
Abstract
Lichen genus Stereocaulon (Schreb.) Hoffm is distributed throughout the world. Although 15 Stereocaulon species have been recorded in Korea, no detailed taxonomic or revisionary research has been conducted for nearly two decades. In this study, we collected 260 putative Stereocaulon spp. samples and identified the species based on morphological, chemical, and molecular characteristics. From the collected samples, 10 species of Stereocaulon were identified, nine of which had already been reported, although this was the first report for the tenth, S. octomerellum Hue, in Korea. General characteristics of Stereocaulon spp. include coralloid phyllocladia and tubercular cephalodia; however, the specimen first collected in Korea was a rare species with tomentum on the pseudopodetia. The specimen of S. octomerellum is characterized by the presence of a primary thallus, granule to short coralloid phyllocladia, and pseudopodetia up to 1 cm in size, with tubercular cephalodia. To determine the phylogeny of the specimens, we compared the ITS sequences of ribosomal DNA and the β-tubulin gene sequences. Phylogenetic analysis showed that the Korean Stereocaulon species were monophyletic and placed in the previous phylogenetic classification. Species of S. intermedium and S. exutum, however, were polyphyletic, and are morphologically variable and widespread species. Overall, we present here detailed morphological and chemical descriptions of each species identified and a revised key of all known Stereocaulon species in South Korea.
1. Introduction
The lichen genus Stereocaulon Hoffm. is a cosmopolitan genus consisting of approximately 125 species of common fruticose lichens [Citation1,Citation2]. Stereocaulon spp. are generally composed of a primary thallus, pseudopodetia, phyllocladia, cephalodia, and apothecia, although only some species have soredia and tomentum. Primary thalli of Stereocaulon spp. are short lived or persistent, consisting of tiny white granules or squamules, and pseudopodetia are erect or decumbent. Cephalodia are chlorococcoid (Trebouxia) or blue-green (Nostoc or Stigonema), and are attached to the primary thallus or pseudopodia, which can be spherical to botryose and sacculate [Citation2,Citation3]. Phyllocladia are classified into coralloid, granular, squamulose, and peltate types [Citation3]. During growth of these lichens, cephalodia appear at an early stage and persist on the basal part of the pseudopodetia, and pseudopodetia arise from the primary thallus by elongation of the thalline tissue as areolae or verrucae [Citation3,Citation4].
A worldwide key, a conspectus, and a classification of the genus Stereocaulon based on morphology, chemistry, and ontogeny were produced by Lamb [Citation3,Citation5,Citation6]. Alongside morphological classification, molecular analysis of the genus Stereocaulon has been studied using the internal transcribed spacer (ITS) regions of ribosomal DNA and partial β-tubulin gene regions [Citation4]. Using these two molecular markers, Högnabba analyzed the phylogenetic status of 49 out of 125 known taxa [Citation4]. This study revealed that Stereocaulon was polyphyletic and the morphological classification of Stereocaulon, suggesting morphological analysis may not be exactly in accord with molecular analysis [Citation4].
To date, 15 Stereocaulon species have been reported in South Korea [Citation7]. Before 1950, Japanese lichenologists, Sato and Ueda, reported three Stereocaulon species, S. coralloides, S. paschale, and S. wrightii [Citation8,Citation9]. Later, Kim reported five different Stereocaulon species, including S. apocalypticum, S. curtatum, S. japonicum, S. paschale, and S. wrightii [Citation10]. Park described nine more species in South Korea, including S. commixtum, S. dendroides, S. exutum, S. intermedium, S. nigrum, S. octomerum, S. sorediiferum, S. verruculigerum, and S. vesuvianum [Citation11]. In 2002, Kasiwadani additionally reported the presence of S. pileatum in Korea [Citation12]. Although a number of studies have been conducted on the genus Stereocaulon in Korea, none has conducted molecular analysis.
In the present study, the taxonomy of Stereocaulon species in Korea is revised based on morphological and chemical characteristics and molecular phylogenetic analysis. A key to all known Stereocaulon species in Korean is also presented.
2. Materials and methods
2.1. Morphological examination
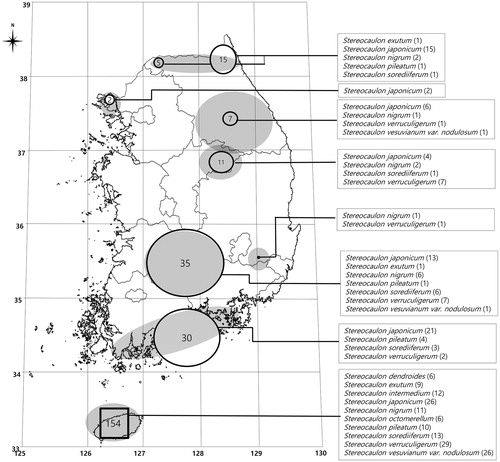
Samples of the genus Stereocaulon were collected in South Korea from 2003 to 2016 (). The specimens were deposited at the herbarium of KoLRI (Korea Lichen Research Institute). Descriptions of the external morphology were based on air-dried materials observed under a Nikon SMZ-645 dissecting microscope (Nikon Corp., Tokyo, Japan) and an Olympus BX-50 compound microscope (Olympus, Tokyo, Japan) was used to study the anatomy of ascomata. Color reaction tests were conducted on phyllocladia and pseudopodetia to check for secondary metabolites. (K = 10% aqueous KOH solution, P = 5% alcoholic p-phenylenediamine solution) [Citation13]. Thin-layer chromatography (TLC) was performed in Solvent A (toluene:dioxin:acetic acid =180:45:5) and solvent C (toluene:acetic acid =85:15) [Citation14].
