
Abstract
Lactifluus (Pers.) Roussel is an ectomycorrhizal genus that was recently recognized to be distinct from the genus Lactarius. To date, 226 Lactifluus species have been reported worldwide. Misidentification of Lactifluus species is common because of intraspecific morphological variation, cryptic diversity, and the limited number of taxonomic keys available. Molecular data are indispensable for species delimitation; a multilocus phylogenetic analysis showed that most Asian Lactifluus species are not conspecific with morphologically similar species present on other continents. In particular, Korea has misused European and North American Lactifluus names. In this study, we evaluated the taxonomy of Lactifluus in Korea using both morphological and multilocus molecular (ITS, nrLSU, rpb1, and rpb2) data. We examined 199 Lactifluus specimens collected between 1980 and 2016, and a total of 24 species across the four Lactifluus subgenera were identified. All Korean species are distinct and clearly separated from European and North American species. Five taxa corresponded to previously described species from Asia and the remaining 19 taxa are confirmed as new species. Herein, we provide keys to the Korean Lactifluus species within their subgenera, molecular phylogenies, a summary of diversity, and detailed description of the new species.
1. Introduction
The ectomycorrhizal genus Lactifluus (Pers.) Roussel was recently recognized as a distinct genus of milkcaps [Citation1–3]. Previously, Lactifluus was considered a part of Lactarius, likely because species in both genera exude latex when the fruiting body is bruised. Molecular data showed that the original genus Lactarus was not monophyletic and warranted the creation of the new milkcap genera Lactifluus and Multifurca (the latter somtimes without latex). Upon closer inspection, Lactifluus and Lactarius have some key morphological differences. Although no synapomorphic characteristic has been found to consistently distinguish both genera, the distinction can usually be made on a combination of characters and trends. First, Lactifluus species, relative to Lactarius, are more likely have dry pilei with a velvety to pruinose texture. Also, Lactifluus species never have zonate, hairy, viscid, or scrobiculate pilei. The genus Lactifluus contains many species with veiled velvety pilei, as well as all known pleurotoid milkcap species [Citation1,Citation4]. So far, no sequestrate species are known within the genus Lactifluus. Microscopically, species of Lactifluus are characterized by having spherical cells (sphaerocytes) in the hymenophoral trama and very often cells with thickened cell walls in the pileipellis structure and/or in the hymenium [Citation5]. In many cases, a clear distinction between Lactifluus and Lactarius relies on genetic data.
To date, 226 Lactifluus species have been described worldwide [Citation6]. The species richness of Lactifluus is higher in tropical compared to temperate regions, with the highest known diversity in Africa [Citation7] and Asia [Citation8–12]. Recent studies indicate that Lactifluus is also well represented in South America [Citation13–20]. In Korea, 21 species of Lactifluus have been recorded, with the first record being Lactifluus volemus (first identified as Lactarius volemus) in 1936 [Citation21–24]. Lactifluus volemus is a typical example of a name that has been used in several continents, but covers a very rich species complex [Citation10,Citation25,Citation26].
Lactifluus species are an important resource for human use. Most Lactifluus species are edible and are important food sources in the Northern Hemisphere (e.g., Lf. piperatus and Lf. volemus). Some species are sold commercially in markets of Europe [Citation27,Citation28], Central & North America [Citation29,Citation30], and Asia [Citation27,Citation31–33]. Also, some species, such as Lf. Vellereus, are the source of useful bioactive secondary metabolites, such as sesquiterpenes with lactarane or marasmane skeletons [Citation34]. Lactifluus volemus (Fr.) Kuntze has a large amount of low-molecular-weight natural rubber in its latex that was used for revealing the mechanism of natural rubber synthesis [Citation35,Citation36].
