
ABSTRACT
Caribbean islands are important hotspots for endemic species, concentrating high biodiversity on exceptionally small areas. Cozumel Island is not an exception, yet little published information exists on its bird diversity. In this study, we assessed the contribution of each major ecosystem of Cozumel Island (i.e., tropical forest, mangrove, palm thicket, and urban setting) to the island’s avian diversity. Avian richness was highest in tropical forests, followed by mangroves, palm thickets, and urban setting. Bird densities were highest in urban settings, followed by tropical forests, mangroves, and palm thickets. Species composition similarity among ecosystems was low, with the most dissimilar ecosystem being the urban setting. Interestingly, we recorded the highest number of unique species in mangroves, followed by tropical forests, urban settings, and palm thickets. Tropical forests and mangroves of Cozumel Island are the main ecosystems harboring high avian diversity, with tropical forests housing high avian densities and endemics and mangroves including the most unique species. As expected, the contribution of urban settings to the avian diversity of Cozumel Island is limited and includes some highly abundant invasive species, of which the Eurasian Collared-Dove could represent a major ecological threat.
RÉSUMÉ
Les îles des Caraïbes sont d’importants points chauds pour les espèces endémiques, concentrant une biodiversité élevée dans des régions exceptionnellement petites. L’île de Cozumel ne fait pas exception, mais peu d’information existe sur sa diversité en oiseaux. Dans cette étude, nous avons évalué la contribution à la diversité aviaire de chacun des principaux écosystèmes de l’île de Cozumel (i.e., forêt tropicale, mangrove, fourré de palmiers, zone urbanisée). La richesse aviaire était plus élevée dans les forêts tropicales, suivies des mangroves, des fourrés de palmiers et des zones urbanisées. Les densités d’oiseaux étaient plus élevées dans les zones urbanisées, suivies des forêts tropicales, des mangroves et des fourrés de palmiers. La composition spécifique entre les écosystèmes avait une faible similarité; les écosystèmes urbanisés étant les plus dissimilaires. Nous avons enregistré le nombre le plus élevé d’espèces uniques dans les mangroves, suivies des forêts tropicales, des zones urbanisées et des fourrés de palmiers. Les forêts tropicales et les mangroves de l’île de Cozumel sont les principaux écosystèmes arborant une diversité aviaire élevée. Les forêts tropicales ont des densités aviaires élevées et abritent des espèces endémiques, tandis que les mangroves ont le plus d’espèces uniques. Tel qu’attendu, la contribution des zones urbaines à la diversité aviaire de l’île de Cozumel est limitée et inclut certaines espèces invasives très abondantes, parmi lesquelles la tourterelle turque, qui pourrait représenter une menace écologique majeure.
Introduction
Given the amount and type of biodiversity they shelter, insular systems are considered hotspots of conservation, with endemic species representing an important proportion of their diversity (Myers et al. Citation2000). The high diversity on islands has often been related to their size, mainly because insular areas have been shown to correlate with more diverse and heterogeneous environments (MacArthur and Wilson Citation1967). Thus, the wildlife diversity of an island is, at least partially, driven by its size and consequent habitat diversity (Scherber et al. Citation2018). Other factors such as isolation, commonly measured by the relative distance to other islands and the mainland, have been shown to determine the diversity of an island (Simberloff Citation1974; Stark et al. Citation2006; Blackburn et al. Citation2016). One important factor related to insular systems is that large and isolated islands have faster speciation rates than small islands closer to the mainland, which is why islands tend to have high endemism (Rosenzweig Citation1975). This is of special concern due to the anthropogenic pressures that are currently affecting these fragile ecosystems (Whittaker and Fernández-Palacios Citation2007).
Human activities have shaped insular ecosystems in a similar fashion as has happened inland (Lugo et al. Citation2012). In particular, the development of mass tourism related to sea, sun, and sand amenities has been shown to represent an important threat to biodiversity (e.g., due to the development of resorts, the use of resources for buildings, transport, pollution by vehicle emissions, and litter), with important damages to coastal ecosystems (Davenport and Davenport Citation2006). Also, the presence of roads has a negative relationship with avian richness and abundance in different tropical regions, causing increased vehicle mortality, behavioral changes due to traffic noise, and increased hunting rates by humans that take advantage of these roads (Pinto et al. Citation2020). Development of tourism can increase the rates of beach erosion and promote human activities in all forms, affecting habitat availability, productivity, and biodiversity (Yasué and Dearden Citation2006). Additionally, hurricanes are an important natural force that has been shown to mold the structure of bird communities on islands, with direct detrimental effects (i.e., individual mortality or geographic displacement), as well as indirect ones (e.g., reduction of food supplies, loss of nests and roost sites, increase in vulnerability to predation, and microclimate change; Wiley and Wunderle Citation1993). Moreover, the presence of introduced invasive species such as the boa (Boa constrictor) has been identified as a threat to a broad variety of native species on islands, including endemics (Martínez-Morales and Cuarón Citation1999). The synergy of disasters including hurricanes or cyclones together with anthropogenic activities is an important trigger for the decline of endemic and endangered insular species (Perdomo-Velázquez et al. Citation2017).
Birds have been widely studied on islands, providing an important model to understand the ecology of such isolated systems (Blondel et al. Citation1988). Birds inhabiting islands are not exempt from the many anthropogenic and naturally related hazards (Webster et al. Citation2005). In response to human disturbances, bird populations have decreased or even disappeared from islands (Macouzet and Escalante Citation2001). Yet, some insular bird species have been shown to be able to overcome hazards and use human-disturbed habitats and related resources. In particular, synanthropic species have been shown to increase their abundances in highly disturbed areas, with an important number of species concentrating at intermediate-disturbance levels and in edge systems (Lugo et al. Citation2012; Irizarry et al. Citation2016; Titoko et al. Citation2019). Furthermore, native vegetation on islands (e.g., mangroves and forests) do not only harbor important avian diversity but are often large-scale sources of diversity, mitigating some of the effects of human activities (Khimoun et al. Citation2017; Wang et al. Citation2017; Titoko et al. Citation2019).