Figure 1. Collection map of Stereocaulon specimens during 2003–2016 in South Korea.
2.2. DNA extraction and PCR amplification
Total DNA was extracted directly from the lichen thalli using the DNeasy Plant Mini Kit (QIAGEN, Dusseldorf, Germany). The partial 18 s rRNA–ITS1–5.8s rRNA–ITS2–partial 28 s rRNA region was amplified with universal primers ITS1F [Citation15], ITS4 [Citation16], or LR5 [Citation17]. PCR amplification was carried out using a Takara JP/TP600 PCR machine (Takara Bio Inc., Kusatsu, Japan) under the following conditions: initial cycle of 5 min at 94 °C, followed by 30 cycles of 30 seconds at 94 °C, 30 seconds at 55 °C, 10 min at 72 °C, and then finally 10 min at 72 °C. The partial β-tubulin sequence was amplified with Bt3-LM and Bt10-LM [Citation18] or Bt-PJS-F1(5′-GCCTCTGGCAAATATGT-3′) and Bt-PJS-R1(5′-AATTCTGGCACGGTGACC-3′).
β-tubulin samples were prepared in the same way as ITS samples. The PCR reaction of β-tubulin was carried out in a 2720 thermal cycler PCR machine (Applied Biosystems, Singapore, Singapore) under the following conditions: initial cycle of 5 min at 94 °C, followed by 30 cycles of 30 seconds at 94 °C, 30 seconds at 55 °C initially, and then 52 °C for the remaining cycles, 1 min at 72 °C, and finally 72 °C for 5 min [Citation4].
PCR products were purified using a PCR quick-spin PCR Product Purification Kit (INTRON Biotechnology, Inc., seoul, South Korea) and then sent to Genotech Cooperation (Daejeon, Korea) for sequencing.
2.3. Sequence alignments and phylogenetic analysis
The sequences were manually edited and assembled using CodonCode Aligner (CodonCode Corporation, Dedham, MA) and BIOEDIT 7.0.9 [Citation19]. All the sequences generated in this study and sequences retrieved from GenBank were initially aligned using Clustal W ver. 1.83 [Citation20]. Two lichen species, Cladonia ahtii (AF45327) and C. rangiformis (AF455172) were used as outgroups. Phylogenetic analysis was conducted using the maximum parsimony (MP) method and neighbor joining (NJ) in MEGA version 6.0 software [Citation21]. Reliability of the inferred tree was tested by 1000 bootstrap replications of the MP and NJ tree. Kimura two-parameter model was used for the analysis of the NJ method.
3. Results and discussion
3.1. Taxonomy and distribution of the genus Stereocaulon in Korea
The genus Stereocaulon was collected all over South Korea along Baekdudaegan from the truce line to Jeju Island (). Taxonomic identification was carried out as per Lamb’s taxonomic keys [Citation6]. The taxonomic characteristics of the genus Stereocaulon in South Korea are described at the end of this report.
A total of 260 Stereocaulon specimens were collected from eight different sampling sites (; marked in gray) and 59.2% of them were collected on Jeju Island. Using morphological and chemical analysis, we identified 10 Stereocaulon species: S. dendroides, S. exutum, S. japonicum, S. intermedium, S. nigrum, S. octomerellum, S. pileatum, S. sorediiferum, S. verruculigerum, and S. vesuvianum var. nodulosum. Nine of these 10 species have already been reported in Korea in previous studies [Citation10]; however, this is the first report for S. octomerellum.
The distribution of the 10 species identified is shown in . S. japonicum (34% of total 260 specimens collected) was the most common Stereocaulon species, distributed throughout South Korea at all sampling sites (). The second most common was S. verruculigerum, with 47 specimens (18%) in South Korea (). Twenty-eight specimens of S. vesuvianum var. nodulosum (11%) (), 24 of S. sorediiferum (9%) (), 23 of S. nigrum (9%) (), and 16 of S. pileatum (6%) () were distributed in various regions. S. intermedium (5%) (), S. exutum (4%) (), and S. dendroides (2%) () were mainly observed in Jeju Island.
Figure 2. Distribution of Stereocaulon species in South Korea. Circle indicates Stereocaulon species that was also found in a previous study. Stars indicates samples collected in this study. (A) S. japonicum; (B) S. verruculigerum; (C) S. vesuvianum var. nodulosum; (D) S. sorediiferum; (E) S. nigrum; (F) S. pileatum; (G) S. intermedium; (H) S. exutum; (I) S. dendroides; (J) S. octomerellum.
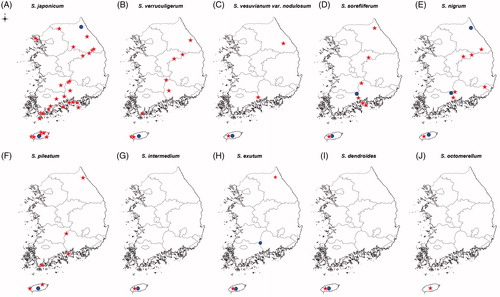
Figure 3. Habit of the Stereocaulon species. (A) S. japonicum Hur130006; (B) S. verruculigerum Hur141605; (C) S. vesuvianum var. nodulosum Hur130060; (D) S. sorediiferum Hur040639; (E) S. nigrum Hur130045; (F) S. pileatum Hur121814; (G) S. intermedium Hur130044; (H) S. exutum Hur130030; (I) S. dendroides Hur130059; (J) S. octomerellum Hur121094; (K) Hypothecium color of S. octomerellum; (L) Spore type and septation of S. octomerellum (Scale bars: K = 100 µm, L = 10 µm).

