
ABSTRACT
This study examines the Holocene evolution of an inland subarctic permafrost peatland located on the north bank of Wiyâshâkimî Lake (Nunavik, northeastern Canada). The analysis of plant macrofossils allowed us to reconstruct the succession of the trophic conditions of a palsa and a filled thermokarst pond. The accumulation of organic matter began at around 6290 cal. y BP. The evolution of the site then followed three stages: a pond (6290–5790 cal. y BP), a minerotrophic peatland (5790–4350 cal. y BP) and an ombrotrophic peatland (from 4350 cal. y BP). The establishment of permafrost caused a palsa to form at around 170 cal. y BP, which corresponds to the coldest period of the Little Ice Age in northeastern Canada. A subsequent degradation of the palsa and the formation of a thermokarst pond were induced by the climate warming that began at the turn of the 20th century. The analysis of plant macrofossils from an adjacent filled thermokarst pond indicated three phases of development over a short 450-year period: subaquatic, minerotrophic, and ombrotrophic phases. When combined with previous studies of filled thermokarst ponds in northern Québec, this result indicates that ponds are rapidly filling in with vegetation and acting as carbon sinks.
RÉSUMÉ
Cette étude vise, à l’aide de l’analyse macrofossile, à reconstituer l’évolution des conditions trophiques d’une tourbière à pergélisol et d’une mare thermokarstique comblée situées sur la rive nord du Lac Wiyâshâkimî (Nunavik, nord-est du Canada). L’accumulation de la matière organique a commencé vers 6290 cal. y BP. Trois phases ont caractérisé l’évolution de la tourbière depuis son origine: un étang (6290-5790 cal. y BP), une tourbière minérotrophe (5790-4350 cal. y BP) puis une tourbière ombrotrophe (à partir de 4350 cal. y BP). L’établissement du pergélisol dans la tourbière a progressivement soulevé sa surface, conduisant à la formation d’une palse vers 170 cal. y BP, lors d’une des périodes les plus froides du Petit Âge glaciaire dans le nord-est du Canada. La dégradation de la palse et la formation d’une mare thermokarstique ont été favorisées par le réchauffement climatique qui a commencé au début du XXe siècle. L’analyse macrofossile d’une mare thermokarstique comblée, située à proximité de la palse a révélé trois phases de développement durant une courte période de 450 ans: subaquatique, minérotrophe et ombrotrophe. Combiné à d’autres études similaires au Québec nordique, ce résultat montre que les mares thermokarstiques sont comblées rapidement par la végétation et agissent comme des puits de carbone.
Introduction
Palsa peatlands and peat plateaus are the most important periglacial features in the circumpolar subarctic and high-boreal regions, in which they form a large northern ecosystem (e.g., Zoltaï and Tarnocai Citation1971; Zoltaï Citation1993, Citation1995; Sollid and Sorbel Citation1998; Payette Citation2001; Bhiry et al. Citation2007; Kuhry Citation2008; Thibault and Payette Citation2009). Their spatiotemporal distribution depends on local and regional climatic factors. The establishment and evolution of these ecosystems during the Holocene were driven chiefly by climate and topography, but autogenic factors such as peat accumulation and acidification also played a significant role (Seppälä Citation1986). Several studies have shown that palsa formation in northeastern Canada occurred during the coldest periods of the Late-Holocene, e.g., the Neoglacial period (which started at about 3500 cal. y BP) and the Little Ice Age (LIA; 1650–1850 A.D), due to cold conditions and the decrease in snow precipitation (e.g., Payette Citation2009; Bhiry and Lavoie Citation2018).
These ecosystems are currently degrading in response to global warming in the 20th to 21st centuries, generating distinct adjacent zones: palsas with varying degrees of degradation, collapsed palsas, thermokarst ponds and filled thermokarst ponds (through terrestrialisation) (Laprise and Payette Citation1988; Fillion et al. Citation2014; Swindles et al. Citation2015). This degradation induces a significant increase in greenhouse carbon gases, principally methane from thermokarst ponds (Moore and Roulet Citation1993; Matveev et al. Citation2016), as well as changes in the local vegetation cover. On the other hand, recent studies have found high carbon accumulation rates in peatlands located in northern regions (high-boreal and subarctic regions) during the last two centuries in response to climate warming (e.g., Treat et al. Citation2016; Piilo et al. Citation2019), while other studies such as Abbott et al. (Citation2016) indicate that ecosystems in the permafrost regions will become a carbon source to the atmosphere by 2100 as a result of climate warming.
In Nunavik (northeastern Canada), studies of the long-term evolution of permafrost peatlands focused on areas along the coasts (James Bay, Hudson Bay, Ungava Bay; ) where fine postglacial marine sediments have been deposited (e.g., Richard Citation1981; Couillard and Payette Citation1985; Allard and Seguin Citation1987; Lavoie and Payette Citation1995; Payette et al. Citation2004; Arlen-Pouliot and Bhiry Citation2005; Bhiry and Robert Citation2006; Bhiry et al. Citation2007; Thibault and Payette Citation2009; Lamarre et al. Citation2012; Fillion et al. Citation2014; Tremblay et al. Citation2014). Using a paleoecological approach that focuses on plant-macrofossils, pollen and testate amoeba, these studies showed the asynchronous succession of several developmental stages of permafrost peatlands, starting with a shallow body of water (pond) that began to fill with vegetation. A rich minerotrophic peatland developed afterward, followed by a poor minerotrophic peatland in which the surface runoff provided the mineral nutrients needed for plant growth. The minerotrophic phase evolved into the ombrotrophic phase, which lasted for a very short period before palsa formation. In the high-boreal zone, the succession from the minerotrophic to the ombrotrophic phase prior to the formation of palsa is the result of the establishment of permafrost in the peatland. The location of the studied palsa peatlands along the coasts of Hudson Bay and James Bay in Nunavik means that the distinctive climate would have had a significant impact on these ecosystems. Specifically, the summers are more temperate, as the cold waters of the bays and the winds from the west affect the region. Hence, for example, the cool summers allow the developing permafrost lenses to persist throughout the thawing season, which may not be the case in more inland areas where summers are relatively warmer with moderate precipitation.
