
Abstract
Understanding the at-sea spatial behaviour of sea turtles is a priority for their conservation. In the present paper, the current information on the distribution and movement patterns of the two species breeding in the Mediterranean, the loggerhead and the green turtle, is reviewed, focusing mainly on the 195 published routes of satellite-tracked turtles. A satisfactory level of knowledge about adults’ migrations and other movements is reached only for loggerheads breeding at Zakynthos Island, Greece, and for green and loggerhead females nesting in Cyprus, while studies at foraging grounds are limited to loggerheads in the western and central parts of the basin. Adult males and females mostly show quite uniform post-breeding migratory patterns, typically moving towards individually specific neritic foraging grounds. A much higher variability is shown by loggerhead juveniles, which is probably associated with differences in their spatial behaviour while in oceanic or neritic waters. Evidence of seasonal migrations driven by lower temperatures in winter is available only for adult and juvenile loggerheads frequenting the two northernmost parts of the basin, i.e. the Ligurian and the northern Adriatic Sea. Current knowledge gaps and priorities for further research in the Mediterranean are discussed.
Introduction
Marine turtles are characterised by an itinerant behaviour at all stages of their life cycle. Immediately after hatching, newborn turtles swim quickly away from their natal beaches and begin large-scale movements lasting several years which can extend over entire ocean basins, being strictly dependent on the course of the main ocean current systems (Musick & Limpus Citation1997). In the following juvenile stage, turtles exhibit a variety of movement behaviours associated with their specific feeding needs. Although variable in different turtle species, the juveniles’ spatial behaviour often involves long-range movements, especially during the initial pelagic phase of development, that occur in the oceanic environment (Musick & Limpus Citation1997). As adults, female turtles perform cyclical shuttling migrations between individually specific foraging and breeding areas (Luschi et al. Citation2003; Godley et al. Citation2008), which are probably undertaken by males as well (Hays et al. Citation2010b; Casale et al. Citation2013a).
Understanding the spatial behaviour of marine turtles during their migrations and prolonged movement phases is therefore central to elucidating key aspects of their complex life cycle and ecology. Furthermore, this knowledge is crucial to define appropriate conservation measures for these species of conservation concern, which are listed in the International Union for Conservation of Nature (IUCN) Red List of Threatened Species (IUCN Citation2012) but whose protection status is often unsatisfactory, especially during their at-sea movements (Hamann et al. Citation2010). However, obtaining information on the movements performed by marine animals like sea turtles is not a straightforward task. Turtles spend the vast majority of their life in the water, often frequenting offshore or inaccessible areas, where even approaching or capturing them can be quite difficult. Moreover, they are known to stay submerged for 90–95% of the time (Lutcavage & Lutz Citation1997), and this additionally creates huge technical problems, e.g. to remotely acquire information through radio signals. Sea turtle movements were initially studied through capture-marking-recapture techniques since the 1950s (Hendrickson Citation1958), with the recoveries of marked individuals away from the capture area providing very general information on the extent and course of the movements performed (e.g. Meylan Citation1982). In the last 15–20 years, satellite telemetry has led to substantial progress in the field, as this technique allows the reconstruction of the actual routes followed by marked individuals (Godley et al. Citation2008). This provides quite detailed information on the spatial behaviour of individuals belonging to various life stages, and on the main features of the movements performed by single individuals in their natural environment (see below for further information on these methods).
Adult females represent the class of individuals that have been most studied in their movements, because they can be easily tagged and/or equipped with satellite transmitters during the nesting process and then tracked during the internesting movements between successive egg-layings and the subsequent migrations after the end of the nesting season (Godley et al. Citation2008). In most species, the prevailing migration pattern consists of a quick and direct movement towards specific foraging areas in the neritic environment (e.g. Papi et al. Citation1997; Limpus & Limpus Citation2001), where females stay for long periods feeding on benthic prey and to which they show fidelity in successive reproductive seasons (Broderick et al. Citation2007). The most recent results have, however, revealed numerous exceptions to this general pattern, including the possible prolonged residence of females in the oceanic habitat (Hatase et al. Citation2002; Hawkes et al. Citation2006; Hatase et al. Citation2007; Rees et al. Citation2010), suggesting the possibility of epipelagic feeding in adults (Reich et al. Citation2010), the presence of seasonal migrations between different feeding and wintering areas (Hawkes et al. Citation2007; Zbinden et al. Citation2008, Citation2011) or convoluted segments in the migratory routes (Dodd & Byles Citation2003; Hatase et al. Citation2007; Hawkes et al. Citation2007) of uncertain biological significance.
Information on the movements made by turtles other than adult females (i.e. adult males and juveniles) is limited (Godley et al. Citation2008), although growing. For males, the main difficulty lies in approaching the animals to tag them and deploy the satellite transmitters, since they very rarely leave the aquatic environment (e.g. Godley et al. Citation2008; Arendt et al. Citation2012a; Casale et al. Citation2013a; Schofield et al. Citation2013 and references therein). For juvenile turtles, the best available evidence on their spatial distribution has been obtained from genetic studies that have allowed the estimation of the contribution of different rookeries to the aggregations of juveniles frequenting a certain foraging area (e.g. Bowen & Karl Citation2007), thus providing indirect but valuable information on the distribution and general movement patterns in the juvenile stages. Tracking experiments on juveniles are fewer than those on adults, and are mostly limited to larger turtles foraging in neritic areas, where they usually settle after spending the first few years of their life in oceanic areas (Bolten Citation2003). Very little information is available on the movements of younger turtles during their pelagic developmental phase (but see Kobayashi et al. Citation2008; Howell et al. Citation2010; Mansfield et al. Citation2014), when in fact it is believed that they move over large oceanic areas (Bolten Citation2003).
Three sea turtle species occur in the Mediterranean (Casale & Margaritoulis Citation2010): the leatherback turtle (Dermochelys coriacea Vandelli, 1761), the green turtle (Chelonia mydas Linnaeus, 1758) and the loggerhead turtle (Caretta caretta Linnaeus, 1758). Just a few leatherback turtles enter the Mediterranean from the Atlantic and they can be found all over the basin, although they do not breed in the Mediterranean (Casale et al. Citation2003). The other two species have Mediterranean populations recognised as regional management units, with the green turtle unit being considered at high risk (Wallace et al. Citation2011). The main identified threats at sea to these two Mediterranean populations are incidental catch in fishing gear, collision with boats and intentional killing (Tomás et al. Citation2008; Casale et al. Citation2010; Casale Citation2011), which as a whole represent a high level of threat (Wallace et al. Citation2011). The impact of other potential threats on the populations, like chemical contaminants (D’Ilio et al. Citation2011) and debris (e.g. Tomás et al. Citation2002; Lazar & Gračan Citation2011), is not clear yet.
The loggerhead turtle is the most common species in the Mediterranean Sea, where it nests mainly in the eastern part, from which juveniles then disperse widely throughout the basin (Margaritoulis et al. Citation2003; Casale & Margaritoulis Citation2010; Hays et al. Citation2010a; Casale & Mariani Citation2014). The Mediterranean loggerhead population shows a genetic divergence from the Atlantic populations (Bowen & Karl Citation2007), and a population substructure (Laurent et al. Citation1998; Carreras et al. Citation2007; Clusa et al. Citation2013), indicating the presence of independent management units for conservation. Loggerhead turtles are carnivorous: for the first period of life they mainly feed upon pelagic prey at the sea surface, then they increasingly shift towards benthic prey (Bjorndal Citation1997), although they also feed through the whole water column (e.g. Narazaki et al. Citation2013). The duration of the first strictly epipelagic period varies according to population and oceanographic features, being most extended in those turtles that disperse over wide oceanic regions (Musick & Limpus Citation1997). In the Mediterranean, where neritic (on the continental shelf) and oceanic areas (out of the continental shelf) are not widely separated, loggerhead turtles start feeding upon benthic prey rather early: an analysis of gut and faecal content has shown that benthic feeding occur in turtles as small as 26 cm of curved carapace length (CCL) (Casale et al. Citation2008a).
Green turtles breed exclusively in the Levantine basin (east of Libya and Crete), and primarily in Turkey, Cyprus and Syria (Casale & Margaritoulis Citation2010). At sea, green turtles have been observed mainly in neritic areas in the Levantine basin (e.g. Oruç Citation2001; Oruç et al. Citation2011), but also in Greece and north Africa (e.g. Broderick et al. Citation2007; Margaritoulis & Panagopoulou Citation2010). A limited number of individuals disperse as far as the Ionian, Adriatic and Tyrrhenian Seas (Lazar et al. Citation2004; Bentivegna et al. Citation2011). The Mediterranean green turtle population shows a genetic divergence from the Atlantic populations (Encalada et al. Citation1996), indicating a degree of isolation, and is affected by strong anthropogenic pressures primarily represented by incidental catches in fishery activities (Casale Citation2011). Like loggerheads, green turtles feed upon pelagic prey in the first period of their life but then shift to a herbivorous diet, feeding upon benthic seagrasses and algae (Bjorndal Citation1997). Eventually, large juveniles become fully herbivorous: in the Mediterranean, such a stage is reached at a size over 62 cm CCL (Cardona et al. Citation2010).
Relatively little is known about the spatial behaviour of Mediterranean loggerhead and green turtles: recapture studies and satellite tracking experiments have so far been conducted in a non-systematic manner and the available data is therefore quite fragmentary. However, taking all these findings together, an overall picture is starting to emerge of the movement routes, the habitat use and the most frequented areas of the Mediterranean populations of the two species, providing crucial information for a desirable management of marine areas and representing a research priority for sea turtle conservation (Hamann et al. Citation2010).
The aim of this review is to present the current state of knowledge on sea turtle movement patterns in the Mediterranean and to identify major gaps to be filled by future research. Only data available from peer-reviewed publications or obtained by the authors are considered, to avoid the methodological, interpretative and data-ownership issues associated with non-reviewed sources. Given the scarce occurrence of leatherback turtles in the Mediterranean and the lack of information on their movements, only information about loggerhead and green turtles is considered.
Methods for studying sea turtle movements
The oldest system employed to study sea turtle movements is to apply a tag to one or more flippers of an individual and to wait for someone to recapture it somewhere at a later date. If the turtle is re-encountered, it can then be individually identified thanks to the unique code of the tag (Hendrickson Citation1958; Balazs Citation1999). This method, by its very nature, only gives general indications on the endpoints of the journeys made by tagged turtles. Also, even the simple recapture of a tagged turtle in the vastness of the sea may be a rather improbable event, and the geographic pattern of turtle captures is consequently biased towards the most populated (or most fishery-exploited) areas. In any case, such a capture-mark-recapture method has provided basic but most useful information on the movement patterns and migrations of many sea turtle populations (e.g. Meylan Citation1982) including Mediterranean ones (Margaritoulis et al. Citation2003; Casale et al. Citation2007a).