Six specimens of S. octomerellum (2%), which had not yet been reported in South Korea, were collected from Mt. Halla in Jeju Island (). This species is characterized by pseudopodetia less than 1 cm in size, a short coralloid or granule form of phyllocladia, and colorless hypothecium (). The S. octomerellum specimens collected were very similar to S. verruculigerum and were found to contain lobaric acid. The main substances of Korean Stereocaulon are stictic acid and lobaric acid, and nine of the 10 species identified in this study were classified as producers of these two substances, whereas S. dendroides produces porphyrilic acid.
Previously, Stereocaulon was collected mainly from Mt. Halla, Mt. Jiri, and Mt. Sorak [Citation11,Citation12]. In addition, more studies on Stereocaulon have been conducted in Japan and China than in South Korea. In Japan, Stereocaulon was classified by Asahina and 33 species and 3 subspecies were reported [Citation22–25]. In China, Stereocaulon was classified by Huang and 44 species were reported [Citation26,Citation27]. Also in Japan and China, Stereocaulon species have been classified on the basis of morphology and secondary metabolite chemistry, but not using molecular biology approaches.
3.2. Phylogenetic analysis
To perform molecular phylogenetic analysis, we randomly selected two representative specimens for each of the 10 Stereocaulon species identified. Two general molecular sequences for the genus Stereocaulon, ITS1–5.8s rRNA–ITS2 regions, and protein-coding gene β-tubulin, were used for the analysis [Citation28]. Length variation of ITS1 and ITS2 was 225 to 238 bp and 93 to154 bp, respectively. All sequences from 5.8 s rRNA were of identical length. For β-tubulin, all intron lengths were the same and the lengths of exon 5 and exon 6 were also the same for all species, except exon 5 of S. exutum and S. sorediiferum, which were shorter than that of the other species. All generated sequences were deposited in NCBI and the accession numbers are listed in and Supplementary data S1.
Table 1. Specimens used in molecular phylogenetic analysis.
For phylogenetic analysis, we retrieved sequences of both the ITS regions and the β-tubulin gene from 58 Stereocaulon species obtained in a previous study [Citation4]. We merged the sequences of both ITS regions and the β-tubulin gene that showed 1338 characters containing 746 conserved characters and 583 variable characters. The MP analyses were applied to generate a phylogenetic tree as previously described [Citation4]. One of the most parsimonious trees is shown in . We also constructed an NJ tree in Supplementary data S2 for comparison with the MP tree. Both phylogenetic trees had the same structure, and the interspecific relationship was also the same.
Figure 4. Maximum parsimony (MP) tree was constructed by MEGA6. The phylogenetic tree generated from internal transcribed spacer (ITS) sequences and part of the β-tubulin gene sequence. The bold letters in black indicate 20 representatives from 10 Stereocaulon species used in this study. Bootstrap = 1000. The numbers at each node represent bootstrap values. Only nodes supported by 50 bootstraps or more are shown.
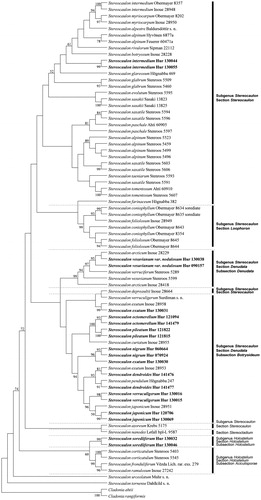
Stereocaulon spp. in South Korea analyzed in this study was found to be polyphyletic with a low support value, as in previous studies [Citation4]. Unlike the previous analyses, however, S. sorediiferum did not exist as a monophyletic clade at the base of the tree but was also grouped with subsection Aciculisporae. This result was probably resulted from the use of only β-tubulin sequences in previous studies [Citation4].
Stereocaulon japonicum was grouped in the same clade with the reference S. japonicum sequence that we used in this study (). However, in a previous study, the reference S. japonicum samples were separated into two different clades, suggesting that it is a polymorphic species. We observed that S. verruculigerum was in the same clade as S. japonicum with a well-supported bootstrap value. However, in a previous study, S. verruculigerum was placed in a different clade from S. japonicum, although it has been reported that S. verruculigerum is highly similar to S. japonicum in their morphology and both species are generally classified by the presence or the absence of stictic acid and lobaric acid [Citation6].
Stereocaulon dendroides was newly sequenced in this study. From previous studies, it was known that S. dendroides and S. pendulum contain the same chemicals [Citation6]. Interestingly, we observed that our S. dendroides species clustered with S. pendulum.
Our two Korean S. exutum samples did not belong to the same clade; one (Hur 130031) of the two specimens was in the same clade as the reference S. exutum (Inoue 28958) but the other was not. Two S. nigrum samples were grouped in the same clade with S. exutum, as it have a highly similar morphology to S. exutum ().
The β-tubulin sequence from S. pileatum and the ITS sequence from S. octomerellum were newly sequenced and analyzed in this study. The phylogenetic analysis showed that these two species were in the same clade with a well-supported bootstrap value. It is known that these two species have pseudopodetia less than 1 cm in size. In previous studies, S. pileatum and S. verruculigerum were grouped in the same clade using only the ITS nucleotide sequence. As a result of using two markers, S. pileatum was grouped in the clades with other lichens that were morphologically similar, such as S. octomerellum.
