
ABSTRACT
The distinctive, tall and slender cyrtoconic shell of the gastropod Pollicina cyathina Koken, 1897, originally established on a poorly preserved internal mould from the Middle Ordovician of the Dalarna region of central Sweden, is redescribed on the basis of additional material from several localities in the vicinity of Rättvik. Repaired injuries preserved in the fine comarginal growth lines of the archinacelloidean gastropod indicate failed durophagous predatory attacks. Unusual apical structures on an internal mould from Sjurberg probably result from closure of the earliest growth stages later in ontogeny.
The slowly expanding, slightly curved and bilaterally symmetrical mollusc Pollicina derives its name from the Latin “pollex” (genitive “pollicis”) meaning thumb and was proposed by Koken in Holzapfel (Citation1895) for an Ordovician species originally described from the Ordovician of the St Petersburg region of Russia: Pollicina corniculum (Eichwald Citation1860). Rare specimens of the upright fossil are known from Ordovician limestones in the Siljan district, Dalarna (), where this “Dala thumb” was given the name Pollicina cyathina by Koken (Citation1897, see also Koken & Perner Citation1925). The species epithet is probably a reflection of the similarity of the inverted shell form to the deep ladle or cup (Latin, “cyathus”) used as a measure to dispense wine in the ancient world. Pollicina has always been a troublesome mollusc. Originally considered to be a cephalopod (Eichwald Citation1842), it has been interpreted subsequently as a hyolith or pteropod (Eichwald Citation1860), a capuliform gastropod (Koken Citation1897; Koken & Perner Citation1925), a scaphopod-like mollusc (Starobogatov Citation1974; Kisselev Citation1994), a tergomyan monoplacophoran (Evans & Cope Citation2003) and a euomphaliform gastropod (Yochelson & Webers Citation2006). After redescribing the type species from Russia and related material from Estonia, Peel (Citation2020) recognised the similarity to some tergomyans but assigned Pollicina to the archinacelloidean gastropods, a group of Palaeozoic patelliform gastropods, but not patellogastropods as defined by Lindberg (Citation1986, Citation1988, Citation2008).
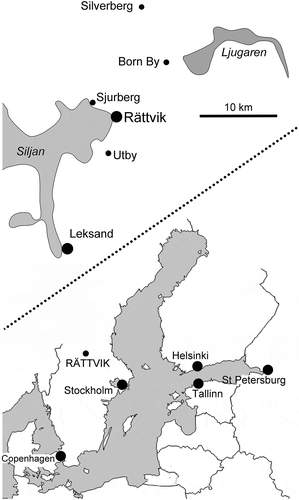
Figure 1. Localities for Pollicina cyathina Koken, Citation1897 in the Rättvik area of Dalarna, central Sweden.
Koken & Perner (Citation1925) recognised nine species of Pollicina from the Baltic Ordovician but only three of these are sufficiently well founded to be retained in the genus at this time. Pollicina corniculum (Eichwald Citation1860), the type species (), was originally described from the St Petersburg region of Russia (Koken Citation1897) but it is known also from the United Kingdom (Evans & Cope Citation2003). Ursula Toom (written communication 13 January 2020) has brought to my attention a well preserved specimen of Pollicina corniculum from the Kunda Stage at Vihula (Vihula I outcrop) in northern Estonia. The specimen (GIT 693–4) is essentially complete, although the shell has been exfoliated in the latest growth stage. It is well illustrated on the Geoscience Collections of Estonia website (http://geocollections.info/specimen/161183).
Figure 2. Pollicina from the Ordovician of Estonia and Russia. A. C–E. Pollicina crassistesta Koken, Citation1897, Ordovician, Darriwilian Series, Kunda Stage, Estonia. A, C. ELM g8:23 [1208g8/23], lectotype, largely exfoliated specimen in lateral view (A) with detail of ornamentation (C), original of Koken & Perner (Citation1925, pl. 38, fig. 9), Domberg (= Toompea). D, E. ELM g8:7 [1208g8/7], paralectotype, internal mould in lateral (D) and sub-apical (E) views, Springthal (= Tondi), original of Koken & Perner (Citation1925, pl. 38, figs. 1, 2). B, F. Pollicina corniculum Koken in Holzapfel, Citation1895, Ordovician, Darriwilian Series, Kunda Stage, Pulkova [Pulkowa], St Petersburg. B. PSM 4/22, lectotype in lateral view. F. CNIGRM 15702 (200/10903), lateral view with deep scratches resulting from preparation on right side. Scale bars: 5 mm.
![Figure 2. Pollicina from the Ordovician of Estonia and Russia. A. C–E. Pollicina crassistesta Koken, Citation1897, Ordovician, Darriwilian Series, Kunda Stage, Estonia. A, C. ELM g8:23 [1208g8/23], lectotype, largely exfoliated specimen in lateral view (A) with detail of ornamentation (C), original of Koken & Perner (Citation1925, pl. 38, fig. 9), Domberg (= Toompea). D, E. ELM g8:7 [1208g8/7], paralectotype, internal mould in lateral (D) and sub-apical (E) views, Springthal (= Tondi), original of Koken & Perner (Citation1925, pl. 38, figs. 1, 2). B, F. Pollicina corniculum Koken in Holzapfel, Citation1895, Ordovician, Darriwilian Series, Kunda Stage, Pulkova [Pulkowa], St Petersburg. B. PSM 4/22, lectotype in lateral view. F. CNIGRM 15702 (200/10903), lateral view with deep scratches resulting from preparation on right side. Scale bars: 5 mm.](/cms/asset/0e672dd9-67ae-40ce-bfe2-1a12898f4f2f/sgff_a_1728374_f0002_b.gif)
Pollicina crassistesta Koken, Citation1897 is an Estonian species () and together with P. corniculum it was recently redescribed by Peel (Citation2020).
