
ABSTRACT
Well-preserved holdfasts and early growth stages of the tubular cnidarian Sphenothallus are described from the early Silurian (Llandovery Series) of Washington Land, North Greenland. The diameter of the holdfast is determined by the formation of a basal disc attached to the substrate, upon which the initially conical, but subsequently tubular shell is formed. The characteristic opposing longitudinal thickenings are developed at each angular margin as the initially circular tube acquires an elliptical form.
Palaeozoic strata have yielded a number of carbonaceous and organophosphatic tubes, although their systematic position is often equivocal on account of their morphologically simple form. Sphenothallus Hall, Citation1847 is probably the most widely distributed, with numerous-cited occurrences from Cambrian to Permian (Zhu et al. Citation2000; Li et al. Citation2004; Muscente & Xiao Citation2015; Dzik et al. Citation2017; Landing et al. Citation2018; Vinn & Mironenko Citation2021). Hall (Citation1847) described Sphenothallus, with type species Sphenothallus angustifolius Hall, Citation1847 from the middle Ordovician of New York and considered it to be a plant. Others have considered it to be a worm (Mason & Yochelson Citation1985; Fauchald et al. Citation1986; Fauchald & Yochelson Citation1990) but the current consensus, summarized by Vinn & Kirsimäe (Citation2015), Dzik et al. (Citation2017) and Landing et al. (Citation2018), is that Sphenothallus is a cnidarian. The entangled nomenclative history of Sphenothallus was reviewed in detail by Mason & Yochelson (Citation1985), Zhu et al. (Citation2000) and Landing et al. (Citation2018).
The main morphological features of Sphenothallus are the long slender phosphatic tube, originating from a small conical holdfast, and an opposing pair of lateral longitudinal thickenings of the elliptical tube wall (Van Iten et al. Citation1992, ; Wang et al. Citation2003, fig. 8). As preserved, the described specimens vary considerably in length, from a few millimetres (Dzik et al. Citation2017, ) to more than 12 cm (Dzik et al. Citation2017, ; Mason & Yochelson Citation1985; Wang et al. Citation2003).
Figure 1. Locality information for GGU sample 216842 Cape Schuchert Formation (Silurian, Llandovery Series), Washington Land, North Greenland.
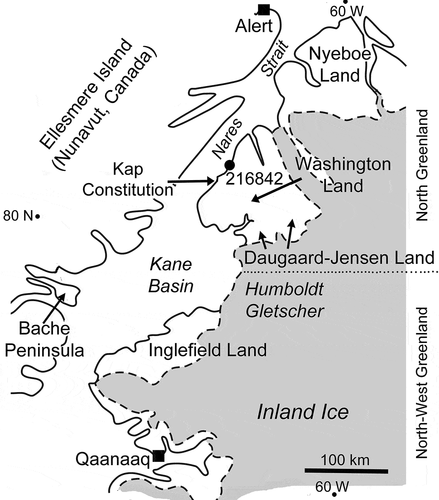
Figure 2. Holdfasts of Sphenothallus sp., GGU sample 216842 Cape Schuchert Formation (Silurian, Llandovery Series), Washington Land, North Greenland. A–C. PMU 38328/1, basal view of the holdfast cone with margin of basal attachment disc preserved at periphery (A and inset). D,E. PMU 38328/2, basal view of holdfast cone (E) showing dense, thin, outer layer and amorphous inner layer; detail in (D). F,G. PMU 38328/3, lateral views showing growth displacement (arrows). H,I. PMU 38328/4, holdfast in plan and oblique views. J. PMU 38328/5, oblique lateral view showing growth lamellae (arrow). K. PMU 38328/6, lateral view. L. PMU 38328/7, lateral view. M. PMU 38328/8, lateral view with irregular holdfast. N. PMU 38328/9, broken tube showing thickening of elliptical shell in apertural view. O. PMU 38328/10, oblique view, showing thickening of shell wall. P. PMU 38328/11, lateral view showing preferential breakage of lateral area. Scale bars: 20 µm (A); 50 µm (D,E); 100 µm (B,C,I,J,O); 200 µm (F–H,K–N,P).