As previously reported, S. vesuvianum var. nodulosum was grouped together with S. arcticum. The two species are very similar in morphology and the only difference is the presence or the absence of Stigonema and Nostoc in the cephalodia [Citation29].
We observed that our S. intermedium specimens were separated from the reference S. intermedium in the phylogenetic tree (). Although previous studies have referred to this species as a single lineage, our results indicate that our S. intermedium is in a different clade, with a well-supported bootstrap value, which suggests that this “species” may be polyphyletic. It could be possible that the genetic sequence variation of the species may result from regional or morphological variation [Citation11].
In conclusion, molecular analysis carried out in this study indicated that the molecular phylogeny of genus Stereocaulon in South Korea is largely in agreement with the previous molecular classification, except for S. intermedium. S. intermedium was not in accordance with previous findings [Citation4]. This might have resulted from differences in the regional distribution of the specimens used in the previous study.
In this survey, we extensively collected specimens of the genus Stereocaulon in South Korea and identified them by morphological, chemical, and molecular analyses. This study will contribute and supplement further studies on lichen classification and ecology.
An updated taxonomic key to the lichen genus Stereocaulon in South Korea.
Soredia present2
- Soredia absent3
Pseudopodetia up to 1 cmS. pileatum
- Pseudopodetia over 1 cmS. sorediiferum
Phyllocladia spatulas apical foliar expansion at the ends of the pseudopodetia
S. apocalypticum
- Not above4
PD + red (stictic acid present)5
- PD– pale yellow (lobaric, porphyrilic acid present)8
Phyllocladia coralloid6
- Phyllocladia peltate, foliose7
Miriquidic acid presentS. commixtum
- Miriquidic acid absentS. japonicum
Phyllocladia peltate
S. vesuvianum var. nodulosum
- Phyllocladia folioseS. wrightii
Primary thallus persist9
- Primary thallus lacking11
Spore broad, 6–7(–9) μm, miriquidic acid presentS. curtatum
- Spore narrow, not over 5 μm, miriquidic acid absent, lobaric acid present10
Pseudopodetia up to 1 cmS. octomerellum
- Pseudopodetia over 1 cm, lobaric acid present, morphologically similar to S. japonicum
S. verruculigerum
Tomentum present12
- Tomentum absent13
Phyllocladia granular, cephalodia scabrid, blackishS. paschale
- Phyllocladia granular to terete coralloid, cephalodia spherical, grayS. intermedium
Lobaric acid present14
- Porphyrilic acid presentS. dendroides
Spore up to seven septate, phyllocladia not over 2 mm longS. octomerum
- Spore usually not over five septate, phyllocladia over 3 mm long15
Hypothecium colorlessS. exutum
- Hypothecium brownS. nigrum
4. Description of the species
4.1. Stereocaulon dendroides Asahina, J. Jap. Bot. 36: 231 (1961)
Morphology: Primary thallus lacking; pseudopodetia dorsiventral, tuft-like, well-branched from the base, thinner toward the top, 3–4.5(–5) cm tall, rust red near the base, ligneous, slightly striate, glabrous, firmly attached to the substrate and attached by one holdfast, phyllocladia coralloid, cylindrical, thin, simple to branched, up to 1–1.5 mm long, densely distributed on the pseudopodetia, cephalodia conspicuous, gray, spherical, tubercular, 0.5–0.7 mm wide, present in the lower part of pseudopodetia. Apothecia terminal, convex, botryose-like, reddish brown, 0.8–1 mm wide, rare, epihymenium pale brown, hymenium hyaline, 30–37.5 μm high, hypothecium brown, 100–150 μm high; spore fusiform, hyaline, 3–5 septate, 20–25 × 2.5–5 μm.
Chemistry: K + yellow, P + pale yellow. TLC: atranorin, porphyrilic acid.
Remark: S. dendroides has unique secondary metabolites compared to other species. This species is close to S. verruculigerum but it has lobaric acid instead of porphyrilic acid.
Representative specimen examined: Jeju-do, Jeju-si, Mt. Halla, alt. 1,658 m, on rock, 33°21′49.09″N, 126°30′42.3″E, 04 July 2012, S. Y. Kondratyuk, L. Lokös, S.-O. Oh, S. Joshi, J.-S. Hur, Hur 121345 (KoLRI 016388).
4.2. Stereocaulon exutum Nyl., Lichenes Japoniae: 18 (1890)
Morphology: Primary thallus lacking; pseudopodetia erect, dorsiventral, tapered, well-branched, 2.5–3.5 cm tall, mostly dichotomous, main branches rather thick, glabrous, exposing gelatinous layer, densely phyllocladiate, phyllocladia numerous, cylindrical-coralloid, thin, simple to delicately branched, dichotomously ramulose, up to 2–3 mm long, cephalodia gray, spherical to botryose, tubercular, 0.5–0.8 mm wide, mostly present in the lower part of pseudopodetia. Apothecia not common, terminal, convex, reddish brown to brown, 1–1.5 mm wide, epihymenium brown, 17–25 μm high, hymenium hyaline, 37.5–50 μm high, hypothecium hyaline, 45–50 μm high; spores not observed.
Chemistry: K + yellow, P + pale yellow. TLC: atranorin, lobaric acid.
Remarks: S. exutum is similar to S. nigrum, but has colorless hypothecium in the apothecia. This species is characterized by containing lobaric acid and having longer and thinner phyllocladia, which are well-developed than in other species.
