract in the central part of the studied area (Aurell et al. Citation2002).
Fig. 1 Location and palaeogeography of the studied sections in E–NE Spain with indication of ammonite records relevant for biostratigraphic correlation. Lithological columns show limestones, marly-limestones and marls from the Loriguilla Formation. Numbers of topographic sheets (1:50,000) for location: 467 (Muniesa), 494 (Calanda), 495 (Castelserás), 518 (Montalbán), 519 (Aguaviva), 543 (Villarluengo). Palaeogeography according to Olóriz (Citation2000).
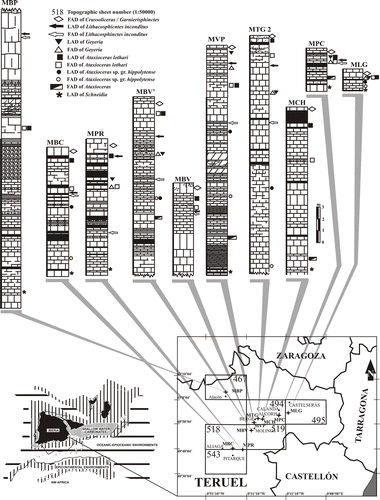
The studied lower Kimmeridgian sediments have been ascribed to an epicontinental marine setting, locally and episodically subjected to open oceanic influences on outer ramp areas (Moliner & Olóriz Citation1999; Olóriz Citation2000; Aurell et al. Citation2002). Moliner & Olóriz (Citation1999) and Olóriz (Citation2000) interpreted depositional depths to range between 30 and 80 m, and sedimentation was influenced by sea-level variation under low energy and relatively high turbidity conditions. These environmental conditions favoured both the accumulation of hemipelagites and imposed strong ecological controls on the composition of the local ammonite community.
In the studied sector, early Kimmeridgian Ataxioceratinae are fairly common, and detailed sampling of favourable sections (Fig. ) allowed identification of evolutionary events that are relevant for biostratigraphy. Ataxioceratinae showing double rib furcations independent of rib density occur slightly above the LAD of Sutneria platynota (Reinecke). This is interpreted as an evolutionary event (cladogenesis), in which novel sculpture is developed as a function of genetic changes (e.g. Callomon in Donovan et al. Citation1981), and this determines the FAD of Ataxioceras Fontannes s.s. In addition, no stratigraphic overlap has been registered between Ataxioceras Fontannes and Parataxioceras Schindewolf with respect to Crusssoliceras Enay and Garnierisphinctes Enay within the study area.
Moliner (Citation1983), Atrops & Meléndez (Citation1984), Finkel (Citation1992), Meléndez et al. (Citation1999) and Moliner & Olóriz (Citation1999) have all carried out ammonite biostratigraphic research on the same interval in this area. The following comments refer only to papers including data on Ataxioceras.
Moliner (Citation1983) interpreted the stratigraphic interval between the LAD of Sutneria platynota (Reinecke) and the common occurrence of Crussoliceras and Garnierisphintes as identifiying the A. hypselocyclum Biozone (using corrected nomenclature here and below according to the International Stratigraphic Guide) in the Gallipuén area, a section at least 24 m thick. He subdivided the A. hypselocyclum Biozone identified into a lower “Hypselocyclum I” and an upper “Hypselocyclum II” interval. Later discovery of Sutneria platynota (Reinecke) within the lower interval led to Moliner & Olóriz (Citation1999) to restrict the use of the name A. hypselocyclum Biozone to the upper subdivision. Rare records of Crussoliceras and Garnierisphintes from the top of the A. hypselocyclum Biozone were reported by Moliner (Citation1983), although these ammonites were all collected from beds above the LAD of Ataxioceras in the studied sections.
In comparatively thick sections from the same region of the eastern Iberian Range, Atrops & Meléndez (Citation1984, p. 384–385) reported the occurrence of ammonite assemblages with Ataxioceras and Parataxioceras as typical elements to identify the A. hypselocyclum Biozone in the area. They also identified typical Crussoliceras species just above the records of Ataxioceratinae from the A. hypselocyclum Biozone, and Garnierisphinctes semigarnieri Geyer below beds with an ammonite fauna typical of the Aspidoceras acanthicum Biozone (all zones labelled as “bio-chronostratigraphic” units, although these authors used biostratigraphic criteria). Remarkably, however, the same authors mentioned an ammonite assemblage containing Garnierisphinctes, Crussoliceras, Ataxioceras and Parataxioceras from a 1 m-thick interval, which they interpreted as the local, thin expression of the A. hypselocyclum Biozone, without discarding the possibility that some beds might belong to the overlying C. divisum Biozone (Atrops & Meléndez Citation1984, p. 381–382).
In relatively extensive deposits of the same area, Moliner & Olóriz (Citation1999) studied a 23 m-thick section and reported an unequivocal but comparatively poor ammonite assemblage with Ataxioceratinae typical of the A. hypselocyclum Biozone but no evidence of crussoliceratins, except in beds above the LAD of Ataxioceras.
