Abstract
Anonyxmolytes lilliput gen. nov., sp. nov. (Curculionidae, Molytinae) is described from Vietnam. Anonyxmolytes lilliput is a blind and wingless weevil, characterized by very small size, presence of a layer of exudate, possibly wax, on the body, thickened on the upper and lower margins of the fore femora and the outer margins of the tibiae, and reduced tarsi, lacking claws. The new genus seems phylogenetically related to Otibazo Morimoto; biogeographical implications are discussed.
Introduction
Recent entomological investigations in the Oriental Region have resulted in the collection of several undescribed taxa of Curculionidae: Molytinae, including many soil‐dwelling species. Molytinae Schoenherr, 1823 is one of the largest subfamilies of Curculionidae, and at least 90 genera are known from the Oriental Region (Alonso‐Zaragaza & Lyal Citation1999). Several tribes have been proposed to accommodate apparently related genera, the majority of which were defined in old works (i.e. Lacordaire Citation1863, Citation1866). The recently erected genera were usually assigned to one of the existing tribes, according to their morphology, but often very doubtfully, as in Zherikhin (Citation1987). Some tribes were recently proposed for single genera, such as Galloisiini (Morimoto 1962) for Galloisia Hustache, 1920 and Microgalloisia Voss, 1962, and Thalasselephantini (Alonso‐Zarazaga & Lyal Citation1999) for Thalasselephas Korotyaev & Egorov, 1976. No discussion based on phylogenetic analyses and biogeographical considerations has ever been attempted, so that many tribes, as listed in Alonso‐Zarazaga and Lyal (Citation1999), have a very heterogeneous composition and probably represent a polyphyletic assemblage of taxa. This is particularly true for the Oriental molytine fauna, about which little is known. Three recent contributions (Meregalli Citation2003a, Citationb, Citation2004) consider specific taxa in the fauna, but a phylogenetic analysis of the relationships of all Molytinae of the Oriental Region is beyond the scope of the present paper, which deals with the diagnosis of a remarkable new genus of soil dwelling weevil from Vietnam, showing a high degree of adaptation to the endogean habitat. It should be mentioned here that Alonso‐Zarazaga and Lyal (Citation1999), following the most recent placement at that time, included several genera of the Oriental fauna in the Anchonini Imhoff, 1856, a tribe recently re‐defined by Voisin (Citation1992, Citation1994). According to Meregalli (Citation2003a, Citationb), these genera should be removed from this tribe, which is native to Central and South America, with only one species of Anchonus Schoenherr, 1825 and one of Acorep Voisin, Citation1992 in Madagascar and Eastern Africa, respectively. The relationships of two genera from Mauritius and the Seychelles Islands (Cycloterinus Kolbe, 1910; Tanyomus Champion, 1914) listed in Anchonini by Alonso‐Zarazaga and Lyal (Citation1999) but not cited in the key to the tribe by Voisin (Citation1994) need to be confirmed.
Material and methods
This study is based on specimens entrusted to us by Dr. Louis Deharveng of the National Museum of Natural History of Paris, France. Specimens of other genera used to recognize phylogenetic relationships of Anonyxmolytes are preserved in the Meregalli collection. All examined specimens with the exception of one female were dissected; female genitalia were embedded in Solacryl (Medika, Prague, Czech Republic) and male genitalia were either embedded in Solacryl or mounted dry. Genitalia preparations are pinned below the specimen from which they were dissected. Drawings were made using a Wild M5A stereomicroscope with the aid of a camera lucida; photographs were taken with a Nikon Coolpix 4500 camera, on the same stereomicroscope and on a Leitz microscope, and were enhanced with Photoshop 7.0 (Adobe Systems Incorporated). The SEM pictures were done at the Service Commun de Microscopie Électronique du Laboratoire des Sciences de la Vie du Muséum National d'Histoire Naturelle, Paris; critical point was obtained on Emitech K850 and samples were gold sputtered on JEOL JFC‐1200; prepared specimens were observed on a JEOL JSM 840A scanning electronic microscope and digitally photographed.
Taxonomic accounts
Anonyxmolytes n. gen.
Gender: masculine
Type species: Anonyxmolytes lilliput Meregalli & Osella, n. sp.