Inaccurate description and misidentification of Lactifluus species is common in the literature due to intraspecific morphological variation and the occurrence of cryptic diversity. Verbeken and Nuytinck [Citation4] predict that cryptic species are more common in Lactifluus, a genus with a very large genetic diversity [Citation20], than in Lactarius. Identification of species is complicated by the limited number of taxonomic keys available. To counter these problems, mycologists have utilized DNA sequence data for species identification and phylogenetic studies. The most commonly used genes for identification of milkcaps (genera Lactifluus and Lactarius in the family Rusulaceae) are the internal transcribed spacer (ITS), the large subunit 28S region (nrLSU), and the region between the conserved domain 6 and 7 of the second largest subunit of RNA polymerase II (rpb2) [Citation1,Citation37,Citation38]. Several studies suggested incorporation of the region between domains A and C of the largest subunit of RNA polymerase II (rpb1) and rpb2 sequences for clearer understanding in infrageneric relationships [Citation5,Citation39,Citation40]. However, DNA data have uncovered new problems—some Lactifluus species have been identified to be species complexes and remain difficult to identify [Citation9,Citation10,Citation25,Citation26,Citation41,Citation42].
As for many other groups of macrofungi, Lactifluus research has primarily been conducted in Europe and North America, where there is a long history of mycology. In the twentieth century, the taxonomy of Lactifluus was well studied under the name of Lactarius sensu lato in Europe and North America [Citation43,Citation44]. Subsequently, mycologists studying Asian fungal diversity used the scientific names originating from Europe and North America to refer to morphologically similar individuals (e.g., Pegler & Fiard [Citation45], Singer et al. [Citation46]). Recent molecular studies, however, showed that most Asian Lactifluus species are not conspecific with morphologically similar species present on other continents [Citation8,Citation10,Citation11,Citation25,Citation42]. Recent work on Korean milkcaps revealed new species and raised doubt of previous identifications based on morphological characters [Citation23,Citation24,Citation47]. Such findings warrant a comprehensive reevaluation of Korean Lactifluus. In this study, we examine all available Lactifluus specimens from Korean herbaria to delineate species using morphological and multilocus molecular (ITS, nrLSU, rpb1, and rpb2) data.
2. Materials and methods
2.1. Sampling
A total of 199 Lactifluus specimens were obtained from four herbaria in South Korea: Kangwon National University (TPML), Korea National Arboretum (KA), National Institute of Agricultural Sciences (HCCN), and Seoul National University (SFC). These specimens were collected across South Korea from 1980 to 2016, with the initial identification based on morphological characters. When available, the collection information of each specimen was obtained from each herbarium.
2.2. Morphological approaches
Morphological characters were recorded from fresh materials, such as size and color of basidiomata, latex features, taste and smell. Specimen colors were recorded during daylight hours with color standard codes from Korneup and Wanscher [Citation48]. For macromorphological characters, we follow the terminology in Verbeken and Walleyn [Citation7]. Most collections were photographed in the field.
All microscopic characters were observed in dried specimens, using a Nikon Eclipse 80i microscope (Nikon, Japan), with the aid of the NIS element BR v3.2 (Nikon, Japan). Measurements and statistics follow Lee et al. [Citation47], and were based on at least 20 spores and 10 hymenial structures (e.g., basidia, cystidia, marginal cells and pseudocystidia). Spore size and ornamentation were measured and observed in Melzer’s reagent. The spores were measured in lateral view and the height of ornamentation was examined separately. The Q value represents the length/width ratio of the spore, referring to its shape. The spore ornamentation was photographed using a scanning electron microscope (SEM) at 5000× and 10,000× magnification (SUPRA 55VP, Carl Zeiss, Germany). Before SEM photography, dried pieces of lamellae were attached onto aluminum stubs, coated with platinum using a sputter coater (EM ACE200, Leica, Austria). Hymenial and pileal elements were mounted in 5% (w/v) KOH and observed in 5% (w/v) Congo red. The width and contents of pseudocystidia were recorded. For the features of hymenial and pileal cystidia, we recorded size, contents, abundance, and thickness of the cell wall. We documented the following pileipellis characters: overall structure, depth, delimitation, stratification, and features of dermatocystidia, if present.
2.3. Molecular approaches
We used a modified CTAB method [Citation49] to extract genomic DNA from dried tissue. Four loci were amplified and sequenced: ITS, nrLSU, rpb1, and rpb2. Molecular approaches were performed in two steps. First, we attempted to amplify and sequence ITS for all samples. Next, we provisionally identified the species based on morphological characters and the ITS-based phylogeny. For each provisional species, one to five representative specimens were selected (more recently collected) for additional amplification and sequencing of nrLSU, rpb1, and rpb2.