All of the above essentially agree with our data concerning the lack of unequivocal records of overlapping ranges of Ataxioceras and Parataxioceras with respect to Crussoliceras and Garnierisphinctes in the area. Additional information was supplied by Finkel (Citation1992), who studied Kimmeridgian ataxioceratins, including Parataxioceras Schindewolf and Ataxioceras Fontannes, collected from the eastern part of the same region. However, Finkel (Citation1992, p. 236–237, Figs. 24, 26, 27, 29, 30) interpreted species typical of the upper part of the Sutneria platynota Biozone [i.e. Schneidia sp. cf. S. guilherandense Atrops, Schneidia sp. cf. S. elmii Atrops, Schneidia sp. cf. S. collignoni Atrops and Lithacosphinctes pseudoachilles (Wegele)] as belonging to the A. hypselocyclum Biozone. Unfortunately, Finkel (Citation1992) gave synthetic biostratigraphic ranges that do not allow interpretation of within-biozone ranges in the area.
Material and methods
Ten sections (MBP–MPR–MBC–MBV′–MBV–MVP–MTG2–MCH–MPC–MLG; Fig. ) were sampled in the eastern Iberian Chain (Aragonese Branch) and northern Maestrazgo region (province of Teruel, Spain). The studied lower Kimmeridgian succession belongs to the Calanda Member (Aurell Citation1990) of the Loriguilla Formation (Gómez & Goy Citation1979), a limestone to marly-limestone rhythmite usually about 20 m thick. Limestone beds reach 0.5 m thick, whereas intercalations of marly limestones and marls may exceed 1 m thick. The stratigraphic interval investigated is included in sequence J3.5 interpreted by Aurell et al. (Citation2002) to have been deposited within an outer ramp environment.
The stratigraphic sections were measured, and bed-by-bed sampling was conducted on fossil macroinvertebrates. A total of 759 ammonites were obtained, 41% of them belonging to subfamily Ataxioceratinae. This material was prepared in the laboratory before its study, and is lodged with the University of Granada.
Conducted under a palaeobiological approach, the revision of Ataxioceratinae systematics at the species level forced grouping of microconchs [m] and macroconchs [M] and, therefore, their taxonomic reference to a single species. In the case of resulting large phenotype spectra, mention of particular “historical” forms (e.g. forms matching the characters to which old species names were applied) is given to facilitate morphological “visualisation” in particular cases. In addition, genus-level systematics included some particular cases of re-interpretation of species distribution based on the high-resolution biostratigraphic controls on the interpreted phyletic lineages (L. Moliner, submitted PhD thesis).
Biostratigraphic analysis favoured subdivision based on a FAD approach for both biozones and their subordinate ranks. Since, the terms “horizon” or “biohorizon” (e.g. Callomon Citation1984) are strongly discouraged by the International Stratigraphic Guide, the informal term “intra-subzone biodivision” is used herein to denote biostratigraphic units of lesser rank than sub-biozones. As reported by Callomon (Citation1984), differential orthography has been applied to identify biostratigraphic units (lower case and italics for the index species) and their chronostratigraphic equivalents (normal case and initial capitals for both the index species and the term zone), and different orthography has been applied by various authors. However, nomenclature in this study follows the International Stratigraphic Guide. Zonal schemes obtained from the bibliography, have been modified according to this standard.
Ammonite assemblages and biostratigraphy
Approaching Ataxioceratinae systematics by giving special relevance to palaeobiology involves the nomenclatural treatment of dimorphs within a single species. Hence, ammonite systematics tends to have more biological meaning than mere differentiation into morphotaxa. Hantzpergue (Citation1989) applied this approach to early Kimmeridgian ataxioceratins of Aquitaine, NW France, and a similar approach has been adopted for coeval material from the eastern Iberian Range.
In the study area, ataxioceratins are the best represented ammonites within the stratigraphic interval containing Ataxioceras s.s. The record of higher taxa of Ammonitina such as Taramelliceratinae, Aspidoceratidae and Glochiceratidae increases with respect to assemblages from the underlying S. platynota Biozone; secondary components are Streblitinae and Passendorferiinae [e.g. Nebrodites; cf. Caracuel et al. (Citation1999) for comments about early Kimmeridgian Passendorferiinae].
In the context of revising the systematics of Ataxioceras Fontannes and related Ataxioceratinae (L. Moliner, PhD thesis submitted), successive ammonite assemblages and biostratigraphic units (zones, subzones and informal “intra-subzone biodivisions”) have been characterised (Fig. ): (i) the A. lothari Biozone, and its chronostratigraphic equivalent Lothari Chronozone; (ii) the A. hyppolytense Subzone in the lower part, and its chronostratigraphic equivalent Hippolytense Chronozone (Fig. ); and (iii) the A. lothari Subzone in the upper part (and the corresponding chronostratigraphic Lothari Subchronozone), subdivided into the A. lothari “intra-subzone biodivision” below (Fig. ) and the Geyericeras aragoniense “intra-subzone biodivision” above (Fig. ). The definition of the A. lothari Biozone, the equivalent Hypselocyclum Chronozone, and their respective subdivisions, together with their correlation potential are discussed further in the section on Biostratigraphic correlation (below). The reference section for all the biostratigraphic units considered below is the MTG2 section in the Gallipuén sector.
Fig. 2 Biostratigraphic correlation chart for the reinterpreted Hypselocyclum Chronozone in epicontinental and oceanic-fringe deposits from southern Europe.