Diagnosis
A genus of Curculionidae, subfamily Molytinae. Size small, less than 1 mm, wingless, blind, weakly convex; integument reddish, smooth, relatively glossy, covered with a translucent layer of exudate, composed of thin polygonal scales, apparently of wax; upper margin of scrobes, outer margin of tibiae and upper and lower margin of fore femora with a thicker layer of exudate; tarsi, including segment 3, reduced, claws absent; vestiture composed of sparse erect setae.
Rostrum stout, cylindrical in cross‐section, of uniform width from base to apex, as long as pronotum, pregenae not expanded below the scrobes and not visible from above; in lateral view rostrum of uniform thickness from base to apex, strongly and regularly curved from base; apex with some setae oriented anteriorly; basal half with a layer of exudate. Epistoma smooth, flat, scarcely developed, apex slightly curved. Mandible small, with one basal seta; maxillae subquadrate, with two setae; maxillary palps short, prementum flat. Upper margin of scrobes slightly keeled, sharply delimiting the lower part of the rostrum, with a scarcely apparent narrow translucent exudate; scrobes not extended beyond antennal insertion, not visible from above, running almost completely along underside of rostrum. Antennae inserted at 2/3 length of rostrum from base, short, funicle 7‐segmented, the first segment much thicker than the second, the remaining segments progressively widened; club oval, segment 1 and 2 with few setae, as long as the remaining part of club, segment 3 more densely setose.
Head globose, only slightly larger than the base of rostrum, more glossy and smooth, unpunctured, well defined by a shallow transversal depression on base of rostrum.
Pronotum subcylindrical, longer than wide, apex wider than base, sides moderately and sub‐linearly widened from base to near apex, base and apex truncate, postocular lobes completely absent.
Scutellum indistinct.
Elytra oval‐oblong, at base as wide as base of pronotum, surface smooth, very uniform, intervals not convex, striae not impressed, apparently 9, 1 to 4 and 8–9 complete from base to apex, 5–7 shortened, irregular, very badly distinct; declivity, in lateral view, abrupt.
Prosternum flat beyond fore coxae; apex truncate; length between fore coxae and apex about half as long as diameter of coxae; length between fore coxae and base as long as diameter of coxae. Fore coxae contiguous at base, globose‐conical. Middle coxae slightly separate, globose‐conical. Hind coxae globose‐transverse, widely separate. Metasternum much shorter than diameter of middle coxae.
Ventrites 1 and 2 fused, suture indistinct, surface weakly depressed in male, slightly convex in female, longer that ventrites 3–5 together; intercoxal process of ventrite 1 truncate, as wide as diameter of hind coxae; ventrites 3–4 very short, sublinear; ventrite 5 semicircular, with some erect setae.
Legs short, fore femora flattened and curved inward, untoothed, upper margin with a few long erect setae; upper and lower margins with a thick layer of exudate; middle and hind femora lacking a layer; tibiae very short, wide, flattened, perpendicularly truncate at apex, with a long, curved apical uncus, inner preapical mucro microscopic, inner preapical comb present, composed of a few setae; outer margin with a thick layer of exudate; tarsi short, narrow, onychium and claws absent.
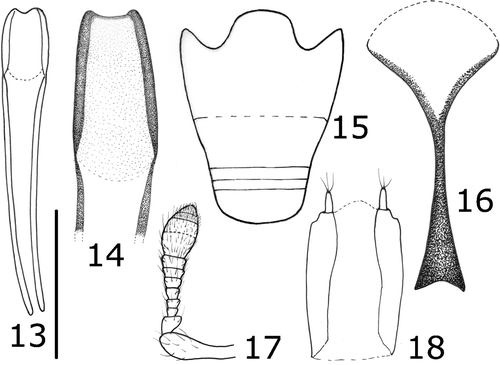
Male genitalia: aedeagus subrectangular, weakly curved, apex canaliculate, dorsum not sclerotized, dorsal structures absent, apodemes slender, three times as long as the median lobe; internal sac without sclerotized pieces; tegmen lacking a tectum, with a long manubrium, nearly as long as aedeagus apodemes; sternite VIII narrow, transverse, with weakly broadened lobes; sternite IX narrow, straight for most of its length, weakly sclerotized.
Female genitalia: sternite VIII with Y‐shaped apodeme, arms divergent, basal part of apodeme long. Coxites very long and robust; styli cylindrical, elongate. Spermatheca very weakly sclerotized, cornu thick.
Etymology
The name of the genus derives from Greek óόνυξ (onyx, claw), preceded by privative alpha, and the name of genus Molytes, meaning a Molytine without claws.