More recently, satellite telemetry has rapidly integrated and complemented the initial basic information obtainable with tag recoveries. This technique is a viable and quite effective solution to track air-breathing animals that spend at least some seconds at the sea surface, and turtles were among the first animals to be studied in this way (Stoneburner Citation1982; Timko & Kolz Citation1982). Satellite telemetry has become a popular method to study sea turtles, despite its non-negligible costs, and the number of turtles so far tracked worldwide lies in the range of a few thousand at least, with individuals being now routinely tracked for several months on the average, and up to a few years in the best cases.
Essentially, marine animal telemetry via satellite is presently only possible through the Franco-American Argos System, which employs a number of polar-orbiting satellites (six in 2014) located at around 850 km above the Earth’s surface. Upon receiving radio signals coming from special transmitters, called Platform Transmitter Terminals (PTTs), the Argos system is able to localise the animal carrying the PTT anywhere on the Earth’s surface, through a complex computational process based on the Doppler effect of the received signals. Successive signals useful for localisation are spaced in time (with 40–50 seconds minimum between them), and this greatly increases the localisation efficiency in surfacing animals like turtles, which can then be localised when they emerge only briefly, then dive and surface again after a few minutes. The determined locations have different accuracies, depending on various factors (especially on the number of PTT signals received within a single satellite overpass) and Argos itself classifies the fixes obtained in six location classes, with an accuracy ranging from 150 to over 1000 m. Recently, new models of Argos-linked PTTs have been developed also featuring a global positioning system (GPS) receiver; these instruments can acquire GPS ephemeris even during the short surfacings of the turtle (thanks to special technologies allowing fast data acquisition, such as Fastloc; Hazel Citation2009). This information is then part-processed on board, and then relayed in a compressed form to the Argos system (thus using Argos satellites only as a communication channel), which subsequently determine the final GPS position. These units, however, have a higher cost, are heavier and their batteries last much less time than standard PTTs, so that their usage in turtle studies is not widespread.
The PTT models currently available can either be attached to the top of the turtle carapace, commonly by an epoxy resin (e.g. Godley et al. Citation2002), or be towed by the turtle through a tether attached to the posterior part of the carapace (e.g. Sasso & Epperly Citation2007). In any case, PTTs will sooner or later detach from the carapace, although the gluing systems commonly employed guarantee quite long tag attachment durations, not uncommonly well over 1 year (e.g. Casale et al. Citation2013a; Schofield et al. Citation2013). PTTs can have an impact on turtle behaviour and energetics, especially during long movements, and carapace-attached tags have indeed been shown to increase the drag of marine turtles (Watson & Granger Citation1998; Jones et al. Citation2013). This effect is relevant only for relatively small turtles (juveniles), while it is minimal for the large adults. Impacts on other activities (e.g. feeding, mating) seem unlikely. Besides turtle localisations, other information can be obtained through the Argos satellites: for instance, various kinds of sensor data (e.g. water temperature or depths reached) can be collected by the PTT on-board sensors and then transmitted every time the turtle surfaces or when the PTT detaches from the turtle as programmed (“pop-up units”; Swimmer et al. Citation2006). Tracking and sensor data can then be processed and displayed through online platforms such as the Satellite Tracking and Analysis Tool (STAT), originally designed for turtle studies (Coyne & Godley Citation2005).
Other tracking systems such as visual, radio or acoustic tracking can also be used (Lohmann et al. Citation2008), but they require maintaining relatively close contact between the animal and the experimenter, and as such are quite tricky to use on animals moving in the marine environment while staying mostly underwater. Data loggers recording GPS positions are useful only when chances are high to recover tagged individuals after some time, and so their application has basically been limited to females tracked during the period between successive egg depositions, the so-called internesting interval (Schofield et al. Citation2007, Citation2010b, Citation2010c).
Genetic methods can additionally contribute to understanding the general movement patterns of turtle populations, usually over broad spatial scales (e.g. Bowen & Karl Citation2007). This is possible because turtles, and especially females, tend to display a natal homing behaviour, i.e. to breed at natal areas (Bowen & Karl Citation2007), and, as a consequence, the different rookeries tend to differentiate genetically with time and to be characterised by unique DNA sequences or by different proportions of specific DNA sequences. Analysing genetic differences can therefore provide information on the relative contribution of different rookeries to a common foraging ground, rather than on the origin of a single individual (e.g. for the Mediterranean: Carreras et al. Citation2006; Garofalo et al. Citation2013).
Finally, other sources of information can provide clues on turtle distribution, such as stranding data (Tomás et al. Citation2008; Casale et al. Citation2010; Türkozan et al. Citation2013), incidental catch in fishing gears (reviewed by Casale Citation2011) and aerial surveys (Gomez de Segura et al. Citation2006; Lauriano et al. Citation2011).
Adult loggerhead turtles
As for the other turtle species, the majority of the available information about movement patterns of adult loggerheads in the Mediterranean concerns nesting females, obtained either from recoveries of tagged individuals (Margaritoulis et al. Citation2003) or from satellite tracking experiments (summarised in ). This is not surprising, since this class of individuals is the only one that comes ashore for extended periods at predictable times and locations during the long and laborious egg-laying process, thus offering an easy way for researchers to approach and tag or equip them with electronic instruments.
Table I. Summary of the 195 loggerhead and green turtles tracked by satellite in the Mediterranean (data presented in published papers, up to 2013).
Adult females in the Mediterranean Sea have been typically tracked during their postnesting migrations, having been equipped with satellite transmitters while at their nesting beach in the final part of the nesting season so to reduce the risk of losing the tag during mating. In a few cases, females have also been tagged early in the season to be tracked also during the internesting intervals, showing movements in the close vicinity of the nesting beach (Hays et al. Citation1991; Godley et al. Citation2003; Schofield et al. Citation2007, Citation2010b, Citation2010c; Zbinden et al. Citation2007; Fuller et al. Citation2008). In Greece, some females have, however, been shown to make excursions away from the nesting area during the internesting interval, with a turtle making a 200-km return trip (Schofield et al. Citation2010b, Citation2010c).
Nearly all postnesting routes tracked so far in Mediterranean loggerheads conform to the most common migratory pattern known in loggerheads, i.e. a direct and rather quick migration away from the breeding site to reach a specific feeding area, usually located in the neritic environment, where the turtle is then localised for months (type A pattern in Godley et al. Citation2008). The most information is available for turtles belonging to the main rookery of Zakynthos Island, in western Greece (), with a total of 36 females having been tracked so far (Zbinden et al. Citation2008, Citation2011; Schofield et al. Citation2013). They have been shown to migrate towards two main destinations in the central Mediterranean (), the Adriatic Sea (especially in its northern portion) and the continental shelf area offshore Tunisia and Libya (often, but rather improperly, indicated as the Gulf of Gabès, which actually only constitutes a small part of this area). These findings confirmed the information obtained through recoveries of turtles tagged in western Greece (Margaritoulis et al. Citation2003). Tracked Zakynthos turtles were distributed about roughly equally between these two foraging areas (Zbinden et al. Citation2011; Schofield et al. Citation2013), which indeed represent the largest neritic areas suitable for turtle foraging in the whole Mediterranean (Schofield et al. Citation2013). Some of the turtles foraging in the northern Adriatic displayed seasonal migrations moving towards lower latitudes during the winter months (Zbinden et al. Citation2011). In a few cases, turtles were tracked for long enough to reconstruct also their movements back to the breeding areas (Zbinden et al. Citation2008; Hays et al. Citation2010b; Schofield et al. Citation2010a). This revealed valuable details of the poorly known pre-breeding migratory phenology of Mediterranean loggerheads. This information is particularly valuable since some migratory features (e.g. timing of pre-breeding migrations) might be flexible and controlled by the environment (e.g. by sea temperatures; Mazaris et al. Citation2013).
Figure 1. General migratory routes of the 63 adult loggerhead turtles satellite tracked in the Mediterranean () and showing type A movement pattern (Godley et al. Citation2008). Only groups of ≥ 10 animals released from the same site are considered, while releases of single individuals () are not shown. Arrows represent the approximate, general courses and do not show the actual reconstructed routes. Circles represent the release breeding sites in Greece (32 females and 23 males) and Cyprus (eight females; two turtles remained along the Cyprus coast). Number of individual females (F) and males (M) reaching different destinations are shown in boxes. The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; CY, Cyprus; EG, Egypt; GR, Greece; HR, Croatia; IL, Israel; IT, Italy; LB, Lebanon; LY, Libya; ME, Montenegro; SI, Slovenia; SY, Syria; TN, Tunisia; TR, Turkey.
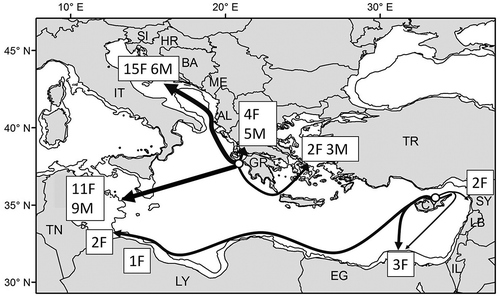
Some Zakynthos turtles, however, migrated towards other sites such as the close-by Amvrakikos Gulf in western Greece, the Aegean Sea and the coasts of Peloponnese, eastern and northern Greece (). Females breeding in Cyprus displayed much the same type A movement pattern, migrating towards more disparate foraging areas along the North African shelf or the Syrian coast or even remaining in Cyprus (Godley et al. Citation2003; Broderick et al. Citation2007) (). In both cases, it is not to be excluded that other foraging sites for these populations may be discovered in the future, given that the number of tracked females is still quite low with respect to the number of females breeding in these rookeries (Schofield et al. Citation2013). Information on the migrations of turtles from the other Mediterranean rookeries is limited and mostly unpublished. A single female migrated from Rethymno, northern Crete, to Mykonos Island in the Aegean Sea, from where she then made a southward seasonal movement frequenting other islands during the winter before returning to Mykonos the following spring (Margaritoulis & Rees Citation2011). Turtles nesting in Calabria, southern Italy, have been shown to migrate directly towards the Tunisian shelf, again following the type A pattern (T. Mingozzi and P. Luschi, unpublished data).
The other postnesting migratory pattern known in loggerheads, that of staying in offshore oceanic waters moving continuously over more or less extended areas (type B pattern in Godley et al. Citation2008), is much less represented in Mediterranean turtles, having so far been shown in only a few turtles tracked from Zakynthos (Zbinden et al. Citation2008; Hays et al. Citation2010b; Schofield et al. Citation2013). For example, a female tracked by Zbinden et al. (Citation2008) stayed on the move for the 189 days of tracking while visiting various areas of the Adriatic and Ionian sea, apparently without heading towards a specific destination (but slowing down its movement rate while in the Northern Adriatic) ().