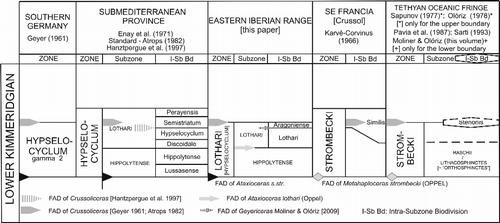
Fig. 3 Three species relevant for biostratigraphy and correlation of the A. lothari Biozone in the eastern Iberian Chain, ENE Spain. A. Ataxioceras lothari lothari (Oppel) [m], MPR.36.5 from the MPR section. B. G. aragoniense Moliner and Olóriz [m], MPC.28.73 from the MPC section. C. Ataxioceras hippolytense Atrops [m], MBV1.13.2 from the MBV section. White stars for the beginning of the body-chamber. All the specimens are housed in the repository of the Palaeontological collection of the University of Granada, Spain.
![Fig. 3 Three species relevant for biostratigraphy and correlation of the A. lothari Biozone in the eastern Iberian Chain, ENE Spain. A. Ataxioceras lothari lothari (Oppel) [m], MPR.36.5 from the MPR section. B. G. aragoniense Moliner and Olóriz [m], MPC.28.73 from the MPC section. C. Ataxioceras hippolytense Atrops [m], MBV1.13.2 from the MBV section. White stars for the beginning of the body-chamber. All the specimens are housed in the repository of the Palaeontological collection of the University of Granada, Spain.](/cms/asset/2802c9ab-6f2f-451d-85c1-6089eb80bdfc/sgff_a_395460_o_f0003g.gif)
Ataxioceras hippolytense subzone
This subzone encompasses the stratigraphic interval between the FAD of Ataxioceras s.s. [m, M], above the LAD of Schneidia Atrops [m, M], and the FADs of Ataxioceras lothari (Oppel) [m, M] and/or endemic Ardescia species [m, M] derived from a conservative lineage documented throughout the S. platynota Biozone. This subzone embraces the total range of Ataxioceras hippolytense Atrops [m, M] in the eastern Iberian range. A bottom-up impoverishment in ammonites has been identified.
The ammonite assemblage comprises endemic species of Ardescia (L. Moliner, PhD thesis submitted), respectively, originating from the older A. desmoides group and from a conservative phyletic lineage registered throughout the older S. platynota Biozone; the oldest records of Lithacosphinctes inconditus (Fontannes) [m, M], Lithacosphinctes sp., Ataxioceras sp. [m, M], and the total range for Ataxioceras hippolytense Atrops [m, M]. In contrast, to the record from SE France (e.g. Atrops Citation1982), the stratigraphic ranges of Ataxioceras hippolytense Atrops [m, M] and Ataxioceras lothari (Oppel) [m, M] do not overlap. Indeterminate Ataxioceratinae [m, M] are also present. Among other ammonites there are Aspidoceras binodum (Oppel), Aspidoceras sesquinodosum (Fontannes), Aspidoceras sp. ex gr. A. linaresi Checa, Aspidoceras spp., Pseudowaagenia micropla (Oppel), Physodoceras wolfi (Neumayr), Physodoceras sp. ex gr. altenense (D'Orbigny), Physodoceras spp.; rare Nebrodites sp. ex gr. hospes (Neumayr); Metahaploceras sp. ex gr. subnereus (Wegele), Metahaploceras sp. ex gr. kobyi (Choffat), Metahaploceras sp. and Taramelliceras spp.; Glochiceras sp. ex gr. nimbatum (Oppel), Glochiceras sp., Lingulaticeras sp. ex gr. lingulatum (Quenstedt), Lingulaticeras sp. ex gr. nudatum (Oppel), Coryceras sp. ex gr. canale (Quenstedt), Ochetoceras canaliferum (Oppel) and Ochetoceras sp. The Streblitinae is represented by Streblites sp.
Ataxioceras lothari subzone
This subzone corresponds to the total range of Ataxioceras lothari (Oppel) [m, M]; the lower boundary coincides with the FADs of this species together with a new endemic, conservative species of Ardescia (L. Moliner, PhD thesis submitted); the upper boundary is marked by the FAD of Crussoliceras Enay [m, M] and Garnierisphinctes Enay [m, M] just above the LAD of Ataxioceras lothari Oppel [m, M]. Ataxioceras lothari (Oppel) [m, M] is typical along with endemic species of Ardescia, which probably also occurs within the lowermost part of the C. divisum zone, and the new genus Geyericeras Moliner & Olóriz (Citation2009b).
The ammonite assemblage is composed of new Ardescia species [m, M] (L. Moliner, PhD thesis submitted), Ataxioceras lothari (Oppel) morphotype lothari Oppel [m, M] and chronotype semistriatum Schneid [m], Ataxioceras n. sp. A [M], Ataxioceras sp. [m, M], Parataxioceras sp. gr. P. evolutum Atrops [m, M], Parataxioceras sp. gr. P. planulatum (Quenstedt) [m], Lithacosphinctes inconditus (Fontannes) [m, M], Lithacosphinctes n. sp. ex gr. L. perayensis (Atrops) [m], Lithacosphinctes sp., G. aragoniense Moliner and Olóriz [m, M] and indeterminate Ataxioceratinae [m, M]. Among ammonites other than the Ataxioceratinae there are Aspidoceras binodum (Oppel), Aspidoceras sesquinodosum (Fontannes) and related forms, Pseudowaagenia micropla (Oppel), Physodoceras wolfi (Neumayr) and indeterminate aspidoceratins; and rare Nebrodites hospes (Neumayr) morphotype minor Quenstedt and Nebrodites sp.; Metahaploceras sp. ex gr. subnereus (Wegele), Metahaploceras sp. ex gr. kobyi (Choffat), Metahaploceras sp. and Taramelliceras sp.; Glochiceras sp. ex gr. nimbatum (Oppel), Glochiceras sp., Lingulaticeras sp. ex gr. lingulatum (Quenstedt), Lingulaticeras sp., Coryceras sp. ex gr. canale (Quenstedt), Coryceras sp. ex gr. modestiforme, Ochetoceras canaliferum (Oppel) and Ochetoceras sp. The Streblitinae is again represented by Streblites sp.