Figure 1. Location of the study site (X) on a small island close to the northern bank of Wiyâshâkimî Lake (Nunavik, Canada) and of sites mentioned in the text: 1) northwestern palsa (NW-palsa); 2) southeastern palsa (SE-palsa); 3) central ombrotrophic peatland (Payette Citation1988); 4) Bear peatland on Lepage Island (Bhiry et al. Citation2019). The ecological zones map is from Payette (Citation1983)
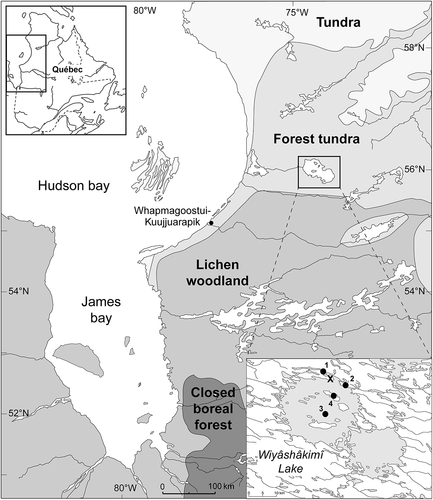
This study focuses on a palsa peatland located in the Wiyâshâkimî Lake (Lac à l’Eau-Claire) region about 150 km east of the Hudson Bay coast (), outside the area of influence of the maritime climate. Many environmental studies (geology, geomorphology, forest ecology, etc.) have been carried out in the Wiyâshâkimî Lake region (e.g., Simard et al. Citation2004; Schmieder et al. Citation2015; Decaulne et al. Citation2018), but only two sites were studied from a paleoecological perspective: the first consists of an ombrotrophic peatland characterised by ice wedges that covers a small island in the center of the western basin of Wiyâshâkimî Lake (Payette et al. Citation1986; Payette Citation1988; ). The development of this peatland over the last 5000 years was driven by climate as well as by cyclic-autogenic succession (Payette Citation1988). The second site is a minerotrophic peatland located on Lepage Island, which is one of the islands that is arranged in a ring shape in the western basin of the lake (). This site is situated close to the coast about 400 m north-east of a 30–50 m-high slope. Debris flow features (i.e., levees, channels and lobes) stretch for about 200 m from the talus slope towards the peatland (Bhiry et al. Citation2019). Local conditions have remained wet since 4900 cal. y BP, which prevented the establishment of permafrost.
According to our observations, these categories of peatland (ombrotrophic peatlands with ice wedges and minerotrophic peatlands) are rare in the region, while the peatland complex formed by palsas, ponds, and minerotrophic stands around the lake and on the islands is much more common. In this study, we used plant-macrofossils to reconstruct the developmental stages of an inland palsa peatland since its origin. The results were then compared with other studies of palsa peatlands that are located on the Hudson Bay and James Bay coasts in order to identify the potential impacts of continental conditions and climate change.
Study area
The study site is located on the north shore of Wiyâshâkimî Lake (Lac à l’Eau-Claire) (56°20ʹ524”N; 74°27ʹ167”W). It lies inland approximately 150 km from Hudson Bay at an altitude of 238 m (). Wiyâshâkimî Lake is made up of two circular basins (western and eastern) that were created by a double meteor impact approximately 290 million years ago (Schmieder et al. Citation2015). The region is part of the Canadian Shield, which consists of granite-gneiss outcrops that formed approximately 2.5 billion years ago (Avramtchev Citation1982). The western basin is distinct from the eastern one as it includes a series of large islands that form a circle and a few small islands situated in the centre ().
The retreat of the glacier would have occurred very quickly on the Hudson Bay coast, between 8000 and 7000 cal. y BP. At Wiyâshâkimî Lake, deglaciation occurred between 6500 and 6000 cal. y BP (Citation1986; Allard and Seguin Citation1987). The ice retreat was followed by the Tyrell Sea transgression, which would have extended to the North River, the Caribou River, the Lac à l’Eau Claire River and the Ouiatchouane River, reaching somewhere between Tasiujaq Lake (Guillaume-Delisle Lake) and Wiyâshâkimî Lake. The presence of a delta in the Eau-Claire River valley at an altitude of 220 m suggests that the maximum limit of the Tyrell Sea was lower than the altitude of Wiyâshâkimî Lake (Begin Citation1986).
The regional climate is subarctic. However, given its location inland and its higher elevation, the Wiyâshâkimî Lake region was not influenced by the Hudson Bay climate in the same way as in the coastal zones. The local climate is characterised by a late spring thaw and strong winds. Weather data was gathered from the SILA weather station that is situated on the northern shore of the lake (SILA is a northern network of climate and environmental observatories run by the Centre d’études nordiques (CEN)). The annual data for 2006, 2007 and 2011–2015 indicate a mean annual temperature of −3.58°C (data available from the SILA network website: http://www.cen.ulaval.ca/sila.php?xml = cartesila< = 72&lg = −79&zm = 3) and 648 mm of precipitation annually, a third of which fell in the form of snow (equivalent to 241 mm of water). The Wiyâshâkimî Lake region is part of the forest tundra, a transition zone between the boreal forest and the arctic tundra (Payette Citation1983). Forest patches dominated by black spruce (Picea mariana) and eastern larch (Larix laricina) are located at sites sheltered from the wind such as depressions and the lower parts of slopes, while the summits are covered by discontinuous lichen-heath community cover. The region also includes many palsa peatlands.
The study site consists of a small palsa peatland located on a small island close the northern shore of Wiyâshâkimî Lake (). The palsa has an area of approximately 100 m2 and is bounded by a wet environment. The combination of a dry environment (palsa), a humid environment (infilled pond) and an aquatic environment (pond) generates a wide diversity of flora (). The filling of the pond was initiated at the edges of the site, beginning with Carex and Sphagnum. Ericaceae, such as Rubus chamaemorus, Vaccinium vitis-idaea, and Rhododendron groenlandicum, dominate the site in addition to lichens. The site is also surrounded by mature trees such as Picea mariana and Larix laricina ().
Figure 2. A view of the study peatland palsa showing distinct sectors: a palsa, a thermokarst pond and a vegetation-filled pond; photo credit: Denis Sarrazin, 2014. The yellow and white dots indicate the sampling location

Methods
Fieldwork
Fieldwork was conducted in August 2013. A survey of the surface vegetation of the palsa, the thermokarst pond and the vegetation-filled pond was carried out (). Species were identified using the Nunavik plant atlas (Blondeau et al. Citation2010) and the Vascan nomenclature (Brouillet et al. Citation2010+).
A 210 cm-long sedimentary core was extracted from the palsa: the upper portion (0–50 cm) consists of a monolith that was removed using a shovel, while the deeper sediments were collected using a STIHL™ 08 S drill fitted with a KB-300 GÖLZ™ transmission specially designed for permafrost zones (Calmels et al. Citation2005). Sediments were wrapped in aluminum foil, placed in plastic bags and then transported to the laboratory where they were stored at 4°C until further processing. After thawing, the core shortened to 186 cm (from 210 cm). A core (47 cm long) was also removed from the filled thermokarst pond (FTP) that is located beside the sampled palsa using a Russian peat corer (). The core was then subdivided into 1-cm-thick subsamples that were stored in bags and kept at a temperature of 4°C until the time of analysis. Stratigraphy of the sediments was described based on a visual scale of peat humification, colour and texture. Two other palsas located in two different palsa fields were also sampled (): one is approximately 2 km northwest of the study site (NW-palsa) and the other is located 3 km southeast of the study site (SE-palsa). Only the basal peat in contact with the lacustrine sediments and the peat at the top of these palsas were collected for radiocarbon dating.