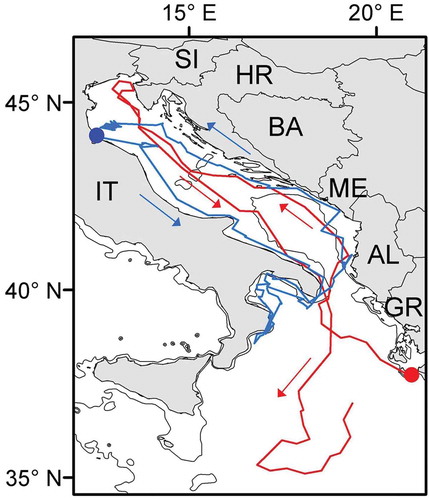
Figure 2. Seasonal migrations in Mediterranean loggerheads. Blue track: migration of a rehabilitated juvenile turtle that left the release site in the North Adriatic (blue dot) in autumn 2007, remained in the Ionian Sea during winter and early spring and returned to northern Adriatic in May 2008 (from Casale et al. Citation2012a). Red track: seasonal migration of an adult female that moved to the northern Adriatic after completing nesting in Zakynthos island (red dot) and then migrated south during winter (from Zbinden et al. Citation2008). Arrows indicate direction of movement. The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; GR, Greece; HR, Croatia; IT, Italy; ME, Montenegro; SI, Slovenia. (Note: the colour version of this Figure is available only in the online edition of this journal).
Quite a large number of adult males have been tracked in the Mediterranean, although to a slightly lesser extent than females (). Males breeding at Zakynthos have been captured while in coastal waters and tracked either during the breeding season or during the successive post-breeding migrations (Schofield et al. Citation2010a, Citation2013). They showed a pattern fully correspondent to that of females (), as they (i) remained in the waters close to the nesting site during the breeding period, which, however, lasted a few days only (Schofield et al. Citation2010b, Citation2010c); (ii) mostly migrated away from Zakynthos towards disparate feeding grounds predominantly located in the Adriatic or on the Tunisian shelf, as well as along the Greek coasts and in Western Turkey, with a few turtles foraging in oceanic sites (Schofield et al. Citation2010a, Citation2013); (iii) displayed a fidelity to the same foraging sites in successive years, often showing a remarkable consistency in the migratory route and an accurate homing behaviour (Schofield et al. Citation2010a); (iv) undertook seasonal migrations when foraging in the Northern Adriatic Sea (Hays et al. Citation2010a, Citation2010b; Schofield et al. Citation2010a). Males were, however, found to remain more often than females in Zakynthos and nearby areas, without embarking on migrations (Schofield et al. Citation2010a, Citation2013). Adult males have also been tracked from their foraging area in the Tunisian shelf area after having been captured by fisheries (Casale et al. Citation2013a). The reconstructed routes showed a combination of short-range foraging movements and of shuttling migrations towards breeding sites along the Libyan coast and even in Greece. In two cases, a strong fidelity to foraging sites on the Tunisian shelf and to a possible breeding site in Libya was revealed, although the four males that were tracked during the complete return migrations followed different routes during the pre- and post-breeding movements. A protracted residency in specific foraging sites, likely indicative of site fidelity, has also been shown in males and females frequenting the enclosed basin of Amvrakikos Gulf, Western Greece (Rees et al. Citation2013).
Finally, a few adult-sized turtles (CCL between 68 and 87 cm) have been tracked upon release after a rehabilitation period in turtle rescue centres in Italy (Bentivegna Citation2002; Mencacci et al. Citation2006; Casale et al. Citation2012a; Luschi et al. Citation2013). These turtles displayed a variety of movements, including seasonal migrations over large spatial scales, often made independently and even against main surface currents (Bentivegna et al. Citation2007). The scarcity of information about the specific individuals prevents us from precisely framing their movements within the turtles’ life cycle, and in only two cases could the reconstructed movements be clearly identified as pre-nesting migrations towards a nesting site (Luschi et al. Citation2013). In many cases, turtles were found to wander over large distances while frequenting different, disparate areas during the non-breeding period: such a lack of a prolonged permanence in a fixed area may indicate that these turtles too displayed a type B movement pattern (Godley et al. Citation2008).
Juvenile loggerhead turtles
Small juveniles: the “lost years”
If movements and distribution of wide-ranging marine species like sea turtles are generally difficult to study, this is particularly true for their first period of life, the so-called “lost-years” (Carr & Meylan Citation1980), when individuals are small, usually live in open waters and are extremely difficult to observe. The Mediterranean turtles are no exception. The various empirical methods employed to study turtle distribution and movements in Mediterranean waters usually observe adults or large juveniles, thus leaving out loggerheads smaller than 40 cm CCL, roughly corresponding to the first seven years of life (Casale et al. Citation2009, Citation2011a, Citation2011b). The only method available to investigate their movement patterns is through simulations of particle dispersals in large-scale oceanographic models (e.g. Hays et al. Citation2010a), although promising new tracking techniques for small turtles are being developed (Mansfield et al. Citation2014).
In the Mediterranean, recent simulations of hatchling dispersal have indicated that hatchlings from the Levantine and central nesting sites are mainly retained in the same areas of origin, while those from the Ionian region disperse widely to the Ionian, Adriatic and south-central Mediterranean areas (Hays et al. Citation2010a; Casale & Mariani Citation2014).
Large juveniles
The numbers of satellite-tracked large juveniles (> 40 cm CCL) in the Mediterranean and the study areas are summarised in . These turtles showed a large variety of movements. Some just wandered across wide areas, with no sign of residence at any specific site (Bentivegna Citation2002; Casale et al. Citation2007a; Eckert et al. Citation2008; Cardona et al. Citation2009; Hochscheid et al. Citation2010) (). Others frequented somewhat smaller areas (Cardona et al. Citation2005; Revelles et al. Citation2007a, Citation2007b; Eckert et al. Citation2008; Cardona et al. Citation2009; Hochscheid et al. Citation2010; Casale et al. Citation2012a, Citation2012b) (), in some cases remaining for a long time in very restricted sites (Hochscheid et al. Citation2010; Mencacci et al. Citation2011). The areas frequented and the routes followed are the result of a combination of surface circulation patterns and the turtles’ active swimming (Bentivegna et al. Citation2007; Revelles et al. Citation2007b).
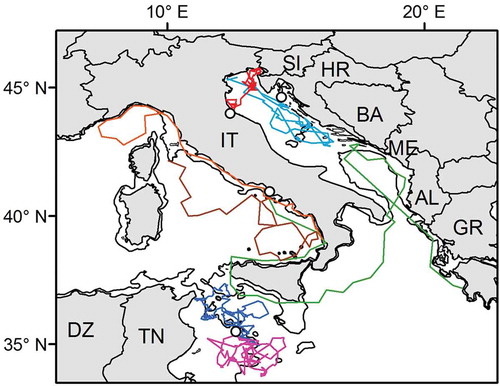
Figure 3. Examples of variable movements in juvenile loggerheads in the Mediterranean. Turtles released from southwestern Italy performed long-distance erratic movements over large oceanic areas (brown, orange and green tracks; Hochscheid et al. Citation2010). Conversely, turtles released from the Tunisian (fuchsia and dark blue; Casale et al. Citation2012b) and Adriatic shelves (red and light blue; Casale et al. Citation2012a) displayed convoluted movement over more restricted areas. Release sites are indicated by open circles. The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; DZ, Algeria; GR, Greece; HR, Croatia; IT, Italy; ME, Montenegro; SI, Slovenia; TN, Tunisia. (Note: the colour version of this Figure is available only in the online edition of this journal).
Juveniles frequenting oceanic areas may either show fidelity to an area or move across multiple areas. A degree of permanence in the same area was indicated by tag returns in the western and eastern Mediterranean (Casale et al. Citation2007a), and was directly observed through satellite tracking in some cases in the western Mediterranean (Revelles et al. Citation2007a) but not in others, either in the western (Cardona et al. Citation2005; Eckert et al. Citation2008; Cardona et al. Citation2009) or in the eastern Mediterranean (Bentivegna Citation2002). In general, resident areas of juveniles in oceanic zones are much wider than in neritic zones (Revelles et al. Citation2007b; Cardona et al. Citation2009; Casale et al. Citation2012a, Citation2012b) (), as also observed for adults in the Mediterranean and elsewhere (Hawkes et al. Citation2006; Schofield et al. Citation2010a). Long-term residence of juveniles in neritic foraging grounds seems more common than in oceanic areas and it has been observed in various locations through tag returns (Casale et al. Citation2007a; Revelles et al. Citation2008) and satellite tracking (Hochscheid et al. Citation2007; Cardona et al. Citation2009; Hochscheid et al. Citation2010; Casale et al. Citation2012a, Citation2012b; Hochscheid et al. Citation2013) (). The juvenile neritic foraging sites are, however, usually larger than the adult neritic sites, with juveniles showing a propensity to wander over quite large areas (e.g. Casale et al. Citation2012b) rather than to settle in a given location as adults usually do (e.g. Schofield et al. Citation2010a; Casale et al. Citation2013a).
Figure 4. Schematic examples of the areas frequented by satellite-tracked juvenile loggerheads in the Mediterranean. All turtles considered were tracked for a minimum of 120 days. The areas frequented by juveniles in the oceanic zone north of Algeria (a; Revelles et al. Citation2007b; Cardona et al. Citation2009) tend to be larger than areas of juveniles in the neritic zones along the Spanish coast (b; Cardona et al. Citation2009), the Tunisian (c; Casale et al. Citation2012b) and Adriatic shelves (d; Casale et al. Citation2012a), and the western Italian coast (e; the arrows indicate the areas frequented by two turtles; Mencacci et al. Citation2011). The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; DZ, Algeria; FR, France; GR, Greece; HR, Croatia; IT, Italy; LY, Libya; MA, Morocco; ME, Montenegro; SI, Slovenia; SP, Spain; TN, Tunisia.
The behavioural response of juvenile turtles to seasons varies per location and per individual. In some areas, like the western Mediterranean (Revelles et al. Citation2007b) and the Tunisian shelf (Casale et al. Citation2012b), no seasonal movement patterns were observed through satellite tracking. In contrast, individuals foraging in the north Adriatic have been found to move southwards in winter, as observed also in adults (see above), and in the Ligurian Sea (Lauriano et al. Citation2011). For instance, Casale et al. (Citation2012a) tracked one individual during a complete migration cycle, with an autumn southwards migration along the Italian coast in the western Adriatic and a northwards spring return to the very same site following a route along the eastern Adriatic coast (). However, the same study also showed that other individuals remained in the northernmost and coldest part of the basin during the whole winter (Casale et al. Citation2012a). Actually, there is evidence from bycatch (Casale et al. Citation2004) and stranding records (Casale et al. Citation2010) that at least some loggerhead turtles, mainly juveniles, frequent the wider north Adriatic area also in winter. Indeed, Mediterranean loggerhead turtles have been observed to maintain some level of activity at temperatures as low as 11.8°C, adopting a specific behavioural strategy known as overwintering (Hochscheid et al. Citation2007). Therefore, it seems that cold winter temperatures induce only some of the turtles to move southwards, and only from the two northernmost areas (), i.e. the north Adriatic (Casale et al. Citation2012a and references therein) and the Ligurian Sea (Lauriano et al. Citation2011), where temperatures drop below 13°C in winter.