Anonyxmolytes lilliput sp. nov.
Type locality: Vietnam: Vietnam, prov. Kien Giang, distr. Kien Luong; Nui Hon Chong [10°10'N 104°37'E]
Holotype ♀: Vietnam, Kien Giang, Kien Luong; Nui Hon Chong, Chua Hang (pagode) sous brousailles—Berlese 500 cm3 leg. Le Cong Man, 10.135662N 104.644000E, 19.I.2000 (Muséum National d'Histoire Naturelle, Paris).
Paratypes: same data, 1 ♂ (coll. Meregalli); Kien Giang, Kien Luong; Nui Hon Chong, soil 3 near Hang I, Berlese, 27.XI.2006, Le Cong Man & Quan leg., 1 ♂ 1 ♀ (Muséum National d'Histoire Naturelle, Paris); Kien Luong: Hon Chong; Nui Hon Chong, col au‐dessus de la pagode, 22.VIII.2003, 10.137176N 104.649018E, 1 ♀, Deharveng & Bedos legg. (coll. Osella); Vietnam, Kien Giang, distr. Kien Luong: Hon Chong; Nui Khoe La, 30.XI.2005, 1 ♂ 1 ♀ (Muséum National d'Histoire Naturelle, Paris)
Diagnosis
A blind endogean Molytinae characterized by very small size, very smooth sculpture of integument and vestiture composed of few erect setae, perpendicularly inserted on rostrum, pronotum and elytra.
Measurement
Length of body (excluding rostrum): 0.92 mm. Rostrum: length 0.33 mm; width 0.11 mm; ratio length/width 3.00. Pronotum: length 0.31 mm; width at base 0.19 mm; maximum width near apex 0.25 mm; width at apex 0.21 mm; ratio length/maximum width 1.24. Elytra: length 0.58 mm; width 0.39 mm; ratio length/width 1.49. Ratio elytral/pronotal length 1.87; ratio elytral/pronotal width 1.36.
Description
Body (Figures , ) ovate‐elliptical, moderately convex in lateral view, integument reddish, smooth, covered on base of rostrum, pronotum and elytra with a shallow layer of scaly exudate.
Rostrum (Figures , ) stout, as long as pronotum, of uniform width and thickness, in lateral view very regularly curved from base to apex and forming a quarter of a circle; surface smooth, very finely, shallowly and minutely wrinkled or vermiculate, with few erect hair‐like setae, spaced and perpendicularly inserted, half as long as rostrum thickness; dorsum in its basal half with a thin layer of exudate; apex beyond antennae more glossy and smooth, with scarcely distinct sculpture and a few converging setae. Upper margin of scrobes slightly keeled, darker, subrectilinear, reaching base of rostrum, with a very narrow, barely visible translucent layer of exudate. Antennae (Figures , ) short, with sparse setae, some of which bifid; scape short, strongly thickened from base; segment 1 of funicle large, thick, conical, isodiametric, about 2 times broader than segment 2; segments 2–7 progressively widened, with segment 7 wider than 1 and approximately as wide as the first segment of club; club ovate, first two segments with few setae, segment 3 more densely setose. Mandible (Figure ) very small, with acute teeth, prementum subquadrate.
Figures 1–5 Anonyxmolytes lilliput, holotype. Body (1–2); rostrum (3). Paratype ♀: fore tibia (4) and detail of fore tarsus (5). Bars: 1–2 = 0.5 mm; 3 = 0.2 mm; 4 = 0.1 mm; 5 = 50 µm.
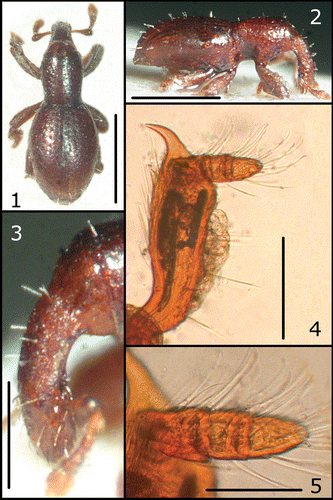
Figures 6–12 Anonyxmolytes lilliput, paratype, Nui Hon Chong. Body (6–7); rostrum, lateral (8) and frontal (10) view; mouth parts (9); hind tibia (11); middle tarsi (12).