Laboratory analyses: plant macrofossil data and 14C dating
A 2 cm interval was used for macrofossil analysis. In total, 95 samples from the palsa and 24 samples from the thermokarst pond were analysed. The samples were prepared according to the method of Bhiry and Filion (Citation2001). Identification of macrofossils was carried out using binocular and light microscopy. The remains of vascular and non-vascular plants were identified using the reference collection of the Centre d’études nordiques (CEN) and other sources (Rousseau Citation1974; Deshaye and Morisset Citation1985; Marie-Victorin Citation1995; Garneau Citation2001; Faubert Citation2013, Citation2014; Ayotte and Rochefort Citation2014). The volume occupied by each botanical group in the organic matrix of the peat (brown mosses, Sphagnum, herbaceous remains, wood remains, vascular leaves, charcoal) was estimated as a percentage by observing the entire sample. To identify bryophytes to the species level, 100 leaves of Sphagnum and 100 leaves of brown mosses from each sample were slide-mounted and examined under a compound microscope. Results for vascular plant remains were expressed as the number of macrofossils per 50 cm3 of sediment. For bryophytes, each species’ percentage representation in a sample was calculated within its botanical group (Sphagnum and brown mosses). Results were compiled into macrofossil diagrams using Palaeo Data Plotter (Juggins Citation2002). The zonation of the macrofossil diagrams is based on plant macrofossil assemblages and the stratigraphy of peat profiles.
Thirteen samples were dated using the AMS radiocarbon method (): eight samples were collected from the palsa, one from the FTP and four from the two nearby palsa peatlands. Samples consisted of plant-macrofossils such as leaves of vascular plants, leaves of brown mosses, and seeds. The samples were dated at the 14C laboratory at Laval University and at the Kerk Laboratory at the University of California. Radiocarbon dates (y BP) were calibrated (cal. y BP) using the INTCAL20 curve (Reimer et al. Citation2020).
Table 1. Radiocarbon and calibrated ages of the samples from the studied palsa and filled thermokarst pond and from two palsas located about 2–3 km from the study site, NW-palsa and SE-palsa
Results
Palsa chronostratigraphy and plant macrofossil data
Five macrofossil zones (Mac-1 to Mac-5) were identified based on the peat litho-stratigraphy (color and degree of humification of the peat) and the plant-macrofossil assemblages (). The Mac-2 and Mac-4 zones were subdivided into two subzones.
Figure 3. Summarized plant macrofossil diagram. Each group of botanical components and charcoal was estimated as a percentage by volume (50 cm3)
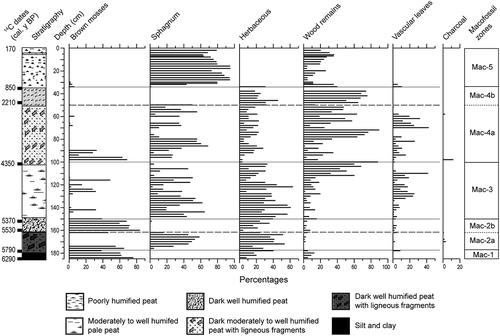
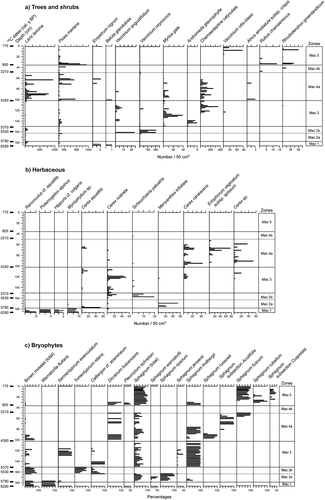
Figure 4. Macrofossil diagram of the studied palsa. (a) tree and shrub macrofossil diagram of the palsa (number of macrofossils per 50 cm3); (b) herbaceous macrofossil diagram of the palsa (number of macrofossils per 50 cm3); (c) brown mosses and Sphagnum macrofossil diagram of the palsa (macrofossil percentages)
Zone Mac-1: 185–178 cm (6290–5790 cal. y BP)
Zone Mac-1 begins at the bottom of the core and is 7 cm thick (185–178 cm). It is composed of greyish silty clay containing decomposed organic matter. The sediment accumulation rate was low (0.12 mm/yr; Figure A, supplemental material). The plant-macrofossil assemblages are dominated by aquatic species (Ranunculus cf. aquatilis, Potamogeton alpinus, Hippuris vulgaris) and subaquatic species (Carex rostrata, Carex aquatilis). Brown mosses were represented by Warnstorfia fluitans and Sarmentypnum exannulatum. Some seeds of Picea mariana, Betula glandulosa and Larix laricina were also identified, but in very small quantities.
Zone Mac-2: 177–150 cm (5790–5370 cal. y BP)
Peat corresponding to this zone is humified and includes wood fragments. The sediment accumulation rate was higher than it had been previously (0.40–0.64 mm/yr). Two subzones were delineated (Mac-2a and Mac-2b). The macrofossil assemblages of subzone 2a are made up of aquatic and rich minerotrophic plants, including Carex aquatilis, Carex rostrata, Myriophyllum sp., Menyanthes trifoliata, Sphagnum riparium and Sphagnum warnstorfii. The upper part of the subzone includes many minerotrophic species (Sphagnum Jensenii, Sphagnum lindbergii), which suggests a rapid infill of the pond. Wood fragments were found in several levels of this subzone (). Subzone Mac-2b is notable for the disappearance of Menyanthes trifoliata. The macroremains data also showed that Larix laricina and Picea mariana were still present locally. The presence of Scheuchzeria palustis, Carex rostrata and Vaccinium oxycoccos as well as the abundance of brown moss (Calliergon cf. stramineum, Tomentypnum nitens) indicates local moist conditions.
Zone Mac 3: 149–102 cm (5360–4350 cal. y BP)
Organic sediments corresponding to Zone Mac-3 are composed of moderately to very decomposed peat, with a significant change in the amount of shrub and herbaceous species remains (). The rate of accumulation was 0.49 cm/yr. The minerotrophic species were replaced by ombrotrophic species (Andromeda glaucophylla, Chamaedaphne calyculata, Vaccinium angustifolium, Vaccinium vitis-idaea, Rubus idaeus). Minerotrophic bryophytes were also replaced by ombrotrophic species (Calliergon cf. stramineum, Sarmentypnum exannulatum, Sphagnum jensenii, Sphagnum lindbergii) ()).
Zone Mac-4: 101–35 cm (4350–850 cal. y BP)
Zone Mac-4 covers a very long period (3500 years). Sediments correspond to well-decomposed brown peat rich in ligneous plant remains (), whereas the rate of accumulation reduced drastically compared to the previous zone (0.10–0.25 mm/yr; Figure A, supplemental material). Two subzones were identified: Mac-4a and Mac-4b.
