
Abstract
Landscape analysis with transects, in the Marina Baja area (province of Alicante, Spain), has contributed to establish the influence of different landscape matrices and some environmental gradients on wild rabbit Oryctolagus cuniculus (Linnaeus, 1758) (Mammalia: Leporidae) abundance (kilometric abundance index, KAI). Transects (n = 396) were developed to estimate the abundance of this species in the study area from 2006 to 2008. Our analysis shows that rabbits have preferences for a specific land use matrix (irrigated: = 3.47 ± 1.14 rabbits/km). They prefer the coastal area (
= 3.82 ± 1.71 rabbits/km), which coincides with thermo-Mediterranean (a bioclimatic belt with a tempered winter and a hot and dry summer with high human density), as opposed to areas in the interior (continental climate with lower human occupation). Their preference for the southern area of the region was also noted (
= 8.22 ± 3.90 rabbits/km), which coincides with the upper semi-arid area, as opposed to the northern and intermediate areas (the north of the region coinciding with the upper dry and the intermediate area with the lower dry). On the other hand, we found that the number of rabbits increased during the 3-year study period, with the highest abundance (
= 2.71 ± 1.30 rabbits/km) in May. Thus, this study will enable more precise knowledge of the ecological factors (habitat variables) that intervene in the distribution of wild rabbit populations in a poorly studied area.
Introduction
Since ancient times, the wild rabbit Oryctolagus cuniculus (Linnaeus, 1758) (Mammalia: Leporidae) has been one of the emblematic species of the Iberian Peninsula, due to its ecological characteristics. At present, and despite its decline (Virgós et al. Citation2007), the wild rabbit continues to be one of the most important small game species, and it is part of the diet of many predators in the Iberian Peninsula, especially predators in danger of extinction such as the Iberian Imperial eagle (Aquila adalberti) and the Iberian lynx (Lynx pardinus) (Ferrer & Negro Citation2004).
Many studies demonstrate that rabbit abundance and distribution are related to the habitat structure and land use that make up the Mediterranean mosaic landscape, which may vary according to potential use of resources (Willot et al. Citation2000) and the risk from predator species of the wild rabbit (Palomares & Delibes Citation1997). Other authors have observed that rabbit is positively related to plant cover (Fa et al. Citation1999), to the presence of patches of shelter (Virgós et al. Citation2003) and to ecotone areas between cereal crops and natural vegetation (Calvete et al. Citation2004).
From the 1950s onwards, the province of Alicante and, in particular, the region of the Marina Baja have undergone important changes, since the appearance of new activities related to tourism, both on the coast and inland. All of these activities have caused a decrease in traditional agricultural practices and, above all in the mountain areas, have caused important disturbances in the naturalized environment of the rural regions of the Mediterranean Basin (Farina Citation1998). Landscape changes have had a great influence on the area, causing changes in the socioeconomics, culture and biodiversity of the area (Martínez-Pérez Citation2000).
The problem that arises is the lack of data on wild rabbit distribution in a coastal region influenced by urban development. Due to the strong territorial transformation, the influence of environmental gradients on this species is unknown.
The aim of this study is to analyse wild rabbit abundance in this coastal zone of the semi-arid peninsula during the period from February 2006 to September 2008, with the following specific objectives:
To analyse the degree of influence of the main landscape matrices on population and abundance of the species.
To describe the influence of the main environmental gradients on a regional level (degree of continentality, human occupation of land, influence of latitude/ombroclimate and of thermotype) and their relation with rabbit distribution and abundance.
To analyse the temporal variation in species abundance according to periods in its annual biological cycle, principally related to reproductive and hunting periods during the 3-year study period.
Materials and methods
Study area
The region of the Marina Baja is located in the southeast of the Iberian Peninsula, in the province of Alicante (). There are 18 municipalities; its area is 580 km2 and it is divided into two sectors, one mountainous and the other coastal. Altitudes range from sea level to 1588 m. As regards hydrography, the two rivers Algar and Guadalest are worthy of mention (Solanas & Crespo Citation2001). The Marina Baja has a semiarid-dry Mediterranean climate, with mild temperatures, a long dry period in the summer and rainfall that is concentrated in spring and autumn (Mas Citation1993).
Figure 1. Location of the Spanish study area.
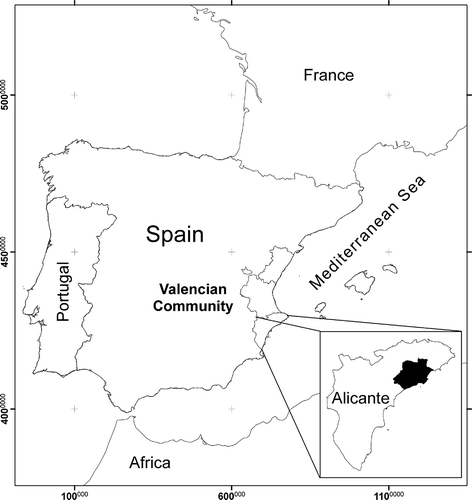
Natural vegetation is the dominant covering in the region of the Marina Baja, covering 54.95% of the surface area (31,717 ha). The territory is a good representation of the cultural landscapes in the Alicante province, from the mountains to the coast, including pieces of padded and thorny oromediterranean vegetation, deciduous forest interspersed between sclerophyll forests, pine forests, thermophilic garrigues, salt and semi-arid steppe communities. This is followed by the three agricultural matrices: irrigated area (loquats, citrus and vegetable patches: 15.09%), abandoned field (old crops and opportunistic vegetation: 10.81%) and dryland farming (olives, carobs, almonds and cereals: 9.94%), and urban land (9.21%) ().