Representative specimen examined: Gangwon-do, Inje-gun, Mt. Sorak, alt. 660 m, on rock, 38°09′37.07″N, 128°19′26.05″E, 15 May 2005, J.-S. Hur, Hur 050254 (KoLRI 003158).
4.3. Stereocaulon japonicum Th. Fr., De Stereocaulis et Pilophoris commentatio: Uppsala: 18 (1857)
Morphology: Primary thallus persistent or soon evanescent; pseudopodetia erect, dorsiventral, 1–2 cm tall, upper part dichotomously branched, main stems 1 mm wide, glabrous, completely decorticated, firmly attached to the substrate, phyllocladia abundant, granular-like to cylindrical-coralloid, simple to branched, gray to dark green, up to 2 mm long, cephalodia common, spherical, tubercular, gray to brown, 0.5–1 mm wide, mostly present on the basal part. Apothecia mostly terminal or rarely lateral on branch, reddish brown to black, convex, 1–2 mm wide, immature apothecia have exiple, which disappear when mature, epihymenium brown, 20–30 μm high, hymenium hyaline, 50–100 μm high, hypothecium brown, 90–100 μm high; spore fusiform, hyaline, 3–5(–7) septate, 30 × 2.5–5 μm.
Chemistry: K + yellow, P + red. TLC: atranorin, norstictic, stictic, cryptostictic, menegazziaic, and constictic acids.
Remarks: S. japonicum is similar to S. commixtum but the latter has miriquidic acid and pseudopodetia are taller than that of S. japonicum. This species is also morphologically close to S. verruculigerum but differs in having stictic acid instead of lobaric acid. S. japonicum has much variation and is widely distributed in South Korea.
Representative specimens examined: Gyeongsangnam-do, Peak Ungseokbong, alt. 848 m, on rock, 35°22′58.08″N, 127°51′29.01″E, 16 October 2007, J.-S. Hur, Hur 070900 (KoLRI 007710); Gangwon-do, Mt. Hwangbyeong, alt. 772 m, on rock, 37°44′47.06″N, 128°37′31.05″E, 14 Jul 2008, Hur 080414 (KoLRI 008661); Jeollanam-do, Yeosu-si, Bulga coast, alt. 12 m, on rock, 34°39′00.04″N, 127°34′04.07″E, 28 April 2012, U. Jayalal, J. S. Park, J.-A. Rye, Hur 120706 (KoLRI 017060).
4.4. Stereocaulon intermedium (Sav.) Magn., Bot. Mater. Inst. Sporov. Rast. Glavn. Bot. Sada RSFSR 2(11 163. 1923): 163 (1923)
Morphology: Primary thallus lacking; pseudopodetia erect, dorsiventral, caespitose, well branched from the base to the top, becoming gradually thinner toward the top. 2.5–3.7 cm tall, 0.5–0.7 mm wide, decorticate, exposing inner layer, not ligneous, glabrous to thin tomentum, firmly attached to the rock, phyllocladia abundant, granular to verrucose in upper part, whereas in basal part terete to coralloid, partly squamulose-like, grayish white to white, 0.5–1 mm long, 0.1–0.2 mm wide, cephalodia abundant, sessile on pseudopodetia, violet-gray to brownish, spherical to botryose, tubercular, 1–2 mm wide, concealed in phyllocladia to conspicuous well on pseudopodetia. Apothecia are not observed.
Chemistry: K + yellow, P– pale yellow. TLC: atranorin, lobaric acid.
Remarks: This species may be confused with other coralloid type species. According to Park, it is close to S. verruculigerum but differs by having a short tomentum and shorter pseudopodetia and is more whitish with verrucose phyllocladia at the young stage [Citation11].
Representative specimen examined: Jeju-do, Jeju-si, Mt. Halla, alt. 1585 m, on rock, 33°22′04.03″N, 126°30′35.09″E, 04 April 2013, S.-O. Oh, J. S. Park, 130055 (KoLRI 018041); the same locality, alt. 1671 m, on rock, 33°21′35.01″N, 126°30′42.30″E, 02 May 2014, S.-O. Oh & J. S. Park, Hur 140098 (KoLRI 021271).
4.5. Stereocaulon nigrum Hue, Nouv. Arch. Mus. Hist. Nat. ser. 3, 10: 248 (sep. 36) (1898)
Morphology: Primary thallus lacking; pseudopodetia 2–4 cm tall, dorsiventral, tuft, well-branched, densely phyllocladiate, decorticate, exposing inner brownish layer, phyllocladia lateral, coralloid, cylindrical, simple to delicately branched, up to 2–3 mm long, cephalodia abundant, spherical to botryose, tubercular, dark gray, 1 mm wide. Apothecia terminal, convex, dark brown to black, epihymenium and hypothecium brown, hymenium hyaline; spores not observed.
Chemistry: K + yellow, P + pale yellow. TLC: atranorin, lobaric acid.
Remarks: This species is similar to S. exutum but differs in having pigmented brown hypothecium. Hypothecium color is the most important key for identification between S. nigrum and S. exutum.
Representative specimens examined: Gyeongsangbuk-do, Danyang-gun, Mt. Sobeak, alt. 1325 m, on rock, 36°57′18.00″N, 128°28′52.03″E, 03 October 2003, J.-S. Hur, Hur 030817 (KoLRI 000608); Jellanam-do, Sancheong-gun, Mt. Jiri, alt. 1346 m, on rock, 35°19′25.02″N, 127°44′25.04″E, 16 September 2006, J.-S. Hur, Hur 060664 (KoLRI 005038).