Head distinctly more glossy and smooth than rostrum, globose, delimited by a shallow transverse impression at base of rostrum.
Pronotum subcylindrical, longer than wide, apex slightly wider than base, maximum width near apex, sides sub‐linearly broadened from base and very regularly and gently curvilinear at point of maximum width; surface, below the exudate, minutely but distinctly vermiculate, lacking any trace of keel, with very shallowly impressed small and sparse punctures, barely visible as minute depressions; vestiture composed of some scattered erect hair‐like setae.
Elytra oval‐oblong, flattened on dorsum, declivity abrupt; base as wide as base of pronotum, sides regularly curvilinear, maximum width slightly before middle of elytral length; integument smooth, microscopically vermiculate as pronotum, all intervals flat, striae not impressed, with very small shallow punctures, striae on sides incomplete, detectable only around middle of length; vestiture composed of few long, hyaline hair‐like setae, spaced and perpendicularly inserted on odd intervals.
Legs (Figures , ) short, fore femora distinctly flattened from base to near apex, slightly curved inward, with some long setae and a thick layer of exudate on upper and lower margins; middle and hind femora scarcely flattened, lacking exudate; tibiae flattened, very short, rectangular, outer margin straight, with 3‐4 long erect setae and a thinner layer of exudate; subapical comb oblique, composed of a few short rigid setae; apex flattened, sub‐perpendicularly truncate, with a series of apical setae; uncus strong, curved inward; premucro microscopic, barely visibile on fore tibiae; tarsi strongly reduced (Figures , ), narrow, short, basal segment triangular, slightly longer than wide, segment 2 short, transverse; segment 3, broad, flattened, as long as wide, conical in lateral view; all segments with slender downwards curved setae, segment 3 with a fringe of long apical setae downwards curved.
Ventrites (Figure ) 1 and 2 with vermiculate surface and with very minute punctures; 3 and 4 narrow, slightly darker; 5 semicircular, with some long setae.
Genitalia: Figures .
Figures 13–18 Anonyxmolytes lilliput, paratype, Nui Hon Chong. Aedeagus, dorsum and detail of median lobe (13–14); ventrites (15); sternite VIII of ♀ (16); ovopositor (18); antenna (17). Bar: Figures : 65 μm; Figure : 0.1 mm; Figures : 0.2 mm; Figure : 0.3 mm.
Variation
The five specimens from Nui Hong Chong are quite uniform; size of the body excluding rostrum varies between 0.92 and 1.05 mm. The two specimens from Nui Khoe La, a hill about 7 km distant, have slightly longer setae on pronotum and elytra. As reported by Deharveng (personal communication), these limestone hills are isolated outcrops, separate from each other by flat wetland, a habitat not suitable for these blind endogean weevils. It is therefore not surprising that the two populations are very slightly distinct.
Etymology
The specific epithet is taken from Lilliput, the island conceived by the writer Jonathan Swift (1667–1745) that was inhabited by very tiny people, in reference to the extremely small size of the new species. It is to be treated as a substantive in apposition.
Distribution and ecology
The type locality lies in north‐western Vietnam. The two hills where the specimens were found are limestone rocky outcrops (Figure ), with caves and low secondary forest. The specimens were collected sifting 500 cm3 of forest soil (Deharveng, personal communication), together with specimens of Acicnemis sp. and a presently undetermined microphthalmic Entiminae.
Figure 19 Nui Hong Chong, the habitat ofAnonyxmolytes lilliput gen. n., sp. n. (photo Deharveng).

Remarks
Adaptation to the endogean habitat in this species has resulted in the modification of several characters, and the phylogenetic affinities of the new genus, even at the family‐group level, were not immediately apparent because of the high level of homoplasy in these character states. The endogean‐only Raymondionymidae, although superficially similar, were excluded because this family has an aedeagus with a tectum and the structure of the apex of the tibiae is different as is the structure of the rostrum. Anonyxmolytes shares with Molytinae the overall structure of the male and female genitalia, as well as the morphology of the rostrum, the mouth parts, the thoracic parts and the legs.
The most striking and peculiar trait of Anonyxmolytes is the extreme reduction of the tarsi, with a complete loss of the claws, a state which does not appear to be associated with endogean habits per se.