Pollicina cyathina Koken, Citation1897 from the Ordovician of Dalarna, central Sweden, is redescribed in the present paper on the basis of the holotype, a poorly preserved internal mould (), and supplementary material showing shell ornamentation () together with unusual features of the shell interior (). Repaired injuries preserved in growth ornamentation () indicate that P. cyathina was subject to durophagous predatory attacks which the specimen in question survived.
Figure 3. Pollicina from the Ordovician of Sweden and Estonia. A–D, G, H. Pollicina cyathina Koken, Citation1897, A–D. PMU 35717 from Rättvik, Dalarna. A. close up sub-apical surface showing fine comarginal growth lines on adaperturally tapering growth lamellae disturbed by penetrative fractures (arrows 1–3) repaired during life. B. view of sub-apical surface showing broken apex. C. detail of broken apex showing circular cross-section and thick shell. D. lateral view. G, H. SMNH Mo 7767, holotype by monotypy, original specimen of Koken & Perner (Citation1925, pl. 38, figs. 6,7). Born [Borns] By, Rättvik, Dalarna. Lower Grey Orthoceras Limestone (Holen Limestone, Darriwilian Series, Kunda Stage; Ebbestad & Högström Citation2007). E, F. Pollicina crassistesta Koken, Citation1897, ELM 1201: g1:2664, Tallinn, Ordovician, Darriwilian Series, Kunda Stage, Estonia. A. slightly oblique view from perspective of sub-apical surface showing laterally flattened internal mould overlain by extremely thick shell (arrows mark lateral margin of shell). F. lateral view of internal mould and enveloping thick shell (arrows mark margin of shell on supra-apical surface). Scale bars: A, C (2 mm), B, D (4 mm), E, F (1 mm), G, H (5 mm).
![Figure 3. Pollicina from the Ordovician of Sweden and Estonia. A–D, G, H. Pollicina cyathina Koken, Citation1897, A–D. PMU 35717 from Rättvik, Dalarna. A. close up sub-apical surface showing fine comarginal growth lines on adaperturally tapering growth lamellae disturbed by penetrative fractures (arrows 1–3) repaired during life. B. view of sub-apical surface showing broken apex. C. detail of broken apex showing circular cross-section and thick shell. D. lateral view. G, H. SMNH Mo 7767, holotype by monotypy, original specimen of Koken & Perner (Citation1925, pl. 38, figs. 6,7). Born [Borns] By, Rättvik, Dalarna. Lower Grey Orthoceras Limestone (Holen Limestone, Darriwilian Series, Kunda Stage; Ebbestad & Högström Citation2007). E, F. Pollicina crassistesta Koken, Citation1897, ELM 1201: g1:2664, Tallinn, Ordovician, Darriwilian Series, Kunda Stage, Estonia. A. slightly oblique view from perspective of sub-apical surface showing laterally flattened internal mould overlain by extremely thick shell (arrows mark lateral margin of shell). F. lateral view of internal mould and enveloping thick shell (arrows mark margin of shell on supra-apical surface). Scale bars: A, C (2 mm), B, D (4 mm), E, F (1 mm), G, H (5 mm).](/cms/asset/dede96a9-92cc-43e4-9937-471ceda1f214/sgff_a_1728374_f0003_b.gif)
Figure 4. Pollicina cyathina Koken, Citation1897 from Dalarna, Sweden, Ordovician, Darriwilian Series, Kunda Stage, Lower Grey Orthoceras Limestone (Holen Limestone; Ebbestad & Högström Citation2007). A–C, G, H. LO 12 441t, internal mould, Utby, Dalarna. A, B. views of smooth nodose tip. C. dorsal view of supra-apical surface showing pointed apex. G. lateral view showing pointed apex. H. detail of raised tip in lateral view. D–F, I. LO 12 442t, internal mould, Sjurberg. D. sub-apical surface showing apical node and periodic slight constrictions (arrows) of lateral profile during growth. E. supra-apical surface. F. lateral view. I. oblique lateral view of apex. Scale bars: A, B, H (1 mm), C (2mm), D–G, I (5 mm).

Systematic Palaeontology
Repositories
– CNIGRM, F.N. Chernyshev Central Geological Survey Research Museum, St Petersburg. ELM, Estonian Natural History Museum, Tallinn. GIT, Geological collections at Tallinn University of Technology. LO, palaeontological type collection, Department of Geology, Lund University. PMU, palaeontological type collections of the Museum of Evolution, Uppsala University. PSM, Museum of the Department of Historical Geology, St Petersburg State University. SMNH, Swedish Museum of Natural History, Stockholm.