Figure 2. Land use matrices: natural, abandoned, irrigated and dry land.
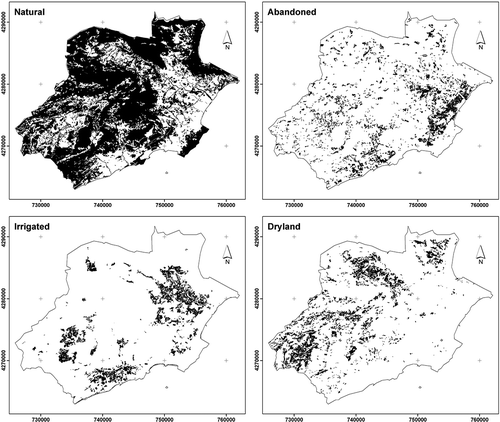
The classification according to thermotype was based on the magnitude of the thermoclimatic index (It, Rivas-Martínez Citation2004), and differentiated transects in three categories: supra-Mediterranean (mountain climate characterized by frequent frosts in winter and high temperatures in summer; T 8 to 13°C, m –4 to –1°C, M 3 to 8°C, It 70 to 200), meso-Mediterranean (bioclimatic belt characterized by a typical Mediterranean climate with frost in winter and summer high temperatures and water stress; T 13 to 17°C, m –1 to 5°C, M 8 to 14°C, It 200 to 360), and thermo-Mediterranean (bioclimatic belt with a tempered winter and a hot and dry summer; T 17 to 19°C, m 5 to 10°C, M 14 to 18°C, It 360 to 470). Thermoclimatic symbols represent T = mean annual temperature, m = average minimum temperature of the coldest month, and M = average maximum temperature of the coldest month and It = (T + m + M) 10. Thus, the thermo-Mediterranean (22,672.71 ha) runs parallel to the coast and the meso-Mediterranean (30,047.64 ha) coincides with the next band towards the interior. The supra-Meditarreanean, with a lower surface area (4987.32 ha), is found in the areas that are furthest away from the coast and coincides with the mountainous zones. To a certain extent, the thermotype coincides greatly with the coastal-inland stratification ().
Figure 3. Environmental gradient: thermo-Mediterranean which coincides with coast, meso-Mediterranean which coincides with intermediate zone, and supra-Mediterranean. The transects were located on the map. Different land cover identified were: N (natural), A (abandonment), S (dry crops) and R (irrigated crops).
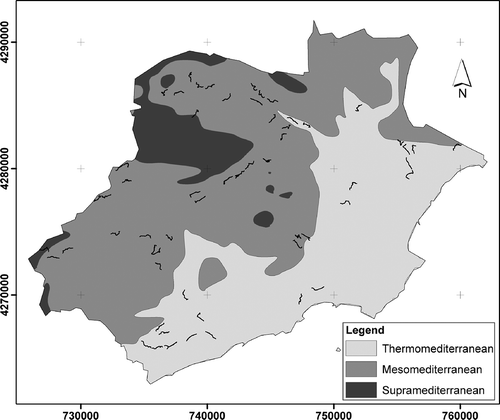
The differences between the coast and the interior of the region are well known. Thus, the degree of human land occupation decreases from the coast to the inland zones of this region (INE Citation2006; Belda et al. Citation2013). The coastal area is the most built up and its human population density is the greatest (8.46 hab/ha). The intermediate area is less built up (1.21 hab/ha) and of a more dispersed type, and the interior of the region is the least populated and urbanized (0.14 hab/ha). On the other hand, ombrotype classification was carried out with the annual ombrothermic index (Io) (Rivas-Martínez Citation2004). The ombrotype coincides to a great extent with the north-south stratification, with the north of the region coinciding with the upper dry with 13,422 ha (Ses, Io: 2.8-3.6), the intermediate area with 27,811 ha of lower dry (Sei, Io: 2.0-2.8), and the south-upper semiarid with 16,620 ha (Sas, Io: 1.5-2.0) ().
Figure 4. Environmental gradient: north/upper dry (Ses), intermediate/lower dry (Sei) and south/upper semiarid (Sas).