Extreme reduction of the tarsus is relatively uncommon in Curculionoidea, occasionally appearing in unrelated taxa (i.e. Diabathrariini Lacordaire, Citation1863; Cryptoplini Lacordaire, Citation1863; some genera of Anoplini Bedel, 1884; some Magdalini Pascoe, 1870, personal observation and data supplied by R. Thompson), but in all these taxa the tarsal segments are regularly developed, and segment 3 is generally broad and bilobed. On the other hand, in Anonyxmolytes all tarsomeres are extremely reduced, onychium and claws are absent and segment 3 presents an apical fringe of long setae. This extreme reduction of the tarsus in Anonyxmolytes, in our knowledge, does not appear in any other weevil and may thus be unique within Curculionoidea. It is therefore regarded as a very peculiar autapomorphic state. Blindness and extreme adaptation to the endogean environment are not very common in Molytinae; several endogean taxa are part of the Lymantini Lacordaire, Citation1866, a molytine tribe endemic to Central America, the Caribbean Islands and some areas of the USA (blind genera are, among others, Caecossonus Gilbert, 1955; Decuanellus Osella, 1977; Pseudoalaocybites Osella, 1980). There are also some presently undescribed very small blind weevils probably referable to Lymantini, from Venezuela, that lack claws as well (Anderson, personal communication); these taxa were not examined, but other morphologic traits of Anonyxmolytes apparently exclude any relationship with that tribe. The most promising candidate to share phylogenetic relationships with Anonyxmolytes seems Otibazo Morimoto, 1961. This genus is composed of a few leaf litter species native to Japan, plus some still unnamed species from the Oriental Region, presently under study (Meregalli, unpublished). Otibazo is a well characterized genus, and all the species examined are very similar to each other. It shares several traits with Anonyxmolytes: integument completely smooth, sparsely punctured; rostrum curved, of equal width and thickness from base to apex; antennal scrobes oblique, mainly running to underside of rostrum and not visible from above; antennal funicle with segment 1 thickened, much thicker than segment 2 and segments 2–7 progressively widened; club with segment 1 and 2 glossy, scarcely setose, segment 3 with dense hair‐like setae; pronotum cylindrical, broadened anteriorly and with slightly sinuate sides; scutellum indistinct; elytra at base not larger than the pronotum, very smooth, with completely flat intervals; tibiae with a strong, inwardly curved uncus; upper margin of the scrobes, upper and lower margins of the fore femora and outer margin of the tibiae with a more or less thick layer of exudate, very similar to that of Anonyxmolytes, included into a thick vestiture of minute setae.
An exudate on the integument, probably a wax coating, is a relatively common feature in Curculionoidea. In Molytinae it appears, for example, in Paipalephorus Jekel, 1873, Peribleptus Schoenherr, 1843, Paramecops Schoenherr, 1825, etc.; it was also documented for some species of Cryptoplini, a tribe presently referred to Curculioninae, which, however, do not have any morphological similitude with the new genus. In all these cases, moreover, the exudate covers elytral and in some cases pronotal integument but is absent from legs.
This very peculiar layer appears in Otibazo also on the apex of the antennal scape and on the upper margin of the middle and hind femora, its extent depending on the species; together with minute setae, it reflects light, and looks like a light and convex broad raised keel contrasting strikingly with the blackish body. This is a very distinctive trait, absent in this aspect and distribution on integument from any other genus of Curculionoidea; its shared presence in Anonyxmolytes and Otibazo might thus be regarded as a synapomorphy, suggesting relationships between the two genera. As indicated by the dark brownish to blackish integument and the presence of eyes, Otibazo species are not as adapted to the endogean habitat as is A. lilliput. Anonyxmolytes differs at the generic rank from Otibazo because it presents several traits of high taxonomic significance: eyes completely absent (eyes present, small, composed of about 20 ommatidia or less in Otibazo); layer of exudate present on base of rostrum, pronotum and elytra, and thickened only on the fore femora and the outer part of the tibia (layer of exudate much more evident on the upper margin of the scrobes and also present on the apex of the antennal scape and on the middle and hind femora in Otibazo); onychium and claws missing (onychium present, although narrow, and minute claws present in Otibazo); ventrites 1 and 2 completely fused, ventrite 2 as long as 1 and ventrites 3 and 4 linear, very short (ventrites 1 and 2 shorter, particularly ventrite 2, not completely fused, suture visible for the whole width; ventrites 3 and 4 longer, relatively similar to ventrite 2 in Otibazo); median lobe of the aedeagus membranous on dorsum (weakly sclerotized); sternum VIII of the female with scarcely sclerotized apical plate (plate regularly sclerotized in Otibazo).