Family Pollicinidae Perner in Koken & Perner, Citation1925
Genus Pollicina Koken in Holzapfel, Citation1895
Type species
– Cyrtolithes corniculum Eichwald, Citation1860 from the Darriwilian Series, Kunda Stage of the St Petersburg region, Russia.
Discussion
– Peel (Citation2020) presented an emended diagnosis and described the type species on the basis of type and other material from Pulkova, St Petersburg, Russia, of Darriwilian (Kunda Stage) age. Specimens of Pollicina corniculum from Russia usually retain the shell (). In contrast, almost all specimens of P. crassitesta Koken, Citation1897 from the Kunda Stage of Estonia are preserved as internal moulds (Peel Citation2020; ), although the lectotype retains patches of shell with ornamentation (). Only one of ten examined specimens of Pollicina from Dalarna retains shell (), but the four specimens described herein permit a redescription of P. cyathina. Specimens of other species are rare. Pollicina conoidea Koken in Koken & Perner, Citation1925 was transferred to Pygmaeoconus conoideus by Yochelson (Citation1977). Peel (Citation2020) excluded P. ampliata Koken in Koken & Perner, Citation1925 from Pollicina, while P. acuta and P. brevis are too poorly known for placement (Perner in Koken & Perner Citation1925; Peel Citation2020).
Scale measurements in the caption accompanying illustrations of the lectotype of P. corniculum by Kisselev (Citation1994, fig. 1) indicated that the specimen is only 9 mm tall, less than half its true height, although other measurements in Kisselev’s (Citation1994) Russian text appear to be correct. The incorrect specimen height was inadvertently restated by Evans & Cope (Citation2003, p. 143).
Pollicina cyathina Koken, Citation1897 ;
1897 Pollicina cyathina Koken, p. 198–199, fig. 38
1925 Pollicina cyathina; Koken & Perner, p. 227, pl. 38, figs. 6, 7
Holotype
– SMNH Mo 7767, the specimen figured in simple sketches by Koken (Citation1897, fig. 38) and stated to be from Clason. Koken & Perner (Citation1925, pl. 38, figs. 6, 7; ) illustrated a single specimen from the collections of the Swedish Natural History Museum, Stockholm, which is compatible with these sketches and also stated to be from Clason. This name is scrawled across the specimen label but most probably indicates the collector, I.G. Clason (1856–1930) of Rättvik. The specimen is from Born By [Borns By], about 10 km north-east of Rättvik, Dalarna () and was collected during 1892 from the Lower Grey Orthoceras Limestone (Holen Limestone, Darriwilian Series, Kunda Stage; Ebbestad & Högström Citation2007).
Figured material
– LO 12 441t, internal mould, Utby, Dalarna; LO 12 442t, internal mould, Sjurberg, Dalarna. Lower Grey Orthoceras Limestone (Holen Limestone, Darriwilian Series, Kunda Stage; Ebbestad & Högström Citation2007). PMU 35717 from Rättvik, Dalarna.
Other material
– Incomplete internal moulds from Silverberg and Utby () in the collections of the Swedish Natural History Museum, Stockholm.
Diagnosis
– Slender species of Pollicina with circular cross-section and ornamentation of fine comarginal growth lines.
Description
– The slowly expanding, bilaterally symmetrical, cyrtoconic shell curves through about 60 degrees, such that the apex of the tall shell overhangs the sub-apical margin. In lateral view (; ) the maximum dimension of the aperture is about two fifths of the total height. The cross-section is almost circular (). The apertural margin is orthocline and the internal mould periodically may be slightly constricted near the aperture (). The shell is thick near the apex () but thins to an acute margin, feather edge, at the aperture (). Ornamentation consists of fine growth lines (). Internal moulds are generally smooth (; ), without indications of transverse rugae, although a comarginal raised band of muscle scars (Peel Citation2020) may be prominent at about half to two thirds of the distance from the apex to the aperture in some of the unfigured material.
Discussion
– The lectotype is a large but poorly preserved internal mould with the apex broken away, although it appears rounded in lateral view in the illustrations due to weathering (). It has the same curvature and circular cross-section as a broken specimen from Rättvik, which retains the shell with its ornamentation of fine comarginal growth lines (). The broken apex demonstrates the almost circular cross-section of the shell interior even in early growth stages and stands in contrast to the strongly laterally compressed apex of P. crassitesta (). However, the shell wall in the latter (indicated by arrows in ) is substantially thicker than in P. cyathina.
Koken (Citation1897, p. 199) described but did not illustrate Pollicina acuta Koken, Citation1897 from the Lower Grey Orthoceras Limestone of Dalarna (Holen Limestone, Darriwilian Series, Kunda Stage; Ebbestad & Högström Citation2007). Koken & Perner (Citation1925, p. 228) noted that P. acuta Koken, Citation1897 was based on material from Dalarna, but Perner in Koken & Perner (Citation1925) commented that the specimen before him did not agree with the original description and the name is not employed. The cross-section of P. acuta was stated to be elliptical even angulated, in the plane of symmetry while that of P. cyathina is circular. Both species were reported from Sjurberg ().