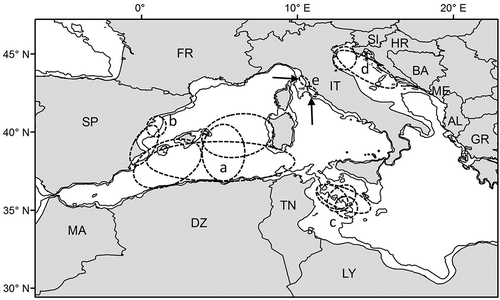
Figure 5. Specific cases of distribution and movements of loggerhead turtles in the Mediterranean. The main movement routes and the areas possibly visited by Atlantic individuals entering the Mediterranean are shown by grey arrows. Black arrows indicate the course of the possible seasonal movements from the two northernmost and coldest areas of the Mediterranean, the Ligurian Sea (a; only juveniles) and the north Adriatic (b; both adults and juveniles). The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; DZ, Algeria; FR, France; GR, Greece; HR, Croatia; IT, Italy; LY, Libya; MA, Morocco; ME, Montenegro; SI, Slovenia; SP, Spain; TN, Tunisia.
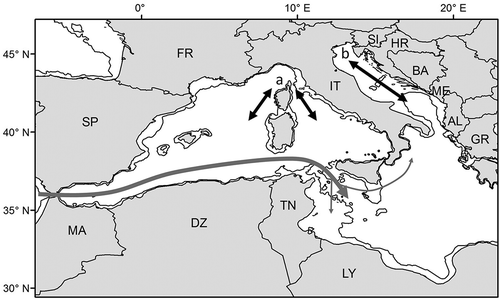
In the western Mediterranean, tag and genetic data indicate that turtle movements are affected by the prevailing currents that define two main “water masses”: the northern and southern parts of the basin (Carreras et al. Citation2006; Revelles et al. Citation2008). It has been suggested that the Atlantic individuals entering from the Strait of Gibraltar (see below) frequent the southern part of the western basin, while Mediterranean loggerheads frequent the northern part (Carreras et al. Citation2006).
Atlantic loggerheads in the Mediterranean
Genetic analyses on mtDNA have shown that the Mediterranean is also frequented by loggerheads originating from the Atlantic Ocean. These turtles derive from western Atlantic rookeries (Carreras et al. Citation2006), and reach the eastern side during their long-distance developmental migrations, helped by the Gulf Stream and related oceanic currents (Musick & Limpus Citation1997). The modal size of loggerheads found on the Atlantic coast of northern Europe is about 23 cm CCL (20.5 cm straight carapace length, Hays & Marsh Citation1997; converted according to Bjorndal et al. Citation2000), suggesting that Atlantic turtles enter the Mediterranean at this size or larger. The smallest turtle found in the eastern Mediterranean and carrying an mtDNA haplotype endemic to the western Atlantic was in the size range 30–34 cm CCL (Laurent et al. Citation1998). Genetic markers have shown that Atlantic loggerheads occur in high numbers, especially in the oceanic zone of the westernmost part of the Mediterranean basin, closer to the Strait of Gibraltar, through which they enter from the Atlantic () (Carreras et al. Citation2006). However, individuals tagged in the Atlantic have been found in the eastern Mediterranean as well, as far as the Sicily Channel (Bolten et al. Citation1992) and the north Ionian Sea (Manzella et al. Citation1988). Genetic studies too have indicated high proportions of Atlantic individuals in the oceanic zone of the Sicily Channel (Laurent et al. Citation1998) as well as their occurrence in the neritic zone of the Tunisian continental shelf (), though at a lower proportion (Casale et al. Citation2008b). Current data suggests that Atlantic turtles do not enter the Adriatic or do so in very low numbers (Giovannotti et al. Citation2010; Garofalo et al. Citation2013). The occurrence of Atlantic loggerheads in other parts of the eastern Mediterranean basin is possible but has not yet been adequately assessed through genetic surveys.
Since genetic flow from the Atlantic to the Mediterranean is estimated to be low, these Atlantic turtles are thought not to breed in the Mediterranean but to return to the Atlantic to breed (Carreras et al. Citation2011). This hypothesis is corroborated by a few empirical findings: one loggerhead tagged in the western Mediterranean (Spain) was found in Cuba (Moncada et al. Citation2010), one turtle tagged in the eastern Mediterranean (north Ionian Sea) was found on the coast of Portugal, just outside the Mediterranean (Argano et al. Citation1992), and one turtle tagged in the central Mediterranean was found in North America (Casale et al. Citation2013b). Moreover, three large juveniles were satellite tracked during their transatlantic migrations starting from the Mediterranean side of the Strait of Gibraltar: two of them stopped transmissions in the middle of the North Atlantic while the third one reached Nicaragua (Eckert et al. Citation2008). The occurrence of Atlantic loggerheads in the Mediterranean represents a good opportunity to understand the species’ behavioural patterns associated with fidelity to different habitats at different life stages, and information about backward migration to the Atlantic basin can contribute to understanding these patterns.
Green turtles
The current knowledge on the distribution and movements of green turtles in the Mediterranean is very limited in comparison to that for loggerhead turtles. As with loggerheads, information on the distribution of small turtles is particularly deficient, but in green turtles this applies to larger juveniles as well, given that no tracking experiment has been ever undertaken. Hatchling dispersal simulations indicated that green turtle hatchlings are mainly retained in the Levantine basin, where the nesting sites are (Casale & Mariani Citation2014). A high proportion of green turtles smaller than 30 cm CCL was reported to occur in southern Turkey waters (Türkozan et al. Citation2013) while low numbers of small turtles < 40 cm CCL frequent the Adriatic (Lazar et al. Citation2004), especially in the southern part, the Ionian and even the Tyrrhenian seas around Italy (Bentivegna et al. Citation2011). Larger juveniles and adults are mainly found in the Levantine zone (Casale & Margaritoulis Citation2010; Nada & Casale Citation2011; Türkozan et al. Citation2013), with foraging habitats having been identified also in the Aegean and Ionian (south Peloponnesus, Greece) Seas (Margaritoulis & Panagopoulou Citation2010), as well as in Libyan and, to a lesser extent, in Tunisian waters (Broderick et al. Citation2007; Bradai & Jribi Citation2010; Hamza Citation2010; Turkecan & Yerli Citation2011).
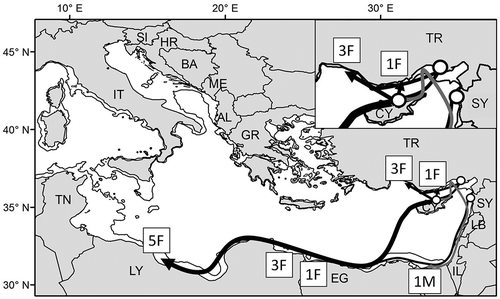
A total of 14 green turtles have been satellite tracked in the Mediterranean (). Thirteen adult females have been monitored from nesting sites in Turkey (Turkecan & Yerli Citation2011; n = 2), Cyprus (Broderick et al. Citation2007; n = 10) and Syria (Rees et al. Citation2008; n = 1). They all followed a type A pattern (Godley et al. Citation2008), performing a direct and rather quick migration away from the breeding site to reach individually specific neritic foraging areas in Libya, Turkey and Egypt (). These data indicate that the north African coast of Egypt and Libya is an important migratory corridor for green turtles (Broderick et al. Citation2007). Cyprus females tracked in different years showed a remarkable fidelity to the same foraging grounds in Libya (Broderick et al. Citation2007). No information on the internesting movements performed is available, unfortunately. Finally, one adult male was tracked from Cyprus and showed a non-direct route to Egypt along the coast, which was interpreted as an evidence of multiple breeding areas (Wright et al. Citation2012) ().
Figure 6. Migratory routes of 13 adult green turtles satellite tracked in the Mediterranean (). Arrows represent the approximate, general courses and do not show the actual reconstructed courses. Routes of females and male are represented by black and grey lines respectively. Circles indicate the release sites of 10 females and one male (Cyprus), two females (Turkey) and one female (Syria). The area of Turkey, Syria and Cyprus is shown enlarged in the top right box. Number of individual females (F) and males (M) reaching different destinations are shown in boxes. The 200 m isobath is shown. AL, Albania; BA, Bosnia and Herzegovina; CY, Cyprus; EG, Egypt; GR, Greece; HR, Croatia; IL, Israel; IT, Italy; LB, Lebanon; LY, Libya; ME, Montenegro; SI, Slovenia; SY, Syria; TN, Tunisia; TR, Turkey.
Discussion
General considerations
Despite the large knowledge gaps still remaining, the available information reviewed above allows us to depict an overall picture of the spatial behaviour of Mediterranean sea turtles, which can be compared with what is known on turtles of other areas. Overall, adult females display a rather uniform migratory behaviour, especially in green turtles, as they leave the nesting beach to quickly move towards discrete foraging zones (type A pattern; Godley et al. Citation2008). These sites are often located quite far away from the breeding area, and are in neritic (frequently coastal) waters, in line with the tendency of both species to feed primarily on algae, seagrass or benthic prey (Bjorndal Citation1997). While the geographic course and extension of these migrations can be quite variable even within the same rookery (e.g. Schofield et al. Citation2013), the overall behavioural pattern is basically the same, and is shared by loggerhead males as well. This migratory behaviour is fully correspondent to that observed in other areas in female loggerhead (e.g. Papi et al. Citation1997; Blumenthal et al. Citation2006; Hawkes et al. Citation2007; Girard et al. Citation2009) and green turtles (e.g. Luschi et al. Citation1996; Cheng Citation2000; Craig et al. Citation2004; Seminoff et al. Citation2008), as are the small-scale internesting movements performed by loggerheads, also typically recorded elsewhere (Stoneburner Citation1982; Limpus & Limpus Citation2001). Given that in the Mediterranean the largest extension of shallow areas offering suitable neritic foraging sites is in the northern Adriatic and the Libyan/Tunisian shelf, these represent key areas for both Mediterranean green and loggerhead turtles, given that both areas are frequented by adults and juveniles. Other factors may, however, play a role in determining the movement patterns observed: for instance, recent evidence suggests that the foraging locations of adult turtles might be influenced by their drift scenarios as hatchlings (Hays et al. Citation2010a; Scott et al. Citationin press), and this would explain, for example, why few adults forage in the western part of Mediterranean, given that rookeries are confined to the Eastern basin (see also Casale & Mariani Citation2014). Mediterranean adult loggerheads do, however, exhibit some variety in their movements, with some turtles foraging in offshore oceanic waters for at least part of their inter-reproductive phase, sometimes keeping on the move without fixing to any specific site (Zbinden et al. Citation2008; Luschi et al. Citation2013; Schofield et al. Citation2013). This pattern too is in general accordance with recent findings obtained elsewhere showing how loggerhead spatial behaviour is actually characterised by a considerable complexity (Hatase et al. Citation2002; Hawkes et al. Citation2006, Citation2007; Rees et al. Citation2010). This variability is likely linked to the loggerhead’s plastic and opportunistic foraging habits (McClellan et al. Citation2010; Reich et al. Citation2010; Narazaki et al. Citation2013), which allow them to exploit both neritic and oceanic foraging grounds.