We used a C1000 thermal cycler (Bio-Rad, Hercules, CA, USA) for PCR amplifications using AccuPower® PCR premix (Bioneer Co., Daejeon, Korea). The primer sets and PCR conditions used to amplify ITS, nrLSU, and rpb2 region followed Lee et al. [Citation47]. For rpb1, we developed six Lactifluus-specific primers using the Primer3 program [Citation50]: rpb1-Lac1F (TTC GGG CAC ATT GAA CTG GCC), rpb1-Lac2F (CAT TGA ACT GGC CAG ACC RG), rpb1-Lac3F (TCC ATC CAG GTG CGT GGC), rpb1-Lac1R (GTC CAT RTA AGT CGC CAC GT), rpb1-Lac2R (RTA AGT CGC CAC GTG GAA CTG), and rpb1-Lac3R (ACC TGR AGR AGT TGY TCR T). The partial rpb1 locus was amplified using two steps. The first amplification of the rpb1 locus was conducted using the primer set rpb1-Lac1F/rpb1-Lac1R under the same conditions of the first rpb2 amplification in Lee et al. [Citation47]. The primer set rpb1-Lac2F/rpb1-Lac2R or rpb1-Lac3F/rpb1-Lac3R was used in the second amplification of rpb1 with the same amplification conditions previously described for nrLSU in Lee et al. [Citation47].
PCR products were visualized on a 1% agarose gel stained with EcoDye (SolGent Co., Daejeon, Korea) and purified with the Expin PCR purification kit (GeneAll Biotechnology, Seoul, Korea), according to the manufacturer’s instructions. All successfully amplified products were sequenced using the respective PCR primers on an ABI3730 automated DNA sequencer by Macrogen (Seoul, Korea). Obtained sequences were checked and manually edited using the software FinchTV v1.4 (Geospiza Inc.) and assembled in MEGA 6 [Citation51]. All concensus sequences generated in this study were deposited in GenBank ().
Table 1. GenBank accession numbers for DNA sequences of four loci, species names, voucher and potential host trees of studied specimens in this study.
2.4. Phylogenetic analyses
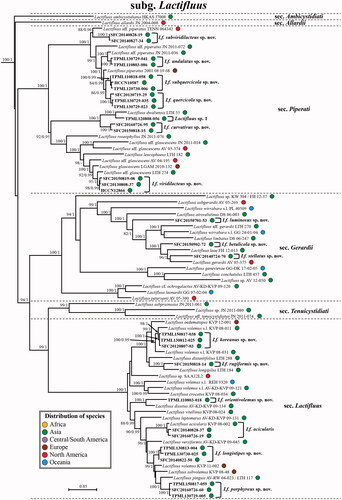
Four datasets (ITS, nrLSU, rpb1, and rpb2) were generated from the representative specimens of each species and used for phylogenetic analyses. Data for relevant species were downloaded from GenBank (shown in ), with the majority being from De Crop et al. [Citation5]. Six Lactarius species were chosen as outgroups: L. fulginosus, L. hatsudake, L. miniatescens, L. olympianus, L. scrobiculatus, and L. tenellus [Citation5]. All DNA datasets were aligned using the online version of MAFFT v7 [Citation52], using the E-INS-I strategy, then trimmed or edited in MEGA 6. Species delimitation was first examined using single locus phylogenies. When significant conflict was not observed among the single locus phylogenies, we concatenated all single locus alignments using MEGA 6. A total of 52 concatenated sequences were generated in this study, representing species in six sections/four subgenera: Lf. subg. Gymnocarpi sect. Luteoli, Lf. subg. Lactariopsis sect. Albati, three sections belonging to Lf. subg. Lactifluus (sect. Gerardii, Lactifluus, and Piperati), and Lf. subg. Pseudogymnocarpi sect. Pseudogymnocarpi.
Figure 1. Maximum Likelihood (ML) phylogenetic tree of Lactifluus subg. Pseudogymnocarpi based on concatenated ITS, nrLSU, rpb2 and rpb1 sequence data. ML bootstrap support values/Bayesian posterior probability greater than 70%/0.95 are indicated. The color of circles after the specimen voucher name indicates the continental origin of each species; Africa (orange), Asia (green), Australasia (blue), Europe (brown), Central/South America (violet), North America (red).