In Subzone Mac-4a, the brown mosses disappeared at 90–91 cm ()). Charcoal fragments were found at two levels (). Remains from several tree and shrub taxa were also found: Larix laricina, Picea mariana, Betula glandulosa, Alnus rugosa, Empetrum nigrum (). The many leaves and seeds of Chamaedaphne calyculata in this zone indicate ombrotrophic conditions. The percentage of bryophyte remains also fluctuated significantly ()). Subzone Mac-4b is distinctive because of the total absence of Sphagnum and brown mosses. Very few macrofossils were retrieved, and the samples primarily contained ligneous material (roots, bark and wood fragments). Picea mariana dominated the site, whereas Larix laricina appears to have disappeared ()). Ombrotrophic species such as Rhododendron groenlandicum and Rubus chamaemorus were also identified.
Zone Mac-5: 34–0 cm (850–170 cal. y BP)
Zone Mac-5 is composed of poorly decomposed Sphagnum peat. The rate of sediment accumulation increased significantly from 0.10 mm/yr in Mac-4b to 0.49 mm/yr. The main Sphagnum species were Sphagnum lindbergii, Sphagnum rubellum and Sphagnum fuscum ()). In addition to Picea mariana, remains from Vaccinium vitis-idaea, Rhododendron groenlandicum and Rubus chamaemorus were identified. The peat stopped accumulating at 170 cal. y BP when the palsa began to form.
Chronostratigraphy and plant macrofossil data of the filled thermokarst pond
A 47 cm-long peat core was extracted from the filled thermokarst pond (FTP). It was composed of dark brown decomposed peat (47–32 cm) overlain by brownish peat composed of moderately decomposed Sphagnum (31–10 cm) and a poorly decomposed pale brown Sphagnum peat (9–0 cm). Three zones were identified: Mac-A (47–32 cm), Mac-B (31–10 cm), and Mac-C (9–0 cm) (). The transition between Mac-A and Mac-B was dated at 360 cal. y BP.
Figure 5. Macrofossil diagram of the studied filled thermokarst pond. (a) Summarized plant macrofossil diagram of the filled thermokarst pond (FTP); (b) brown mosses and sphagnum macrofossil diagram. Each group of botanical components and charcoal was estimated as a percentage by volume (15 cm3)

Zone Mac-A: 47–32 cm
Zone Mac-A is rich in ligneous macroremains and Sphagnum leaves and stems. A very large quantity of Picea mariana remains were found (600 to 1800). Macrofossils from Empetrum nigrum, Vaccinium angustifolium, Chamaedaphne calyculata and Rubus chamaemorus (ombrotrophic Ericacaeae) were abundant. A few remains of Carex canescens, Carex sp. and Eriophroum sp. were also found. Remains from bryophytes, which are typically found in ombrotrophic conditions, were identified in all of the samples. The macrofossil assemblages of this zone are similar to those of Mac-5.
Zone Mac-B: 31–10 cm
The macrofossil assemblages revealed two subzones: Mac-Ba and Mac-Bb. Subzone Mac-Ba is characterised by the abundance of macrofossils from Sphagnum and herbaceous taxa. There were very few remains from Ericacaeae (Empetrum nigrum, Vaccinium oxycoccos) and from black spruce. Seeds of Carex canescens, Carex cf. aquatilis and Carex sp. were also found, which are indicative of wetter conditions. Sphagnum cf. angustifolia was identified at the end of the subzone.
Subzone Mac-Bb is distinctive because of the increase in Sphagnum (50–95%), represented by Sphagnum russowii, Sphagnum riparium and Sphagnum fuscum. Betula glandulosa, Larix laricina and Alnus alnobetula subsp. crispa were also present, but no remains of Picea mariana were found. Several ombrotrophic species such as Empetrum nigrum, Rubus chamaemorus, Rhododendron tomentosum and Vaccinium oxycoccos were present, but only in small quantities. The mixture of wet and dry tax would indicate a gradual change in local conditions towards ombrotrophic conditions.
Zone MAC 3: 9–0 cm
Remains from several ombrotrophic species were identified in this zone, including Empetrum nigrum, Vaccinium angustifolium, Rubus chamaemorus and Rhododendron tomentosum. The presence of Larix laricina, Vaccinium uliginosum, Vaccinium oxycoccos, Alnus alnobetula subsp. crispa, Carex canescens and Carex cf. aquatilis indicates that the local conditions remained humid. Sphagnum fuscum, which is indicative of ombrotrophic conditions, dominated all of the samples (95–100%), while Straminergon cf. stramineum was present in small quantities (3–5%).
Dating of peatland establishment and palsa formation in the Wiyâshâkimî Lake region
Two additional palsas were sampled for dating. The results indicate that the peatlands formed at 6290 (NW-palsa) and 5960 cal. y BP (SE-palsa) respectively, while the palsa was formed at around 170 (studied palsa) and 400 cal. y BP (SE-palsa) respectively, i.e., during the Little Ice Age (). The modern age of the peat surface of the NW-palsa () could be the result of contamination by contemporary plant materials.
Discussion
Palsa evolution
The evolution of the palsa peatland began at approximately 6300 cal. y BP. The site underwent a succession of several trophic phases: an initial pond, a minerotrophic phase, then an ombrotrophic phase. Subsequently, the palsa formed but began to collapse at the end of the Little Ice Age. The palsa degradation was followed by the creation of thermokarst ponds that filled in with vegetation ().
Figure 6. Paleoenvironmental succession of the studied palsa peatland (palsa and filled thermokarst pond) from about 6300 cal. y BP to present
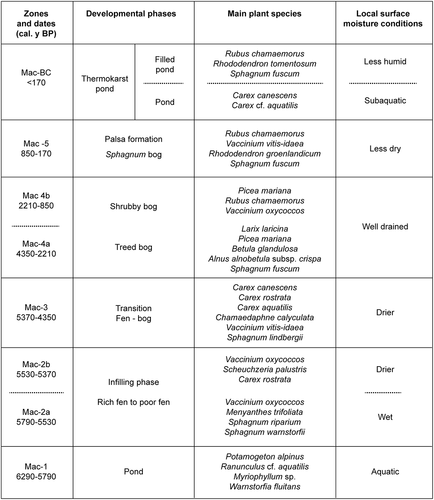
Aquatic phase and peatland initiation (6290–5790 cal. y BP)
The deglaciation of the Wiyâshâkimî Lake region occurred between 8000 and 7000 cal. y BP. At that time, the Wiyâshâkimî Lake was larger than it is today, as it would have included the study site, which is located approximately 100 m from the shore of the lake. After the level of the lake decreased, smaller lakes or ponds were left behind. The study site is one of those ponds, which is indicated by the presence of greyish clay-silty sediments at the base of the peat core. The earliest remains from aquatic and subaquatic taxa accumulated at around 6290 cal. y BP.