Pollicina corniculum usually has a more rapidly expanding shell than P. cyathina and lamellose growth lines (). Pollicina crassitesta was illustrated by Koken (Citation1897, p. 198) with a schematic drawing based on a specimen (Koken & Perner Citation1925, pl. 38, fig. 9) designated as the lectotype by Peel (Citation2020; ). Internal moulds of P. crassitesta are similar in lateral profile to P. cyathina, although the latter may be more slender, but they are more rapidly expanding when viewed in the plane of symmetry (compare with ). Pollicina crassitesta differs from P. cyathina in having an oval cross-section. In the earliest growth stages, the lateral surfaces of the internal mould in P. crassitesta are strongly flattened such that the length of the whorl cross-section is about twice its width (), whereas the cross-section is almost circular in P. cyathina (). The ornamentation of P. crassistesta has spiral lirae not seen in P. cyathina.
Shell growth in Pollicina
Most specimens assigned to Pollicina by Koken (Citation1897) and Koken & Perner (Citation1925) are internal moulds without details of ornamentation and great care is required in making deductions based on such material. However, the shell exterior surface with ornamentation is usually preserved in the type species, Pollicina corniculum (). The lectotype of Pollicina crassitesta () and one specimen of P. cyathina described herein () are rare specimens of their respective species which also retain shell, but information concerning shell at the apex is lacking in both.
The apex of Pollicina corniculum is bluntly rounded without discernible changes in the nature of the ornamentation (). Evidence of septation or closure of the early parts of the shell is not known in 3-dimensionally preserved material of P. corniculum from East Baltica although Evans & Cope (Citation2003) noted that an internal plug or septum was present in some, but not all, of available crushed specimens from the United Kingdom.
In general, the apex of Pollicina crassitesta is known only from internal moulds that show the strong laterally compressed form characteristic of the species (Peel Citation2020; ). However, where patches of shell material are preserved, it is evident that there has been very substantial thickening of the shell at the apex ( where arrows indicate the outer surface of the shell) leading to significant reduction in the volume of the shell interior at the apex. The lack of constrictions or discontinuities in the profile of the internal moulds indicates that this secondary thickening in the apical area passed imperceptibly into the normal shell growth as the aperture was approached. Thus, shell thickness at the apex may be the same as that around the aperture. A similar situation is evident in P. cyathina where the shell near the apex () is almost as thick as that in the latest preserved growth stage, but not as massively thickened as in P. crassitesta. At this latest stage, however, the shell in P. cyathina is clearly seen to have been built up from successive feather-edged lamellae ().
Two internal moulds from the Törnquist Collection (Lund University), here assigned to Pollicina cyathina, are preserved in black limestone and their smooth glossy surface retains no trace of adherent shell material (). Periodicity in growth of the largest specimen, possibly of a seasonal nature, is suggested by shallow constrictions in the lateral margins in sub-apical view (, arrows). The apex in this specimen is uniformly and smoothly rounded with a weakly developed terminal node (). In contrast, the internal mould of the smaller specimen narrows abruptly to a distinct node at the apex (). In detail (), this apical area is delimited in part from the later growth area by its smooth texture and a narrow comarginal groove (). The apical node is transversely elongate and its upper surface is flattened, with acute edges suggestive of breakage.
The delimitation of the distinct apical portion of the internal mould may have several causes. It might represent the internal core of a small protoconch but it is not known how this was reflected, if at all, in the external morphology of the apex. A structure similar to the apical node is preserved on an internal mould of the tryblidioid tergomyan Pilina cheyennica Peel, Citation1977 from the Late Ordovician of Oklahoma, U.S.A. (Peel Citation1977, text-fig. 2A, B). A corresponding pit on the shell interior was illustrated by Lindström (Citation1884, pl. 1, fig. 36) in Pilina unguis (Lindström in Angelin & Lindström Citation1880) and by Angelin & Lindström (Citation1880, pl. 10, fig. 9) in Tryblidium reticulatum Lindström in Angelin & Lindström, Citation1880 from the Silurian of Gotland, Sweden. A low conical species described from Gotland by (Lindström Citation1884, pl. 19, fig. 3) as Palaeacmaea (?) solarium Lindström, Citation1884 also preserves a distinct apical node on the internal mould; it is not a species of Palaeacmaea Hall & Whitfield, Citation1872 originally described from the Cambrian (Furongian) of the U.S.A. (Webers & Yochelson Citation1999). Recognition of a protoconch could be extended to also include the initial, smooth, conical section of the internal mould from which the apical node arises (), but both interpretations fall if there was substantial ontogenetic thickening of the shell at the apex, as in P. crassitesta.
A more fanciful interpretation is that the node could represent a trace of the infilling of a siphuncle and that Pollicina is therefore a cephalopod. But there is scarcely room for camerae in the apical portion of the shell and this interpretation is dismissed. Kisselev (Citation1994; see discussion in Peel Citation2020) did not record septa in the early sectioned portion of the lectotype of Pollicina corniculum, although they are present in some specimens figured by Evans & Cope (Citation2003). Septa are of common occurrence in slowly expanding Palaeozoic gastropod shells (Yochelson Citation1971; Linsley Citation1978; Cook Citation1994; Peel Citation2019) and usually result from the closing off of narrow, unwanted, early growth stages as growth progresses. Internal thickening of the apical shell or the deposition of internal apical plugs may have a similar function, but can also serve to close off corroded early growth stages, or to strengthen weak, originally thin-shelled, early growth stages. The last case is supported by the similarity in shell thickness of the strengthened apex and the adult shell ().