Although Caribbean islands are considered important hotspots for endemic species, concentrating high biodiversity on exceptionally small areas, they are among the most threatened ecosystems worldwide (Myers et al. Citation2000). Despite the existence of bird studies in Cozumel Island (Buffa and Morris Citation1999; Howell Citation2004; Martínez-Morales et al. Citation2009; Sosa-López and Mennill Citation2014; Morales-Contreras et al. Citation2019), there is little published information regarding bird diversity and distribution across Cozumel Island. In this study, we contrasted ecological snapshots of each major ecosystem of Cozumel Island (i.e., tropical forest, mangrove, palm thicket, and urban setting) to assess their contribution to the island’s avian diversity. Considering the relevance and uniqueness of mangroves, forests, and palm thickets on islands (Khimoun et al. Citation2017; Wang et al. Citation2017; Titoko et al. Citation2019), we predicted that the bird assemblages in these ecosystems would be more diverse and unique than those from urban settings.
Material and methods
Study area
We conducted this study on Cozumel Island (which means ‘Land of Swallows’ in Maya; i.e., Kosom Lumil), a system of coral origin located in front of the northeastern coast of the Yucatan Peninsula, in the state of Quintana Roo, Mexico (20°30ʹ13”N 86°56ʹ26”W; ~1 m asl). The island was formed between the Oligocene and Pleistocene, with no recognized land bridge, so it is classified as oceanic (Weidie Citation1985). It is practically flat, without geographical barriers or superficial bodies of water, except for some cenotes (natural wells in limestone soil). Cozumel Island is the largest island in the Mexican Caribbean, with an approximate territory of 478 km2. Human population in the island is ~86,000 inhabitants, most of whom live in San Miguel de Cozumel. The main economic activity in the island is tourism, which is concentrated on its western coast. The island receives ~50,000 tourists monthly (INEGI Citation2017). Agriculture, while important in the past, is now of minor importance.
Cozumel Island is a priority area for biodiversity conservation, being the Mexican island with the highest number of endemic taxa (UNESCO Citation2016). However, these endemics are subject to several threats, such as the introduction of potentially invasive species, genetic introgression, the expansion of the road system, loss and fragmentation of native vegetation, human population growth, and tourism development, which bring changes in land use (Cuarón Citation2009). In addition, hurricanes are natural disturbances with a high capacity to modify the previous natural conditions, representing an important threat to the island (Wiley and Wunderle Citation1993).
Cozumel Island has over 500 plant species, representing 40% of the vegetation of the State of Quintana Roo (Téllez-Valdés and Cabrera Citation1987). Dominant vegetation types across the island are tropical forests (both deciduous and semi-deciduous) composed mainly of two arboreal strata of 8–20 m and a scarce shrub and herbaceous strata. After tropical forests, mangroves (5–10 m high) follow. Finally, palm thickets (3–10 m high) are scattered across the island. Most of the plant associations on the island are considered as secondary vegetation, as they often replace tropical forests when disturbed (both naturally and anthropogenically; Téllez-Valdés et al. Citation1989; Cuarón Citation2009; Vázquez-Domínguez and Espindola Citation2013) and coastal dune vegetation (Collantes-Chávez-Costa et al. Citation2019). The dominant plant species in each ecosystem are as follows: Acacia gaumeri, Leucaena leucocephala, Pithecellobium dulce, Caesalpinia platyloba, Cassia racemosa, Cordia sebestena, Guaiacum sanctum, Capparis incana, Colubrina arborescens, Bursera simaruba, Guazuma ulmifolia, Coccoloba cozumelensis, Cedrella mexicana, and Randia longiloba in deciduous forests; Rhizophora mangle and Avicennia germinans in mangroves; and Thrinax radiata and Acoelorrhaphe wrightii in palm thickets (Flores-Guido Citation1983).
Bird surveys
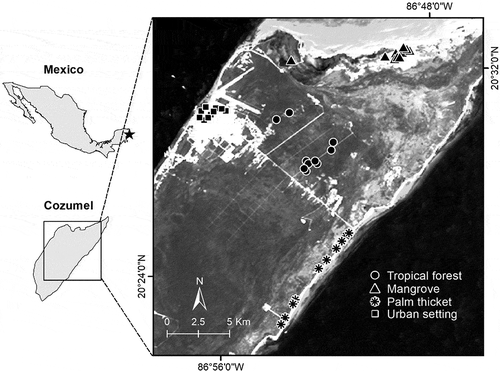
We carried out standardized bird surveys in the four main ecosystems on the island (i.e., mangrove, tropical forest, palm thicket, and urban setting; ) in August 2019. We placed 10 point-counts (50 m radius and 10 min) in each ecosystem. Briefly, point-counts consist of fixed circular areas, often with a defined radius and during a limited period of time, in which birds are recorded (both visually and auditory). We established the location of all point-counts following the presence of the ecosystems of interest and a criterion of accessibility. We placed points at least 200 m from each other to avoid recording the same individuals twice. We performed surveys from sunrise up to four hours after (following Ralph et al. Citation1996). At each point-count location, we recorded all birds seen or heard through a double-observer approach (sensu Nichols et al. Citation2000). Both observers were present during all the survey points. For each count, the ‘primary’ observer (IM-F) indicated the location of detected birds to the ‘secondary’ observer (MAG-M) for him to record distances between them and the recorded individuals, using a laser rangefinder (Leupold RX-650). Notably, the ‘secondary’ observer also alerted the ‘primary’ one about non-detected bird individuals, increasing the overall bird detection and allowing for more precise estimates of avian presence and densities (Nichols et al. Citation2000). This approach was suitable, considering the structure of vegetation in Cozumel Island, which can hamper the detection ability of only one observer. Therefore, we followed this approach with the aim of increasing avian detectability.