Methodology
The relative abundance of wild rabbits (sightings) has been obtained by using stratified transects (linear path on a plot to sample) 1000 m long (n = 44), with a detectability width limit of 100 m and sampling on foot for the same person for 3 hours after dawn or before dusk (Belda & Munera et al. Citation2012; Belda & Zaragozí et al. Citation2012). The average time to sample each transect was 20 min. Sampling transects were chosen at random and according to availability, taking into account the mosaic landscape typology proposed by Jiménez-García (Citation2007), covering the different land cover matrices (): natural (12 transects), abandoned (12 transects), dry land (11 transects) and irrigated area (nine transects) and the different environmental gradients present in the region latitude/ombroclimate, distance from the coast and thermotype. During the field sessions, a series of transects covering the different environmental gradients was carried out. In this way, the following checks were made (). The distance from the coast/human occupation gradient was defined by three strips with the same width (8292.26 m) (Belda & Munera et al. Citation2012). The latitude/ombroclimate and thermotype gradients were sampled and selected according to the criteria of Rivas-Martínez (Citation2004) (see Study area and ).
Table I. Distribution of transects by sampling period in different environmental gradients.
There were three sampling periods without overlapping in each year (2006–2008): post-hunting or pre-reproductive (spring), post-reproductive (summer) and pre-hunting (autumn), for a period of 3 years, for a total of 396 transects. A global positioning system (GPS) Trimble XM® was used in the field sessions, with the aim of georeferencing sampling points and detected wild rabbits. This information about rabbit distribution was exported to a geographic information system (GIS) (ArcGis 9.2®).
Land-cover maps (natural, abandoned, dry land and irrigated areas) were obtained by digitization and georeferencing of aerial photographs (scale 1:5,000) from the Instituto Cartográfico Valenciano (ICV) (Jiménez-García Citation2007). The obtained covers were exported to ArcGIS® and Idrisi Andes® formats in order to create the GIS database and obtain thematic maps (thermotype, continentality, ombroclimate, etc.). Therefore, the cartographic database has the following variables: hunting reserve limits (source: CMAAUV Citation2007), land uses (source: the authors) and the field census (linear transects). In this way, starting from land cover, sampling points were chosen by random, taking into account the typology of the landscape, with the condition of being 1 km long.
Spatial variation of temperature and rainfall was modeled from data in the climatic atlas of the Valencia Region (Pérez-Cueva Citation1994) using Kriging (Oliver & Webster Citation1990), and the resulting estimates were used to calculate the Thermicity and Ombrothermic indices as defined by Rivas-Martínez (Citation1997).
The data of the rabbit sightings detected during the census, in each of the land cover matrices and sampling periods, are presented using the kilometric abundance index (KAI) (Ferry & Frochot Citation1958; Peiro & Blanc Citation2011), expressed as individuals per linear kilometer. In this way, the mean KAI has been obtained by dividing the total number of individuals detected by the total number of transects undertaken in a particular matrix (natural, abandoned, dry and irrigated). This enables comparison between different areas with a different number of transects undertaken.
Data on relative wild rabbit abundances (dependent variable) were subjected to a two-factor analysis of variance (ANOVA). The variance was related to the main land uses (natural, abandonment, dryland and irrigated areas) and to the interaction between the factors thermotype, distance from the coast or latitude (independent variables). The main effects and the interaction between factors have been calculated. Means ± SD were calculated and, when the F ratio was significant, least significant differences were evaluated by the Tukey-b test as available in the SPSS statistical software (version 17.0). Significance levels were always set at 5%. Tukey’s range test is a single-step multiple comparison procedure and statistical test generally used in conjunction with an ANOVA to find which means are significantly different from one another (Liancourt et al. Citation2013). The test identifies where the difference between two means is greater than the standard error (SE) would be expected to allow.
Results
The census carried out in different parts of the region shows that there are significant differences in relative wild rabbit abundance between the different land-cover matrices. The data showed a greater presence of rabbits in the irrigated area ( = 3.47 ± 1.14 rabbits/km), followed by abandoned field and the natural environment. Finally, the dryland farming registered the lowest abundance (). The ANOVA results indicated a significant interaction (p ≤ 0.05) between landscape categories ().
Table II. Abundance of wild rabbit (kilometric abundance index, KAI: rabbits/km) in different land-covers and different environmental gradients.
Table III. Abundance of wild rabbit (kilometric abundance index, KAI: rabbits/km) in different land uses: abandonment (A), natural (N), irrigated areas (R) and dry lands (D) were compared with thermotype: Thermo-Mediterranean (T) and Meso-Mediterranean (Me). Different land uses were compared with the latitude: north (N), intermediate (In) and south (S). Finally, different land uses were compared with the distance from the coast: interior (I), intermediate (M) and coast (C). Data were analysed with two-way analysis of variance (ANOVA) with land uses and above-mentioned factors as the main effects. Means ± SE were calculated and, when the F ratio was significant, least significances were evaluated by the Tukey-b test. Values followed by a common letter are not significantly different. Moreover, ns, *, **, *** indicated non-significant or significant at 5%, 1% and 0.1% levels.