As regards cephalopods, however, it is the presence of a siphuncle penetrating the septa that defines a cephalopod and not the presence of septa alone (Teichert Citation1988; Peel Citation1991). Yochelson et al. (Citation1973) proposed that the cephalopod siphuncle was developed from a strand of tissue that remained attached to the shell apex as the mollusc body mass migrated in its narrow shell towards the aperture with growth, with septa forming astride this remnant tissue. Adherents of this theory may find support in the apical node of Pollicina cyathina but this is a Middle Ordovician taxon, which bears little similarity in detail to the Cambrian (Furongian) ancestor of cephalopods proposed by Yochelson et al. (Citation1973) and Webers & Yochelson (Citation1989). A morphological comparison of the two was given by Peel (Citation2020).
At the present day, the minute (1–2 mm) gastropod Caecum Fleming, Citation1813 has a tubular, slightly curved shell that is not disimilar in curvature to Pollicina, but is parallel-sided (Bandel Citation1996). Its protoconch is usually trochospirally coiled and is decollated during ontogeny from the later, tubular, teleoconch at a septum that closes the apical extremity of the adult shell (de Folin Citation1868; Bandel Citation1996; Pizzini et al. Citation1998; Pastorino & Chiesa Citation2014). The septum is usually convex and often develops an adapically directed protruberance, termed the mucro, well illustrated by Pastorino & Chiesa (Citation2014, figs. 7,8). Their illustrations show a transversely elongate mucro, with a flattened upper surface with angular edges that is morphologically similar to the apical node in P. cyathina (). Pizzini et al. (Citation1998, figs. 12–19) demonstrated that the septation and decollation process in Caecum involved the formation of two septa. An initial temporary convex septum is followed adaperturally by a second, mucronate, septum. Thus, the mucro is occurs in the space between the two septa and may have contact with the temporary septum. The mucronate septum eventually forms the permanent septum following loss of the adapical temporary septum. While any taxonomic relationship between Pollicina and Caecum is obviously discounted, the morphological similarity of the mucro-bearing septa may indicate comparable mechanisms for the closing off of unwanted earlier portions of the shell. Septum formation in Caecum is followed by decollation of the protoconch, while in Pollicina cyathina it precedes closure of the apex by a septum or plug.
Peel (Citation2020) concluded that the frequent displacement of the lamellose growing margin in Pollicina corniculum indicated a limpet-like mode of life, although such a tall shell form is not found in limpets at the present day (Vermeij Citation2017). Comparison can be made with the frequent development of tall, but irregular, clamping shells of Orthonychia Hall, Citation1843 amongst the anisostrophic platyceratoidean gastropods in the Silurian–Carboniferous (Perner Citation1903; Frýda et al. Citation2008, Citation2009; Peel Citation2020), although no taxonomic relationship is implied. The uniformly thick shell of P. crassitesta might support the limpet interpretation, suggesting life in a higher energy environment than P. corniculum, but the extreme height of the shell does not suggest stability under such conditions. A simple protective function seems inadequate in P. cyathina where the apex is relatively thick shelled () but the growing edge of the aperture is a feather-edge lamella susceptible to breakage (). However, such an edge functions equally well as a seal against the substrate and may just indicate that soft parts never extended beyond the apertural margin (Peel Citation2020).
Many Cambrian (Furongian)–Early Ordovician molluscs develop tall shells (Stinchcomb & Echols Citation1966; Stinchcomb Citation1980, Citation1986; Stinchcomb & Angeli Citation2002; Webers & Yochelson Citation1989; Webers et al. Citation1992; Yochelson & Webers Citation2006) and Stinchcomb (Citation1975) suggested association of some of these with sheltered sites within digitate stromatolites. At the present day, siliquariid gastropods live within sponges (Savazzi Citation1996; Pansini et al. Citation1999) but their irregularly uncoiled anisostrophic shells are dissimilar to the bilaterally symmetrical cones of Pollicina.
Shell breakage and repair
Numerous irregularities and dislocations were noted by Peel (Citation2020) in the growth lines of Pollicina corniculum () but penetrative fractures of the margin or shell repairs were not observed. In contrast, the single available specimen of Pollicina cyathina with preserved shell shows a series of repaired injuries on the sub-apical surface (). The regular pattern of fine growth lines is largely undisturbed on other surfaces with the exception of minor chipping (, arrow). The apex of the specimen is missing, almost certainly broken away during collection: the rest of the specimen has been re-assembled from two broken pieces, The aperture is also broken, but there is some evidence that the sub-apical margin was broken prior to burial.
The fractures on the sub-apical surface are penetrative embayments into the apertural margin that have left scars with raised, scarp-like edges at their deepest, adapical, point. This relief is quickly lost laterally where traces of the breakage events are absent. Three major scars are visible (, arrows 1–3), each infilled by subsequent shell growth. In each scar the thin growth lamella of the shell is seen to taper to an acute margin, which produces just a normal, fine growth line on the adjacent lateral surface. Growth lines in the shell filling the fractures are initially irregular but gradually return to their normal transverse pattern as the embayment is filled. Repaired injuries of this kind are well known in Palaeozoic molluscs (Vermeij et al. Citation1981; Peel Citation1984, Citation2015; Vermeij Citation1987, Citation2013; Ebbestad & Peel Citation1997; Horný Citation1997; Ebbestad Citation1998; Alexander & Dietl Citation2003; Lindström & Peel Citation2005; Ebbestad et al. Citation2009; Stafford et al. Citation2015) and may result from physical as well as biological agencies (Peel Citation2020).