Figure 1. Study area and survey sites on Cozumel Island indicating point-count distribution in each ecosystem type.
Data analysis
We contrasted the avian species richness values for each studied ecosystem using rarefaction procedures with EstimateS 9, which is based on a re-sampling framework that outputs interpolated species richness statistical expectations (Colwell Citation2013). We also calculated distance-corrected densities for all birds recorded at each condition (Alldredge et al. Citation2007), using Distance 6.2. Distance calculates the number of individuals per hectare based on the probability of detection at increasing distances from the observer, afterwards standardizing detection rates along concentric surveyed distances (Buckland et al. Citation2001; Thomas et al. Citation2010). For species richness and density comparisons among conditions, we contrasted 84% confidence intervals and considered statistical differences with an α = 0.05 when confidence intervals did not overlap (following MacGregor-Fors and Payton Citation2013). We used 84% confidence intervals as 95% confidence intervals fail to indicate statistical differences with α = 0.05 (MacGregor-Fors and Payton Citation2013). We finally performed a multivariate Bray–Curtis cluster analysis (average linkage) using the package vegan (Oksanen et al. Citation2016) in R (R Development Core Team Citation2019) to assess differences in species composition among the birds of the studied ecosystems.
Results
We recorded a total of 50 species across our surveys (). Tropical forests were dominated by the Black Catbird (Melanoptila glabrirostris), Yellow-faced Grassquit (Tiaris olivaceus), and Yellow Warbler (Setophaga petechia). The most abundant species recorded in mangroves were the Yellow Warbler, White-Crowned Pigeon (Patagioenas leucocephala), and Reddish Egret (Egretta rufescens). Palm thickets were dominated by the Tropical Mockingbird (Mimus gilvus), Barn Swallow (Hirundo rustica), and Great-Tailed Grackle (Quiscalus mexicanus). Finally, the most abundant species recorded in urban settings were the Rock Pigeon (Columba livia), Great-tailed Grackle, and Eurasian Collared-Dove (Streptopelia decaocto).
Table 1. Bird species recorded in Cozumel Island, ordered in agreement with the AOS Checklist of North and Middle American Birds (Chesser et al. Citation2020). Conservation status: NOM = SEMARNAT (Citation2010, Citation2019 update); IUCN = IUCN (Citation2019); Pr = Subject to special protection; A = Threatened; NT = Near threatened. Endemism: EN = Endemic, QE = Quasi-endemic, SE = Semi-endemic (Berlanga et al. Citation2019)
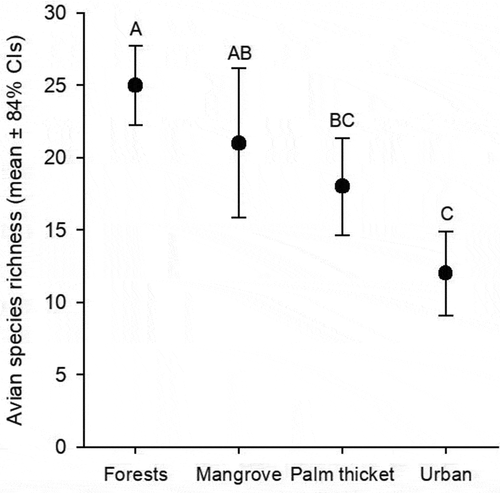
Avian species richness showed to be significantly higher in tropical forests (25 ± 2.8 species), not being statistically different to that recorded in mangroves (21 ± 5.2 species). In turn, species richness in palm thickets was significantly lower than in tropical forests, but not statistically different from that of mangroves (18 ± 3.4 species). Finally, urban settings had the lowest species richness (12 ± 2.9 species), showing significant differences with tropical forests and mangroves, but not with palm thickets ().
Figure 2. Rarefied bird species richness (mean ± 84% confidence intervals) of the studied ecosystems on Cozumel Island. Letters above error bars represent statistical differences between forests, mangroves, palm thickets, and urban settings.
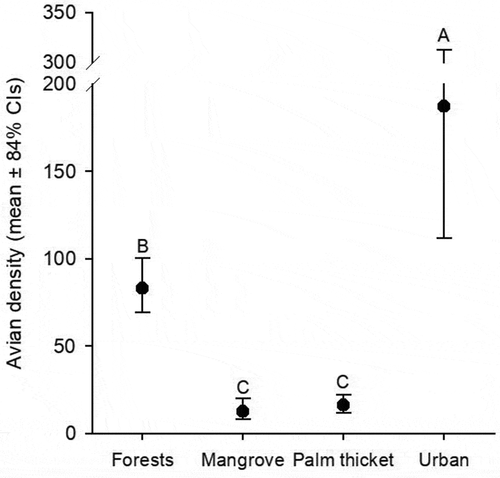
We recorded a significantly higher density in urban settings (187 ind/ha, 84% CIs: 112.0–313.0) compared to the other studied ecosystems. Following urban settings, tropical forests had a statistically higher avian density (83 ind/ha, 84% CIs: 69.4–100.4) than mangroves (12.7 ind/ha, 84% CIs: 8.1–20.2) and palm thickets (16.1 ind/ha, 84% CIs: 11.7–22.2), which did not statistically differ ().
Figure 3. Estimated avian density (mean ± 84% confidence intervals) of the four studied ecosystems on Cozumel Island. Letters above error bars represent statistical differences. The scale-break in the y-axis is used for visual purposes.