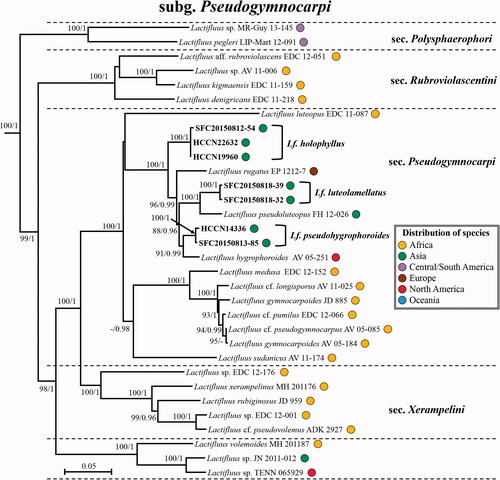
Figure 2. Maximum likelihood (ML) phylogenetic tree of Lactifluus subg. Lactariopsis based on concatenated ITS, nrLSU, rpb2 and rpb1 sequence data. ML bootstrap support values/Bayesian posterior probability greater than 70%/0.95 are indicated. The color of circles after the specimen voucher name indicates the continental origin of each species; Africa (orange), Asia (green), Australasia (blue), Europe (brown), Central/South America (violet), North America (red).
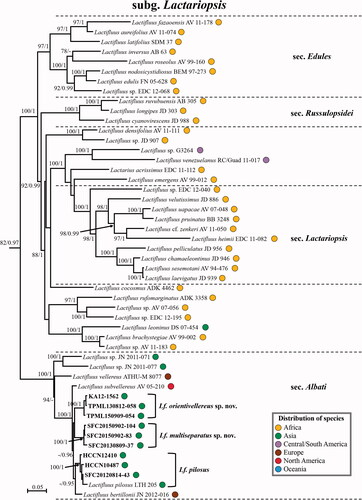
Figure 3. Maximum Likelihood (ML) phylogenetic tree of Lactifluus subg. Gymnocarpi based on concatenated ITS, nrLSU, rpb2 and rpb1 sequence data. ML bootstrap support values/Bayesian posterior probability greater than 70%/0.95 are indicated. The color of circles after the specimen voucher name indicates the continental origin of each species; Africa (orange), Asia (green), Australasia (blue), Europe (brown), Central/South America (violet), North America (red).
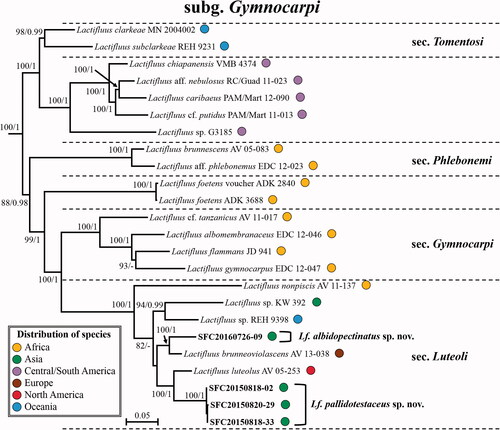
Figure 4. Maximum Likelihood (ML) phylogenetic tree of Lactifluus subg. Lactifluus based on concatenated ITS, nrLSU, rpb2 and rpb1 sequence data. ML bootstrap support values/Bayesian posterior probability greater than 70%/0.95 are indicated. The color of circles after the specimen voucher name indicates the continental origin of each species; Africa (orange), Asia (green), Australasia (blue), Europe (brown), Central/South America (violet), North America (red).
All phylogenetic analyses were performed in the CIPRES Science Gateway [Citation53]. Maximum likelihood (ML) analyses were done using RAxML 8.2.10 [Citation54]. jModeltest 2.1.7 [Citation55] was used to find best substitution model for each gene in the concatenated dataset. Models found by jModeltest 2.1.7 under Bayesian information criterion (BIC) were: K80 + I + G (ITS and nrLSU), HKY + I + G (rpb1), and SYM + I + G (rpb2). Four parallel runs of Bayesian Inference (BI), each containing one cold and three heated chains, were carried out using MrBayes v.3.2.6 [Citation56]. Each analysis was run for 20 million generations, sampling every 1000th generation. The convergence and burn-in value were visualized in Tracer 1.6 [Citation57].