In contrast, ANOVA results indicated, in general, significant differences between rabbit abundances in the coastal gradient. These analyses show that significant differences (Tukey-b test; p < 0.001) exist between the coastal and interior areas (). The highest number of rabbits was detected in the coastal area ( = 3.82 ± 1.71 rabbits/km) and the lowest in the interior zone (
= 0.42 ± 0.30 rabbits/km). Moreover, there was a significant effect between land use and distance from the coast (). During the sampling period, the coastal maximums were obtained in the irrigated area (
= 5.91 ± 2.11rabbits/km) and abandoned matrix (
= 4.67 ± 3.50 rabbits/km). The natural matrix had lower abundance (
= 2.28 ± 1.64 rabbits/km).
ANOVA results indicated an important significant difference in the KAI obtained for the latitude/ombroclimate (p < 0.001). The greatest abundance of rabbits is in the south/upper semiarid area ( = 8.22 ± 3.90 rabbits/km), and much lower KAI values occur in the north/upper dry area (
= 0.44 ± 0.36 rabbits/km). In the south, the greatest abundance of rabbits was in the irrigated lands (
= 14.5 ± 5.40 rabbits/km). The Tukey-b test shows that there are highly significant differences between the latitude categories ().
As regards rabbit distribution according to thermoclimate gradient, data provided by the census show significant differences between the two predominating belts of the region (Tukey-b test, p < 0.001). A greater abundance is found in the thermo-Mediterranean belt ( = 3.44 ± 1.53 rabbits/km) with maximums in the agricultural abandoned zone (
= 4.67 ± 3.50 rabbits/km) and in irrigated areas (
= 4.38 ± 1.50 rabbits/km). In the meso-Mediterranean layer, the mean value obtained is much lower (
= 0.63 ± 0.33 rabbits/km).
As regards rabbit abundance during the study period, this was observed to be increasing (2006 = 1.71 ± 1.45;
2007 = 1.8 ± 1.23;
2008 = 2.25 ± 1.54 rabbits/km), according to one-way variance analysis (UNIANOVA) (F (2.6) = 27.705, p > 0.05).
From the nine seasons analysed, the greatest number of rabbits was obtained in the month of May ( = 2.71 ± 1.30 rabbits/km), followed by February (
= 1.52 ± 1.34 rabbits/km) and September (
= 1.53 ± 1.15 rabbits/km) (UNIANOVA) (F (2.6) = 19.599, p > 0.05). The maximum KAI from the whole period was obtained in May 2008 (3.04 ± 1.54 rabbits/km).
Discussion
Analyses that check rabbit abundance in different landscapes in Marina Baja show that there are significant differences. This region has a structure that is formed by the combination of agricultural and natural matrices that are linked by corridors of natural vegetation. It is, therefore, an area of great contrasts, which goes from the coastal zone to valleys and sheer areas over a relatively short distance, thus providing a rich landscape system (Peña et al. Citation2007; Belda et al. Citation2011; Zaragozí et al. Citation2012). Furthermore, comparable studies have been conducted on game species in nearby regions, using similar methods (Arques Citation2000; Jiménez-García Citation2007; Peiro & Blanc Citation2011; Belda & Zaragozí et al. Citation2012). In agricultural landscapes like ours, KAIs can be used, depending on the aims of study. Thus, wild rabbit KAIs are appropriate when relative abundances or population trends are required (Barrio et al. Citation2010). However, there are several studies which demonstrated that the directed observation has to be corrected for observability along the transect itself (Tizzani et al. Citation2014), but we assume that detection is homogeneous, due to the structure of the vegetation, and include natural and agricultural environments, therefore, the correction is not necessary (Belda & Zaragozí et al. Citation2012).
The greatest presence of rabbits in the census is found in the irrigated lands, dominated by citrus and medlar orchards, despite being a matrix that only occupies 15.09% of the total surface area (Belda et al. Citation2011). In contrast, drip irrigation enables fresh grass to be present all year round; therefore, the rabbits have more food of a better quality here than in other areas. Finally, the abundant margins of the terraced areas are used by the rabbits to build their burrows and warrens for protection against predator species, and to reproduce (Moreno et al. Citation1996). These results coincide with those obtained by Arques (Citation2000) in the south of the province of Alicante, where two neighbouring areas were compared, one with abandoned and the other with irrigated land, and the differences in rabbit abundance were significantly in favour of the irrigated land cultivation.
The next matrix with the greatest number of rabbits is abandoned land (10.81% of total area), principally affecting dryland fruit farming. Natural hedges in cultivated lands and abandoned thickets benefit the rabbits (Lombardi et al. Citation2003; Jiménez-García Citation2007), as they provide the rabbits with corridors and areas of protection. These areas are slowly being covered by thickets, which benefits the rabbits (Lombardi et al. Citation2003; Calvete et al. Citation2004).