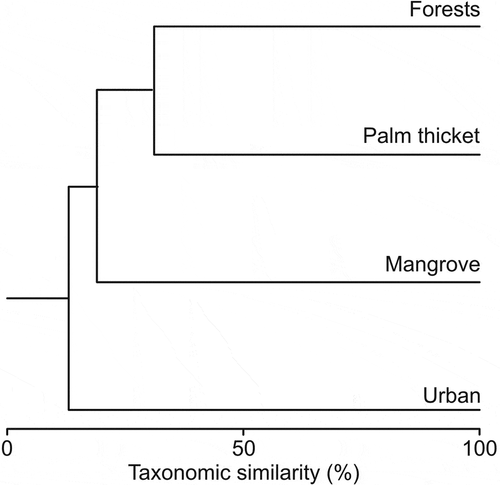
We found sites to be quite different, with high average paired dissimilarities (i.e., ~82%). The most dissimilar ecosystem was the urban setting, with an average 87.3% dissimilarity with the other studied ecosystems, while the most similar ecosystems were tropical forests and palm thickets (68.4% dissimilarity; ). Interestingly, we recorded the highest number of unique species – only recorded in that ecosystem – in mangroves (S = 20, representing 66.6% of the recorded species in that ecosystem), followed by tropical forests (S = 13, representing 43.3% of the recorded species in that ecosystem). The number of unique species in palm thickets and urban settings was low (3 and 5, respectively), representing a low proportion of the recorded species in those ecosystems (16.6% and 23.8%, respectively; ).
Figure 4. Bray–Curtis multivariate analysis showing bird community composition similarity in the four studied ecosystems on Cozumel Island.
Discussion
Studies shedding light on the relative role of ecosystems in vulnerable systems, such as heavily visited islands, can provide information for land managers and planners (Coccossis Citation2002). The studied ecosystems showed overall low species composition similarity; yet, tropical forests and mangroves, where we recorded the highest species richness (86% of the total recorded species), were highly dissimilar in terms of their species composition (84% abundance-based dissimilarity), showing their complementary importance in adding to the avian richness of the island.
Tropical forests were the most diverse of the studied ecosystems in the island in terms of species richness and density. This is in agreement with the plethora of studies documenting the biodiversity sheltered in these ecosystems, estimated to have more than half of the terrestrial biodiversity (Wilson Citation1988; Gardner et al. Citation2009). But the conservation importance of the forests of Cozumel Island does relate not only to the high diversity of birds that they shelter but also to the presence of endemic and quasi-endemic species, some of which are of conservation concern (i.e., Cozumel Vireo – Vireo bairdi; Yucatan Woodpecker – Melanerpes pygmaeus; Caribbean Elaenia – Elaenia martinica). Notably, these species can be vulnerable to natural disasters, such as the critically endangered and endemic Cozumel Thrasher (Toxostoma guttatum), which was considered associated with dense vegetation areas of forest before the impact of Hurricane Gilbert (Macouzet and Escalante Citation2001). Other species, such as the cryptic and highly endangered Great (Cozumel) Curassow (Crax rubra), have been estimated to be more abundant in the island’s tropical forests due to the presence of cenotes and reduced human disturbance (Martínez-Morales Citation1999).
Finding higher avian species richness in the surveyed tropical forests compared to palm thickets was not surprising, considering that higher bird richness can be associated with more complex vegetation structure (Melles et al. Citation2003; Ghadiri Khanaposhtani et al. Citation2012). Tropical forests have more complex vegetation structure than palm thickets, which consist of low diverse communities of secondary vegetation that grew and replaced the original forests that existed before natural or anthropogenic disasters such as hurricanes or wildfires (Téllez-Valdés et al. Citation1989). However, we did record some species that highlight their relative conservation value, including endemic and quasi-endemic species (Cozumel Emerald – Cynanthus forficatus; Caribbean Dove – Leptotila jamaicensis, Black Catbird – Melanoptila glabirostris), as well as species considered of conservation concern, nationally or internationally (i.e., Zenaida Dove – Zenaida aurita, Chimney Swift – Chaetura pelagica, Black Catbird). We recorded 10 times less average avian density in palm thickets compared with tropical forests, suggesting that habitat structure in palm thickets offers less resources for birds (Jayson and Mathew Citation2003). This was not surprising, considering that landscape heterogeneity, vegetation density, and structure are essential environmental characteristics, which often determine bird diversity and distribution (Cody Citation1981).
Besides recording high bird species richness in the studied mangroves, we recorded very low avian densities. Mangroves have been shown to shelter important avian diversity globally (Mohd-Azlan et al. Citation2015; Titoko et al. Citation2019), but the low densities we recorded were not necessarily expected, given that many associated species are social (e.g., White Ibis – Eudocimus albus; Wilson’s Plover – Charadrius wilsonia; Kushlan et al. Citation2017). Additionally, we recorded the highest number of unique species in mangroves. Among the species recorded only in mangroves, two are of conservation concern: the White-crowned Pigeon – Patagienas leucocephala and Reddish Egret – Egretta rufescens.