On account of their depth, form and location, the three scars described here in Pollicina cyathina are readily interpreted as the result of non-lethal predatory attacks that have been repaired by subsequent shell growth (). However, discrimination between physical and biological breakage is rarely clear cut (Stafford et al. Citation2015). The scars are unusual in that they form a series on the sub-apical surface of the shell while evidence of attacks is absent from other areas. It might be expected that if breakage was a result of physical agencies similar scars would be present on other parts of the feather-edge apertural margin, but this is not the case. Most of the remaining shell surface is characterised by very fine, undisturbed growth lines (). However, on the basis of a single specimen it is not profitable to assess if this pattern is coincidental or if there is an underlying cause. The location of predatory attacks is often stimulated by features of shell morphology in the prey, e.g., focus on the apertural emargination in bellerophontiform and pleurotomarioidean gastropods (Lindström Citation2003; Ebbestad et al. Citation2009), but could reflect features of the behaviour of either the prey or attacker, or a response to the reception of chemical signals from an exhalant current from the mantle cavity (Kohn Citation1961; Croll Citation1983; Sakata Citation1989; Villanueva et al. Citation2017).
Acknowledgments
Per Ahlberg and Mats Eriksson (Lund University), Mare Isakar (Tartu Museum, Estonia) and Christian Skovsted (Swedish Museum of Natural History) kindly arranged the loan of specimens in their care. Guido Pastorino (Museo Argentino de Ciencias Naturales) supplied information concerning Caecum. Jan Ove R. Ebbestad (Museum of Evolution, Uppsala University) provided information concerning I.G. Clason. Ulla Toom (Tallinn University of Technology) is thanked for recording Pollicina corniculum from Estonia. Gregory P. Dietl (Cornell University) and an anonymous reviewer are thanked for comments concerning the submitted manuscript.
Disclosure statement
No potential conflict of interest was reported by the author.
Additional information
Funding
References
- Alexander, R.R. & Dietl, G.P., 2003: The fossil record of shell-breaking predation on marine bivalves and gastropods. In P.H. Kelley, M. Kowalewski & T.A. Hansen (eds.): Predator-prey Interactions in the Fossil Record, 141‒176. Kluwer Academic/Plenum Publishers, New York.
- Angelin, N.P. & Lindström, G., 1880: Fragmenta silurica. Samson & Wallin, Stockholm. 39 pp.
- Bandel, K., 1996: Phylogeny of Caecidae (Caenogastropoda). Mitteilungen aus dem Geologisch-Paläontologischen Institut der Universität Hamburg 79, 53–115.
- Cook, A.G., 1994: A septate gastropod from the Silurian Bungonia Limestone, New South Wales. Memoirs of the Queensland Museum 35, 27–30.
- Croll, R.P., 1983: Gastropod chemoreception. Biological Reviews 58, 293–319. doi:10.1111/brv.1983.58.issue-2.
- de Folin, L., 1868: Observations on the septum of the Caecidae; and some remarks on the subject of the suppression of the genera Brochina and Strebloceras or Phleboceras. Zoology Journal of the Linnean Society of London 10, 254–264. doi:10.1111/j.1096-3642.1868.tb02235.x.
- Ebbestad, J.O.R., 1998: Multiple attempted predation in the Middle Ordovician gastropod Bucania gracillima. GFF 120, 27‒33. doi:10.1080/11035899801201027.
- Ebbestad, J.O.R. & Högström, A.E.S., 2007: Ordovician of the Siljan District, Sweden. Sveriges Geologiska Undersökning, Rapporter Och Meddelanden 128, 7–26.
- Ebbestad, J.O.R., Lindström, A. & Peel, J.S., 2009: Predation on bellerophontiform molluscs in the Palaeozoic. Lethaia 42, 469‒485. doi:10.1111/j.1502-3931.2009.00167.x.
- Ebbestad, J.O.R. & Peel, J.S., 1997: Attempted predation and shell repair in Middle and Upper Ordovician gastropods from Sweden. Journal of Paleontology 71, 1007–1019. doi:10.1017/S002233600003599X.
- Eichwald, E., 1842: Die Urwelt Russlands. Zweites Heft. Aus den beitragen zur Kenntnis des Russischen Reichs. Herausgegeben von K. E. Baer. Besonders Abgedrüct der Wissenschaften, St. Petersburg. 100 pp.
- Eichwald, E., 1860: Lethaea Rossica ou paléontologie de la Russié, décrite et figurée. Vol. 1, seconde sectione de l’ancienne période, 681–1657. E. Schweizbart, Stuttgart.
- Evans, D.H. & Cope, J.C.W., 2003: Systematic position of Pollicina corniculum (Eichwald, 1860) (Mollusca, Tergomya) from the Middle Ordovician of the United Kingdom. Palaeontology 46, 139–149. doi:10.1111/1475-4983.00291.