3. Results
3.1. Diversity of Lactifluus in South Korea and multigene analysis
Of the 199 Korean Lactifluus specimens, ITS sequences were successfully obtained from 143 specimens. The 56 specimens without sequence data were excluded from the study. A total of 24 putative taxa were identified based on morphological characters and the ITS-based phylogenetic tree. To confirm species identity and to infer the phylogenetic relationships of Korean Lactifluus, three additional loci (nrLSU, rpb1, and rpb2) were amplified and sequenced from the 52 representative specimens of the 24 taxa. The final phylogenetic analyses were performed with datasets of four loci from 134 Lactifluus samples and six Lactarius species as outgroups. The concatenated dataset contained 3,519 bp. The ML and BI analyses generated nearly identical tree topologies with minimal variation in statistical support values. A simplified tree detailing the relationships between the subgenera of Lactifluus is shown in . In line with previous studies, the monophyly of the genus Lactifluus is well-supported (ML: 100, BI: 1.0), with four well-supported subgenera: Lf. subg. Pseudogymnocarpi (ML: 100, BI: 1.0), Lf. subg. Lactariopsis (ML: 82, BI: 0.97), Lf. subg. Gymnocarpi (ML: 100, BI: 1), and Lf. subg. Lactifluus (ML: 100, BI: 1). Each subgenus can be further divided into several sections, which is described below with their known morphological characteristics.
Of the 24 putative taxa, five correspond to previously described species: Lf. acicularis, Lf. holophyllus, Lf. luteolamellatus, Lf. pilosus, and Lf. pseudohygrophoroides. The other 19 taxa did not correspond to any known species. One of these taxa lacks micro-morphological data because of immature basiomata and thus remains unnamed until additional data are collected. The other 18 taxa are confirmed as new species after morphological comparison with phylogenetically closely related species. The 24 Korean species identified in this study are distributed across the four Lactifluus subgenera: 3 species in Lf. subg. Pseudogymnocarpi, 3 in Lf. subg. Lactariopsis, 2 in Lf. subg. Gymnocarpi, and 16 in Lf. subg. Lactifluus ().
3.2. Classification and morphological characterization of Korean Lactifluus species
Latifluus subg. Gymnocarpi, originating from Africa and Central-South America, includes four well-supported sections and two unnamed clades (). The two Korean species found in this study (Lf. albidopectinatus sp. nov. and Lf. pallidotestaceus sp. nov.) belonging to Lf. sect. Luteoli. This section contains species from all continents except Antarctica and South America. Korean species under this section are characterized by capitate elements in the pileipellis and/or marginal cells, brownish discoloring white latex, and absence of true cystidia.
Lactifluus subg. Lactariopsis has four well supported sections, some unnamed clades, and several isolated species (). The three Korean species (Lf. multiseparatus sp. nov., Lf. orientivellereus sp. nov., and Lf. pilosus) belong to Lf. sect. Albati. Species in this section are characterized by a whitish and velutinous pileus, a lamprotrichoderm as pileipellis structure, and the presence of pleuromacrocystidia.
Lactifluus subg. Lactifluus is the most diverse subgenus with four well-supported sections and two isolated clades in our analysis. This subgenus lacks African taxa. Three Korean species (Lf. betulicola sp. nov., Lf. luminosus sp. nov., and Lf. stellatus sp. nov.) belong to Lf. sect. Gerardii. These Korean species share characters like spore ornamentation forming a complete reticulum, a palisade as pileipellis structure, the absence of macrocystidia, distant lamellae, and unchanging white latex. Six Korean species (Lf. acicularis, Lf. koreanus sp. nov., Lf. longistipes sp. nov., Lf. orientivolemus sp. nov., Lf. porphyreus sp. nov., and Lf. rugiformis sp. nov.) are members of Lf. sect. Lactifluus. This section contains species occurring throughout Europe, North America, and Asia that were previously referred to as Lf. volemus. Korean species in this section have an orange to orange brown pileus, a lampropalisade as pileipellis structure, pleurolamprocystidia, and brownish discoloring latex. The remaining seven Korean species (Lf. curvativus sp. nov., Lf. quercicola sp. nov., Lf. subquercicola sp. nov., Lf. subviridilacteus sp. nov., Lf. undulatus sp. nov., Lf. viridilacteus sp. nov., and Lactifluus sp. 1) belong to Lf. sect. Piperati. All Korean species in this section are characterized by firm and whitish basidiocarps, thin-walled macrocystidia, a hyphoepithelium as pileipellis structure comprising dermatocystidia in the suprapellis, a low spore ornamentation forming an incomplete reticulum, and white latex.