A. lothari subzone–Lothari “intra subzone biodivision”
The lothari “intra-subzone biodivision” is defined by the lower boundary of the lothari Subzone below and the FAD of Geyericeras Moliner & Olóriz (Citation2009b) above.
The ammonite assemblage is composed of Lithacosphinctes inconditus (Fontannes) [m, M], Parataxioceras sp. gr. P. evolutum Atrops [m, M], Ataxioceras lothari (Oppel) morphotype lothari OPPEL [m, M], a new endemic species of Ardescia (L. Moliner, PhD thesis submitted) and indeterminate Ataxioceratinae [m, M]. Other ammonites are Aspidoceras sp. ex gr. A. linaresi Checa, Physodoceras wolfi (Neumayr) and indeterminate aspidoceratins; rare Nebrodites hospes (Neumayr) morphotype minor Quenstedt, Nebrodites sp., and indeterminate Rasenidae; Glochiceras sp. ex gr. G. nimbatum Oppel, Lingu-laticeras sp. ex gr. L. lingulatum (Quenstedt) and Lingulaticeras sp.; Metahaploceras sp. ex gr. subnereus (Wegele), Metahaploceras sp. and Streblites sp.
A. lothari subzone–G. aragoniense “intra-subzone biodivision”
The aragoniense “intra-subzone biodivision” encompasses the stratigraphic interval between the FAD of Geyericeras Moliner & Olóriz (Citation2009b) and the FADs of Crussoliceras Enay [m, M] and Garnierisphinctes Enay [m, M].
The ammonite assemblage is made up of Lithacosphinctes inconditus (Fontannes) [m, M], Lithacosphinctes n. sp. ex gr. L. perayensis (Atrops) [m], Lithacosphinctes sp., Parataxioceras sp. ex gr. P. evolutum Atrops [m, M], Ataxioceras lothari (Oppel) morphotypes lothari Oppel [m, M] and semistriatum Schneid [m], Ataxioceras sp. [m, M], G. aragoniense Moliner and Olóriz [m, M], a new and morphologically conservative species of Ardescia (L. Moliner, PhD thesis submitted) and indeterminate Ataxioceratinae [m, M]. Among other ammonites there are Aspidoceras sp. gr. A. linaresi Checa, Aspidoceras sp. gr. A. binodum (Oppel) – sesquinodosum (Fontannes), Aspidoceras sesquinodosum (Fontannes), Pseudowaagenia micropla (Oppel), Physodoceras wolfi (Neumayr) and indeterminate aspidoceratins; rare Nebrodites sp.; Glochiceras s.l., Metahaploceras sp. ex gr. subnereus (Wegele), Metahaploceras sp. and Streblites sp.
Biostratigraphic correlation
As explained in the introductory section, three major alternative definitions have been applied to the A. hypselocyclum Biozone in different regions of sub-Mediterranean Europe, resulting in different application of the respective, local biostratigraphic and corresponding chronostratigraphic units (Hypselocyclum Zone). According to the reinterpretation of Ataxioceras Fontannes mentioned above, i.e. based on the evolutionary significance given to the occurrence of clear double furcations unrelated to rib density above the LAD of Sutneria platynota (Reinecke), the lower boundary of the Standard Hypselocyclum Chronozone is clearly recognised by the FADs of Ataxioceras s.s. and Parataxioceras (but see Atrops Citation1982). Hence, a primary distinction between our results and the aforementioned interpretations is that the base of the proposed Hypselocyclum Chronozone is defined on the basis of FADs instead of acmes. The remaining difference between the three alternative hypotheses resides in the interpretation of the upper boundary, for which we recommend the use of a FAD amongst ammonite species. Geyer (Citation1961) used the FAD of katroliceratins (“Katroliceras” and Crussoliceras), which are characteristic fossils in terms of sculpture. Atrops (Citation1982) favoured the use of the LADs of Ataxioceras and Parataxioceras. As usual when the use of LADs is preferred for definition of biostratigraphic units, it involves an eco-bioevent that can be considered as comparatively volatile with respect to FADs in terms of precise biostratigraphy – even in proven cases of FAD diachrony (e.g. see Marques Citation1983; Atrops & Marques Citation1986; Olóriz et al. Citation1992; Marques et al. Citation1998 for Ardescia; and Karvé-Corvinus Citation1966 for the acme of Ataxioceras in SE France and southern Germany). Meanwhile, LADs are assumed to be comparatively prone to local variation due to ecological factors. In addition, LADs are used within a common context of progressive impoverishment in the particular record of the given ammonite species.
Another alternative for definition of the A. hypselocyclum Biozone results from the revision of the systematics of lower Kimmeridgian Ataxioceratinae (L. Moliner, PhD thesis submitted) and the application of biostratigraphic units defined by lower boundaries based on the priority of FADs (Fig. ). Thus, updated correlation with the Standard Hypselocyclum Chronozone can be evoked by using FADs: (i) the lower boundary is marked by the FAD of Ataxioceras s.s., which most probably occurred slightly after the LAD of Sutneria platynota (Reinecke) according to available biostratigraphy; and, (ii) the upper boundary is marked by the FAD of Crussoliceras.