Otibazo has also been reported from Pliocene deposits from the Yukon Territory, Canada (Matthews & Telka Citation1997), when large stands of mixed coniferous forests existed in Beringia. Identification of fossil insects is always complex, because of their usually fragmentary nature, and Matthews and Telka (Citation1997) acknowledged this difficulty. A photograph of the fossil attributed to Otibazo (Matthews & Telka Citation1997, Figure , p. 942) shows the head and rostrum of a small weevil. Noteworthy is the very small round eye, but the fossil differs from the extant species of this genus by the lack of the peculiar layer of exudate on the sides of the rostrum (this absence might be expected on a fossil, but a trace on integument, such as a scar as seen in extant specimens when exudate is removed, should be visible) and by the longitudinally furrowed distinctly stronger and deeper sculpture. Length and aspect of the rostrum, microphthalmy and morphology of the scrobes are similar to Otibazo, but the fossil species cannot be definitely attributed to this genus. It may indeed belong to a taxon close to it, but the alternative possibility that it belongs to a microphthalmic species of Anchonini or Molytini (Plinthina) cannot be completely ruled out. If this fossil species is truly related to Otibazo, it poses challenging biogeographical and paleoecological questions: Matthews and Telka (Citation1997) and Porch and Elias (Citation2000) suggest in fact that Pliocene faunas from the western North American arctic include taxa which are now restricted to the Palaearctic Region. All the genera of weevils found in the Alaskan sites (Matthews & Telka Citation1997) presently have a Nearctic or Holarctic distribution, the fragment referred to Otibazo being the only representative of a fauna now absent from the Nearctic Region. Thus, it was taken to be an example of taxa once having a Holarctic range but now restricted to the Palaearctic Region. However, if the Alaskan taxon truly belongs to this group, then it would have affinities with the temperate and tropical Oriental fauna, and this would be quite surprising: these soil‐adapted weevils are nowadays associated with broadleaved forests in warm and humid habitats, and it seems very likely that their ancestors were similarly specialized. Hence, to reach Alaska they would have expanded their range much earlier than the Pliocene, probably in mid‐Miocene, when temperate broadleaved forests with Quercus, Juglans and Liriodendron reached the Bering latitude (Leopold & Liu Citation1994; White & Ager Citation1994), permitting continuous habitat between North America and eastern Asia along the Bering land bridge. The Alaskan representatives of this Oriental lineage would have survived the shift to colder and drier climatic conditions, with prevalence of coniferous forests, that occurred during the late Miocene and early Pliocene, probably helped by their adaptation to the relatively more protected and stable soil environment. They eventually became extinct when the climate drastically deteriorated with the onset of the first glaciation. This interesting, but purely hypothetical distribution model requires verification. More data on the Alaskan fossils, confirming their suggested affinities with Otibazo, or the discovery of other taxa with south‐eastern Asiatic affinities, are required.
The placement of Otibazo and Anonyxmolytes within Molytinae is uncertain, because their morphologic peculiarities, derived by adaptation to the soil habitat, distinguish them from any other genera of the subfamily. Otibazo was included in Anchonini (cf. Morimoto Citation1982), but as previously noted it does not belong to this tribe. Morphological similarities in the aspect of the body, the glossiness and sculpture of the integument, the shape of the pronotum and elytra are shared with Leiosoma Stephens, 1829, but as they are not supported by similarities in other traits and in the genitalia they are regarded as homoplasy. The extremely peculiar characters of Anonyxmolytes suggest that it should be referred to a new, self‐standing tribe. However, as previously said, taxonomy of Molytinae at the tribe level is still in a state of chaos, and even monophyly of the subfamily is discussed (Marvaldi et al. Citation2002); moreover, relationships between Otibazo and Anonyxmolytes are likely, but cannot be demonstrated beyond any doubt. Thus, at the moment we prefer to avoid proposing a new taxon at tribe rank for these two genera; the tribe which seems more appropriate to accommodate them is Molytini.