Lactifluus subg. Pseudogymnocarpi contains four well supported sections and one unnamed clade (). The three Korean species (Lf. holophyllus, Lf. Luteolamellatus, and Lf. pseudohygrophoroides) are members of Lf. sect. Pseudogymnocarpi. This section contains African, European, North American, and Asian species. These Korean species all have a yellowish orange to orange colored pileus, a lampropalisade as pileipellis structure, and lack of true cystidia.
4. Taxonomy
Taxonomic keys to species in South Korea and the full descriptions of the new species are provided below. Abbreviations in and are as follows: ba basidia, cc cheilocystidia, clc cheilolamprocystidia, cmc cheilomacrocystidia, dc dermatocystidia, mc marginal cells, pc pseudocystidia, plc pleurolamprocystidia, pmc pleuromacrocystidia, pp pileipellis. All scale bars are 10 µm in length, horizontal scale bars are for pileipellis, and vertical scale bars are for other microscopic features.
4.1. Key to the species of Lactifluus subgenus Pseudogymnocarpi section Pseudogymnocarpi in South Korea
- Lamellae adnate to slightly decurrent; pileipellis a lampropalisade; lack of true cystidia
1. Lamellae yellowish; latex white, slowly changing brownishLf. luteolamellatus
1. Lamellae whitish to pale cream; latex white, unchanging2
2. Associated with Abies; pseudocystidia 3–6 µm broadLf. holophyllus
2. Associated with Pinus or Quercus; pseudocystidia 6–8 µm broadLf. pseudohygrophoroides
4.2. Key to the species of Lactifluus subgenus Gymnocarpi section Luteoli
- Brownish discoloration of latex; no true cystidia; capitate marginal cells and/or pileipellis terminal elements
1. Pileus whitish; pileus margin radially folded to pectinate in ageLf. albidopectinatus
1. Pileus grayish red to reddish brown; pileus margin entireLf. pallidotestaceus
4.3. Lactifluus subgenus Gymnocarpi section Luteoli
Lactifluus albidopectinatus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832062
Etymology: “albido” means a little soiled white color and “pectinatus” means comb-shaped. The name refers to the pileus color and pectinate margin.
Holotypus: SOUTH KOREA: Incheon-si, Ongjin-gun, Jangbong Island, alt. 70 m, N37°31′55″ E126°21′11″, 26 Jul 2016, SFC20160726-09 (holotypus, SFC)
Pileus 25–60 mm diam., convex to depressed with decurved margin; margin even and entire when young, radially folded to pectinate in age; surface dry, slightly tomentose, white (1 A-) often tinged with faint ochraceous to light brown. Lamellae adnate, sometimes slightly decurrent, rather crowded, never forked, up to 3 mm broad, with 2–4 series of lamellulae between lamellae, whitish. Stipe 30–70 × 10–20 mm, cylindrical, sometimes tapering downwards, slightly velvety; surface whitish to concolorous with pileus; base with white tomentosum. Latex abundant, white changing slowly to brownish.