As expected, the urban area of San Miguel de Cozumel was where we recorded the lowest richness of bird species. Actually, urban settings have been shown to represent a semi-permeable filter for birds globally and at different scales, with their assemblages representing non-random subsets of regional species pools (Croci et al. Citation2008; MacGregor-Fors Citation2010; Aronson et al. Citation2014; Sanz and Caula Citation2015; La Sorte et al. Citation2018). As it has been shown in empirical studies across the urban ecology literature, avian diversity in human settlements depends on vegetation cover and structure (Sultana et al. Citation2021). Similarly, we recorded higher bird richness close to greenspaces of San Miguel de Cozumel. In terms of the urban settings contributing unique species to the island diversity, we only recorded three doves, two exotic (i.e., Rock Pigeon – Columba livia, Eurasian Collared-dove – Streptopelia decaocto) and one native (i.e., White-winged Dove – Zenaida asiatica). It is important to note that the third most abundant species in San Miguel de Cozumel was the Eurasian Collared-Dove, a species that recently colonized Mexican grounds (González-Zamora et al. Citation2016; Ingenloff et al. Citation2017). This dove has increased its populations in San Miguel de Cozumel only since 2008 (with only sporadic records in the island in 1991, 1992, and 2004; GBIF Citation2020). It is currently widespread across town, representing the most frequent species (present in 80% of our point counts located within the urban setting). This is not only worrisome due to antagonistic interactions with other species in the Yucatan Peninsula (Ruddy Ground Dove – Columbina talpacoti, White-winged Dove – Zenaida asiatica; Chablé-Santos et al. Citation2012), but for two additional reasons: (i) it is a potential reservoir of many diseases such as the avian chlamydiosis (Chlamydia psittaci; Donati et al. Citation2015), West Nile Virus (Panella et al. Citation2013), and Newcastle disease (Terregino et al. Citation2003), to name a few; and (ii) because the species has been shown to invade non-urban areas after a lag-phase in the United States, with records spreading through agricultural fields, and even forest edges (eBird Citation2020). It is notable that this species, as an invasive, does not occur in high densities in all islands, as is the case in Japan, where it is considered an endangered species because of decreased population numbers due to reduced genetic diversity (Nagai et al. Citation2020).
The size of the island and the fact that tropical forests cover most of its territory, with important heterogeneity due to both natural and anthropogenic disturbances, could be some of the factors behind the concentration of avian diversity in these ecosystems. This is in agreement with previous studies that native ecosystems in large islands can promote habitat diversity and heterogeneity and consequently biodiversity (MacArthur and Wilson Citation1967; Simberloff Citation1974; Scherber et al. Citation2018). This pattern does not only apply to birds and has been shown to be quite generalizable (e.g., plants, terrestrial mammals, and bats; Power Citation1976; Kohn and Walsh Citation1994; Fox and Fox Citation2000; Davidar et al. Citation2001; Hoffman et al. Citation2019).
Cozumel is a highly biodiverse island, even when contrasted with nearby insular systems. For instance, the avifauna recorded on Cozumel Island – compiled from different sources for a considerable period of time by Lepage (Citation2021) – reaches 333 species, representing ~58% of the total avifauna of the Yucatan Peninsula (566 species; MacKinnon-Haskins Citation2018). In contrast, the avifauna of Isla Mujeres (Quintana Roo), an island one-tenth of Cozumel’s size, is of 123 species (Barrientos-Medina and Ceballos-Povedano Citation2019), comparable to that of Contoy Island, another smaller island, where 97 bird species have been recorded (MacKinnon-Haskins Citation1993). It is noteworthy to mention that such insular avifaunas are heavily affected by seasonal changes, and our surveys were conducted on a specific window of time in August (at the beginning of the arrival of winter migrants); thus, our results should be contextualized in relation to the surveyed time of year. In summary, our results reveal that the surveyed tropical forests and mangroves of the island of Cozumel are the main ecosystems harboring high avian diversity on the island and thus need to be protected. Notably, our results need to be considered cautiously, given our limited sampling effort. Thus, we strongly suggest future studies to pursue long-term surveys to confirm if the relations that we found hold.
On the one hand, tropical forests were shown to house high avian densities and endemics (different from urban settings where we recorded high densities of few species), while on the other hand, we recorded the most unique species in mangroves. Yet, all four studied ecosystems showed overall low species composition similarity, indicating the importance of palm thickets in adding to the avian diversity of the island. Finally, as expected, the contribution of urban settings to the avian diversity of the island is limited and includes some highly abundant invasive species, of which the Eurasian Collared-Dove could represent a major ecological threat.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Alldredge MW, Pollock KH, Simons TR, Shriner SA. 2007. Multiple-species analysis of point count data: a more parsimonious modelling framework: modelling multiple-species count data. J Appl Ecol. 44:281–290. doi:https://doi.org/10.1111/j.1365-2664.2006.01271.x.
- Aronson MFJ, La Sorte FA, Nilon CH, Katti M, Goddard MA, Lepczyk CA, Warren PS, Williams NSG, Cilliers S, Clarkson B, et al. 2014. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. P Roy Soc B. 281:20133330.
- Barrientos-Medina RC, Ceballos-Povedano R. 2019. Aves urbanas, la biodiversidad no visible de Isla Mujeres. Ecofronteras. 23:26–29.
- Berlanga H, Gómez de Silva H, Vargas-Canales VM, Rodríguez-Contreras V, Sánchez-González LA, Ortega-Álvarez R, Calderón-Parra R. 2019. Aves de México: lista actualizada de especies y nombres comunes. Mexico: CONABIO.
- Blackburn TM, Delean S, Pyšek P, Cassey P, Field R. 2016. On the island biogeography of aliens: a global analysis of the richness of plant and bird species on oceanic islands. Global Ecol Biogeogr. 25:859–868. doi:https://doi.org/10.1111/geb.12339.
- Blondel J, Chessel D, Frochot B. 1988. Bird species impoverishment, niche expansion, and density inflation in Mediterranean island habitats. Ecology. 69:1899–1917. doi:https://doi.org/10.2307/1941167.
- Buckland ST, Anderson D, Burnham K. 2001. Introduction to distance sampling: estimating abundance of biological populations. New York: Oxford University Press.
- Buffa J, Morris C. 1999. Occurrence of four Neotropical migrants on Cozumel Island, Mexico. Cotinga. 11:14–16.