The natural category of land use is formed by thickets and Aleppo pine on the coast and in the south of the region, and by not very dense pine formations in mountainous areas, and it is the matrix which occupies most surface area in the region (54.95%). In other parts of the province, it has been demonstrated that lower rabbit abundance in areas with dense pine is due to the fact that this land use reduces the heterogeneity of the environment, above all in the mountain reserves, as can be found in the region of the Marina Alta in the province of Alicante (Jiménez-García Citation2007). On the other hand, it has been demonstrated that altitude affects rabbit abundance and distribution negatively (Trout et al. Citation2000; Cabrera-Rodriguez Citation2006). Jiménez-García (Citation2007) has also proved the existence of a negative relation between rabbit abundance and areas of natural vegetation with no tree cover in the region of the Marina Alta (Alicante).
The dryland farming area, mainly dominated by almonds and olives, counts for 9.94% of the total surface area and has the lowest number of rabbits on the census for the region. The dryland orchards have a negative effect on the global density of huntable species (Jiménez-García Citation2007), although dryland farming benefits granivorous birds and rabbits (Lombardi et al. Citation2003). The rabbit appears in heterogeneous landscapes, where it can find habitats for reproduction, refuge and feeding areas. A rabbit’s territory is no greater than 3 ha, according to some studies (Arques Citation2000; Lombardi et al. Citation2003), and for this reason, rabbits need to have the majority of the resources that they require within a small surface area.
Previous studies indicate that rabbits are influenced by landscape shape index (p = 0.015; R = 0.580) and are distributed irregularly over the surface of the region (Belda et al. Citation2011). This species prefer ecotone areas, where they can find the basic requirements for their survival in a small space. Despite the many changes that have taken place over the last few decades in the region of the Marina Baja, micro-plots still exist and, in general, landscapes are heterogeneous. Habitats that are suitable for rabbits must have abundant and good-quality food during the reproductive period, and be located in areas that are close and with safe access to the warrens. Rabbits are most abundant in the patch areas with shelter (Rogers Citation1981; Virgós et al. Citation2003).
In the study area, there are areas suitable for rabbits; however, rabbits are absent from them. Therefore, stocking programs should be conducted in areas with heterogeneous landscapes, which provide the rabbits, in a small space, with food, suitable areas for breeding and shelter. Restocking should be made with farmed rabbits, possibly genetically pure, or rabbits caught in surrounding areas. Thus, with the establishment of new populations or enlargement of existing ones, we would get a more uniform distribution of the rabbits, and hunters and predators in danger of extinction will benefit.
Rabbit abundance in the region differs statistically according to the distance from the coast. Thus, the coastal area is characterised by having the greatest human population density of the region, with figures above 114,377 inhabitants, and coincides with the thermo-Mediterranean area. Urban areas are an important component in the landscape mosaic of this region and they account for a surface area of 5318 ha (9.21%), mainly situated on the coast. In this area, the greatest rabbit abundance is found in the abandoned area, which has a surface area of 16.76% of the total, and on the irrigated land, which accounts for 14.37% of the surface area. In contrast, the natural matrix of the coastal strip is the one that covers most surface area (64.71%) and has the lowest abundance. Rabbit abundance reduces considerably with the presence of humans (Fa et al. Citation1999; Trout et al. Citation2000) and the species prefers to avoid areas with a dense population (Trout et al. Citation2000). Therefore, the greatest rabbit abundance in the region of the Marina Alta, located to the north and adjoining our study area, is found in areas with a high degree of dispersed urban impact. This type of urban use allows resources to be parcelled, and an increase in the border effect of cultivated land, in the ecotone zones, which are the elements that favour the presence of rabbits (Lombardi et al. Citation2003; Calvete et al. Citation2004; Fernández Citation2005). Changes in the landscape cause their heterogeneity, enabling a large quantity of resources in a small space, and this benefits species such as the rabbit that have small territories (Arques Citation2000; Lombardi et al. Citation2003). In contrast, the coastal zone of the study area coincides with the thermo-Mediterranean layer of the region, with maximums in the abandoned and irrigated areas. These results coincide with those obtained by Jiménez-García (Citation2007), who demonstrated that rabbits showed a preference for habitats located in this belt, in the province of Alicante.
The intermediate area practically coincides with the meso-Mediterranean of the region. This belt is characterised by the presence of cultivated areas, thickets, oak groves and pine woods. What is more, this area is less densely populated by humans than the coastal area, with the kind of disperse urban land which permits conservation of the original patches and even adds new elements such as gardens and areas with trees that allow corridors to be maintained that connect the different patches. These characteristics imply that the abundance of rabbits in this zone is lower than in the coastal area but greater than in the interior of the region.
The interior of the region is where the fewest rabbits are found. It is located towards the west and corresponds to the sheerest mountainous area of the region. Rabbits prefer moderate slopes and avoid areas that are extremely uneven (Jiménez-García Citation2007). The interior coincides with the meso-Mediterranean (30,207.67 ha) and the supra-Mediterranean. It is a belt with low urban density and natural vegetation. Landscape heterogeneity is considerably reduced in hunting reserves located in mountainous areas with pine woods; therefore, the presence of rabbit populations is lower. In previous studies, small game density has been correlated to the landscape heterogeneity using Shannon’s diversity index (p = 0.029; R = 0.708) (Belda et al. Citation2011).