Peatlands were established in the Whapmagoostui-Kuujjuarapik region on the Hudson Bay coast between 6000 and 5000 cal. y BP (Arlen-Pouliot and Bhiry Citation2005; Bhiry and Robert Citation2006; Bhiry et al. Citation2011; Fillion et al. Citation2014). Peatlands also formed at the tree line on the banks of the Boniface River between 6800 and 6300 cal. y BP (Asselin and Payette Citation2006; Bhiry et al. Citation2007) and around 5310 cal. y BP further south in the James Bay lowlands (Tremblay et al. Citation2014). All of these peatlands were established in depressions that contained postglacial marine clay. At Wiyâshâkimî Lake, the studied peatland formed at roughly the same time, even though it is located further inland (150 km from the coast) in a zone that had not been submerged by the sea. Similarly, peat started to accumulate on the small island at the center of the lake at 5800 cal. yr BP (Payette Citation1988) and on Lepage Island by 4900 cal. y BP (Bhiry et al. Citation2019; ).
Fen phase and fen-bog transition phase (5790–4350 cal. y BP)
The decrease in the water level of the pond around 5800 cal. y BP favoured greater plant diversity over about a 1440-year period. Similarly, an increase in the rate of peat accumulation also occurred (0.52 mm/yr on average). The drop in water level between 5790 and 5370 cal. y BP could be linked to the isostatic uplift (Lafortune et al. Citation2006; Lavoie et al. Citation2012); in this regard, several raised lacustrine terraces were observed on islands in the region. At about 5370 cal. y BP, changes in the macrofossil assemblages indicate the transition to ombrotrophic conditions. The rapid accumulation of peat contributed to the diminished influence of the groundwater, leading to further Sphagnum growth and, in turn, a gradual ombrotrophication of the site between 5370 and 4350 cal. y BP.
Ombrotrophic phase (4350–850 cal. y BP)
During this phase, black spruce and eastern larch extensively colonized the peatland; the abundance of needles from these two species indicates the growth of forested areas on the periphery of the peatland and probably also on the peatland itself. The presence of Rhododendron groenlandicum and Rubus chamaemorus also points to ombrotrophic conditions. Between 2210 and 850 cal. y BP, trees and shrubs seem to have disappeared or significantly declined, but conditions remained ombrotrophic. The high degree of peat humification and the scarcity of identifiable plant macrofossils, combined with the significant decrease in peat accumulation, indicate well-drained conditions during this period.
This change in the local conditions is inconsistent with other studies conducted at Wiyâshâkimî Lake and elsewhere in northern Québec. According to Payette (Citation1988) and Bhiry et al. (Citation2019), the Wiyâshâkimî Lake region was characterised by an increase in atmospheric humidity at around 3900–3500 cal. y BP and cold temperatures. Similar conditions were also reported in several studies conducted in northern Quebec and Nunavik, such as a higher water table between 4200 and 3000 cal. y BP (Payette and Filion Citation1993; Lavoie and Payette Citation1995; Miousse et al. Citation2003; Bhiry and Robert Citation2006; Tremblay et al. Citation2014). On the other hand, the local disappearance of the trees from the site coincides with the findings of Payette (Citation1988), demonstrating a progressive but significant decline in spruce trees at around 2000 cal. y BP. This decline was linked to colder and drier conditions (less snow) during the Neoglacial period (Viau and Gajewski Citation2009). The disappearance of trees and shrubs at the study site would have caused a decrease in snow accumulation at the surface of the peatland, as it was more exposed to winter winds.
Return of humid conditions and subsequent palsa formation
After 850 cal. y BP, the local conditions became relatively wetter than before, as revealed by the reappearance of several species of Sphagnum and a significant increase in the peat accumulation rate. This change was likely triggered by the colder climate of the Little Ice Age, which caused an uplifting of the peat surface following permafrost growth at the site. At around 170 cal. y BP, peat accumulation either stopped or declined dramatically until today. This date should be considered maximal, as it is possible that in some cases the uppermost peat was oxidized or eroded by drifting snow and ice crystals (Seppälä Citation1983). The Sphagnum peat would have provided a cover that served to preserve the permafrost, acting as insulation against the heat in the summer and as a conductor of cold air in winter (Couillard and Payette Citation1985; Bhiry and Robert Citation2006). This is quite different from the data collected from the permafrost peatland covering the small island at the center of the lake (Payette Citation1988). In fact, permafrost aggradation and ice wedge development on the surface of the peatland followed the progressive removal of spruce at around 2000 cal. y BP (Payette et al. Citation1986). The deforestation of the site would have induced a major decrease in snow accumulation and exposed the surface of the peatland to cold winds. This means that autogenic factors were the primary cause of permafrost aggradation.
With regard to the peatlands located on the coasts of northern Quebec (subarctic and high-boreal regions), the formation of palsas would have occurred during a different period of the Holocene, depending on the location of the site (Payette Citation2009; Bhiry and Lavoie Citation2018). For example, in the Boniface River region at the northern limit of the discontinuous permafrost zone, the early formation dates back to 3000 cal. y BP and coincides with the Neoglacial cooling; palsa development resumed during the LIA (Asselin and Payette Citation2006; Bhiry and Lavoie Citation2018). Further south in Whapmagoostui-Kuujjuarapik (Hudson Bay), the formation of palsas occurred during the Little Ice Age, between 600 and 340 cal. y BP. In James Bay, 300 km to the south, the formation of palsas also occurred just before the end of the Little Ice Age (Tremblay et al. Citation2014).
Palsa degradation and formation of thermokarst ponds
The recent climate warming, which began in the 20th century, has caused the degradation of permafrost in palsas and induced the formation of thermokarst ponds (Sollid and Sorbel Citation1998; Zuidhoff and Kolstrup Citation2000; Payette et al. Citation2004; Arlen-Pouliot and Bhiry Citation2005; Thibault and Payette Citation2009; Tremblay et al. Citation2014). The collapse of palsas on the coast of Hudson Bay and James Bay (e.g., Arlen-Pouliot and Bhiry Citation2005; Bhiry et al. Citation2011; Fillion et al. Citation2014; Tremblay et al. Citation2014) and in the Wiyâshâkimî Lake region (this study) began at the end of the Little Ice Age, leading to the formation of thermokarst ponds. In the core of the FTP, the transition from palsa to filled thermokarst pond was dated to 360 cal. y BP, which is earlier than the formation of the palsa (170 cal. y BP). This inconsistency may have several explanations, including: (i) the contamination of dated samples, either by older organic material (FTP sample) or by newer organic material (palsa sample), despite our taking precautions to avoid this; (ii) the presence of a previous cycle of permafrost aggradation and degradation at the site, prior to the present-day palsa (although there was no evidence to support this justification, such as the alternation of xerophilic peat and wet peat); and (iii) the differential uplifting and the differential collapse of the peatland surface. This last explanation is supported by the fact that the current peatlands in subarctic regions are formed by different features: palsas, thermokarst ponds and filled thermokarst, minerotrophic areas. In any case, the macrofossil record indicates that the thermokarst pond filled in rapidly, given that the transition from subaquatic environment to poorly minerotrophic peatland occurred over 450 years. The rate of peat accumulation in the filled thermokarst pond (1.9 mm/yr) was as high as was reported in other studies (Robinson and Moore Citation2000; Arlen-Pouliot and Bhiry Citation2005; Coulombe et al. Citation2016). Payette et al. (Citation2004) had similar findings in their spatio-temporal study of the 50-year evolution of a subarctic palsa peatland. The presence of thermokarst ponds was associated with the rapid accumulation of peat through natural infilling processes.