- Chablé-Santos J, Gómez-Uc E, Hernández-Betancourt S. 2012. Registros reproductivos de la paloma de collar (Streptopelia decaocto) en Yucatán, México. Huitzil. 13:1–5.
- Chesser RT, Billerman SM, Burns KJ, Cicero C, Dunn JL, Kratter AW, Lovette IJ, Mason NA, Rasmussen PC, Remsen JV Jr, et al. 2020. Sixty-first supplement to the American Ornithological Society’s Check-list of North American birds. Auk. 137:1–24. doi:https://doi.org/10.1093/auk/ukaa030.
- Coccossis H. 2002. Island tourism development and carrying capacity. In: Apostolopoulos Y, Gayle DJ, editors. Island tourism and sustainable development. Westport (CT): Praeger Publishers; p. 131–143.
- Cody ML. 1981. Habitat selection in birds: the roles of vegetation structure, competitors, and productivity. BioScience. 31:107–113. doi:https://doi.org/10.2307/1308252.
- Collantes-Chávez-Costa A, Alanis-Rodríguez E, Yam-Uicab O, López-Contreras C, Sarmiento-Muñoz T, Tapia-Muñoz JL. 2019. Composition, structure and diversity of coastal vegetation in the northeastern of Cozumel, Mexico. Bot Sci. 97:135–147. doi:https://doi.org/10.17129/botsci.2044.
- Colwell R. 2013. EstimateS, Version 9.1: statistical estimation of species richness and shared species from samples. Freeware. http://viceroyeebuconnedu/estimates.
- Croci S, Butet A, Clergeau P. 2008. Does urbanization filter birds on the basis of their biological traits? Condor. 110:223–240. doi:https://doi.org/10.1525/cond.2008.8409.
- Cuarón AD. 2009. Cozumel. In: Gillespie RG, Clague DA, editors. Encyclopedia of islands. Berkeley: University of California Press; p. 203–206.
- Davenport J, Davenport JL. 2006. The impact of tourism and personal leisure transport on coastal environments: a review. Estuar Coast Shelf Sci. 67:280–292. doi:https://doi.org/10.1016/j.ecss.2005.11.026.
- Davidar P, Yoganand K, Ganesh T. 2001. Distribution of forest birds in the Andaman Islands: importance of key habitats: distribution of forest birds in the Andaman Islands. J Biogeogr. 28:663–671. doi:https://doi.org/10.1046/j.1365-2699.2001.00584.x.
- Donati M, Laroucau K, Delogu M, Vorimore F, Aaziz R, Cremonini E, Biondi R, Cotti C, Baldelli R, Di Francesco A. 2015. Chlamydia psittaci in Eurasian collared doves (Streptopelia decaocto) in Italy. J Wildlife Dis. 51:214–217. doi:https://doi.org/10.7589/2014-01-010.
- eBird. 2020. The Cornell Lab of Ornithology. https://ebird.org.
- Flores-Guido JS. 1983. Vegetación insular de la península de Yucatán. Bol Soc Bot Mex. 45:23–37.
- Fox BJ, Fox MD. 2000. Factors determining mammal species richness on habitat islands and isolates: habitat diversity, disturbance, species interactions and guild assembly rules. Global Ecol Biogeogr. 9:19–37. doi:https://doi.org/10.1046/j.1365-2699.2000.00184.x.
- Gardner TA, Barlow J, Chazdon R, Ewers RM, Harvey CA, Peres CA, Sodhi NS. 2009. Prospects for tropical forest biodiversity in a human-modified world. Ecol Lett. 12:561–582. doi:https://doi.org/10.1111/j.1461-0248.2009.01294.x.
- GBIF. 2020. GBIF home page. [accessed 2020 Oct 1]. https://www.gbif.org.
- Ghadiri Khanaposhtani M, Kaboli M, Karami M, Etemad V. 2012. Effect of habitat complexity on richness, abundance and distributional pattern of forest birds. Environ Manage. 50:296–303. doi:https://doi.org/10.1007/s00267-012-9877-7.
- González-Zamora DA, Angulo-Castellanos LF, Hernández-Vázquez S, Almanza-Rodríguez H, Aguilar Nuño LA. 2016. Presencia de la paloma turca (Streptopelia decaocto) en el archipiélago de Revillagigedo, México. Huitzil. 17:151–154.
- Hoffman JD, Kadlubar G, Pedersen SC, Larsen RJ, Larsen PA, Phillips CJ, Kwiecinski GG, Genoways HH. 2019. Predictors of bat species richness within the islands of the Caribbean basin. Lubbock (TX): Museum of Texas Tech University, Special Publications; p. 337–377.
- Howell SNG. 2004. An update on status of birds from Isla Cozumel, Mexico. Cotinga. 22:15–19.
- INEGI. 2017. Anuario estadístico y geográfico de Quintana Roo 2017. Quintana Roo (México): Instituto Nacional de Estadística y Geografía (INEGI).
- Ingenloff K, Hensz CM, Anamza T, Barve V, Campbell LP, Cooper JC, Komp E, Jimenez L, Olson KV, Osorio-Olvera L, et al. 2017. Predictable invasion dynamics in North American populations of the Eurasian collared dove Streptopelia decaocto. P Roy Soc B. 284:20171157.
- Irizarry JI, Collazo JA, Dinsmore SJ. 2016. Occupancy dynamics in human-modified landscapes in a tropical island: implications for conservation design. Divers Distrib. 22:410–421. doi:https://doi.org/10.1111/ddi.12415.
- IUCN. 2019. The IUCN red list of threatened species. [accessed 2020 Oct 1]. http://www.iucnredlist.org/.
- Jayson EA, Mathew DN. 2003. Vertical stratification and its relation to foliage in tropical forest birds in western Ghats (India). Acta Ornithol. 38:111–116. doi:https://doi.org/10.3161/068.038.0207.