Figure 3. Sphenothallus sp., GGU sample 216842, Cape Schuchert Formation (Silurian, Llandovery Series), Washington Land, North Greenland. A,B PMU 38328/11, growth lamellae, with A a detail of B, the latter showing decrease in thickness of lamella from arrow a to arrow b. C,F,I. PMU 38328/12, channel-like fragment of longitudinal thickening showing growth lamellae. D. PMU 38328/10, apertural view of specimen illustrated as Fig. 2O showing thickening of shell wall and comarginal growth lamellae. E. PMU 38328/13, fragment of tube showing delimitation of longitudinal thickenings by groove (aperture to left). G,H. PMU 38328/14, inner surface of basal attachment disc with down-stepping growth lamellae (arrow in H). Scale bars: 50 µ m (A,C,D,H); 100 µm (B,G,I); 200 µm (E,F).

This paper describes holdfasts and the early growth stages of Sphenothallus specimens etched from Silurian limestones of the Cape Schuchert Formation (Silurian, Llandovery), Washington Land, North Greenland (). While providing additional information on the nature of the attachment structures in Sphenothallus, the lack of later growth stages due to apertural breakage inhibits assignment of the tiny (3 mm) specimens to species, although the characteristic lateral thickenings of the tube are well developed.
Background
The Cape Schuchert Formation of Washington Land was proposed by Koch (Citation1929) and its fauna described by Poulsen (Citation1934). Silurian stratigraphy of the Washington Land area was described by Hurst (Citation1980) and Hurst & Surlyk (Citation1982), while Higgins et al. (Citation1991) gave an integrated overview of the development of the Lower Palaeozoic transarctic Franklinian Basin succession across North Greenland. A geological map and description was published by Dawes (Citation2004).
Material
GGU 216842 was collected from the Cape Schuchert Formation by J.M. Hurst (GGU) in July 1976 from just south of Kap Schuchert, Washington Land (80°45ꞌN, 65°15ꞌW; ). A geological section showing the stratigraphic derivation of the sample was given by Armstrong (Citation1990, fig. 21); Silurian, early Llandovery Series, early Aeronian Stage.
The black, sooty, thin-bedded limestone sample was rich in silicified rhynchonelliformean brachiopods, which were etched free with dilute hydrochloric acid, and are now stored in the geological collection of the Natural History Museum of Denmark, Copenhagen. Currently described phosphatic material was released from the matrix by treatment of a small sample of the limestone in weak acetic acid. The residue contained abundant black, lustrous specimens of Sphenothallus sp., associated with linguliformean brachiopods and conodonts, the latter described by Armstrong (Citation1990). Selected specimens were coated with gold and examined using scanning electron microscopy, prior to the assembly of images in Photoshop CS4.
Abbreviations and repositories
GGU prefix indicates a sample made by Grønlands Geologiske Undersøgelse (Geological Survey of Greenland, now a part of the Geological Survey of Denmark and Greenland; GEUS), Copenhagen, Denmark; PMU prefix, palaeontological type collection of the Museum of Evolution, Uppsala University, Sweden.
Morphology
Specimens of Sphenthallus from GGU sample 216842 achieve a maximum preserved length of about 3 mm, but are always broken at the apertural margin (). Individual specimens consist of a basal holdfast with a hollow tube perpendicular to its surface ().
Holdfast
The holdfast is usually circular in plan view, most frequently with a planar peripheral margin reflecting attachment to a flat surface (), although some specimens preserve an irregular holdfast due to their attachment to irregular surfaces (). The holdfast is conical () with height varying from one third to half of the overall diameter; its sides are shallowly convex. There is a smooth, but abrupt, transition to the tube, which initally rises perpendicular to the holdfast but subsequently often curves slightly to one side (). In terms of outer dimensions, the tube varies from between half and three quarters of the diameter of the holdfast. In all available specimens, the under surface of the holdfast is a funnel that passes gradually into the interior of the tube (), but its inner surface is usually irregular (), with a more granular texture than the thin, dense, outer layer.