- Fleming, J., 1813: Conchology. In D. Brewster (ed.): The Edinburgh encyclopedia conducted by David Brewster with assistance of gentlemen eminent in science and literature Vol. 7, 55–107. William Blackwood, Edinburgh.
- Frýda, J., Racheboeuf, P.R. & Frýdová, B., 2008: Mode of life of Early Devonian Orthonychia protei (Neritimorpha, Gastropoda) inferred from its post-larval shell ontogeny and muscle scars. Bulletin of Geosciences 83, 491–502. doi:10.3140/bull.geosci.2008.04.491.
- Frýda, J., Racheboeuf, P.R., Frýdova, B., Ferrová, L., Mergl, M. & Beryková, S., 2009: Platyceratid gastropods – stem group or patellogastropods, neritomorphs or something else? Bulletin of Geosciences 84, 107–120. doi:10.3140/bull.geosci.1125.
- Hall, J., 1843: Geology of New York. Part IV, comprising the survey of the fourth geological district. Natural History of New York 4, 1–683.
- Hall, J. & Whitfield, R.P., 1872: Notice of two new species of fossil shells from the Potsdam sandstone of New York. Annual Report of the Regents of the University of the State of New York 23, 241–242.
- Holzapfel, E., 1895: Das Obere Mitteldevon (Schichten mit Stringocephalus Burtini und Maeneceras terebratum) im rheinischen Gebirge. Abhandlungen der Königlich Preußischen Geologischen Landesanstalt 16, 1–460.
- Horný, R.J., 1997: Shell breakage and repair in explanate bellerophontoidean gastropods from the Middle Ordovician of Bohemia. Věstník Českého Geologického Ústavu 72, 157–168.
- Kisselev, G.N., 1994: The systematic position of the Early Paleozoic mollusk “Cyrtolithes” corniculum (Eichwald, 1860) (Xenoconchia, Toxeumorphorida). Ruthenica 4, 35–37.
- Kohn, A.J., 1961: Chemoreception in gastropod molluscs. American Zoologist 1, 291–308. doi:10.1093/icb/1.2.291.
- Koken, E., 1897: Die Gastropoden des baltischen Untersilurs. Bulletin de l’Académie Impériale des Sciences de St Petersburg 7, 97–214.
- Koken, E. & Perner, J., 1925: Die Gastropoden des baltischen Untersilurs. Mémoires de l’Académie des Sciences de Russie, Série. 8, Classe Physico-mathématique 37, 1–326.
- Lindberg, D.R., 1986: Radular evolution in the Patellogastropoda. American Malacological Bulletin 4, 115.
- Lindberg, D.R., 1988: The Patellogastropoda. Malacological Review, Supplement 4, 35–63.
- Lindberg, D.R., 2008: Patellogastropoda, Neritomorpha, and Cocculinoidea. In W.F. Ponder & D.R. Lindberg (eds.): Phylogeny and Evolution of the Mollusca, 271–296. University of California Press, Berkeley.
- Lindström, A., 2003: Shell breakage in two pleurotomarioid gastropods from the Upper Carboniferous of Texas, and its relation to shell morphology. GFF 125, 39–46. doi:10.1080/11035890301251039.
- Lindström, A. & Peel, J.S., 2005: Repaired injuries and shell form in some Palaeozoic pleurotomarioid gastropods. Acta Palaeontologica Polonica 50, 697–704.
- Lindström, G., 1884: On the Silurian Gastropoda and Pteropoda of Gotland. Kongliga Svenska Vetenskaps-Akademiens Handlingar 19 (6), 1–250.
- Linsley, R.M., 1978: The Omphalocirridaae: a new family of Palaeozoic Gastropoda which exhibits sexual dimorphism. Memoirs of the National Museum of Victoria 39, 33–54. doi:10.24199/j.mmv.1978.39.03.
- Pansini, M., Cattanea Vietti, R. & Shiaporelli, S., 1999: Relationship between sponges and a taxon of obligatory inqilines: the siliquariid molluscs. Memoirs of the Queensland Museum 44, 427–437.
- Pastorino, G. & Chiesa, I.L., 2014: The family Caecidae (Gastropoda: caenogastropoda) in Argentine waters. The Nautilus 128, 40–50.
- Peel, J.S., 1977: Relationship and internal structure of a new Pilina (Monoplacophora) from the late Ordovician of Oklahoma. Journal of Paleontology 51, 116–122.
- Peel, J.S., 1984: Attempted predation and shell repair in Euomphalopterus (Gastropoda) from the Silurian of Gotland. Bulletin of the Geological Society of Denmark 32, 163–168.
- Peel, J.S., 1991: The Classes Tergomya and Helcionelloida, and early molluscan evolution. Bulletin Grønlands Geologiske Undersøgelse 161, 11–65.
- Peel, J.S., 2015: Operculum regeneration and failed predation in the silurian gastropod oriostoma. Palaeontology 58, 229–237. doi:10.1111/pala.12154.
- Peel, J.S., 2019: Muscle scars in euomphaline gastropods from the Ordovician of Baltica. Estonian Journal of Earth Sciences 68, 88–100. doi:10.3176/earth.2019.08.