The variety in spatial patterns reaches its peak in the case of Mediterranean loggerhead juveniles, which have been found to perform diverse movement patterns such as erratic movements over pelagic or neritic areas, prolonged stays in restricted zones or even long-distance migrations, frequently with a seasonal periodicity. Once again, this is in accordance with tracking data obtained on other loggerhead populations outside the Mediterranean (McClellan & Read Citation2007; Kobayashi et al. Citation2008; Mansfield et al. Citation2009; Arendt et al. Citation2012b). Based on juvenile foraging ecology (Casale et al. Citation2008a) and on the difference in size of the residential oceanic and neritic areas, an ecological-behavioural model has been recently put forward (Casale et al. Citation2012b). The model proposes a gradual rather than an abrupt shift from a pelagic-vagile to a benthic-sedentary life style leading to a progressive reduction of home ranges. This reduction in home ranges may be due to the higher availability and energy content of the benthic prey accessible in the neritic zone with respect to the epipelagic prey that are the only ones available to loggerheads in the oceanic environment. This model has been developed especially for loggerheads living in the Mediterranean Sea, whose oceanographic features, with a great extent of oceanic-neritic edges, would produce an “edge effect” that may favour opportunistic feeding, early frequentation, settlement and residence in neritic feeding grounds. However, such a non-rigid, “relaxed” model could be well applied to loggerheads of other areas as well; actually, the “classic” models of a loggerhead life cycle with specific ontogenetic stages (Musick & Limpus Citation1997; Bolten Citation2003) probably represent just specific cases of a greater behavioural plasticity.
Research priorities
Only a few of the 42 major sea turtle nesting sites in the Mediterranean (Casale & Margaritoulis Citation2010) have been investigated regarding their connections with the marine areas, which is where turtles originating from specific nesting sites spend the vast majority of their time. Due to the well-known phenomenon of sea turtle natal homing, in particular of females (Bowen & Karl Citation2007), each breeding site should be regarded as an independent management unit (Moritz Citation1994; Bowen et al. Citation2005), i.e. the fate of one nesting site has few or no consequences on other nesting sites. In such a context, one research priority is to assess the localisation of the specific foraging areas for each major nesting site. Genetic markers can provide rough patterns of distribution from rookery aggregates (Garofalo et al. Citation2013; Clusa et al. Citation2014). However, this approach is made difficult by the relatively poor genetic characterisation of Mediterranean rookeries (i.e. the presence of shared genetic markers), partly due to the recent colonisation of the Mediterranean Sea by loggerheads (Clusa et al. Citation2013), the gaps in the sampled rookeries and other intrinsic methodological limits, which prevent a complete assessment of the dispersal and migratory patterns of individual rookeries. Satellite tracking, although intrinsically limited in sample size, can be helpful on this aspect and can also identify migratory connectivity, including the routes followed by single individuals. So far, however, satellite tracking experiments on Mediterranean loggerheads have been conducted in a non-systematic manner usually with a few individuals tracked each time (), thus limiting the significance of the findings obtained. Two complementary approaches can be used to overcome this limitation: tracking a large number of adults from one breeding site to identify multiple foraging areas (e.g. Schofield et al. Citation2013) and tracking adults from one foraging area to identify multiple breeding sites (e.g. Casale et al. Citation2013a). This strategy should not be limited to females, but more attention and effort should be given to adult males as well. Considering the previous studies (), the next priority nesting areas should be eastern Greece, Turkey, Libya and Syria.
Also, it has to be considered that sea turtle populations are made up mostly of juveniles; thus, anthropogenic threats at sea, mainly fishing, mostly affect the juvenile class, and the consequent reduction of survival rate (e.g. Casale et al. Citation2007b; Casale et al. in press) can be detrimental for the population (Heppell Citation1998). Hence, a comprehensive knowledge of the distribution of juveniles is fundamental for implementing sound conservation measures. In order to identify important oceanic and neritic foraging grounds, satellite tracking studies should complement other sources of information, such as stranding and bycatch records, which may be intrinsically biased. A priority goal should be identifying movement patterns and neritic foraging areas of juveniles of both species, especially in the Levantine basin (e.g. Turkey, Egypt, Cyprus and Libya), for which information is particularly lacking ().
Finally, it should be noted that many unpublished tracks do exist in addition to those reviewed here. For instance, we estimate that roughly 140 Mediterranean tracks of those available on seaturtle.org’s “Satellite tracking” service as of April 2014 have not been included in any peer-reviewed paper. It is therefore to be expected that an amount of information nearly as great as that reviewed here still awaits proper publication. This represents a missed opportunity to improve the overall picture of Mediterranean turtle spatial behaviour and may even cause incidental redundancy and waste of resources when planning future research. We therefore recommend the prompt publication of satellite tracking data in peer-reviewed journals.
Acknowledgements
Resi Mencacci helped greatly with comments on initial versions of the manuscript and with the preparation of figures. We thank Graeme Hays and two anonymous reviewers for their valuable comments on a previous version of this paper.
References
- Arendt MD, Segars AL, Byrd JI, Boynton J, Schwenter JA, Whitaker JD, Parker L. 2012a. Migration, distribution, and diving behavior of adult male loggerhead sea turtles (Caretta caretta) following dispersal from a major breeding aggregation in the Western North Atlantic. Marine Biology 159:113–125. doi:10.1007/s00227-011-1826-0.
- Arendt MD, Segars AL, Byrd JI, Boynton J, Whitaker JD, Parker L, Owens DW, Blanvillain G, Quattro JM, Roberts MA. 2012b. Seasonal distribution patterns of juvenile loggerhead sea turtles (Caretta caretta) following capture from a shipping channel in the Northwest Atlantic Ocean. Marine Biology 159:127–139. doi:10.1007/s00227-011-1829-x.
- Argano R, Basso R, Cocco M, Gerosa G. 1992. New data on loggerhead (Caretta caretta) movements within Mediterranean. Bollettino del Museo dell’Ististuto di Biologia dell’Università di Genova 56-57:137–163.
- Balazs GH. 1999. Factors to consider in the tagging of sea turtles. In: Eckert KL, Bjorndal KA, Abreu-Grobois FA, Donnelly M, editors. Research and management techniques for the conservation of sea turtles, Gland: IUCN/SSC Marine Turtle Specialist Group Publication No. 4. pp. 101–109.
- Bentivegna F, Ciampa M, Hochscheid S. 2011. The presence of the green turtle, Chelonia mydas, in Italian coastal waters during the last two decades. Marine Turtle Newsletter 131:41–46.
- Bentivegna F, Valentino F, Falco P, Zambianchi E, Hochscheid S. 2007. The relationship between loggerhead turtle (Caretta caretta) movement patterns and Mediterranean currents. Marine Biology 151:1605–1614. doi:10.1007/s00227-006-0600-1.
- Bjorndal KA. 1997. Foraging ecology and nutrition of sea turtles. In: Lutz PL, Musick JA, editors. The biology of sea turtles, Boca Raton, FL: CRC Marine Science Series, CRC Press. pp. 199–231.
- Bjorndal KA, Bolten AB, Martins HR. 2000. Somatic growth model of juvenile loggerhead sea turtles Caretta caretta: Duration of pelagic stage. Marine Ecology-Progress Series 202:265–272. doi:10.3354/meps202265.
- Blumenthal JM, Solomon JL, Bell CD, Austin TJ, Ebanks-Petrie G, Coyne MS, Broderick AC, Godley BJ. 2006. Satellite tracking highlights the need for international cooperation in marine turtle management. Endangered Species Research 2:51–61. doi:10.3354/esr002051.
- Bolten AB. 2003. Variation in sea turtle life history patterns: Neritic vs. oceanic developmental stages. In: Lutz PL, Musick JA, Wyneken J, editors. The biology of sea turtles. Volume II. Boca Raton, London, New York, Washington D.C: CRC Press. pp. 243–257.
- Bolten AB, Martins HR, Bjorndal KA, Cocco M, Gerosa G. 1992. Caretta caretta (loggerhead) pelagic movement and growth. Herpetological Review 23:116.
- Bowen BW, Bass AL, Soares L, Toonen RJ. 2005. Conservation implications of complex population structure: Lessons from the loggerhead turtle (Caretta caretta). Molecular Ecology 14:2389–2402. doi:10.1111/j.1365-294X.2005.02598.x.
- Bowen BW, Karl SA. 2007. Population genetics and phylogeography of sea turtles. Molecular Ecology 16:4886–4907. doi:10.1111/j.1365-294X.2007.03542.x.
- Bradai MN, Jribi I. 2010. Tunisia. In: Casale P, Margaritoulis D, editors. Sea turtles in the Mediterranean: Distribution, threats and conservation priorities, Gland, Switzerland: IUCN. pp. 245–256.
- Broderick AC, Coyne MS, Fuller WJ, Glen F, Godley BJ. 2007. Fidelity and over-wintering of sea turtles. Proceedings of the Royal Society B-Biological Sciences 274:1533–1538.
- Cardona L, Campos P, Levy Y, Demetropoulos A, Margaritoulis D. 2010. Asynchrony between dietary and nutritional shifts during the ontogeny of green turtles (Chelonia mydas) in the Mediterranean. Journal of Experimental Marine Biology and Ecology 393:83–89. doi:10.1016/j.jembe.2010.07.004.
- Cardona L, Fernández G, Revelles M, Aguilar A. 2012. Readaptation to the wild of rehabilitated loggerhead sea turtles (Caretta caretta) assessed by satellite telemetry. Aquatic Conservation: Marine and Freshwater Ecosystems 22:104–112. doi:10.1002/aqc.1242.
- Cardona L, Revelles M, Carreras C, SanFélix M, Gazo M, Aguilar A. 2005. Western Mediterranean immature loggerhead turtles: Habitat use in spring and summer assessed through satellite tracking and aerial surveys. Marine Biology 147:583–591. doi:10.1007/s00227-005-1578-9.
- Cardona L, Revelles M, Parga ML, Tomás J, Aguilar A, Alegre F, Raga A, Ferrer X. 2009. Habitat use by loggerhead sea turtles Caretta caretta off the coast of eastern Spain results in a high vulnerability to neritic fishing gear. Marine Biology 156:2621–2630. doi:10.1007/s00227-009-1288-9.
- Carr A, Meylan AB. 1980. Evidence of passive migration of green turtle hatchlings in Sargassum. Copeia 1980:366–368. doi:10.2307/1444022.
- Carreras C, Pascual M, Cardona L, Aguilar A, Margaritoulis D, Rees A, Turkozan O, Levy Y, Gasith A, Aureggi M, Khalil M. 2007. The genetic structure of the loggerhead sea turtle (Caretta caretta) in the Mediterranean as revealed by nuclear and mitochondrial DNA and its conservation implications. Conservation Genetics 8:761–775. doi:10.1007/s10592-006-9224-8.