In one specimen (–C), a distinct, narrow, peripheral brim abuts against the periphery of a basal attachment disc (), with both displaying a fine radial ornamentation of broad rounded radial ridges separated by sharp, narrow, grooves (). In the former specimen ( and inset), the broken inner margin of this radially ornamented zone is wrinkled and appears to have been flexible prior to final mineralization. The displayed surface of the detached attachment disc preserves two layers centrally but traces of comarginal laminae show a down-stepping towards the periphery in the specimen as illustrated, with overlying lamellae appearing towards the centre (, arrow). Comparison of the ornamentation of the disc surface and the traces preserved in the marginal zone of the holdfast ( and inset) indicates that the disc surface represents the internal surface of the attachment disc and not the actual attachment surface, which is the opposite (under side) side of the specimen as illustrated.
The down-stepping and lamellar overlap on the inner surface of the attachment disc seem to indicate that the disc was initially formed at its maximum present diameter and subsequently was thickened by the internal addition of lamellae on its upper (inner) surface. This is the method of growth described by Müller et al. (Citation1974) in the phosphatic ring Phosphannulus Müller et al., Citation1974, a likely junior synonym of Sphenothallus (Landing et al. Citation2018). The radial ornamentation of rounded ridges alternating with narrow grooves is a feature of the disc rather than an impression of the substrate. It is not known if the central oval perforation is original or diagenetic in origin. It is between about a quarter and a fifth of the diameter of the disc but its margins lie within the plane of the disc, not curving in towards the base of the tube.
Tube
The sides of the tube are sub-parallel with only slight overall expansion towards the aperture. Initially, the tube is circular in cross-section, just distal of the holdfast (), but the cross-section quickly becomes elliptical with the development of angulations at each end of the long axis of the ellipse (). Internally, however, the circular shape is usually maintained, with the shell ellipse becoming massively thickened at its angulations (). Fine, transverse growth lines and weak comarginal undulations are most clearly developed on the lateral areas between the thickened angulations and indicate addition of thin-shell laminae on the inside of the tube (, arrow). The junction between the thin lateral areas and the thick angulations may be marked by a low longitudinal ridge, groove or combination of both (, 3E).
In broken specimens, the thickened longitudinal angulations to the tube are often preserved as characteristic channel-shaped fragments with a concave inner surface in which the margins display the fine lamination of the tube wall (). Breakage of the thinner lateral areas of the tube often produces deep invaginations in the wall between the thickened angulations ().
Shell structure
While growth lamination is visible on the outer surface of some specimens (, arrow), it is most conspicuous on broken surfaces (–F,I). Individual laminae may thin considerably from the thickened extremities of the shell profile towards the thinner lateral areas (, from arrow a to arrow b). Uniform thickness of laminae is maintained in longitudinal profile, such as along the broken edges of the channel-shaped fragments (), although the transverse thickness may be variable. Vinn & Mironenko (Citation2021) described a plywood-like structure in the wall of Sphenothallus from the Carboniferous of central Russia. While this cannot be demonstrated in the Greenland specimens, it is consistent with the textural alternation seen on broken surfaces (). Skovsted & Peel (Citation2011, ,5) described similar lamination with layers of alternating texture in Hyolithellus Billings, Citation1871 from the Cambrian (Series 2, Stage 4) of Greenland.
Breakage and repair
One specimen shows a sudden change in the direction of growth and a transverse dislocation in growth lines that probably represents re-alignment of the tube following injury or disturbance (, arrows). It is possible, however, that the disturbance results from budding, and eventual loss, of a side branch since branching is reported in other specimens assigned to Sphenothallus (Van Iten et al. Citation2019).