Acknowledgements
We acknowledge Louis Deharveng (Muséum national d'Histoire Naturelle, Paris, France) for having entrusted to us these very interesting weevils for study and Céline Houssin (Muséum national d'Histoire Naturelle, Paris, France) for the SEM photos. The collecting trips were organized by Truong Quang Tam (Institute of Tropical Biology, Ho Chi Minh City, Vietnam), Le Cong Man and Le Cong Kiet (Ho Chi Minh University, Ho Chi Minh City, Vietnam), who also participated to the soil fauna collection. We are also grateful to Emilio Balletto (University of Torino, Italy) and Robert Anderson (University of Ottawa, Canada) for a discussion, respectively, about biogeographical aspects and taxonomy, and morphological peculiarities of Anonyxmolytes. Richard Thompson (Natural History Museum, London, UK) and an anonymous reviewer gave us very useful suggestions for improving the paper.
References
- Alonso‐Zarazaga , M. A. and Lyal , C. H. C. 1999 . A world catalogue of families and genera of Curculionoidea (Insecta: Coleoptera) (Excepting Scolytidae and Platypodidae). , Barcelona : Entomopraxis .
- Lacordaire , T. 1863 . “ Histoire Naturelle des Insectes. ” . In Genera de Coléoptères ou exposé méthodique et critique de tous les genres proposés jusqu'ici dans cet ordre d'insectes , Roret : Paris . Vol. 6
- Lacordaire , T. 1866 . “ Histoire Naturelle des Insectes. ” . In Genera de Coléoptères ou exposé méthodique et critique de tous les genres proposés jusqu'ici dans cet ordre d'insectes , Roret : Paris . Vol. 7
- Leopold , E. B. and Liu , G. 1994 . A long pollen sequence of Neogene age, Alaska Range. . Quaternary International , 22/23 : 103 – 140 .
- Marvaldi , A. E. , Sequeira , A. S. , O'Brien , C. W. and Farrell , B. D. 2002 . Molecular and morphological phylogenetics of weevils (Coleoptera, Curculionoidea): Do niche shifts accompany diversification? . Systematic Biology , 51 : 761 – 785 .
- Matthews , Jr. J. V. and Telka , A. 1997 . “ Insect fossil from the Yukon. ” . In Insects of the Yukon. Biological Survey of Canada (Terrestrial Arthropods) Edited by: Danks , H. V and Downes , J. A . 911 – 962 . Ottawa, Canada: Monograph Series no. 2 Ottawa
- Meregalli , M. 2003a . Stenanchonus Voss 1937, a junior synonym of Euthycus Pascoe 1885 (Coleoptera: Curculionidae, Molytinae). . Senckenbergiana Biologica , 82 : 127 – 133 .
- Meregalli , M. 2003b . “ The genus Falsanchonus Zherichin, 1987, with description of six new species (Insecta: Coleoptera: Curculionidae: Molytinae). ” . In Biodiversität und Naturausstattung im Himalaya Edited by: Hartmann , M and Baumbach , H . 323 – 335 . Verein der Freunde und Förderer des Naturkundesmuseum Erfurt e. V
- Meregalli , M. 2004 . The genus Microplinthus, with description of four new species (Coleoptera: Curculionidae, Molytinae). . European Journal of Entomology , 101 : 609 – 627 .
- Morimoto , K. 1982 . The family Curculionidae of Japan. 1. Subfamily Hylobiinae. . Esakia , 19 : 51 – 121 .
- Porch , N. and Elias , S. 2000 . Quaternary beetles: A review and issues for Australian studies. . Australian Journal of Entomology , 39 : 1 – 9 .
- Voisin , J. F. 1992 . Notes sur les genres de la tribu des Anchonini. 1. Généralités, redéfinition du genre Anchonus Schönherr et description de cinq genres nouveaux et de deux sous‐genres nouveaux (Coleoptera, Curculionidae). . Nouvelle Revue d'Entomologie (N.S.) , 9 : 259 – 271 .
- Voisin , J. F. 1994 . Notes sur la tribu des Anchonini. 2. Description de quatre genres nouveaux et revue de six anciens (Coleoptera, Curculionidae). . Nouvelle Revue d'Entomologie (N.S.) , 10 : 327 – 340 .
- White , J. M. and Ager , T. A. 1994 . Palynology, paleoclimatology and correlation of Middle Miocene beds from Porcupine River (Locality 90–1), Alaska. . Quaternary International , 22/23 : 43 – 77 .
- Zherikhin , V. V. 1987 . Curculionidae from the Nepal Himalayas. Part I. Molytinae (Insecta: Coleoptera). . Stuttgarter Beiträge zur Naturkunde A , 411 : 1 – 43 .