Perisphinctes (Ataxioceras) hypselocyclus (Fontannes Citation1879) is interpreted to be the macroconchiate counterpart of Ammonites lothari (Oppel Citation1863) and, therefore, represents a junior synonym of the latter. Hence, the name of the corresponding biostratigraphic unit in the eastern Iberian Chain is here re-labelled as the A. lothari Biozone. This scheme formalises and emends a “lothari” biostratigraphic unit used long ago to denote “Sequanian” stratigraphic intervals of variable order but characterised by Lower Kimmeridgian Ataxioceratinae above beds with Sutneria platynota (e.g. Buticaz Citation1943; Arkell Citation1956).
The epicontinental environment–sub-Mediterranean shelves
The biostratigraphic range of Ataxioceras s.s. and Parataxioceras, i.e. collected above the LAD of Sutneria platynota Reinecke, is useful for identifying the Standard Hypselocyclum Chronozone (Geyer Citation1961; Hantzpergue et al. Citation1997), in which ammonites belonging to the Ataxioceras hypselocyclum Fontannes group are clearly distinguished among ataxioceratins. Whatever the alternative preferred to interpret this Standard chronozone (see above), the lower boundary is indeed related to the combination of the FAD of Ataxioceras s.s. and the more or less closely related and slightly earlier LAD of Sutneria platynota (Reinecke). In contrast, the upper boundary, which is defined by the base of the overlying C. divisum zone, alternatively includes or excludes the FADs of crussoliceratins (e.g. see above and Hantzpergue et al. Citation1997), ammonites featuring very distinctive sculpture.
In the study area, the lower boundary of the stratigraphic interval yielding Ataxioceras s.s. is defined by the FAD of that genus. Its upper boundary is related to the FAD of Crussoliceras, which seems to occur slightly later in Spain than in SE France according to the ammonite record in the area (see above; Fig. ). This interpretation is an intermediate option between those promoted by Geyer (Citation1961) and Atrops (Citation1982); it favours correlations with biostratigraphic schemes proposed for condensed successions showing ammonitico rosso and related facies, i.e. typical Tethyan oceanic-fringe deposits (Fig. ). Thus, the A. lothari Biozone recognised in the eastern Iberian Chain is interpreted as the Assemblage Zone of Ataxioceras + Parataxioceras (as interpreted here), excluding the semistriatum pro parte and perayensis “horizons” described by Atrops (Citation1982) from the uppermost A. hypselocyclum Biozone he interpreted, since they contain Crussoliceras (see Hantzpergue et al. Citation1997).
Deserving mention is the reinterpretation made by Hantzpergue et al. (Citation1997), who argued that the range overlap between Ataxioceras and Crussoliceras was restricted to the A. semistriatum “intra-subzone biodivision”, as the L. perayensis “intra-subzone biodivision” has neither Ataxioceras nor Parataxioceras. This proposal implies that the upper boundary of the A. lothari Biozone in the eastern Iberian Chain does not correspond to two significant bioevents: (i) the LAD of Ataxioceras + Parataxioceras; and (ii) the FAD of Crussoliceras. The latter would occur within the A. semistriatum “intra-subzone biodivision” defined by Atrops (Citation1982), or at its base (Hantzpergue et al. Citation1997), but above the LAD of Ataxioceras + Parataxioceras. Furthermore, this proposal brought to light two possibilities: (i) the occurrence of minor differences between biostratigraphic ranges in SE France and the eastern Iberian Chain; and (ii) the occurrence of a small stratigraphic gap truncating taxon ranges close to the FAD of Crussoliceras, i.e. close to the base of the C. divisum Biozone in the eastern Iberian Chain.
Relevant for correlation with oceanic-fringe areas is the biostratigraphic range of Metahaploceras strombecki (Oppel) reported by Karvé-Corvinus (Citation1966) from Crussol (SE France), who interpreted its stratigraphic equivalence with the A. hypselocyclum Biozone in Swabia, southern Germany. On this basis, Karvé-Corvinus (Citation1966) identified the M. strombecki Biozone, held to be the precise equivalent of the Standard Hypselocyclum Chronozone (e.g. Atrops Citation1982; Hantzpergue et al. Citation1997), exemplifying the third alternative mentioned above (Fig. ). Although Karvé-Corvinus (Citation1966) did not provide a bed-by-bed biostratigraphy of his ca. 20.5 m-thick Similis-Schichten (levels 33: 335 cm; 34: 640 cm; 35: 290 cm; 36: 405 cm; and 37: 375 cm), in the upper part of his M. strombecki Biozone, he commented on the occurrence of the earliest Katroliceras (mainly interpreted as Crussoliceras at present) within the upper part of his Similis-Schichten. The possibility that these “katroliceratins” = crussoliceratins (i.e. Crussoliceras) did not overlap with Ataxioceras – Parataxioceras in the type section at Crussol, SE France, remains unresolved. However, Atrops (Citation1982) demonstrated their rare co-occurrence in the area (i.e. Crussoliceras was registered from the uppermost A. semistriatum but not from the overlying L. perayensis “intra-subzone biodivision”) with no mention of stratigraphic condensation. It remains to be proven whether this biostratigraphic overlap characterises local taxon ranges or, rather, represents a consistent pattern in epicontinental shelves from the sub-Mediterranean province.