Discussion
The longitudinal thickening of the extremities of the tube is a characteristic feature of Sphenothallus, noted originally by Hall (Citation1847), and incorporated into the emended diagnosis by Zhu et al. (Citation2000). They are stated to be present in some material referred to Torellella Holm, Citation1893, which may be distinguished by a lateral furrow (Vinn Citation2006) although seemingly not in the type species Torellella laevigata (Linnarsson, Citation1871) from the lower Cambrian of Sweden. Holm (Citation1893) did not mention thickening of the lateral extremities of the elliptical tube in the type species, but lateral ridges are seen in his illustrated cross-section of Torellella taenia (Holm, Citation1893) from the middle Ordovician of Västergötland, Sweden. The latter species clearly formed the basis for the definition of Torellella given by Fischer (Citation1962, p. W133, fig. 75.3), although this Ordovician species bears little resemblance to the type species Torellella laevigata from the lower Cambrian. Fisher (Citation1962) incorrectly referred the illustrated specimen to Torellella laevigata although it depicts the only known specimen, and therefore holotype, of the Ordovician Torellella taenia Holm, Citation1893, illustrated by Holm (Citation1893, pl. 3, figs. 59–62).
Landing et al. (Citation2018) noted that Torellella lacks a holdfast of the type seen in Sphenothallus and suggested it may be an annelid, making comparisons with Hyolithellus as described by Skovsted & Peel (Citation2011) from North Greenland.
Phosphatic rings representing the basal holdfast of Sphenothallus (and possibly other morphologically similar organisms) have been described in detail under the name Phosphannulus Müller et al., Citation1974 from Cambrian–Devonian localities (Müller et al. Citation1974; Ferretti et al. Citation2013). Dzik et al. (Citation2017) and Landing et al. (Citation2018) noted that holdfasts of this type did not appear to be associated with the similar tube Torellella laevigata in the Cambrian. The tubular Byronia Matthew, Citation1899 and Tubulella Howell, Citation1949 did develop such a holdfast (Matthew, Citation1899; Mierzejewski Citation1986) but see discussion by Dzik (Citation1994) and Skovsted & Peel Citation2011). Peel (Citation2014) described circular or elliptical scars of holdfasts of similar size etched into the surface of organophosphatic linguliformean brachiopods from the Kap Troedsson Formation (Cambrian Series 2, Stage 4), but such scars may have been produced by a number of unrelated organisms (Peel Citation2014). Meshkova (Citation1974) and Skovsted et al. (Citation2021) illustrated the expanded base of tubes of Hyolithellus from Cambrian Series 2 that may have produced circular scars of the type described by Peel (Citation2014) and similar specimens occur in equivalent strata in North Greenland. There does not appear to be a published record of undoubted Sphenothallus holdfasts etched into their substrate, although Phosphannulus rings may experience bioimmurization on crinoid stems (Welch Citation1976; Werle et al. Citation1984). There are numerous records of Sphenothallus holdfasts but few adequately show detailed morphology (Linnarsson Citation1871; Mason & Yochelson Citation1985; Van Iten et al. Citation1992; Neal & Hannibal Citation2000; Bodenbender et al. Citation1989; Lerner & Lucas Citation2005, Citation2011).
The inner surface of a basal attachment disc, attached to its substrate, was illustrated by Van Iten et al. (Citation1992, fig. 7B) from the Carboniferous (Namurian) of Montana but its surface details are obscure. The observations of Bodenbender et al. (Citation1989) and Van Iten et al. (Citation1992) concerning the form of the holdfast are generally confirmed by the Washington Land material as regards the presence of the basal attachment disc () and the conical holdfast, although it is not demonstrated that the central part of the holdfast was completely mineralized. However, the outer surface of the holdfast is formed by a thin, dense layer of uniform thickness (3–4 µm) that covers a thicker, rather amorphous and poorly defined layer that extends at least to the base of the tube (). This differentiation of a thin hard, outer layer and an amorphous inner layer does not appear to extend far up into the tube, where the wall is compact and dense like the outer layer of the holdfast.