As for the latitude/ombroclimate distribution in the region, greater rabbit abundance has been observed in the southern zone, which coincides with the upper semiarid, followed by the intermediate zone and the north, which coincide with lower dry and upper dry, respectively. In the southern part of the region, the greatest presence of rabbits was found in the irrigated and abandoned land. Nevertheless, it should be mentioned that, in this belt of the region, it was impossible to locate a representative number of transects for the dryland matrix, as this agricultural use has no part to play in the socioeconomic development of the area, where land use is mainly urban or irrigated farming. Similarly, it was impossible to locate sampling points in the supra-Mediterranean area, due to its difficult access (steep slopes), lack of paths and areas with restricted access (military zone).
In general, interannual variations of rabbits are related to various factors, such as the incidence of myxomatosis and viral hemorrhagic disease (RHD) (Villafuerte et al. Citation1995), the availability of food (Villafuerte et al. Citation1997; Gibb & Fitzgerald Citation1998), periods of drought (Myers & Parker Citation1975) or the greater or lesser presence of predators (Pech et al. Citation1992; Banks Citation2000). In the study area, a progressive increase in rabbit abundance was observed during the years under analysis. This tendency has been observed in different parts of the province of Alicante, where unusual demographic explosions have been noted. This has meant an increase in the number of rabbits captured. This increase in rabbit populations is also general in the Iberian Peninsula (Villafuerte et al. Citation1995, Citation1998; Fa et al. Citation1999).
Analysis of rabbit abundance in the three annual periods of the 3-year study demonstrates that the greatest presence of rabbits occurs in the months of May, just when the reproductive period has ended or is ending, a fact also demonstrated by other authors (Arques Citation2000; Lombardi et al. Citation2003). Data from September show a marked decrease in rabbit abundance, mainly due to a specific hunting season in the month of July, the high death rate of young rabbits and the effects of myxomatosis in summer. Moreover, in this month, the reproductive period has not yet started in the province (Arques et al. Citation1999; Arques Citation2000). In February, the number of rabbits increases in relation to the census in September, and this is due to the general hunting period, which begins in the middle of October and finishes the first week of January, and the rabbit population increases as it is in the middle of the reproductive period (Arques Citation2000). The differences between the KAIs in February of different years, 0.73 ± 0.72 rabbits/km in 2006, 1.43 ± 1.12 in 2007 and 2.40 ± 1.91 in 2008, may be related to the different dates when the reproductive season starts in Mediterranean ecosystems and to the increase in population detected in interannual evolution. The start of the reproductive period each year is related to temperature and rainfall (Arques et al. Citation1999; Arques Citation2000). In the province of Alicante, this period usually commences in a period of time between October and the beginning of January, when there is enough rain to make grass grow, and it is at this time that the grass has the sufficient hydric and protein content required by rabbits for reproduction (Lazo Citation1992; Arques Citation2000). Other circumstances, such as the effect of predators and that of viral diseases, that affect rabbits do so in a similar way throughout the year.
In conclusion, our results show that in the region of the Marina Baja, rabbits have specific preferences for the irrigated agricultural matrix. They prefer the coastal zone, which coincides with the thermo-Mediterranean layer, as opposed to the interior zones (meso-Mediterranean), despite the latter being the most densely populated belt. In the first line of the coast, and the following areas, rabbits find very heterogeneous areas, that is, ecotone zones, which encourage their presence (Belda et al. Citation2011). Their preference for the southern part of the region has also been demonstrated, which coincides with the upper semiarid area, as opposed to the northern and intermediate areas whose ombrotypes coincide with upper dry and lower dry. In contrast, an increase in the number of rabbits over the 3-year study period was observed, with intra-annual variations and maximums each year in the month of May. Rabbit abundance and distribution in this region are basically related to diversified landscapes (patches) where the rabbits can find the basic requirements for their survival in a small space (Virgós et al. Citation2003).
Acknowledgements
We thank all the hunting managers for their useful comments and their collaborative attitude. Also, we would like to thank the Consellería de Medio Ambiente, Agua, Urbanismo y Vivienda staff, who have been very helpful. The research was supported by the Ministerio de Educación y Ciencia (CGL2004-00202) and Generalitat Valenciana (GV-04B-732).
References
- Arques J. 2000. Ecología y gestión cinegética de una población de conejos en el sur de la provincia de Alicante. Tesis doctoral. Universidad de Alicante. 323 pp.
- Arques J, Peiro V, Candela MC. 1999. Analyse des facteurs qui conditionnent la saison de reproduction du lapin de garenne dans le sud-est de l’Espagne. Thessaloniki: Proceedings of the International Union of Game Biologists. XXIVth Congress. pp. 474–482.
- Banks PB. 2000. Can foxes regulate rabbit populations? The Journal of Wildlife Management 64:401–406. doi:10.2307/3803238.
- Barrio IC, Acevedo P, Tortosa FS. 2010. Assessment of methods for estimating wild rabbit population abundance in agricultural landscapes. European Journal of Wildlife Research 56:335–340. doi:10.1007/s10344-009-0327-7.