4.6. Stereocaulon octomerellum Müll. Arg., J. Jap. Bot. 45(3): 68 (1970)
Morphology: Primary thallus persistent consisting of coarse granules; pseudopodetia 0.5–1 cm tall, erect, not dorsiventral, rarely branched, decorticate, exposing layer, firmly attached the substrate, growing on rock, phyllocladia abundant, lateral, short coralloid to granule-like, 0.1–0.5 mm long, grayish white to gray, cephalodia spherical, tubercular, gray, distributed in the basal part. Apothecia abundant, terminal, convex, brown, 1–2 mm wide, epihymenium brown, hymenium brown, 60–70 μm high, hypothecium hyaline 70–75 μm high; spore fusiform, hyaline, mostly 3–7 septate (), rarely 9–10 septate, 25–53 × 5–6 μm.
Chemistry: K + yellow, P– pale yellow. TLC: atranorin, lobaric acid.
Remarks: This is the first record of the presence of S. octomerellum in South Korea. This species is similar to S. curtatum, but the latter produces miriquidic acid. Most of coralloid group cannot be easily distinguished. Immature S. verruculigerum can be easily confused with S. octomerellum. S. octomerellum is more erect than S. verruculigerum, has short pseudopodetia (up to 1 cm), and hypothecium is almost colorless to pale brown but S. verruculigerum has brown hypothecium.
Representative specimen examined: Jeju-do, Jeju-si, Mt. Halla, alt. 1446 m, on rock, 33°22′21.09″N, 126°30′10.03″E, 04 April 2013, S.-O. Oh, J. S. Park, Hur 130063 (KoLRI 018049); the same locality, alt. 1630 m, on rock, 33°21′49.09″N, 126°30′52.06″E, 08 November 2014, J. S. Park, J.-J. Woo, D. Liu, Hur 141479 (KoLRI 032152).
4.7. Stereocaulon pileatum Ach., Lich. Univ.: 1–696 (1810)
Morphology: Primary thallus persistent consisting of wart-like, pseudopodetia erect, not dorsiventral, rarely branched, 1–4 mm tall, short and stubby, decorticate, attached directly to rock, phyllocladia granular to wart-like, short, soredia at tip of pseudopodetia, apical, capitate and powder-like, sometimes apothecia with soredia, cephalodia spherical, granular to tubercular, gray to violet, 0.5–1 mm diam., small, interspersed basal part on pseudopodetia. Apothecia rare, terminal, 0.2–0.5 mm wide, brown, subglobe to oval, epihymenium brown, 10–15 μm high, hymenium hyaline, 50–60 μm high, hypothecium brown, 40–50 μm high; spore hyaline, 3–4 septate, 23–26 × 3–4 μm.
Chemistry: K + yellow, P–pale yellow. TLC: atranorin, lobaric acid.
Remarks: S. pileatum is a very tiny species that is closely related to S. octomerellum but differs in having capitate soredia tip of pseudopodetia.
Representative specimens examined: Gyeongsan-gnam-do, Geoje-si, Mt. Geumwon, alt. 1313 m, on rock, 35°43′07.70″N, 127°45′08.26″E, 25 May 2010, X. Y. Wang, H. S. Jeon & G. S. Han, Hur 100545; Jeju-do, Jeju-si, Mt. Halla, alt. 1670 m, on rock, 33°21′15.69″N, 126°31′46.43″E, 04 Jul 2012, S. Y. Kondratyuk, L. Lokös, S.-O. Oh, S. Joshi, Hur 121822 (KoLRI 016759).
4.8. Stereocaulon sorediiferum Hue, Nouv. Arch. Mus. Hist. Nat. ser. 3, 10: 250 (sep. 38) (1898)
Morphology: Primary thallus lacking, pseudopodetia erect, not dorsiventral, well-branched, 0.8–4 cm tall, grayish, corticated, main stems rather thick, attached directly to rock, phyllocladia lateral, well-branched, coralloid, 2–2.5 mm, soredia distributed at tip of branch, capitate, powder-like, whitish, cephalodia conspicuous, common, sacculate (sack-like), gray to dark gray. Apothecia not observed.
Chemistry: K + yellow, P–Pale yellow. TLC: atranorin, lobaric acid.
Remarks: S. sorediiferum is characterized by sack-like cephalodia and soredia at the tip of phyllocladia and long pseudopodetia. S. meyery is a similar species but has perlatolic acid instead of lobaric acid. S. pileatum also has soredia but is differently located at the tip of pseudopodetia.
Representative specimen examined: Gangwon-do, Inje-gun, Temple Beakdam, alt. 465 m, on rock, 38°10′25.08″N, 128°22′21.06″E, 11 October 2004, J.-S. Hur, Hur 041554; Jeju-do, Jeju-si, Mt. Halla, alt. 1313 m, on rock, 33°21′13.00″N, 126°29′51.03″E, 04 April 2013, S.-O. Oh, J. S. Park, Hur 130032 (KoLRI 018017).
4.9. Stereocaulon verruculigerum Hue, J. Hattori bot. Lab. 43: 267 (1977)
Morphology: Primary thallus persistent as wart-like; pseudopodetia erect, 1–2 cm tall, dorsiventral, decorticate, exposing inner layer, little branched but becoming well-branched toward apices, densely phyllocladiate, phyllocladia abundant, coralloid, cylindrical, simple to branched, up to 1–2 mm long, cephalodia common, mostly located on basal part, spherical, tubercular, gray to black, 0.5–2 mm wide. Apothecia terminal, convex, reddish brown to black, 1–2 mm wide, abundant, epihymenium brown, 50–100 μm high, hymenium hyaline, 50–100 μm high, hypothecium brown, 50–100 μm high; spore fusiform, hyaline, 3–4(–6) septate, 25–50 × 2.5–5 μm.