- Carreras C, Pascual M, Cardona L, Marco A, Bellido JJ, Castillo JJ, Tomas J, Raga JA, Sanfelix M, Fernandez G, Aguilar A. 2011. Living together but remaining apart: Atlantic and Mediterranean loggerhead sea turtles (Caretta caretta) in shared feeding grounds. Journal of Heredity 102:666–677. doi:10.1093/jhered/esr089.
- Carreras C, Pont S, Maffucci F, Pascual M, Barceló A, Bentivegna F, Cardona L, Alegre F, SanFélix M, Fernández G, Aguilar A. 2006. Genetic structuring of immature loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea reflects water circulation patterns. Marine Biology 149:1269–1279. doi:10.1007/s00227-006-0282-8.
- Casale P. 2011. Sea turtle by-catch in the Mediterranean. Fish and Fisheries 12:299–316. doi:10.1111/j.1467-2979.2010.00394.x.
- Casale P, Abbate G, Freggi D, Conte N, Oliverio M, Argano R. 2008a. Foraging ecology of loggerhead sea turtles Caretta caretta in the central Mediterranean Sea: Evidence for a relaxed life history model. Marine Ecology Progress Series 372:265–276. doi:10.3354/meps07702.
- Casale P, Affronte M, Insacco G, Freggi D, Vallini C, d’Astore PP, Basso R, Paolillo G, Abbate G, Argano R. 2010. Sea turtle strandings reveal high anthropogenic mortality in Italian waters. Aquatic Conservation: Marine and Freshwater Ecosystems 20:611–620. doi:10.1002/aqc.1133.
- Casale P, Affronte M, Scaravelli D, Lazar B, Vallini C, Luschi P. 2012a. Foraging grounds, movement patterns and habitat connectivity of juvenile loggerhead turtles (Caretta caretta) tracked from the Adriatic Sea. Marine Biology 159:1527–1535. doi:10.1007/s00227-012-1937-2.
- Casale P, Broderick AC, Freggi D, Mencacci R, Fuller WJ, Godley BJ, Luschi P. 2012b. Long-term residence of juvenile loggerhead turtles to foraging grounds: A potential conservation hotspot in the Mediterranean. Aquatic Conservation: Marine and Freshwater Ecosystems 22:144–154. doi:10.1002/aqc.2222.
- Casale P, Conte N, Freggi D, Cioni C, Argano R. 2011a. Age and growth determination by skeletochronology in loggerhead sea turtles (Caretta caretta) from the Mediterranean Sea. Scientia Marina 75:197–203. doi:10.3989/scimar.2011.75n1197.
- Casale P, Freggi D, Basso R, Vallini C, Argano R. 2007a. A model of area fidelity, nomadism, and distribution patterns of loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea. Marine Biology 152:1039–1049. doi:10.1007/s00227-007-0752-7.
- Casale P, Freggi D, Cinà A, Rocco M. 2013a. Spatio-temporal distribution and migration of adult male loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea: Further evidence of the importance of neritic habitats off North Africa. Marine Biology 160:703–718. doi:10.1007/s00227-012-2125-0.
- Casale P, Freggi D, Dourdeville KM, Prescot R. 2013b. First evidence of migration by loggerhead sea turtles, Caretta caretta, from the eastern Mediterranean to North America. Vie et Milieu 63:93–96.
- Casale P, Freggi D, Furii G, Vallini C, Salvemini P, Deflorio M, Totaro G, Raimondi S, Fortuna C, Godley B. 2014. Annual survival probabilities of juvenile loggerhead sea turtles indicate high anthropogenic impact to Mediterranean populations. Aquatic Conservation: Marine and Freshwater Ecosystems. doi:10.1002/aqc.2467.
- Casale P, Freggi D, Gratton P, Argano R, Oliverio M. 2008b. Mitochondrial DNA reveals regional and interregional importance of the central Mediterranean African shelf for loggerhead sea turtles (Caretta caretta). Scientia Marina 72:541–548. doi:10.3989/scimar.2008.72n3541
- Casale P, Laurent L, De Metrio G. 2004. Incidental capture of marine turtles by the Italian trawl fishery in the north Adriatic Sea. Biological Conservation 119:287–295. doi:10.1016/j.biocon.2003.11.013.
- Casale P, Margaritoulis D. 2010. Sea turtles in the Mediterranean: Distribution, threats and conservation priorities. Gland, Switzerland: IUCN.
- Casale P, Mariani P. 2014. The first ‘lost year’ of Mediterranean Sea turtles: Dispersal patterns indicate subregional management units for conservation. Marine Ecology Progress Series 498:263–274. doi:10.3354/meps10640.
- Casale P, Mazaris AD, Freggi D. 2011b. Estimation of age at maturity of loggerhead sea turtles Caretta caretta in the Mediterranean using length-frequency data. Endangered Species Research 13:123–129. doi:10.3354/esr00319.
- Casale P, Mazaris AD, Freggi D, Basso R, Argano R. 2007b. Survival probabilities of loggerhead sea turtles (Caretta caretta) estimated from capture-mark-recapture data in the Mediterranean Sea. Scientia Marina 71:365–372.
- Casale P, Mazaris AD, Freggi D, Vallini C, Argano R. 2009. Growth rates and age at adult size of loggerhead sea turtles (Caretta caretta) in the Mediterranean Sea, estimated through capture-mark-recapture records. Scientia Marina 73:589–595. doi:10.3989/scimar.2009.73n3589.
- Casale P, Nicolosi P, Freggi D, Turchetto M, Argano R. 2003. Leatherback turtles (Dermochelys coriacea) in Italy and in the Mediterranean basin. Herpetological Journal 13:135–139.
- Cheng I-J. 2000. Post-nesting migrations of green turtles (Chelonia mydas) at Wan-An Island, Penghu Archipelago, Taiwan. Marine Biology 137:747–754. doi:10.1007/s002270000375.
- Clusa M, Carreras C, Pascual M, Demetropoulos A, Margaritoulis D, Rees AF, Hamza AA, Khalil M, Aureggi M, Levy Y, Türkozan O, Marco A, Aguilar A, Cardona L. 2013. Mitochondrial DNA reveals Pleistocenic colonisation of the Mediterranean by loggerhead turtles (Caretta caretta). Journal of Experimental Marine Biology and Ecology 439:15–24. doi:10.1016/j.jembe.2012.10.011.
- Clusa M, Carreras C, Pascual M, Gaughran S, Piovano S, Giacoma C, Fernández G, Levy Y, Tomás J, Raga J, Maffucci F, Hochscheid S, Aguilar A, Cardona L. 2014. Fine-scale distribution of juvenile Atlantic and Mediterranean loggerhead turtles (Caretta caretta) in the Mediterranean Sea. Marine Biology 161:509–519. doi:10.1007/s00227-013-2353-y
- Coyne MS, Godley BJ. 2005. Satellite Tracking and Analysis Tool (STAT): An integrated system for archiving, analyzing and mapping animal tracking data. Marine Ecology Progress Series 301:1–7. doi:10.3354/meps301001.
- Craig P, Parker D, Brainard R, Rice M, Balazs G. 2004. Migrations of green turtles in the central South Pacific. Biological Conservation 116:433–438. doi:10.1016/S0006-3207(03)00217-9.
- D’Ilio S, Mattei D, Blasi MF, Alimonti A, Bogialli S. 2011. The occurrence of chemical elements and POPs in loggerhead turtles (Caretta caretta): An overview. Marine Pollution Bulletin 62:1606–1615. doi:10.1016/j.marpolbul.2011.05.022.
- Dodd CK, Byles R Jr. 2003. Post-nesting movements and behavior of loggerhead sea turtles (Caretta caretta) departing from east-central Florida nesting beaches. Chelonian Conservation and Biology 4:530–536.
- Eckert SA, Moore JE, Dunn DC, van Buiten RS, Eckert KL, Halpin PN. 2008. Modeling loggerhead turtle movement in the Mediterranean: Importance of body size and oceanography. Ecological Applications 18:290–308. doi:10.1890/06-2107.1.
- Encalada SE, Lahanas PN, Bjorndal KA, Bolten AB, Miyamoto MM, Bowen BW. 1996. Phylogeography and population structure of the Atlantic and Mediterranean green turtle Chelonia mydas: A mitochondrial DNA control region sequence assessment. Molecular Ecology 5:473–483. doi:10.1111/j.1365-294X.1996.tb00340.x.
- Bentivegna F. 2002. Intra-Mediterranean migrations of loggerhead sea turtles (Caretta caretta) monitored by satellite telemetry. Marine Biology 141:795–800. doi:10.1007/s00227-002-0856-z.
- Fuller WJ, Broderick AC, Phillips RA, Silk JRD, Godley BJ. 2008. Utility of geolocating light loggers for indicating at-sea movements in sea turtles. Endangered Species Research 4:139–146. doi:10.3354/esr00048.
- Garofalo L, Mastrogiacomo A, Casale P, Carlini R, Eleni C, Freggi D, Gelli D, Knittweis L, Mifsud C, Mingozzi T, Novarini N, Scaravelli D, Scillitani G, Oliverio M, Novelletto A. 2013. Genetic characterization of central Mediterranean stocks of the loggerhead turtle (Caretta caretta) using mitochondrial and nuclear markers, and conservation implications. Aquatic Conservation: Marine and Freshwater Ecosystems 23:868–884. doi:10.1002/aqc.2338.
- Girard C, Tucker AD, Calmettes B. 2009. Post-nesting migrations of loggerhead sea turtles in the Gulf of Mexico: Dispersal in highly dynamic conditions. Marine Biology 156:1827–1839. doi:10.1007/s00227-009-1216-z.
- Godley BJ, Blumenthal JM, Broderick AC, Coyne MS, Godfrey MH, Hawkes LA, Witt MJ. 2008. Satellite tracking of sea turtles: Where have we been and where do we go next? Endangered Species Research 4:3–22. doi:10.3354/esr00060.
- Godley BJ, Broderick AC, Glen F, Hays GC. 2003. Post-nesting movements and submergence patterns of loggerhead marine turtles in the Mediterranean assessed by satellite tracking. Journal of Experimental Marine Biology and Ecology 287:119–134. doi:10.1016/S0022-0981(02)00547-6.
- Godley BJ, Richardson S, Broderick AC, Coyne MS, Glen F, Hays GC. 2002. Long-term satellite telemetry of the movements and habitat utilisation by green turtles in the Mediterranean. Ecography 25:352–362. doi:10.1034/j.1600-0587.2002.250312.x.
- Gomez de Segura A, Tomas J, Pedraza SN, Crespo EA, Raga JA. 2006. Abundance and distribution of the endangered loggerhead turtle in Spanish Mediterranean waters and the conservation implications. Animal Conservation 9:199–206. doi:10.1111/j.1469-1795.2005.00014.x.
- Hamann M, Godfrey MH, Seminoff JA, Arthur K, Barata PCR, Bjorndal KA, Bolten AB, Broderick A, Campbell LM, Carreras C, Casale P, Chaloupka M, Chan SKF, Coyne MS, Crowder LB, Diez CE, Dutton PH, Epperly SP, FitzSimmons NN, Formia A, Girondot M, Hays GC, Cheng IS, Kaska Y, Lewison R, Mortimer JA, Nichols WJ, Reina RD, Shanker K, Spotila JR, Tomas J, Wallace BP, Work TM, Zbinden J, Godley BJ. 2010. Global research priorities for sea turtles: Informing management and conservation in the 21st century. Endangered Species Research 11:245–269.