- Belda A, Martinez-Perez JE, Peiro V, Seva E, Arques J. 2011. Main landscape metrics affecting abundance and diversity of game species in a semi-arid agroecosystem in the Mediterranean region. Spanish Journal of Agricultural Research 9:1197–1212. doi:http://dx.doi.org/10.5424/sjar/20110904-470-10.
- Belda A, Munera P, Peiró V, Martínez JE, Seva E. 2012. Abundancia y distribución de especies depredadoras sometidas a control en gestión cinegética: estudio de caso representativo de la urraca (Pica pica) en la comarca de la Marina Baja. Mediterránea. Serie de Estudios Biológicos. Época II 23:32–65.
- Belda A, Peiro V, Seva E, Martin J, Martinez-Perez JE. 2013. Ecological gradients and landscape structure affecting seasonal woodpigeon Columba palumbus densities in a coastal region (South-East Spain). Revie d’Ecologie (La Terre et la Vie) 67:1–13.
- Belda A, Zaragozí B, Martínez-Pérez JE, Peiró V, Ramón A, Seva E, Arques J. 2012. Use of GIS to predict potential distribution areas for wild boar (Sus scrofa Linnaeus 1758) in Mediterranean regions (SE Spain). Italian Journal of Zoology 79:252–265. doi:10.1080/11250003.2011.631944.
- Cabrera-Rodriguez F. 2006. Microhabitat selection of the European rabbit on La Palma, Canary Islands, Spain. Acta Theriologica 51:435–442. doi:10.1007/BF03195190.
- Calvete C, Estrada R, Angulo E, Cabezas-Ruiz S. 2004. Habitat factors related to wild rabbit conservation in an agricultural landscape. Landscape Ecology 19:533–544. doi:10.1023/B:LAND.0000036139.04466.06.
- CMAAUV (Consellería de Medio Ambiente, Agua, Urbanismo y Vivienda) (editor). 2007. Cartografía temática de la Comunidad Valenciana. Available: http://orto.cma.gva.es/website/urbanismo/viewer.htm.
- Fa JE, Sharples CM, Bell DJ. 1999. Habitat correlates of European rabbit (Oryctolagus cuniculus) distribution after the spread of RVHD in Cadiz Province, Spain. Journal of Zoology 249:83–96. doi:10.1111/j.1469-7998.1999.tb01062.x.
- Farina A. 1998. Principles and methods in Landscape Ecology. London: Chapman & Hall. 235 pp.
- Fernández N. 2005. Spatial patterns in European rabbit abundance after a population collapse. Landscape Ecology 20:897–910. doi:10.1007/s10980-004-3976-7.
- Ferrer M, Negro JJ. 2004. The near extinction of two large European predators: Super specialists pay a price. Conservation Biology 18:344–349. doi:10.1111/j.1523-1739.2004.00096.x.
- Ferry C, Frochot B. 1958. Une méthode pour dénombrer les oiseaux nicheurs. Revie d’Ecologie (La Terre et la Vie) 105:85–102.
- Gibb JA, Fitzgerald BM. 1998. Dynamics of sparse rabbits (Oryctolagus cuniculus), Orongorongo Valley, New Zealand. New Zealand Journal of Zoology 25:231–243. doi:10.1080/03014223.1998.9518153.
- INE. 2006. Instituto Nacional de Estadística, Madrid. Available: http://www.ine.es/inebase/cgi. Accessed Jan 2006 18.
- Jiménez-García D. 2007. Paisaje, biodiversidad y gestión sostenible de recursos cinegéticos a escala regional en agrosistemas mediterráneos mediante el uso de tecnologías SIG y GPS. Tesis doctoral. Universidad de Alicante. 274 pp.
- Lazo A. 1992. Socioecología del ganado bovino asilvestrado de la Reserva Biológica de Doñana. Tesis doctoral. Univ. Sevilla. 371 pp.
- Liancourt P, Spence LA, Song DS, Lkhagva A, Sharkhuu A, Boldgiv B, Helliker BR, Petraitis PS, Casper BB. 2013. Plant response to climate change varies with topography interactions with neighbors, and ecotype. Ecology 94:444–453. doi:10.1890/12-0780.1.
- Lombardi L, Fernández N, Moreno S, Villafuerte R. 2003. Habitat-related differences in rabbit (Oryctolagus cuniculus) abundance, distribution, and activity. Journal of Mammalogy 84:26–36. doi:10.1644/1545-1542(2003)084<0026:HRDIRO>2.0.CO;2.
- Martínez-Pérez JE. 2000. Paisajes rurales cambiantes: la amenaza del abandono sobre los espacios montañosos de agricultura tradicional mediterránea. Aplicación del SIG en el estudio de los cambios, de los usos del suelo, en el municipio alicantino de Vall de Gallinera (1956–1998). Tesis Doctoral. Departamento de Ecología. Universidad de Alicante. 350 pp.
- Mas LMA. 1993. Atlas de la Marina Alta. Alicante: Ed. Fundación cultural de la Caja de Ahorros del Mediterráneo. 116 pp.