Similarities and differences between peatlands in the Wiyâshâkimî Lake region and in Nunavik
Several studies have found that subarctic palsa peatlands and high boreal peatlands follow a similar hydroseral development: an initial pond transitioned to a minerotrophic peatland, then an ombrotrophic peatland, which then transformed into a palsa after the establishment of permafrost (Lavoie and Payette Citation1995; Fillion et al. Citation2014; Arlen-Pouliot and Bhiry Citation2005; Bhiry and Robert Citation2006; Tremblay et al. Citation2014). This general pattern also applies to our study site. The Boniface River site is an exception because it lacked an ombrotrophic stage. This difference may be accounted for by the flooding that occurred at the Boniface River site as a result of it being situated between two lakes, Bhiry et al., Citation2007. Despite this general similarity, the duration and timing of each trophic phase was quite different at each site. For example, the peatland palsa in the studied site included a very brief minerotrophic phase (420 years: 5790–5370 cal. y BP), while the coastal peatlands had a much longer minerotrophic phase of almost 3000 years. This difference may be explained by the rapid accumulation of peat at Wiyâshâkimî Lake that caused early ombrotrophication (5000 cal. y BP).
On the other hand, the three studies conducted on peatlands located in the Wiyâshâkimî Lake region (Payette Citation1988; Bhiry et al. Citation2019 and this study) showed that each of the sites evolved differently in terms of their trophic, ecological and geomorphological conditions. In fact, the permafrost peatland covering the central island of the lake was composed of ombrotrophic peat from its onset (5050 cal. y BP) to the present day. It is formed of alternating layers of Sphagnum and spruce peat. This long-term cyclical replacement was made possible by the nutrient-poor and cold peaty soils. Other factors supporting this process included the location of this small site at the centre of such a vast lake, which prevented the establishment of diversified flora taxa, and the influence of fire (Payette Citation1988). Finally, permafrost aggradation at the site would have been triggered by site deforestation and a decrease in snow cover. In the peatland located on Lepage Island (Caribou peatland; ), local conditions remained wet and minerotrophic with an absence of permafrost. This infrequent peatland succession was attributed to its geomorphic setting downstream of a debris-covered talus slope; the continuous inflow of water and sand contributed to the maintenance of minerotrophic conditions.
Conclusion
The macrofossil data from a permafrost peatland on the north bank of Wiyâshâkimî Lake was used to reconstruct the temporal evolution of trophic conditions and plant populations. Five principle stages were identified: a shallow pond (from 6290 cal. y BP), a minerotrophic peatland, an ombrotrophic peatland, the formation of a palsa at around 170 cal. y BP, and the subsequent degradation of the palsa. The macrofossil data from the filled thermokarst pond demonstrated that the trophic evolution of the site occurred very rapidly, shifting from pond to fen then to a bog over a 450-year period. The recent and present climate conditions induced rapid peat accumulation and high carbon accumulation rates (Piilo et al. Citation2019), although the trophic conditions of the environment would also have played a role in this process.
The development of a peatland palsa at Wiyâshâkimî Lake was compared with several peatland palsas in Hudson Bay and James Bay in order to determine whether the local maritime climate on the coast influenced its development. Our data indicate that the Wiyâshâkimî Lake peatland evolved in a similar manner as the coastal peatlands. The following phases of development were identified: pond, minerotrophic, ombrotrophic, and palsa. However, these phases were asynchronous with neighbouring peatlands. The peatland palsa at Wiyâshâkimî Lake was significantly influenced by the rapid rate of peat accumulation and entered the ombrotrophic peat stage earlier. The process was significantly influenced by autogenic factors, notably the production of organic matter that led to peat accumulation.
Supplemental Material
Download TIFF Image (6.1 MB)Acknowledgments
Thanks are extended to Y. Bégin, L. Noël and D. Sarrazin for fieldwork assistance and sampling the peat cores, to M. Bourgon Desroches, G. Filteau, J. Lebrun and G. Labrecque for laboratory assistance and to L. Burns for editing and insightful comments on the manuscript. We are also grateful to Annie Novalinga and Véronique Nadeau from Tursujuq National Park, the community of Umiujaq and to the Centre d’études nordiques for logistics and support.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
Related Research Data
References
- Abbott BW, Jones JB, Schuur EA, Chapin FS, Bowden WB, Bret-Harte MS, Epstein HE, Flannigan MD, Harms TK, Hollingsworth TN, et al. 2016. Biomass offsets little or none of permafrost carbon release from soils, streams, and wildfire: an expert assessment. Environ Res Lett. 11(3):034014.
- Allard M, Seguin MK. 1987. La déglaciation d’une partie du versant hudsonien québécois: bassins des rivières Nastapoca, Sheldrake et à l’Eau Claire. Géographie physique et Quaternaire. 39(1):13–24. doi:https://doi.org/10.7202/032581ar.
- Arlen-Pouliot Y, Bhiry N. 2005. Paleoecology of a palsa and a filled thermokarst pond in a permafrost peatland, subarctic Québec, Canada. Holocene. 15(3):408–419. doi:https://doi.org/10.1191/0959683605hl818rp.
- Asselin H, Payette S. 2006. Origin and long-term dynamics of a subarctic tree line. Écoscience. 13(2):135–142. doi:https://doi.org/10.2980/i1195-6860-13-2-135.1.
- Avramtchev L. 1982. Catalogue des gîtes minéraux du Québec, région de la baie d’Hudson, Québec. Québec (QC): Ministère de l’Énergie et des Ressources, Service de la géoinformation. DPV-926. p. 13.
- Ayotte G, Rochefort L. 2014. Les sphaignes du Québec, du Labrador et des Maritimes (à l’exception de l’Île de Terre-Neuve). Clés visuelles d’identification. Québec (QC): Université Laval. 209 p.
- Bégin Y. 1986. Dynamique de la végétation riveraine du lac à l'Eau-Claire, Québec subarctique. Ph.D. thesis. Québec: Université Laval.