- Peel, J.S., 2020: Muscle scars, mode of life and systematics of Pollicina (Mollusca) from the Ordovician of Baltica. Estonian Journal of Earth Sciences 69, 20–36.
- Perner, J., 1903: Systême Silurien du centre de la Bohême par Joachim Barrande. Vol. IV Gastéropodes 1, Texte (Patellidae et Bellerophontidae) et Planches 1 à 89. C., 1–164. Bellman, Prague.
- Pizzini, M., Oliverio, M. & Nofroni, I., 1998: Contribution to the knowledge of the family Caecidae. 4. The temporary septum formation of some caecid species (Caenogastropoda: rissooidea). Sociedad Española de Malacologia 16, 133–140.
- Sakata, K., 1989: Feeding attractants and stimulants for marine gastropods. Bioorganic Marine Chemistry 3, 115–129.
- Savazzi, E., 1996: Adaptations of vermetid and siliquariid gastropods. Palaeontology 39, 157–177.
- Stafford, E.S., Dietl, G.P., Murray, P., Gingras, P. & Leighton, L.R., 2015: Caedichnus, a new ichnogenus representing predatory attack on the gastropod shell aperture. Ichnos 22, 87–102. doi:10.1080/10420940.2015.1031899.
- Starobogatov, Y., 1974: Xenoconchias and their bearing on the phylogeny and systematics of some molluscan classes. Paleontological Zhurnal 1974, 1–13.
- Stinchcomb, B.L., 1975: Paleoecology of two new species of Late Cambrian Hypseloconus (Monoplacophora) from Missouri. Journal of Paleontology 49, 416–421.
- Stinchcomb, B.L., 1980: New information on Late Cambrian Monoplacophora Hypseloconus and Shelbyoceras (Mollusca). Journal of Paleontology 54, 45–49.
- Stinchcomb, B.L., 1986: New Monoplacophora (Mollusca) from Late Cambrian and Early Ordovician of Missouri. Journal of Paleontology 60, 606–626. doi:10.1017/S0022336000022149.
- Stinchcomb, B.L. & Angeli, N.A., 2002: New Cambrian and Lower Ordovician monoplacophorans from the Ozark Uplift, Missouri. Journal of Paleontology 76, 965–974. doi:10.1017/S0022336000057802.
- Stinchcomb, B.L. & Echols, D.J., 1966: Missouri Upper Cambrian Monoplacophora previously considered cephalopods. Journal of Paleontology 40, 647–650.
- Teichert, C., 1988: Main features of cephalopod evolution. In M.R. Clarke & E.R. Trueman (eds.): The Mollusca 12, palaeontology and neontology of cephalopods, 11–75. Academic Press, Orlando.
- Vermeij, G.J., 1987: Evolution & Escalation. An ecological history of life, 1–527. Princeton University Press, Princeton, NJ.
- Vermeij, G.J., 2013: On escalation. Annual Review of Earth and Planetary Sciences 41, 1–19. doi:10.1146/annurev-earth-050212-124123.
- Vermeij, G.J., 2017: The limpet form in gastropods: evolution, distribution, and implications for the comparative study of history. Biological Journal of the Linnean Society 120, 22–37.
- Vermeij, G.J., Schindel, D.E. & Zipser, E., 1981: Predation through geological time: evidence from gastropod shell repair. Science 214, 1024–1026. doi:10.1126/science.214.4524.1024.
- Villanueva, R., Perricone, V. & Fiorito, G., 2017: Cephalopods as predators: a short journey among behavioural flexibilites, adaptations, and feeding habits. Frontiers in Physiology 8. 17 August 2017. doi:10.3389/fphys.2017.00598.
- Webers, G.F., Pojeta, J., Jr. & Yochelson, E.L., 1992: Cambrian Mollusca from the Minaret Formation, Ellsworth Mountains, West Antarctica. Geological Society of America Memoir 170, 181–248.
- Webers, G.F. & Yochelson, E.L., 1989: Late Cambrian molluscan faunas and the origin of the Cephalopoda. Geological Society, London, Special Publications 47, 29–42. doi:10.1144/GSL.SP.1989.047.01.04.
- Webers, G.F. & Yochelson, E.L., 1999: A revision of Palaeacmaea (Upper Cambrian) (?Cnidaria). Journal of Paleontology 73, 598–607. doi:10.1017/S002233600003242X.
- Yochelson, E.L., 1971: A new Late Devonian gastropod and its bearing on problems of open coiling and septation. Smithsonian Contributions to Paleobiology 3, 231–241.
- Yochelson, E.L., 1977: Monoplacophora (Mollusca) from the Upper Ordovician of Norway. Norsk Geologisk Tidsskrift 47, 297–312.
- Yochelson, E.L., Flower, R.H. & Webers, G.F., 1973: The bearing of the new Late Cambrian monoplacophoran genus Knightoconus upon the origin of the Cephalopoda. Lethaia 6, 275–310. doi:10.1111/let.1973.6.issue-3.
- Yochelson, E.L. & Webers, G.F., 2006: A restudy of the Late Cambrian molluscan fauna of Berkey (1898) from Taylor Falls, Minnesota. Minnesota Geological Survey Report of Investigations 64, 1–66.