- Moreno S, Delibes M, Villafuerte R. 1996. Cover is safe during the day but dangerous at night: The use of vegetation by European wild rabbits. Canadian Journal of Zoology 74:1656–1660. doi:10.1139/z96-183.
- Myers K, Parker BS. 1975. Effect of severe drought on rabbit numbers and distribution in a refuge area in semiarid north-western New South Wales. Australian Wildlife Research 2:103–120. doi:10.1071/WR9750103.
- Oliver MA, Webster R. 1990. Kriging: A method of interpolation for geographical information systems. International Journal of Geographical Information Systems 4:313–332. doi:10.1080/02693799008941549.
- Palomares F, Delibes M. 1997. Predation upon European rabbits and their use of open and closed patches in Mediterranean habitats. Oikos 80:407–410. doi:10.2307/3546609.
- Pech RP, Sinclair ARE, Newsome AE, Catling PC. 1992. Limits to predator regulation of rabbits in Australia: Evidence from predator-removal experiments. Oecologia 89:102–112. doi:10.1007/BF00319021.
- Peiro V, Blanc P. 2011. Predicting the spring abundance distribution of red-legged partridge populations in agricultural regions using environmental models and an application for game management. Folia Zoologica 60:203–213.
- Peña J, Bonet A, Bellot J, Sanchez JR, Eisenhuth D, Hallett S, Aledo A. 2007. Driving forces of land-use change in a cultural landscape of Spain. A preliminary assessment of the human-mediated influences. In: Koomen E, Stillwell J, Bakema A, Scholten H, editors. Modelling Land-Use Change. Progress and applications. Dordrecht: Springer. pp. 97–115.
- Pérez-Cueva A. 1994. Atlas climático de la Comunidad Valenciana. Conselleria de Territori i Habitatge. Valencia: Generalitat Valenciana. 206 pp.
- Rivas-Martínez S. 1997. Syntaxonomical sinopsis of the North American natural potential vegetation communities I. Itinera Geobotanica 10:5–148.
- Rivas-Martínez S. 2004. Clasificación bioclimática de la tierra. Available: http://www.ucm.es/info/cif/book/bioc/global_bioclimatics_htm. Madrid: Phytosociological Research Center.
- Rogers PM. 1981. Ecology of the European wild rabbit Oryctolagus cuniculus (L.) in Mediterranean habitats. II. Distribution in the landscape of the Camargue, S. France. The Journal of Applied Ecology 18:355–371. doi:10.2307/2402400.
- Solanas JL, Crespo MB. 2001. Medi físic i flora de la Marina Baixa. Servei de Publicacions Universitat d’Alacant, 420 pp.
- Tizzani P, Menzano A, Dematteis A, Meneguz PG. 2014. Methodological problems related to spotlight count as a census technique for Lepus europaeus in an alpine environment. Acta Theriologica 59:271–276.
- Trout RC, Langton S, Smith GC, Haines RH. 2000. Factors affecting the abundance of rabbits (Oryctolagus cuniculus) in England and Wales. Journal of Zoology 252:227–238. doi:10.1111/j.1469-7998.2000.tb00617.x.
- Villafuerte R, Calvete C, Blanco JC, Lucientes J. 1995. Incidence of viral hemorrhagic disease in wild rabbit populations in Spain. Mammalia 59:651–659. doi:10.1515/mamm.1995.59.4.651.
- Villafuerte R, Lazo A, Moreno S. 1997. Influence of food abundance and quality on rabbit fluctuations: Conservation and management implications in Doñana National Park (SW Spain). Revie d’Ecologie (La Terre et la Vie) 52:345–355.
- Villafuerte R, Viñuela J, Blanco JC. 1998. Extensive predator persecution caused by population crash in a game species: The case of red kites and rabbits in Spain. Biological Conservation 84:181–188. doi:10.1016/S0006-3207(97)00094-3.
- Virgós E, Cabezas-Díaz S, Lozano J. 2007. Is the wild rabbit (Oryctolagus cuniculus) a threatened species in Spain? Sociological constraints in the conservation of species. Biodiversity and Conservation 16:3489–3504. doi:10.1007/s10531-006-9054-5.
- Virgós E, Cabezas-Díaz S, Malo A, Lozano J, López-Huertas D. 2003. Factors shaping European rabbit abundance in continuous and fragmented populations of central Spain. Acta Theriologica 48:113–122. doi:10.1007/BF03194271.
- Willot SJ, Miller AJ, Incoll LD, Compton SG. 2000. The contribution of rabbits (Oryctolagus cuniculus L.) to soil fertility in semi-arid Spain. Biology and Fertility of Soils 31:379–384. doi:10.1007/s003749900183.
- Zaragozí B, Rabasa A, Rodríguez-Sala JJ, Navarro JT, Belda A, Ramón A. 2012. Modelling farmland abandonment: A study combining GIS and data mining techniques. Agriculture, Ecosystems & Environment 155:124–132. doi:http://dx.doi.org/10.1016/j.agee.2012.03.019.