Specimens of Sphenothallus from Washington Land can be compared to isolated phosphatic specimens described by Dzik et al. (Citation2017) from the Ordovician (early Floian) Fenxiang Formation, Hubei Province, China. Both assemblages contain holdfasts with coplanar () and irregular bases (.M), and a flat basal surface (Dzik et al. Citation2017, fig. 4; ). The short longitudinal ridges within the tube reported by Dzik et al. (Citation2017, fig. 6)) in the Chinese material have not been observed in specimens from Greenland.
Nathorst (Citation1883) referred to Sphenothallus cf. angustifolius a cluster of crushed specimens with tubes up to 60 mm in length and with associated holdfasts from the Silurian of Västergötland, Sweden. Brood (Citation1988) described under the name Campylites pleijeli Brood, Citation1988 a slender species measuring 20–40 mm in length from the Silurian Hemse Formation of Gotland, Sweden. However, the cross-section of the inner tube differs from material illustrated here in being elliptical, rather than circular, with the opposing longitudinal thickenings located on the sides of the thick-walled tube rather than on its long axis. The holdfast is not known. Wang et al. (Citation2003) described numerous tubes and associated holdfasts attributed to Sphenothallus angustifolius from the Lungmachi Formation (Silurian, Llandovery) of Guizhou Province, China, though their preservation inhibits comparison with the Greenland specimens.
Acknowledgments
The sample was collected during the Washington Land Project (1975–1977) of Grønlands Geologiske Undersøgelse. Christian Skovsted, Olev Vinn and an anonymous reviewer are thanked for comments.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Armstrong, H.A., 1990: Conodonts from the Upper Ordovician – lower Silurian carbonate platform of North Greenland. Bulletin Grønlands Geologiske Undersøgelse 159, 151 pp. https://doi.org/10.34194/bullggu.v159.6709.
- Billings, E., 1871: On some new species of Palaeozoic fossils. Canadian Naturalist 6, 213–233.
- Bodenbender, B.E., Wilson, M.A., & Palmer, T.J., 1989: Paleoecology of Sphenothallus on an Upper Ordovician hardground. Lethaia 22(2), 217–225. https://doi.org/10.1111/j.1502-3931.1989.tb01685.x.
- Brood, K., 1988: A new species of Campylites from Gotland. Geologiska Föreningens i Stockholm Förhandlingar 110(1), 83–85. https://doi.org/10.1080/11035898809453125.
- Dawes, P.R., 2004: Explanatory notes to the geological map of Greenland, 1:500 000. Humboldt Gletscher sheet 6. Geological Survey of Denmark and Greenland Map Series 1, 48 pp. https://doi.org/10.34194/geusb.v1.4615.
- Dzik, J., 1994: Evolution of 'small shelly fossils' assemblages of the Early Paleozoic. Acta Palaeontographica Polonica 39, 247–313.
- Dzik, J., Balinski, A., & Sun, Y., 2017: The origin of tetraradial symmetry in cnidarians. Lethaia 30, 306–321. https://doi.org/10.1111/let.12199.
- Fauchald, K., Stürmer, W., & Yochelson, E.L., 1986: Sphenothallus »Vermes« in the Early Devonian Hunsrück Slate, West Germany. Paläontologische Zeitschrift 60(1–2), 57–64. https://doi.org/10.1007/BF02989422.
- Fauchald, K. & Yochelson, E.L., 1990: A tubiculous animal from the Hunsrück Slate (Lower Devonian, southern Germany). Paläontologische Zeitschrift 64(1–2), 15–23. https://doi.org/10.1007/BF02985918.
- Ferretti, A., Cardini, A., Crampton, J.S., Serpagli, E., Sheets, H.D., & Štorch, P., 2013: Rings without a lord? Enigmatic fossils from the lower Palaeozoic of Bohemia and the Carnic Alps. Lethaia 46(2), 211–221. https://doi.org/10.1111/let.12004.
- Fisher, D.W., 1962: Small conoidal fossils of uncertain affinities. In R.C. Moore (ed.): Treatise on Invertebrate Paleontology part W, W98–W143. Geological Society of America and University of Kansas Press , Boulder, CO and, Lawrence, KS, 259 pp.