- Hamza A. 2010. Libya. In: Casale P, Margaritoulis D, editors. Sea turtles in the Mediterranean: Distribution, threats and conservation priorities, Gland, Switzerland: IUCN. pp. 157–170.
- Hatase H, Omuta K, Tsukamoto K. 2007. Bottom or midwater: Alternative foraging behaviours in adult female loggerhead sea turtles. Journal of Zoology 273:46–55. doi:10.1111/j.1469-7998.2007.00298.x.
- Hatase H, Takai N, Matsuzawa Y, Sakamoto W, Omuta K, Goto K, Arai N, Fujiwara T. 2002. Size-related differences in feeding habitat use of adult female loggerhead turtles Caretta caretta around Japan determined by stable isotope analyses and satellite telemetry. Marine Ecology-Progress Series 233:273–281. doi:10.3354/meps233273.
- Hawkes LA, Broderick AC, Coyne MS, Godfrey MH, Godley BJ. 2007. Only some like it hot - quantifying the environmental niche of the loggerhead sea turtle. Diversity and Distributions 13:447–457. doi:10.1111/j.1472-4642.2007.00354.x.
- Hawkes LA, Broderick AC, Coyne MS, Godfrey MH, Lopez-Jurado L-F., Lopez-Suarez P, Merino SE, Varo-Cruz N, Godley BJ. 2006. Phenotypically linked dichotomy in sea turtle foraging requires multiple conservation approaches. Current Biology 16:990–995. doi:10.1016/j.cub.2006.03.063.
- Hays GC, Fossette S, Katselidis KA, Mariani P, Schofield G. 2010a. Ontogenetic development of migration: Lagrangian drift trajectories suggest a new paradigm for sea turtles. Journal of the Royal Society Interface 7:1319–1327.
- Hays GC, Fossette S, Katselidis KA, Schofield G, Gravenor MB. 2010b. Breeding periodicity for male sea turtles, operational sex ratios, and implications in the face of climate change. Conservation Biology 24:1636–1643. doi:10.1111/j.1523-1739.2010.01531.x.
- Hays GC, Marsh R. 1997. Estimating the age of juvenile loggerhead sea turtles in the North Atlantic. Canadian Journal of Zoology-Revue Canadienne De Zoologie 75:40–46. doi:10.1139/z97-005.
- Hays GC, Webb PI, Hayes JP, Priede IG, French J. 1991. Satellite tracking of a loggerhead turtle (Caretta caretta) in the Mediterranean. Journal of the Marine Biological Association of the United Kingdom 71:743–746. doi:10.1017/S0025315400053261.
- Hazel J. 2009. Evaluation of fast-acquisition GPS in stationary tests and fine-scale tracking of green turtles. Journal of Experimental Marine Biology and Ecology 374:58–68. doi:10.1016/j.jembe.2009.04.009.
- Hendrickson JR. 1958. The green sea turtle, Chelonia mydas (Linn.) in Malaya and Sarawak. Proceedings of the Zoological Society of London 130:455–535. doi:10.1111/j.1096-3642.1958.tb00583.x.
- Heppell SS. 1998. Application of life-history theory and population model analysis to turtle conservation. Copeia 1998:367–375. doi:10.2307/1447430.
- Hochscheid S, Bentivegna F, Bradai MN, Hays GC. 2007. Overwintering behaviour in sea turtles: Dormancy is optional. Marine Ecology Progress Series 340:287–298. doi:10.3354/meps340287.
- Hochscheid S, Bentivegna F, Hamza A, Hays GC. 2010. When surfacers do not dive: Multiple significance of extended surface times in marine turtles. Journal of Experimental Biology 213:1328–1337. doi:10.1242/jeb.037184.
- Hochscheid S, Travaglini A, Maffucci F, Hays GC, Bentivegna F. 2013. Since turtles cannot talk: What beak movement sensors can tell us about the feeding ecology of neritic loggerhead turtles, Caretta caretta. Marine Ecology 34:321–333. doi:10.1111/maec.12018.
- Howell EA, Dutton PH, Polovina JJ, Bailey H, Parker DM, Balazs GH. 2010. Oceanographic influences on the dive behavior of juvenile loggerhead turtles (Caretta caretta) in the North Pacific Ocean. Marine Biology 157:1011–1026. doi:10.1007/s00227-009-1381-0.
- IUCN. 2012. IUCN Red List of Threatened Species. Version 2012.2. Available: http://www.iucnredlist.org/. Accessed Sept 2014 3.
- Jones TT, Van Houtan KS, Bostrom BL, Ostafichuk P, Mikkelsen J, Tezcan E, Carey M, Imlach B, Seminoff JA. 2013. Calculating the ecological impacts of animal-borne instruments on aquatic organisms. Methods in Ecology and Evolution 4:1178–1186. doi:10.1111/2041-210X.12109.
- Kobayashi DR, Polovina JJ, Parker DM, Kamezaki N, Cheng I-J., Uchida I, Dutton PH, Balazs GH. 2008. Pelagic habitat characterization of loggerhead sea turtles, Caretta caretta, in the North Pacific Ocean (1997-2006): Insights from satellite tag tracking and remotely sensed data. Journal of Experimental Marine Biology and Ecology 356:96–114. doi:10.1016/j.jembe.2007.12.019.
- Laurent L, Casale P, Bradai MN, Godley BJ, Gerosa G, Broderick AC, Schroth W, Schierwater B, Levy AM, Freggi D, Abd El-Mawla EM, Hadoud DA, Gomati HE, Domingo M, Hadjichristophorou M, Kornaraky L, Demirayak F, Gautier C. 1998. Molecular resolution of marine turtle stock composition in fishery bycatch: A case study in the Mediterranean. Molecular Ecology 7:1529–1542. doi:10.1046/j.1365-294x.1998.00471.x.
- Lauriano G, Panigada S, Casale P, Pierantonio N, Donovan G. 2011. Aerial survey abundance estimates of the loggerhead sea turtle Caretta caretta in the Pelagos Sanctuary, northwestern Mediterranean Sea. Marine Ecology Progress Series 437:291–302. doi:10.3354/meps09261.
- Lazar B, Casale P, Tvrtkovic N, Kozul V, Tutman P, Glavic N. 2004. The presence of the green sea turtle, Chelonia mydas, in the Adriatic Sea. Herpetological Journal 14:143–147.
- Lazar B, Gračan R. 2011. Ingestion of marine debris by loggerhead sea turtles, Caretta caretta, in the Adriatic Sea. Marine Pollution Bulletin 62:43–47. doi:10.1016/j.marpolbul.2010.09.013.
- Limpus CJ, Limpus DJ. 2001. The loggerhead turtle, Caretta caretta, in Queensland: Breeding migrations and fidelity to a warm temperate feeding area. Chelonian Conservation and Biology 4:142–153.
- Lohmann KJ, Luschi P, Hays GC. 2008. Goal navigation and island-finding in sea turtles. Journal of Experimental Marine Biology and Ecology 356:83–95. doi:10.1016/j.jembe.2007.12.017.
- Luschi P, Hays GC, Papi F. 2003. A review of long-distance movements by marine turtles, and the possible role of ocean currents. Oikos 103:293–302. doi:10.1034/j.1600-0706.2003.12123.x.
- Luschi P, Mencacci R, Vallini C, Ligas A, Lambardi P, Benvenuti S. 2013. Long-term tracking of adult loggerhead turtles (Caretta caretta) in the Mediterranean Sea. Journal of Herpetology 47:227–231. doi:10.1670/11-173.
- Luschi P, Papi F, Liew HC, Chan EH, Bonadonna F. 1996. Long-distance migration and homing after displacement in the green turtle (Chelonia mydas): A satellite tracking study. Journal of Comparative Physiology a-Sensory Neural and Behavioral Physiology 178:447–452.
- Lutcavage ME, Lutz PL. 1997. Diving physiology. In: Lutz PL, Musick JA, editors. The biology of sea turtles, Boca Raton, FL: CRC Press, Inc. pp. 277–296.
- Mansfield KL, Saba VS, Keinath JA, Musick JA. 2009. Satellite tracking reveals a dichotomy in migration strategies among juvenile loggerhead turtles in the Northwest Atlantic. Marine Biology 156:2555–2570. doi:10.1007/s00227-009-1279-x.
- Mansfield KL, Wyneken J, Porter W, Luo J. 2014. First satellite tracks of neonate sea turtles redefine the “lost years” oceanic niche. Proceedings of the Royal Society B: Biological Sciences 281. doi:10.1098/rspb.2013.3039.
- Manzella SA, Fontaine CT, Schroeder BA. 1988. Loggerhead sea turtle travels from Padre Island, Texas, to the mouth of the Adriatic Sea. Marine Turtle Newsletter 42:7.
- Margaritoulis D, Argano R, Baran I, Bentivegna F, Bradai MN, Caminas JA, Casale P, De Metrio G, Demetropoulos A, Gerosa G, Godley B, Houghton J, Laurent L, Lazar B. 2003. Loggerhead turtles in the Mediterranean Sea: Present knowledge and conservation perspectives. In: Bolten AB, Witherington B, editors. Biology and conservation of loggerhead sea turtles, Washington, DC: Smithsonian Institution Press. pp. 175–198.
- Margaritoulis D, Panagopoulou A. 2010. Greece. In: Casale P, Margaritoulis D, editors. Sea turtles in the Mediterranean: Distribution, threats and conservation priorities. Gland, Switzerland: IUCN. pp. 85–112.
- Margaritoulis D, Rees AF. 2011. Loggerhead turtles nesting at Rethymno, Greece, prefer the Aegean Sea as their main foraging area. Marine Turtle Newsletter 131:12–14.
- Mazaris AD, Kallimanis AS, Pantis JD, Hays GC. 2013. Phenological response of sea turtles to environmental variation across a species’ northern range. Proceedings of the Royal Society B: Biological Sciences 280. doi:10.1098/rspb.2012.2397.
- McClellan CM, Braun-McNeill J, Avens L, Wallace BP, Read AJ. 2010. Stable isotopes confirm a foraging dichotomy in juvenile loggerhead sea turtles. Journal of Experimental Marine Biology and Ecology 387:44–51. doi:10.1016/j.jembe.2010.02.020.
- McClellan CM, Read AJ. 2007. Complexity and variation in loggerhead sea turtle life history. Biology Letters 3:592–594. doi:10.1098/rsbl.2007.0355.
- Mencacci R, Ligas A, Meschini P, Luschi P. 2011. Movements of three loggerhead sea turtles in Tuscany waters. Atti Società toscana di Scienze naturali, Memorie, Serie B 118:117–120.