The oceanic-fringe environment–Mediterranean swell-slope settings
Ataxioceras is rare in condensed sections comprising ammonitico rosso and related facies incorporating numerous hiatuses, developed along the Tethyan oceanic fringes (the epioceanic environment of Olóriz Citation2000). Based on ammonite biostratigraphy from southern Spain, Olóriz (Citation1978) promoted the use of the Strombecki Chronozone to correspond to the stratigraphic interval between Platynota Chronozone deposits and the acme of “Katroliceras” and Crussoliceras. In fact, the lower boundary of the M. strombecki Biozone is marked by the FAD of the index species Metahaploceras strombecki (Oppel); see Moliner & Olóriz (Citation2009a, this volume) for an updated interpretation of the S. platynota Biozone in oceanic-fringe areas. Based on reinterpretation of lower Kimmeridgian “Katroliceras” recorded below beds belonging to the C. divisum Zone (see below) and the occurrence of Crussoliceras in Olóriz (Citation1978), the upper boundary of the M. strombecki Biozone proposed by Olóriz (Citation1978) is marked by the FAD of Crussoliceras (see below; Fig. ).
Like the diverse interpretations of biostratigraphic correlations with the Standard Hypselocyclum Chronozone in epicontinental shelves from southern Europe, the Strombecki Chronozone in oceanic-fringe deposits has been subjected to various biostratigraphic definitions (e.g. Olóriz Citation1978; Pavia et al. Citation1987; Sarti Citation1993; Caracuel et al. Citation1998). Thus, the lower and upper boundaries were interpreted according to the availability of ammonites in sections showing variable stratigraphic condensation regionally (e.g. Olóriz Citation1978; Pavia et al. Citation1987; Sarti Citation1993; Caracuel et al. Citation1998). Yet local taphonomic condensation could also inhibit identification, where local ammonite ranges and sedimentary hiatuses are incompletely documented. Whatever the case, the Strombecki Chronozone represented in ocean-fringe settings was correlated with a significant but variable part of the Standard Hypselocyclum Chronozone identified in epicontinental, sub-Mediterranean shelves.
The use of the Hypselocyclum Chronozone was promoted by Sapunov (Citation1977) for the Forebalkans, but is notably equivalent to the Strombecki Chronozone interpreted by Olóriz (Citation1978) in southern Spain, at least insofar as the upper boundary is defined by the FAD of Crussoliceras. The lower boundary of the Hypselocyclum Chronozone as promoted by Sapunov (Citation1977) is difficult to reinterpret; see Moliner & Olóriz (Citation2009a, this volume) for an updated interpretation of the lower Ataxioceras (Parataxioceras) desmoides Chronozone promoted by Sapunov (Citation1977). Thus, only the upper boundary of Hypselocyclum Chronozone proposed by Sapunov (Citation1977) can be of use for correlation.
Moreover, the biostratigraphic identification and correlation of the lower boundary of the Strombecki Chronozone in oceanic-fringe successions presents no difficulties, in view of both the reinterpretation of the S. platynota Biozone boundaries and the assumption of a small stratigraphic interval between the FAD of Metahaploceras strombecki (Oppel) and the acme of Methaploceras in oceanic-fringe successions (see Moliner & Olóriz Citation2009a, this volume; Fig. ). Hence, the difference noted by Sarti (Citation1993) between the lower boundary of the Strombecki Chronozone in NE Italy and southern Spain (Olóriz Citation1978) is insubstantial.
The biostratigraphic definition of the upper boundary of the Strombecki Chronozone in oceanic-fringe successions must consider the rare records of the youngest Ataxioceras, if present at all, and Metahaploceras, together with the more common records of crussoliceratins, especially Crussoliceras. The FAD of the last of these is, therefore, of value for placing the lower boundary of the overlying Divisum Chronozone based on ammonite biostratigraphy. In such a biostratigraphic context, this option is favoured over some comparatively less consistent alternatives subject to local, environmental forcing factors: (i) the use of acme records (see Karvé-Corvinus Citation1966, for stratigraphic differences between the Ataxioceras acme in Crussol and southern Germany); and (ii) the use of LADs (see above). Additionally, it promotes the use of FADs for defining zone boundaries and utilises a biostratigraphic event, the FAD of Crussoliceras, that has higher correlation potential with epicontinental records (Fig. ) and, therefore, chronostratigraphic significance.
The assumed occurrence of Crussoliceras below the Divisum Chronozone in oceanic-fringe deposits belonging to the Strombecki Chronozone cannot be proven, because the illustrated specimens could belong to Lithacosphinctes (e.g. Pavia et al. Citation1987, pl. 5) and because they show both very shallow umbilica and a particular ribbing style unknown for epicontinental Crussoliceras. In some cases, they could possibly be considered a new genus (e.g. Sarti Citation1993, pl. 9). Furthermore, the early reference to “Katroliceras” occurrence in stratigraphic levels below the Divisum Chronozone in S–SE Spain (Olóriz Citation1978) was rightly reinterpreted by Pavia et al. (Citation1987) and, therefore, does not apply to the interpretation made in this paper. On this basis, the Pseudosimoceras stenonis Sub-biozone (Pavia et al. Citation1987) embraces the upper part of the Strombecki Chronozone in oceanic-fringe ammonitico rosso facies plus an imprecisely known part of the overlying Divisum Chronozone in accordance with both the record of “Katroliceras” in S–SE Spain (Olóriz Citation1978) and its reinterpretation (Pavia et al. Citation1987), together with later data from NE Italy (Sarti Citation1993). As the precise chronostratigraphic interpretation of the P. stenonis subzone is not conclusive, as is its potential for subdivision, its interpretation as an informal “intra-subzone biodivision” is favoured at present.