- Hall, J., 1847: Palaeontology of New York, containing descriptions of the organic remains of the Lower Division of the New York System, vol. 1, 338 pp. C. Van Benthuysen, Albany.
- Higgins, A.K., Ineson, J.R., Peel, J.S., Surlyk, F., & Sønderholm, M., 1991: Lower Palaeozoic Franklinian Basin of North Greenland. Bulletin Grønlands Geologiske Undersøgelse 160, 71–139. https://doi.org/10.34194/bullggu.v160.6714.
- Holm, G., 1893: Sveriges kambrisk-siluriska Hyolithidae och Conulariidae. Sveriges Geologiska Undersökning, Afhandlingar Och Uppsatser C 112, 172 pp.
- Howell, B.F., 1949: New hydrozoan and brachiopod and new genus of worms from the Ordovician Schenectady Formation of New York. Bulletin of the Wagner Free Institute of Science 24, 8 pp.
- Hurst, J.M., 1980: Silurian stratigraphy and facies distribution in Washington Land and western Hall Land, North Greenland. Bulletin Grønlands Geologiske Undersøgelse 138, 95 pp. https://doi.org/10.34194/bullggu.v138.6680.
- Hurst, J.M. & Surlyk, F., 1982: Stratigraphy of the Silurian turbidite sequence of North Greenland. Bulletin Grønlands Geologiske Undersøgelse 145, 121 pp. https://doi.org/10.34194/bullggu.v145.6687.
- Koch, L., 1929; Stratigraphy of Greenland. Meddelelser Om Grønland 73(2), 205–320.
- Landing, E., Antcliffe, J.B., Geyer, G., Kouchinsky, A., Bowser, S.S., & Andreas, A., 2018: Early evolution of colonial animals (Ediacaran evolutionary radiation–Cambrian evolutionary radiation–great Ordovician biodiversification interval). Earth-Science Reviews 178, 105–135.
- Lerner, A.J. & Lucas, S.G., 2005: First New Mexico record of Sphenothallus from the Upper Pennsylvanian (Missourian) Atrasado Formation of Socorro County, New Mexico. New Mexico Geology 24, 60–61.
- Lerner, A. & Lucas, S.G., 2011: Allocthonous Sphenothallus (Cnidaria) from a lacustrine lagertsätte, Carboniferous of New Mexico, USA. Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin 53, 86–89.
- Li, G., Zhu, M.Y., Van Iten, H., & Li, C., 2004: Occurrence of the earliest known Sphenothallus Hall in the Lower Cambrian of southern Shaanxi Province, China. Geobios 37(2), 229–237. https://doi.org/10.1016/j.geobios.2003.04.002.
- Linnarsson, J.G.O., 1871: Geognostiska och palæontologiska iakttagelser öfver Eophytonsandstenen i Vestergötland. Kongliga Svenska Vetenskaps-Akademiens Handlingar 9, 1–19.
- Mason, C. & Yochelson, E.L., 1985: Some tubular fossils (Sphenothallus: “Vermes”) from the middle and late Paleozoic of the United States. Journal of Paleontology 59, 85–95.
- Matthew, G.F., 1899; Upper Cambrian of Mount Stephen, British Columbia: the trilobites and worms. Royal Society of Canada, Transactions Series 2 5(4), 39–56.
- Meshkova, N.P., 1974: Hyolithelminthes. In B.S. Sokolov, and L.N. Repina (eds.): Biostratigraphy and fauna of the lower Cambrian of Charaulach (ridge Tuora-Sis), 70–73. Trudy Instituta Geologii i Geofyziki, Akademiya Nauk SSSR, Sibirskoe Otdelenie 235, 239 pp. [in Russian].
- Mierzejewski, P., 1986: Ultrastructure, taxonomy and affinities of some Ordovician and Silurian organic microfossils. Palaeontologia Polonica 47, 129–220.
- Müller, K.J., Nogami, Y., & Lenz, H., 1974: Phosphatische Ringe aus Mikrofossilien im Altpaläozikum. Palaeontographica Abt. A 146, 79–99.