- Bhiry N, Filion L. 2001. Analyse des macrorestes végétaux. In: Payette S, Rochefort L, editors. Écologie des tourbières du Québec-Labrador. Québec (QC): Presses de l’Université Laval; p. 259–273.
- Bhiry N, Lavoie M. 2018. Timeline of permafrost peatland formation in subarctic Quebec (Canada). In: Deline P, Bodin X, Ravanel L, Barboux C, Delaloye R, Lambiel C, Magnin F, Pogliotti P, Schoeneich P, editors. 5th European Conference on Permafrost Book of Abstracts; Jun 23–Jul 1; Chamonix, France. p. 687–688.
- Bhiry N, Decaulne A, Bourgon-Desroches M. 2019. Development of a subarctic peatland linked to slope dynamics at Lac Wiyâshâkimî (Nunavik, Canada). Holocene. 29(9):1459–1467. doi:https://doi.org/10.1177/0959683619854516.
- Bhiry N, Delwaide A, Allard M, Bégin Y, Filion L, Lavoie M, Nozais C, Payette S, Pienitz R, Saulnier-Talbot É, et al. 2011. Environmental change in the Great Whale River region, Hudson Bay: five decades of multidisciplinary research by Centre d’études nordiques (CEN). Écoscience. 18(3):182–203. doi:https://doi.org/10.2980/18-3-3469.
- Bhiry N, Payette S, Robert É. 2007. Peatland development at the arctic tree line (Québec, Canada) influenced by flooding and permafrost. Quat Res. 67(9):426–437. doi:https://doi.org/10.1016/j.yqres.2006.11.009.
- Bhiry N, Robert É. 2006. Reconstruction of changes in vegetation and trophic conditions of a palsa in a permafrost peatland, subarctic Québec, Canada. Écoscience. 13(1):56–65. doi:https://doi.org/10.2980/1195-6860(2006)13[56:ROCIVA]2.0.CO;2.
- Blondeau M, Cuerrier A, Roy C. 2010. Atlas des plantes des villages du Nunavik. 2e éd., Institut culturel Avataq. Sainte-Foy (QC): Éditions Multimondes. 737 p.
- Brouillet L, Coursol F, Meades SJ, Favreau M, Anions M, Bélisle P, Desmet P. 2010+. VASCAN, la Base de données des plantes vasculaires du Canada. http://data.canadensys.net/vascan/vernacular/25475.
- Calmels F, Gagnon O, Allard M. 2005. A portable earth-drill system for permafrost studies. Permafrost Periglac. 16(3):311–315. doi:https://doi.org/10.1002/ppp.529.
- Couillard L, Payette S. 1985. Évolution holocène d’une tourbière à pergélisol (Québec nordique). Can J Bot. 63(6):1104–1121. doi:https://doi.org/10.1139/b85-152.
- Coulombe O, Bouchard F, Pienitz R. 2016. Coupling of sedimentological and limnological dynamics in subarctic thermokarst ponds in Northern Québec (Canada) on an interannual basis. Sediment Geol. 340(S1):15–24. doi:https://doi.org/10.1016/j.sedgeo.2016.01.012.
- Decaulne A, Bhiry N, Lebrun J, Veilleux S, Sarrazin D. 2018. Geomorphic evidence of Holocene slope dynamics on the Canadian shield – a study from Lac à l’Eau-Claire, western Nunavik. Écoscience. 25(4):343–357. doi:https://doi.org/10.1080/11956860.2018.1431376.
- Deshaye J, Morisset P. 1985. La flore vasculaire du lac à l’Eau Claire, Nouveau Québec. Provancheria 18. Québec (QC): Presses de l’Université Laval. 52 p.
- Faubert J. 2013. Flore des bryophytes du Québec-Labrador. Vol. 2, Mousses, première partie. Saint-Valérien (QC): Société québécoise de bryologie. 402 p.
- Faubert J. 2014. Flore des bryophytes du Québec-Labrador. Vol. 3, Mousses, deuxième partie. Saint-Valérien (QC): Société québécoise de bryologie. 456 p.
- Fillion M-È, Bhiry N, Touazi M. 2014. Differential development of two palsa fields in a peatland located near Whapmagoostui-Kuujjuarapik, Northern Québec, Canada. Arct Antarct Alp Res. 46(1):40–54. doi:https://doi.org/10.1657/1938-4246-46.1.40.
- Garneau M. 2001. Les taxons préférentiels des tourbières (Annexe 1). In: Payette S, Rochefort L, editors. Écologie des tourbières du Québec-Labrador. Québec (QC): Presses de l’Université Laval; p. 523–532.
- Juggins S. 2002. Palaeo Data Plotter. Beta test version 1.0. Newcastle upon Tyne: University of Newcastle.
- Kuhry P. 2008. Palsa and peat plateau development in the Hudson Bay Lowlands, Canada: timing, pathways and causes. Boreas. 37(3):316–327. doi:https://doi.org/10.1111/j.1502-3885.2007.00022.x.
- Lafortune V, Filion L, Hétu B. 2006. Émersion des terres et développement des sols bien drainés au lac Guillaume-Delisle, Québec subarctique. Géographie physique et Quaternaire. 60(2):165–181. doi:https://doi.org/10.7202/016827ar.
- Lamarre A, Garneau M, Asnong H. 2012. Holocene paleohydrological reconstruction and carbon accumulation of a permafrost peatland using testate amoeba and macrofossil analyses, Kuujjuarapik, subarctic Québec, Canada. Rev Palaeobot Palynol. 186:131–141. doi:https://doi.org/10.1016/j.revpalbo.2012.04.009.
- Laprise D, Payette S. 1988. Évolution récente d’une tourbière à palses (Québec subarctique): analyse cartographique et dendrochronologique. Can J Bot. 66(6):2217–2227. doi:https://doi.org/10.1139/b88-304.
- Lavoie C, Allard M, Duhamel D. 2012. Deglaciation landforms and C-14 chronology of the Lac Guillaume-Delisle area, eastern Hudson Bay: a report on field evidence. Geomorphology. 159:142–155. doi:https://doi.org/10.1016/j.geomorph.2012.03.015.
- Lavoie C, Payette S. 1995. Analyse macrofossile d’une palse subarctique (Québec nordique). Can J Bot. 73(4):527–537. doi:https://doi.org/10.1139/b95-054.
- Marie-Victorin F. 1995. La flore laurentienne. 3e éd. Montréal (QC): Presses de l’Université de Montréal. 1093 p.
- Matveev A, Laurion I, Deshpande BN, Bhiry N, Vincent WF. 2016. High methane emissions from thermokarst lakes in subarctic peatlands. Limnol Oceanogr. 61(S1):S150–S164. doi:https://doi.org/10.1002/lno.10311.
- Miousse L, Bhiry N, Lavoie M. 2003. Isolation and water level fluctuations of lake Kachishayoot, northern Quebec, Canada. Quat Res. 60(2):149–161. doi:https://doi.org/10.1016/S0033-5894(03)00094-2.