- Khimoun A, Peterman W, Eraud C, Faivre B, Navarro N, Garnier S. 2017. Landscape genetic analyses reveal fine-scale effects of forest fragmentation in an insular tropical bird. Mol Ecol. 26:4906–4919. doi:https://doi.org/10.1111/mec.14233.
- Kohn DD, Walsh DM. 1994. Plant species richness – the effect of island size and habitat diversity. J Ecol. 82:367–377. doi:https://doi.org/10.2307/2261304.
- Kushlan JA, Angehr GR, Hines K. 2017. Seabirds and other colonial waterbirds of the Caribbean coast of Panama. J Caribbean Ornithol. 30:145–153.
- La Sorte FA, Lepczyk CA, Aronson MFJ, Goddard MA, Hedblom M, Katti M, MacGregor-Fors I, Mörtberg U, Nilon CH, Warren PS, et al. 2018. The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization. Divers Distrib. 24:928–938. doi:https://doi.org/10.1111/ddi.12738.
- Lepage D. 2021. Avibase – bird checklists of the world: Cozumel island. [accessed 2021 Mar 23]. https://avibase.bsc-eoc.org/checklist.jsp?region=MXqncz.
- Lugo AE, Carlo TA, Wunderle JM. 2012. Natural mixing of species: novel plant–animal communities on Caribbean Islands. Anim Conserv. 15:233–241. doi:https://doi.org/10.1111/j.1469-1795.2012.00523.x.
- MacArthur RH, Wilson EO. 1967. The theory of island biogeography. 1st ed. Princeton (USA): Princeton University Press.
- MacGregor-Fors I. 2010. How to measure the urban-wildland ecotone: redefining ‘peri-urban’ areas. Ecol Res. 25:883–887. doi:https://doi.org/10.1007/s11284-010-0717-z.
- MacGregor-Fors I, Payton ME. 2013. Contrasting diversity values: statistical inferences based on overlapping confidence intervals. PLoS ONE. 8:e56794. doi:https://doi.org/10.1371/journal.pone.0056794.
- MacKinnon-Haskins B. 1993. Listado de las aves en Isla Contoy. Quintana Roo (México): Amigos de Sian Ka’an.
- MacKinnon-Haskins B. 2018. Check-list of the birds of the Yucatan Peninsula. Quintana Roo (México): Amigos de Sian Ka’an.
- Macouzet TF, Escalante PP. 2001. Registros del Cuitlacoche de Cozumel Toxostoma guttatum posteriores al huracán Gilberto. Cotinga. 15:32–33.
- Martínez-Morales MA. 1999. Conservation status and habitat preferences of the Cozumel Curassow. Condor. 101:14–20.
- Martínez-Morales MA, Cruz PC, Cuarón AD. 2009. Predicted population trends for Cozumel Curassows (Crax rubra griscomi): empirical evidence and predictive models in the face of climate change. J Field Ornithol. 80:317–327. doi:https://doi.org/10.1111/j.1557-9263.2009.00237.x.
- Martínez-Morales MA, Cuarón AD. 1999. Boa constrictor, an introduced predator threatening the endemic fauna on Cozumel Island, Mexico. Biodiv Conserv. 8(7):957–963. doi:https://doi.org/10.1023/A:1008815004072.
- Melles S, Glenn SM, Martin K. 2003. Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conserv Ecol. 7:5.
- Mohd-Azlan J, Noske R, Lawes M. 2015. The role of habitat heterogeneity in structuring mangrove bird assemblages. Diversity. 7:118–136. doi:https://doi.org/10.3390/d7020118.
- Morales-Contreras J, Escalante P, Matías-Ferrer N. 2019. Genetic diversity in Mexican wild populations of the Great Curassow (Crax rubra). Biota Neotrop. 19:e20180649. doi:https://doi.org/10.1590/1676-0611-bn-2018-0649.
- Myers N, Mittermeier RA, Mittermeier CG, Da Fonseca GA, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature. 403:853–858. doi:https://doi.org/10.1038/35002501.
- Nagai K, Takahashi Y, Tokita K, Anzai T, Uchida K, Nakayama F. 2020. Genetic diversity in Japanese populations of the Eurasian Collared Dove. Avian Res. 11:1–6. doi:https://doi.org/10.1186/s40657-020-00207-8.
- Nichols JD, Hines JE, Sauer JR, Fallon FW, Fallon JE, Heglund PJ. 2000. A double-observer approach for estimating detection probability and abundance from point counts. Auk. 117:393–408. doi:https://doi.org/10.1093/auk/117.2.393.
- Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara R, Simpson GL, Solymos P. 2016. vegan: Community ecology package. R package version 2.4-1.
- Panella NA, Young G, Komar N. 2013. Experimental infection of Eurasian collared-dove (Streptopelia decaocto) with West Nile virus. J Vector Ecol. 38:210–214. doi:https://doi.org/10.1111/j.1948-7134.2013.12032.x.
- Perdomo-Velázquez H, Andresen E, Vega E, Schondube JE, Cuarón AD. 2017. Effects of hurricanes on the understory forest birds of Cozumel Island. Trop Conserv Sci. 10:1–14.
- Pinto FAS, Clevenger AP, Grilo C. 2020. Effects of roads on terrestrial vertebrate species in Latin America. Environ Impact Asses. 81:106337. doi:https://doi.org/10.1016/j.eiar.2019.106337.
- Power DM. 1976. Avifauna richness on the California Channel Islands. Condor. 78:394–398. doi:https://doi.org/10.2307/1367701.
- R Development Core Team. 2019. R: a language and environment for statistical computing. Vienna (Austria): R Foundation for Statistical Computing. http://www.R-project.org/.
- Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF, Milá B. 1996. Manual de métodos de campo para el monitoreo de aves terrestres. Albany (CA): Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture. Gen. Tech. Rep. PSW-GTR-159.
- Rosenzweig ML. 1975. On continental steady states of species diversity. In: Cody ML, MacArthur RH, Diamond JM, editors. Ecology and evolution of communities. Cambridge: Harvard University Press; p. 121–141.
- Sanz V, Caula S. 2015. Assessing bird assemblages along an urban gradient in a Caribbean island (Margarita, Venezuela). Urban Ecosyst. 18:729–746. doi:https://doi.org/10.1007/s11252-014-0426-4.
- Scherber C, Andert H, Niedringhaus R, Tscharntke T. 2018. A barrier island perspective on species–area relationships. Ecol Evol. 8:1–11. doi:https://doi.org/10.1002/ece3.4726.
- SEMARNAT. 2010. Norma Oficial Mexicana NOM-059-SEMARNAT. [accessed 2021 Mar 23]. https://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm.
- SEMARNAT. 2019. Norma Oficial Mexicana NOM-059-SEMARNAT (Update). [accessed 2021 Mar 23]. http://www.dof.gob.mx/nota_detalle.php?codigo=5576689&fecha=28/10/2019.
- Simberloff DS. 1974. Equilibrium theory of island biogeography and ecology. Annu Rev Ecol Syst. 5:161–182. doi:https://doi.org/10.1146/annurev.es.05.110174.001113.
- Sosa-López JR, Mennill DJ. 2014. Vocal behaviour of the island-endemic Cozumel Wren (Troglodytes aedon beani): song structure, repertoires, and song sharing. J Ornithol. 155:337–346. doi:https://doi.org/10.1007/s10336-013-1008-6.
- Stark SC, Bunker DE, Carson WP. 2006. A null model of exotic plant diversity tested with exotic and native species-area relationships: exotic and native species-area relationships. Ecol Lett. 9:136–141. doi:https://doi.org/10.1111/j.1461-0248.2005.00859.x.
- Sultana M, Corlatti L, Storch I. 2021. The interaction of imperviousness and habitat heterogeneity drives bird richness patterns in south Asian cities. Urban Ecosyst. 24(2):335–344. doi:https://doi.org/10.1007/s11252-020-01037-8.
- Téllez-Valdés O, Cabrera EF. 1987. Florula de la Isla Cozumel, Quintana Roo. Listados florísticos de México. Mexico: Instituto de Biología, Universidad Nacional Autónoma de México.
- Téllez-Valdés O, Cabrera EF, Linares E, Bye R. 1989. Las plantas de Cozumel. Guía botánico-turística de la Isla de Cozumel. Quintana Roo (Mexico): Instituto de Biología, Universidad Nacional Autónoma de México.
- Terregino C, Cattoli G, Grossele B, Bertoli E, Tisato E, Capua I. 2003. Characterization of Newcastle disease virus isolates obtained from Eurasian collared doves (Streptopelia decaocto) in Italy. Avian Pathol. 32:63–68. doi:https://doi.org/10.1080/0307945021000070732.
- Thomas L, Buckland ST, Rexstad EA, Laake JL, Strindberg S, Hedley SL, Bishop JRB, Marques TA, Burnham KP. 2010. Distance software: design and analysis of distance sampling surveys for estimating population size. J Appl Ecol. 47:5–14. doi:https://doi.org/10.1111/j.1365-2664.2009.01737.x.
- Titoko R, Lowry JH, Osborne T, Naikatini A, Comely J, Riley R. 2019. Relationship of bird richness, abundance and assemblage to the built environment in a small island tropical urban setting: a Suva, Fiji case study. Urban Ecosyst. 22:709–719. doi:https://doi.org/10.1007/s11252-019-00861-x.
- UNESCO. 2016. Isla Cozumel. Ecological sciences for sustainable development. [accessed 2020 Oct 1]. http://www.unesco.org/new/en/natural-sciences/environment/ecological-sciences/biosphere-reserves/latin-america-and-the-caribbean/mexico/isla-cozumel/.
- Vázquez-Domínguez E, Espindola S. 2013. Characterization of ten new microsatellite loci from the endangered endemic rodent, Reithrodontomys spectabilis. Conserv Genet Resour. 5:251–253. doi:https://doi.org/10.1007/s12686-012-9780-9.
- Wang E, Van Wijk RE, Braun MS, Wink M. 2017. Gene flow and genetic drift contribute to high genetic diversity with low phylogeographical structure in European hoopoes (Upupa epops). Mol Phylogenet Evol. 113:113–125. doi:https://doi.org/10.1016/j.ympev.2017.05.018.
- Webster PJ, Holland GJ, Curry JA, Chang H-R. 2005. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science. 309:1844–1846. doi:https://doi.org/10.1126/science.1116448.
- Weidie AE. 1985. Geology of the Yucatan Platform, part 1. In: Ward WC, Weidie AE, Back W, editors. Geology and hydrogeology of the Yucatan and quaternary geology of Northeastern Yucatan Peninsula. New Orleans (Louisiana): New Orleans Geological Society; p. 1–9.
- Whittaker RJ, Fernández-Palacios JM. 2007. Island biogeography: ecology, evolution, and conservation. Oxford: Oxford University Press.
- Wiley JW, Wunderle JM. 1993. The effects of hurricanes on birds, with special reference to Caribbean islands. Bird Conserv Int. 3:319–349. doi:https://doi.org/10.1017/S0959270900002598.
- Wilson EO. 1988. The current state of biological diversity. Biodiversity. 521:3–18.
- Yasué M, Dearden P. 2006. The potential impact of tourism development on habitat availability and productivity of Malaysian plovers Charadrius peronii. J Appl Ecol. 43:978–989.