- Muscente, A.D. & Xiao, S., 2015: New occurrences of Sphenothallus in the lower Cambrian of South China: implications for its affinities and taphonomic demineralization of shelly fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 437, 141–164. https://doi.org/10.1016/j.palaeo.2015.07.041.
- Nathorst, A.G., 1883: Om förekomsten af Sphenothallus cfr angustifolius Hall i silurisk skiffer i Vestergötland. Geologiska Föreningens i Stockholm Förhandlingar 6(8), 315–319. https://doi.org/10.1080/11035898309444062.
- Neal, M.L. & Hannibal, J.T., 2000: Paleoecologic and taxonomic implications of Sphenothallus and Sphenothallus-like specimens from Ohio and areas adjacent to Ohio. Journal of Paleontology 74(3), 369–380. https://doi.org/10.1017/S0022336000031644.
- Peel, J.S., 2014: Failed predation, commensalism and parasitism on lower Cambrian linguliformean brachiopods. Alcheringa 39, 149‒163.
- Poulsen, C., 1934: The Silurian faunas of North Greenland. I. The fauna of the Cape Schuchert Formation. Meddelelser Om Grønland 72,2,1, 467 pp.
- Skovsted, C.B., Balthasar, U., Vinther, J., Sperling, E.A., & Álvaro, J., 2021: Small shelly fossils and carbon isotopes from the early Cambrian (Stages 3–4) Mural Formation of western Laurentia. Papers in Palaeontology 7(2), 961–983. https://doi.org/10.1002/spp2.1313.
- Skovsted, C.B. & Peel, J.S., 2011: Hyolithellus in life position from the Lower Cambrian of North Greenland. Journal of Paleontology 85(1), 37–47. https://doi.org/10.1666/10-065.1.
- Van Iten, H., Cox, R.S., & Mapes, R.H., 1992: New data on the morphology of Sphenothallus Hall: implications for its affinities. Lethaia 25(2), 135–144. https://doi.org/10.1111/j.1502-3931.1992.tb01378.x.
- Van Iten, H., Leme, J.M., Simões, M.G., & Cournoyer, M., 2019: Clonal colony in the Early Devonian cnidarian Sphenothallus from Brazil. Acta Palaeontologica Polonica 64, 409–416.
- Vinn, O., 2006: Possible cnidarian affinities of Torellella (Hyolithelminthes, Upper Cambrian, Estonia). Paläontologische Zeitschrift 80(4), 384–389. https://doi.org/10.1007/BF02990210.
- Vinn, O. & Kirsimäe, K., 2015: Alleged cnidarian Sphenothallus in the Late Ordovician of Baltica, its mineral composition and microstructure. Acta Palaeontologica Polonica 60, 1001–1008.
- Vinn, O. & Mironenko, A.A., 2021: Discovery of plywood structure in Sphenothallus from Gurovo Formation (Mississippian), central Russia. Annales Societatis Geologorum Poloniae 91, 67–74.
- Wang, Y., Hao, S.-G., Chen, X., Rong, J.-Y., Li, G.-X., Liu, J., & Xu, H., 2003: Sphenothallus from the Lower Silurian of China. Journal of Paleontology 77(3), 583–588. https://doi.org/10.1017/S0022336000044267.
- Welch, J.R., 1976: Phosphannulus on Paleozoic crinoid stems. Journal of Paleontology 50, 218–225.
- Werle, N.G., Frest, T.J., & Mapes, R.H., 1984: The epizoan Phosphannulus on a Pennsylvanian crinoid stem from Texas. Journal of Paleontology 58, 1163–1166.
- Zhu, M.-Y., Van Iten, H., Cox, R.S., Zhao, Y.-L., & Erdtmann, B.D., 2000: Occurrence of Byronia Matthew and Sphenothallus Hall in the Lower Cambrian of China. Paläontologische Zeitschrift 74(3), 227–238. https://doi.org/10.1007/BF02988098.