- Moore TR, Roulet NT. 1993. Methane flux: water table relations in northern wetlands. Geophys Res Lett. 20(7):587–590. doi:https://doi.org/10.1029/93GL00208.
- Payette S. 1983. The forest tundra and present tree-lines of the northern Québec-Labrador peninsula. Nordicana. 47:3–23.
- Payette S. 1988. Late-Holocene development of subarctic ombrotrophic peatlands: allogenic and autogenic succession. Ecology. 69(2):516–531. doi:https://doi.org/10.2307/1940450.
- Payette S. 2001. Les processus et les formes périglaciaires. In: Payette S, Rochefort L, editors. Écologie des tourbières du Québec-Labrador. Québec (QC): Presses de l’Université Laval; p. 199–240.
- Payette S. 2009. Rise and fall of palsas and peat plateaus in eastern Canada. In: Fronzek S, Johansson M, Christensen TR, Carter TR, Thomas Friborg T, Miska Luoto M, editors. Climate change impacts on sub-arctic palsa mires and greenhouse gas feedbacks. Proceedings of the PALSALARM symposium. Abisko, Sweden. p. 56–58.
- Payette S, Delwaide A, Caccianiga M, Beauchemin M. 2004. Accelerated thawing of subarctic peatland permafrost over the last 50 years. Geophys Res Lett. 31(18):L18208. doi:https://doi.org/10.1029/2004GL020358.
- Payette S, Filion L. 1993. Holocene water-level fluctuations of a subarctic lake at the tree-line in northern Québec. Boreas. 22(1):7–14. doi:https://doi.org/10.1111/j.1502-3885.1993.tb00159.x.
- Payette S, Gauthier L, Grenier I. 1986. Dating ice-wedge growth in subarctic peatland following deforestation. Nature. 322(6081):724–727. doi:https://doi.org/10.1038/322724a0.
- Piilo SR, Zhang H, Garneau M, Gallego-Sala A, Amesbury MJ, Väliranta MM. 2019. Recent peat and carbon accumulation following the Little Ice Age in northwestern Québec, Canada. Environ Res Lett. 14:075002. doi:https://doi.org/10.1088/1748-9326/ab11ec.
- Reimer PJ, Austin WE, Bard E, Bayliss A, Blackwell PG, Ramsey CB, Butzin M, Cheng H, Edwards RL, Friedrich M, et al. 2020. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon. 62(4):725–757. doi:https://doi.org/10.1017/RDC.2020.41.
- Richard PJH. 1981. Paléophytogéographie postglaciaire en Ungava par l’analyse pollinique. Paléo-Québec 13. Montréal (QC): Recherches amérindiennes au Québec. 153 p.
- Robinson SD, Moore TR. 2000. The influence of permafrost and fire upon carbon accumulation in high boreal peatlands, Northwest Territories, Canada. Arct Antarct Alp Res. 32(2):155–166. doi:https://doi.org/10.1080/15230430.2000.12003351.
- Rousseau C. 1974. Géographie floristique du Québec-Labrador: distribution des principales espèces vasculaires. Québec (QC): Presses de l’Université Laval. 798 p.
- Schmieder M, Schwarz WH, Trieloff M, Tohver E, Buchner E, Hopp J, Osinski GR. 2015. New Ar-40/Ar-39 dating of the Clearwater Lake impact structures (Québec, Canada) – not the binary asteroid impact it seems? Geochim Cosmochim Ac. 148:304–324. doi:https://doi.org/10.1016/j.gca.2014.09.037.
- Seppälä M. 1983. Surface abrasion of palsas by wind action in Finnish Lapland. Geomorphology. 52(1–2):141–148. doi:https://doi.org/10.1016/S0169-555X(02)00254-4.
- Seppälä M. 1986. The origin of palsas. Geogr Ann. 68A:141–147. doi:https://doi.org/10.1080/04353676.1986.11880167.
- Simard M, Parent M, Thériault R, David J, Lacoste P, Sharma KNM. 2004. Géologie de la région du lac à l’Eau Claire (34B et 34C). Québec (QC): Ministère des ressources naturelles. 46 p.
- Sollid JL, Sorbel L. 1998. Palsa bogs as a climate indicator: examples from Dovrefjell, southern Norway. Ambio. 27(4):287–291.
- Swindles GT, Amesbury MJ, Turner TE, Carrivick JL, Woulds C, Raby C, Mullan D, Roland TP, Galloway JM, Parry L, et al. 2015. Evaluating the use of testate amoebae for palaeohydrological reconstruction in permafrost peatlands. Palaeogeogr Palaeoclimatol Palaeoecol. 424:111–122. doi:https://doi.org/10.1016/j.palaeo.2015.02.004.
- Thibault S, Payette S. 2009. Recent permafrost degradation in bogs of the James Bay area, northern Quebec, Canada. Permafrost Periglac. 20(4):383–389. doi:https://doi.org/10.1002/ppp.660.
- Treat CC, Jones MC, Camill P, Gallego-Sala A, Garneau M, Harden JW, Hugelius G, Klein ES, Kokfelt U, Kuhry P, et al. 2016. Effects of permafrost aggradation on peat properties as determined from a pan-Arctic synthesis of plant macrofossils. J Geophys Res Biogeosci. 121:78–94. doi:https://doi.org/10.1002/2015JG003061.
- Tremblay S, Bhiry N, Lavoie M. 2014. Long-term dynamics of a palsa in the sporadic permafrost zone of northwestern Quebec (Canada). Can J Earth Sci. 51(5):500–509. doi:https://doi.org/10.1139/cjes-2013-0123.
- Viau AE, Gajewski K. 2009. Reconstructing millennial-scale, regional paleoclimates of boreal Canada during the Holocene. J Climate. 22(2):316–330. doi:https://doi.org/10.1175/2008JCLI2342.1.
- Zoltaï SC. 1993. Cyclic development of permafrost in the peatlands of northwestern Alberta, Canada. Arct Alp Res. 25(3):240–246. doi:https://doi.org/10.2307/1551820.
- Zoltaï SC. 1995. Permafrost distribution in peatlands of west-central Canada during the Holocene warm period 6000 years BP. Géographie physique et Quaternaire. 49(1):45–54. doi:https://doi.org/10.7202/033029ar.
- Zoltaï SC, Tarnocai C. 1971. Properties of a wooded palsa in northern Manitoba. Arct Antarct Alp Res. 3(2):115–129. doi:https://doi.org/10.2307/1549981.
- Zuidhoff FS, Kolstrup E. 2000. Change in palsa distribution in relation to climate change in Laivadalen northern Sweden, especially 1960-1997. Permafrost Periglac. 11(2):55–69. doi:https://doi.org/10.1002/(SICI)1099-1530(200001/03)11:1<55::AID-PPP338>3.0.CO;2-T.