Existing proposals for biostratigraphic differentiation of the lower part of the Strombecki Chronozone in oceanic-fringe sections seem to be based on local distribution patterns (e.g. the Lessiniceras raschii Sub-biozone proposed by Sarti Citation1993). They are difficult to correlate, or even require revision (Pavia et al. Citation1987) to clarify their correlation potential with both oceanic-fringe and epicontinental records. Hence, their precise interpretation in biostratigraphic (e.g. Pavia et al. Citation1987; Sarti Citation1993) and chronostratigraphic terms cannot be considered conclusive.
Acknowledgements
This research has been supported by Project BTE2005-01316 and the EMMI Group 178 J.A., Spain. This is a contribution to IGCP 506: Marine/Non-marine Jurassic correlation. We are indebted to the anonymous reviewers and the editors for suggestions and comments on an early draft of this paper.
References
- Arkell , W.J. 1956 . Jurassic Geology of the World , 806 Edinburgh : Oliver & Boyd .
- Atrops , F. 1982 . La sous-Famille des Ataxioceratinae (Ammonitina) dans le Kimméridgien inférieur de Sud-Est de la France: Systématique, évolution, chronostratigraphie des genres Orthosphinctes et Ataxioceras . Documents des Laboratoires de Géologie Lyon , 83 : 1 – 463 .
- Atrops , F. and Marques , B. 1986 . Mise en évidence de la Zone à Platynota (Kimmeridgien inférieur) dans le massif du Montejunto (Portugal); conséquences stratigraphiques et paléontologiques . Geobios , 19 : 537 – 547 .
- Atrops , F. and Meléndez , G. 1984 . “ Kimmeridgian and lower Tithonian from the Calanda–Berge area (Iberian Chain, Spain): Some biostratigraphic remarks ” . In Proceedings of the 1st International Symposium on Jurassic Stratigraphy Edited by: Michelsen , O. and Zeiss , A. Vol. 2 , 377 – 392 . Erlangen
- Aurell, M., 1990: El Jurásico superior de la Cordillera Ibérica Central (provincias de Zaragoza y Teruel). Análisis de Cuenca. Ph.D. Thesis, University of Zaragoza. 389 pp
- Aurell , M. , Meléndez , M. and Olóriz , F. 2002 . “ The Jurassic ” . In The Geology of Spain , Edited by: Gibbons , W. and Moreno , T. 213 – 253 . London : The Geological Society of London . (coords.)
- Buticaz, P., 1943: Ammonites séquaniennnes de Reculet (Jura Français). Ph.D. Thesis, no. 1053, Université de Genevé. Mémoires suisses de Paléontologie 64, 31 pp
- Callomon , J.H. 1984 . “ Biostratigraphy, chronostratigraphy and all that – again! ” . In International Symposium on Jurassic stratigraphy (Erlangen 1984) 3 , Edited by: Michelsen , O. and Zeiss , A. 611 – 624 . Copenhagen : Geological Survey of Denmark .
- Caracuel , J. , Olóriz , F. and Sarti , C. 1998 . Updated biostratigraphy of the Kimmeridgian and Lower Tithonian at Lavarone (Trento Plateau, Italy). Correlation for epioceanic Western Tethys . Geologica et Palaeontologica , 32 : 235 – 251 .
- Caracuel , J. , Olóriz , F. and Rodríguez-Tovar , F.J. 1999 . Ammonite biostratigraphy from the Planula and Platynota Zones in the Lugar section (External Subbetic, southern Spain) . Profil , 16 : 107 – 120 .
- Donovan , D.T. , Callomon , J.H.C. and Howart , M.K. 1981 . “ Classification of the Jurassic Ammonitina ” . In The Ammonoidea. The evolution, classification, mode of life and geological usefulness of a major fossil group , Edited by: House , M.R. and Senior , J.R. 101 – 155 . London : Systematics Association Special Volume 18 .
- Enay , R. , Tintant , H. and Rioult , M. 1971 . “ Kimméridgien ” . In (Groupe Français d'Etude du Jurassique): Les Zones du Jurassique en France. Comptes Rendus Sommaire des Séances de la Société géologique de France , Edited by: Mouterde , R. , Enay , R. , Cariou , E. , Contini , D. , Elmi , S. , Gabilly , J. , Mangold , C. , Mattei , J. , Rioult , M. , Thierry , J. and Tintant , H. Vol. 6 , 97 – 98 . Nancy : Berger-Levraut .
- Finkel , R. 1992 . Eine Ammoniten-Fauna aus dem Kimmeridgium des nordöstlichen Keltiberikums (Spanien) . Profil , 3 : 227 – 297 .
- Fontannes , F. 1879 . Description des ammonites des calcaires du Château de Crussol, Ardèche (zones à Oppelia tenuilobata et Waagenia beckeri) , 123 Paris : Georg., Lyon et Savy F .
- Geyer , O.F. 1961 . Monographie des Perisphinctidae des unteren Unterkimmeridgium (Weisser Jura gamma, Badenerschichten) im süddeutschen Jura . Palaeontographica , 117 : 1 – 157 .