- Mencacci R, Vallini C, Rubini S, Funes L, Sarti A, Benvenuti S, Luschi P. 2006. Movements of a male loggerhead sea turtle (Caretta caretta) tracked by satellite in the Adriatic Sea. In: Zuffi MAL, editor. Atti del V Congresso nazionale della Societas Herpetologica Italica. Florence: Firenze University Press. pp. 167–171.
- Meylan A. 1982. Sea turtle migration. Evidence from tag returns. In: Bjorndal KA, editor. Biology and conservation of sea turtles. Washington, DC: Smithsonian Institution Press. pp. 91–100.
- Moncada F, Abreu-Grobois FA, Bagley D, Bjorndal KA, Bolten AB, Camiñas JA, Ehrhart L, Muhlia-Melo A, Nodarse G, Schroeder BA, Zurita J, Hawkes LA. 2010. Movement patterns of loggerhead turtles Caretta caretta in Cuban waters inferred from flipper tag recaptures. Endangered Species Research 11:61–68. doi:10.3354/esr00248.
- Moritz C. 1994. Defining ‘Evolutionarily Significant Units’ for conservation. Trends in Ecology and Evolution 9:373–375. doi:10.1016/0169-5347(94)90057-4.
- Musick JA, Limpus CJ. 1997. Habitat utilization and migration in juvenile sea turtles. In: Lutz PL, Musick JA, editors. The Biology of Sea Turtles, Boca Raton, FL: CRC Marine Science Series, CRC Press, Inc. pp. 137–163.
- Nada M, Casale P. 2011. Sea turtle bycatch and consumption in Egypt threatens Mediterranean turtle populations. Oryx 45:143–149. doi:10.1017/S0030605310001286.
- Narazaki T, Sato K, Abernathy KJ, Marshall GJ, Miyazaki N. 2013. Loggerhead turtles (Caretta caretta) use vision to forage on gelatinous prey in mid-water. PLoS ONE 8:e66043. doi:10.1371/journal.pone.0066043.
- Oruç A. 2001. Trawl fisheries in the eastern Mediterranean and their impact on marine turtles. Zoology in the Middle East 24:119–125. doi:10.1080/09397140.2001.10637890
- Oruç A, Tural U, Cesur T. 2011. A review of potential marine habitats for marine turtles in Turkey. Marine Turtle Newsletter 131:23–25.
- Papi F, Luschi P, Crosio E, Hughes GR. 1997. Satellite tracking experiments on the navigational ability and migratory behaviour of the loggerhead turtle Caretta caretta. Marine Biology 129:215–220. doi:10.1007/s002270050162.
- Rees AF, Al Saady S, Broderick AC, Coyne MS, Papathanasopoulou N, Godley BJ. 2010. Behavioural polymorphism in one of the world’s largest populations of loggerhead sea turtles Caretta caretta. Marine Ecology Progress Series 418:201–212. doi:10.3354/meps08767.
- Rees AF, Jony M, Margaritoulis D, Godley BJ. 2008. Satellite tracking of a Green Turtle, Chelonia mydas, from Syria further highlights importance of North Africa for Mediterranean turtles. Zoology in the Middle East 45:49–54. doi:10.1080/09397140.2008.10638306.
- Rees AF, Margaritoulis D, Newman R, Riggall TE, Tsaros P, Zbinden JA, Godley BJ. 2013. Ecology of loggerhead marine turtles Caretta caretta in a neritic foraging habitat: Movements, sex ratios and growth rates. Marine Biology 160:519–529. doi:10.1007/s00227-012-2107-2.
- Reich KJ, Bjorndal KA, Frick MG, Witherington BE, Johnson C, Bolten AB. 2010. Polymodal foraging in adult female loggerheads (Caretta caretta). Marine Biology 157:113–121. doi:10.1007/s00227-009-1300-4.
- Revelles M, Caminas JA, Cardona L, Parga M, Tomas J, Aguilar A, Alegre F, Raga A, Bertolero A, Oliver G. 2008. Tagging reveals limited exchange of immature loggerhead sea turtles (Caretta caretta) between regions in the western Mediterranean. Scientia Marina 72:511–518.
- Revelles M, Cardona L, Aguilar A, SanFélix M, Fernández G. 2007a. Habitat use by immature loggerhead sea turtles in the Algerian Basin (western Mediterranean): Swimming behaviour, seasonality and dispersal pattern. Marine Biology 151:1501–1515. doi:10.1007/s00227-006-0602-z.
- Revelles M, Isern-Fontanet J, Cardona L, San Félix MS, Carreras C, Aguilar A. 2007b. Mesoscale eddies, surface circulation and the scale of habitat selection by immature loggerhead sea turtles. Journal of Experimental Marine Biology and Ecology 347:41–57. doi:10.1016/j.jembe.2007.03.013.
- Sasso CR, Epperly SP. 2007. Survival of pelagic juvenile loggerhead turtles in the open ocean. Journal of Wildlife Management 71:1830–1835. doi:10.2193/2006-448.
- Schofield G, Bishop CM, MacLean G, Brown P, Baker M, Katselidis KA, Dimopoulos P, Pantis JD, Hays GC. 2007. Novel GPS tracking of sea turtles as a tool for conservation management. Journal of Experimental Marine Biology and Ecology 347:58–68. doi:10.1016/j.jembe.2007.03.009.
- Schofield G, Dimadi A, Fossette S, Katselidis KA, Koutsoubas D, Lilley MKS, Luckman A, Pantis JD, Karagouni AD, Hays GC. 2013. Satellite tracking large numbers of individuals to infer population level dispersal and core areas for the protection of an endangered species. Diversity and Distributions 19:834–844. doi:10.1111/ddi.12077.
- Schofield G, Hobson VJ, Fossette S, Lilley MKS, Katselidis KA, Hays GC. 2010a. Fidelity to foraging sites, consistency of migration routes and habitat modulation of home range by sea turtles. Diversity and Distributions 16:840–853. doi:10.1111/j.1472-4642.2010.00694.x
- Schofield G, Hobson VJ, Lilley MKS, Katselidis KA, Bishop CM, Brown P, Hays GC. 2010b. Inter-annual variability in the home range of breeding turtles: Implications for current and future conservation managementì. Biological Conservation 143:722–730. doi:10.1016/j.biocon.2009.12.011.
- Schofield G, Lilley MKS, Bishop CM, Brown P, Katselidis KA, Dimopoulos P, Pantis JD, Hays GC. 2010c. Conservation hotspots: Implications of intense spatial area use by breeding male and female loggerheads at the Mediterranean’s largest rookery. Endangered Species Research 10:191–202. doi:10.3354/esr00137.
- Scott R, Marsh R, Hays GC. in press. Ontogeny of long distance migration. Ecology. Available: http://dx.doi.org/10.1890/13-2164.1. Accessed Sept 2014 3.
- Seminoff JA, Zárate P, Coyne M, Foley DG, Parker D, Lyon BN, Dutton PH. 2008. Post-nesting migrations of Galápagos green turtles Chelonia mydas in relation to oceanographic conditions: Integrating satellite telemetry with remotely sensed ocean data. Endangered Species Research 4:57–72. doi:10.3354/esr00066.
- Stoneburner DL. 1982. Satellite telemetry of loggerhead sea turtle movement in the Georgia bight USA. Copeia 1982:400–408. doi:10.2307/1444621.
- Swimmer Y, Arauz R, McCracken M, McNaughton L, Ballestero J, Musyl M, Bigelow K, Brill R. 2006. Diving behavior and delayed mortality of olive ridley sea turtles Lepidochelys olivacea after their release from longline fishing gear. Marine Ecology Progress Series 323:253–261. doi:10.3354/meps323253.
- Timko RE, Kolz AL. 1982. Satellite sea turtle tracking. Marine Fisheries Review 44:19–24.
- Giovannotti M, Franzellitti S, Cerioni PN, Fabbri E, Guccione S, Vallini C, Tinti F, Caputo V. 2010. Genetic characterization of loggerhead turtle (Caretta caretta) individuals stranded and caught as bycatch from the North-Central Adriatic Sea. Amphibia-Reptilia 31:127–133. doi:10.1163/156853810790457902.
- Tomás J, Gozalbes P, Raga JA, Godley BJ. 2008. Bycatch of loggerhead sea turtles: Insights from 14 years of stranding data. Endangered Species Research 5:161–169. doi:10.3354/esr00116.
- Tomás J, Guitart R, Mateo R, Raga JA. 2002. Marine debris ingestion in loggerhead sea turtles, Caretta caretta from the Western Mediterranean. Marine Pollution Bulletin 44:211–216. doi:10.1016/S0025-326X(01)00236-3.
- Turkecan O, Yerli SV. 2011. Satellite tracking of adult green sea turtles from Turkey: A long distance diary. Marine Turtle Newsletter 131:38–41.
- Türkozan O, Özdilek SY, Ergene S, Uçar AH, Sönmez B, Yilmaz C, Kaçar Y, Aymak C. 2013. Strandings of loggerhead (Caretta caretta) and green (Chelonia mydas) sea turtles along the eastern Mediterranean coast of Turkey. Herpetological Journal 23:11–15.
- Wallace BP, DiMatteo AD, Bolten AB, Chaloupka MY, Hutchinson BJ, Abreu-Grobois FA, Mortimer JA, Seminoff JA, Amorocho D, Bjorndal KA, Bourjea J, Bowen BW, Briseño Dueñas R, Casale P, Choudhury BC, Costa A, Dutton PH, Fallabrino A, Finkbeiner EM, Girard A, Girondot M, Hamann M, Hurley BJ, López-Mendilaharsu M, Marcovaldi MA, Musick JA, Nel R, Pilcher NJ, Troëng S, Witherington B, Mast RB. 2011. Global conservation priorities for marine turtles. PLoS ONE 6:e24510.
- Watson KP, Granger RA. 1998. Hydrodynamic effect of a satellite transmitter on a juvenile green turtle (Chelonia mydas). Journal of Experimental Biology 201:2497–2505.
- Wright LI, Stokes KL, Fuller WJ, Godley BJ, McGowan A, Snape R, Tregenza T, Broderick AC. 2012. Turtle mating patterns buffer against disruptive effects of climate change. Proceedings of the Royal Society B: Biological Sciences 279:2122–2127.
- Zbinden JA, Aebischer A, Margaritoulis D, Arlettaz R. 2007. Insights into the management of sea turtle internesting area through satellite telemetry. Biological Conservation 137:157–162. doi:10.1016/j.biocon.2007.01.022.
- Zbinden JA, Aebischer A, Margaritoulis D, Arlettaz R. 2008. Important areas at sea for adult loggerhead sea turtles in the Mediterranean Sea: Satellite tracking corroborates findings from potentially biased sources. Marine Biology 153:899–906. doi:10.1007/s00227-007-0862-2.
- Zbinden JA, Bearhop S, Bradshaw P, Gill B, Margaritoulis D, Newton J, Godley BJ. 2011. Migratory dichotomy and associated phenotypic variation in marine turtles revealed by satellite tracking and stable isotope analysis. Marine Ecology Progress Series 421:291–302. doi:10.3354/meps08871.