Chemistry: K + yellow, P–pale yellow. TLC: atranorin, lobaric acid.
Remarks: This species is very similar to S. japonicum, but contains stictic acid instead of lobaric acid. S. octomerellum is also a similar species but has short pseudopodetia (up to 1 cm long). According to the Lamb key, S. verruculigerum has a primary thallus but according to the Japanese key it does not [Citation6]. “The Macrolichen flora of South Korea” states that this species has a primary thallus [Citation11]. It is very difficult to distinguish these species from S. japonicum without a TLC test.
Representative specimen examined: Jeollabuk-do, Muju-gun, Mt. Deokyu, alt. 1550 m, on rock, 35°50′22.04″N, 127°44′48.00″E, 30 April 2005, J.-S. Hur, Hur 050185 (KoLRI 003088); Gyeongsangbuk-do, Yeongju-si, Mt. Sobeak, alt. 1270 m, on rock, 36°57′19.08″N, 128°09′17.00″E, 30 Mar 2013, U. Jayalal, J. S. Park, Hur 130015 (KoLRI 018000).
4.10. Stereocaulon vesuvianum var. nodulosum (Wallr.) I.M. Lamb, Best. europ. Flecht. (Vaduz): 633 (1969)
Morphology: Primary thallus lacking, pseudopodetia decumbent, well-branched, 1.5–2.5 cm tall, decorticate, brown exposing layer, woody, twisted, main stem rather thick and robust than other branches, 1–1.5 mm wide, densely phyllocladiate, firmly attached on the rock, phyllocladia peltate with dark center (greenish gray) and pale margin (grayish white to whitish), 0.5–1 mm wide, at first convex then flattened and finally developing a depressed, slightly tumid, squamulose-like, cephalodia not seen. Apothecia terminal to lateral, brown to black, 0.5–1 mm wide, convex, epihymenium brown, 12–12.5 μm high, hymenium and hypothecium hyaline to brownish; spore hyaline, 12.5 × 2.5 μm, no septate.
Chemistry: K + yellow, P + red. TLC: atranorin, norstictic acid, stictic acid, cryptostictic acid, menegazziaic acid, and constictic acid.
Remarks: This species has peltate phyllocladia and can be easier to separate than others. This species appear near a volcano in South Korea and has many variations.
Representative specimens examined: Jeju-do, Jeju-si, Mt. Halla, alt. 1547 m, on rock, 33°22′06.09″N, 126°30′32.09″E, 04 April 2013, S.-O. Oh, J. S. Park, Hur 130060 (KoLRI 018046); Jeollanam-do, Mt. Baekun, 915 m, 35°04′09.06″ N, 127°39′24.04″E, 25 September 2004, J.-S. Hur, Hur 041296.
4.11. Species excluded from this study
According to the literature, the species of S. apocalypticum Nyl., S. commixtum (Asahina) Asahina, S. curtatum Nyl., S. octomerum Müll. Arg., S. paschale (L.) Hoffm., and S. wrightii Tuck. have been reported in Korea. However, this time we did not detect these species. We have descriptions based on previous literature.
4.11.1. Stereocaulon apocalypticum Nyl., in Middendorff, Reise Sibir. 4(6): LV (1867)
This species is characterized by phyllocladia. Phyllocladia represented by spathulate apical foliar expansions at the end of the pseudopodetia [Citation6]. It has a very distinct phyllocladia form which can be easily characterized in the field. It has previously been reported by Kim [Citation10] and has also been found in China [Citation30] and Japan [Citation13].
4.11.2. Stereocaulon commixtum (Asahina) Asahina, (1969)
This species is characterized by having miriquidic acid, densely coralloid phyllocladia and tubercular cephalodia gray to blackish. It closely resembles S. japonicum. Both species have stictic acid, but S. commixtum additionally produces miriquidic acid. S. verruculigerum is also morphologically similar, but S. verruculigerum has lobaric acid instead of stictic acid. Although the three aforementioned species are morphologically very similar, they can be distinguished using the TLC method. S. commixtum has been reported by Park in Korea [Citation11] and has also been found in Japan [Citation13].
4.11.3. Stereocaulon curtatum Nyl., Lich. Japon.: 18 (1890)
This species is characterized by having coralloid phyllocladia, broad spore size (6–9 μm) and contains miriquidic acid. It is similar to S. octomerellum but the latter has a grain-like or verruculose phyllocladia and contains lobaric acid instead of miriquidic acid. This species has been reported by Kim [Citation10] and it has been also found in Japan [Citation13].
4.11.4. Stereocaulon octomerum Müll. Arg., Flora, Regensburg 74(1): 109 (1891)
This species is characterized by evanescent primary thallus, presence of minutely granule-like to short coralloid phyllocladia and usually seven septate spores. This species is also very similar to other coralloid type Stereocaulon, but it has seven septate spores, whereas the other species have three to five septate spores. This species has been reported by Park [Citation11] and has also been found in Alaska [Citation31], Japan [Citation13], and Taiwan [Citation32].
4.11.5. Stereocaulon paschale (L.) Hoffm., Deutschl. Fl., Zweiter Theil (Erlangen): 130 (1796) [1795]
This species is characterized by having scabrid cephalodia, verrucose to digitate-squamulose phyllocladia, and thinly tomentum. It is a cosmopolitan species, widely distributed in the American arctic [Citation33], Finland [Citation34], North America [Citation35], Patagonia [Citation36], Canada [Citation2], and British Isles [Citation37]. In East Asia, it has been found in China [Citation27] and Japan [Citation13].