- Gómez , J.J. and Goy , A. 1979 . Las unidades litoestratigráficas del Jurásico medio y superior en facies carbonatadas del Sector Levantino de la Cordillera Ibérica . Estudios Geológicos , 35 : 569 – 598 .
- Hantzpergue , P. 1989 . Les ammonites kimméridgiennes du Haut-Fond d'Europe occidentale. Biochronologie, systématique, évolution, paléobiogéographie. Cahiers de Paleontologie , 1 – 428 . Paris : Centre National de la Recherche Scientifique (éd.) .
- Hantzpergue , P. , Atrops , F. and Enay , R. 1997 . “ Kimméridgien ” . In (coords.): Biostratigraphie du Jurassique Ouest-Européen et Méditerranéen. Zonations parallèles et distribution des invertébrés et microfossiles. Groupe Français Etude Jurassique. Bulletin du centre de recherches Elf Exploration–Production, Mémoir Edited by: Cariou , E. and Hantzpergue , P. Vol. 17 , 87 – 96 .
- Karvé-Corvinus , G. 1966 . Biostratigraphie des Oxfordium und untersten Kimmeridgium am Mont Crussol, Ardèche, im Vergleich mit Süddeutschland . Neues Jarhbuch für Geologie und Paläontologie Abhandlungen , 126 ( 2 ) : 101 – 141 .
- Marques, B., 1983: O Oxfordiano–Kimmeridgiano do Algarve oriental: estratigrafia, paleobiologia (Ammonoidea) e paleobiogeografia. Ph.D. These, Universidade Nova de Lisboa, Lisbon, Portugal. 547 pp
- Marques , B. , Olóriz , F. and Rodríguez-Tovar , F.J. 1998 . Oxfordian–Kimmeridgian boundary as determined by an index species of ammonite (Sutneria) (Algarve, Portugal) . Comptes Rendus de l'Académie des sciences Paris, Sciences de la Terre et des planètes , 326 : 641 – 645 .
- Meléndez , G. , Bello , J. , Delvene , G. , Pérez-Urresti , I. , Ramajo , J. and Atrops , F. 1999 . Middle and Upper Jurassic at the Calanda-Mas de las Matas area, in the region of River Guadalope (NE Iberian Chain, E Spain) . Profil , 16 : 275 – 296 .
- Moliner, L., 1983: El Jurásico superior en el sector de Alcorisa–Berge (Provincia de Teruel). M.Sc. Thesis, Universidad de Granada, Granada, Spain, 197 pp. [inedit.]
- Moliner , L. and Olóriz , F. 1999 . The Lower Kimmeridgian Section at Gallipuén. Ammonite Biochronostratigraphy and Ecostratigraphic Remarks . Profil , 16 : 95 – 106 .
- Moliner , L. and Olóriz , F. 2009a . Correlation potential of Lower Kimmeridgian Platynota Zone deposits in northeast Spain . GFF , 131 (this issue)
- Moliner , L. and Olóriz , F. 2009b . New Lower Kimmeridgian ammonites (Ataxioceratinae) from the eastern Iberian Chain, E–NE Spain. Systematic, biogeographic and biostratigraphic relevance . Acta Palaeontologica Polonica , [in litt.]
- Olóriz, F., 1978: Kimmeridgiense-Tithónico inferior en el Sector Central de las Cordilleras Béticas (Zona Subbética). Paleontología. Bioestratigrafía. Ph.D. Thesis No. 184, Universidad de Granada, Granada, Spain. 758 pp
- Olóriz , F. 1997 . “ Interpretaciones ecoestratigráficas. Aplicaciones a los materiales del Jurásico superior ” . In Vida y ambientes del Jurásico , Edited by: Gámez Vintaned , J.A. and Linán , E. 33 – 57 . Zaragoza : Institución Fernando el Católico .
- Olóriz , F. 2000 . Time-Averaging and long-term palaeoecology in macroinvertebrate assemblages with ammonites (Upper Jurassic) . Revue de Paleobiologie, Vólume Spécial , 8 : 123 – 140 .
- Olóriz , F. , Rodríguez-Tovar , F.J. and Schairer , G. 1992 . New record of Barthelia subbetica Olóriz & Schairer (Jurassic Ammonitina) from the South Iberian paleomargin (Prebetic Zone, Spain) . Neues Jahrbuch für Geologie und Paläontologie, Monatshefte , 6 : 343 – 350 .
- Oppel , A. 1863 . Palaeontologische Mittheilungen – III. Ueber jurassische Cephalopoden . Palaeontologische Mittheilungen aus dem museum des Königlichen Bayerische Staates, Stuttgart , 1 : 163 – 266 .
- Pavia , G. , Benetti , A. and Minetti , C. 1987 . Il Rosso Ammonitico dei Monti Lessini Veronesi (Italia NE). Faune ad Ammoniti e discontinuità stratigrafiche nel Kimmeridgiano inferiore . Bolletino della Societá Paleontologica Italiana , 26 ( 1–2 ) : 63 – 92 .
- Sapunov , I.G. 1977 . Ammonite Stratigraphy of the Upper Jurassic in Bulgaria. III. Kimmeridgian, Substages, Zones and Subzones . Geologica Balcanica , 7 : 63 – 80 .
- Sarti , C. 1993 . “ Il Kimmeridgiano delle Prealpi Veneto–Trentine: Fauna e Biostratigrafia ” . In Memorie del Museo Civico di Storia Naturale di Verona (IIa serie), Sezione Scienze della Terra Vol. 5 , 144