4.11.6. Stereocaulon wrightii Tuck., Amer. J. Sci. Arts, Ser. 2 28: 202 (1859)
This species is similar to S. apocalypticum but is distinguished by the presence of stictic acid instead of lobaric acid. It is characterized by the presence of pseudopodetia 1 to 4 cm, solitary or caespitose, branched above, apical foliar expansions of phyllocladia and cephalodia doubtful [Citation38]. It has reported by Sato [Citation9] and Kim [Citation10] and in East Asia, and has been found in Japan [Citation13].
Supplemental Material
Download PDF (84.6 KB)Supplemental Material
Download TIFF Image (1.9 MB)Acknowledgements
We are grateful to Jeong-Ae Ryu (Sunchon, South Korea) and Dr Xin Yu Wang (Kunming, China) for their specimen collection.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Smith CW, Aptroot A, Coppins BJ, et al. Lichens of Great Britain and Ireland. London: British Lichen Society; 2009.
- Goward T. The lichens of British Columbia. Illustrated Keys-Part 2: fruticose species. British Columbia: Ministry of Forests Research Branch; 1999.
- Lamb IM. On the morphology, phylogeny, and taxonomy of the lichen genus Stereocaulon. Can J Bot. 1951;29:522–584.
- Högnabba F. Molecular phylogeny of the genus Stereocaulon (Stereocaulaceae, lichenized ascomycetes). Mycol Res. 2006;110:1080–1092.
- Lamb IM. A conspectus of the lichen genus Stereocaulon (Schreb.) Hoffm. J Hattori Bot Lab. 1977;43:191–355.
- Lamb IM. Key to the species of the lichen genus Stereocaulon (Schreb.) Hoffm. J Hattori Bot Lab. 1978;44:209–250.
- Hur J-S, Koh YJ, Harada H. A checklist of Korean lichens. Lichenology. 2005;4:65–95.
- Ueda T. Lichens of Paetu. J Muse, Corea. 1934;17:61–63.
- Sato M. Index plantarum Nipponicarum IV. Tokyo: Tokyo Sci Mus; 1943.
- Kim SH. Floral studies on the lichens in Korea. Bull Kongju Teachers Col. 1981;17:279.
- Park YS. The macrolichen flora of South Korea. Bryologist. 1990;93:155–157.
- Kashiwadani H. Lichen of the Cheju island, Republic of Korea 1. The Macrolichens. National Science Museum Monographs. 2002;115–135.
- Yoshimura I. Lichen flora of Japan in color. Hoikusha Publ; 1982.
- Orange A, James PW, White FJ. Microchemical methods for the identification of lichens. London: British Lichen Society; 2010.
- Gardes M, Bruns TD. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol Ecol. 1993;2:113–118.
- White TJ, Bruns T, Lee S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Orlando (FL): Academic Press; 1990.
- Vilgalys R, Hester M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol. 1990;172:4238–4246.
- Myllys L, Lohtander K, Tehler A. β-tubulin, ITS and group I intron sequences challenge the species pair concept in Physcia aipolia and P. caesia. Mycologia. 2001;93:335–343.
- Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Acids Symp Ser. 1999;41:95–98.
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–4680.
- Tamura K, Stecher G, Peterson D, et al. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol Evol. 2013;30:2725–2729.
- Asahina Y. Lichens collected in Aogashima and Mikurazima. J Jpn Bot. 1955;30:222–224.
- Asahina Y. Lichenologische Notizen. J Jpn Bot. 1960;35:289–295.
- Asahina Y. Lichenologische Notizen. J Jpn Bot. 1961;36:46–50.
- Asahina Y. Lichenologische Notizen. J Jpn Bot. 1961;36:225–232.
- Huang MR. Noteworthy species of Stereocaulon from China. Mycosystema. 2008;27:85–90.
- Huang MR, Wei JC. Overlooked taxa of Stereocaulon (Stereocaulaceae, Lecanorales) in China. Nova Hedw. 2006;82:435–445.
- Högnabba F, Raquel PB, Nordin A, et al. Phylogenetic position of the crustose Stereocaulon species. Lichenologist. 2014;46:103–114.
- Dahl E. Studies on the macrolichens flora of Southwest Greenland. Medd Grønl. 1950;150:1–176.
- Kurokawa S. Checklist of Japanese Lichens. Tokyo: TNSM; 2003.
- Krog H. The macrolichens of Alaska. Norsk Polarinstitutt: Universitetsforlaget; 1968.
- Lai MJ. The lichen family Stereocaulaceae of Taiwan. Taiwan J For Sci. 2001;16:175–180.
- Thompson JW. American Arctic lichens. 1. The macrolichens. New York: Columbia University Press; 1984.
- Kivisto L. Taxonomy of Stereocaulon paschale and allied species in Finland. Sauteria. 1998;9:25–36.
- Brodo IM, Sharnoff SD, Sharnoff S. Lichens of North America. New Heaven (CT): Yale University Press; 2001.
- Lamb IM. New Lichens from Northern Patagonia with Notes on some related species. Farlowia. 1955;4:423–471.
- Kershaw KA. The Genus Stereocaulon Schreb. in The British Isles. Lichenologist. 1960;1:184–203.
- Riddle LW. The North American species of Stereocaulon. Bot Gaz. 1910;50:285–304.