Basidiospores (n = 20), (8.2–)8.4–9.3–10.1 × 7.6–8.5–9.3 µm, Q = 1.03–1.09–1.17, mostly subglobose, occasionally globose or broadly ellipsoid; ornamentation amyloid, composed of coarse ridges and spines connected by lower lines forming an almost completely reticulum, up to 0.4 µm high, sharp and irregular, often branched; plage non-amyloid to centrally amyloid. Basidia 42.5–53.5 × 8.5–9.5 µm, 4-spored, subclavate, filled with guttulate and granular contents. Pleuromacrocystidia absent. Lamellar edge consisting of marginal cells; marginal cells 9–60.5 × 2.5–5.5 µm, often projecting with slightly inflated and rounded apex with brownish contents. Pseudocystidia numerous, 3–5 µm diam., aseptate, distinctly emergent beyond the hymenium, cylindrical, never branched, with refringent contents; apex tapering. Pileipellis 180–280 µm thick, a lampropalisade; terminal elements 25–88 × 3–5 µm, parallel to ascending to upright, cylindrical, sometimes with slightly inflated apex, thin to thick-walled; subpellis composed of isodiametric cells, 10–40 µm in diam.
Habitat: Scattered to gregarious on soil in mixed forests dominated by Quercus.
Comments: Lactifluus albidopectinatus has a whitish colored pileus, which is a rare character of species in sect. Luteoli. This species is phylogenetically closely related to Lf. brunneoviolascens (), a species found in Europe, but differs in the size and shape of basidiospores. The ellipsoid basidiospores of Lf. brunneoviolascens are smaller (8–9 × 5.5–6.5 µm) than the subglobose spores of Lf. albidopectinatus [Citation58] and the terminal elements of the pileipellis are more distinctly capitate.
Lactifluus pallidotestaceus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832063
Etymology: “pallido” means pale and “testaceus” means brick-red color. The name refers to the color of the basidiomata.
Holotypus: SOUTH KOREA: Incheon-si, Ganghwa-gun, Mt. Mani, alt. 133 m, N37°36′30″ E126°25′51″, 18 Aug 2015, SFC20150818-02 (holotypus, SFC)
Pileus 30–50 mm diam., convex to depressed with decurved margin; margin even and entire; surface dry, velutinous, grayish red (7B3) to reddish brown (8D6) with whitish to faint ochraceous tinges. Lamellae adnate to broadly adnate, rather crowded, up to 5 mm broad, with 2–4 series of lamellulae between lamellae, whitish to pale cream. Stipe 45–80 × 8–15 mm, cylindrical, sometimes tapering upwards, sometimes irregularly compressed; surface slightly velvety, concolorous with pileus; base with dense white tomentosum. Latex abundant, white changing brownish slowly, staining lamellae brownish.
Basidiospores (n = 40), (7.1–)7.2–7.7–7.8–8.5 × 6.4–6.8–6.9–7.4 µm, Q = 1.03–1.13–1.25, subglobose to broadly ellipsoid; ornamentation amyloid up to 0.5 µm high, composed of lower short ridges and warts; rarely branched, never forming a reticulum; plage centrally amyloid. Basidia 56.5–64 × 9–10.5 µm, 4-spored, subclavate, mostly filled with guttulate and granular contents. Pleuromacrocystidia absent. Lamellar edge consisting of marginal cells; marginal cells 10–40 × 4–7.5 µm, subcylindrical to subclavate, some projecting with strongly inflated, capitate apex with brownish contents. Pseudocystidia rare, 2.5–5 µm diam., aseptate, distinctly emergent beyond the hymenium, cylindrical, never branched, with refringent contents; apex tapering, Pileipellis 210–300 µm thick, a lampropalisade; terminal elements ascending to upright, cylindrical with inflated and rounded, capitate apex, thin- to thick-walled, 19.5–50 × 3–6 (up to 8.5 at the apex) µm; subpellis composed of isodiametric cells, 5–18.5 µm in diam.
Habitat: Scattered to gregarious on soil in mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Incheon-si, Ganghwa-gun, Mt. Mani, alt. 183 m, N37°36′32″ E126°26′05″, 18 Aug 2015, SFC20150818-33 (SFC); Gyeonggi-do, Gwacheon-si, Mt. Cheonggye, alt. 537 m, N37°25′15″ E127°02′37″, 20 Aug 2015, SFC2015082-29 (SFC).
Comments: In the field, Lf. pallidotestaceus is easily recognized by its medium-sized, apricot colored velutinose pileus, brown staining latex, and warty basidiospores. This species differs from the only other Korean species in the same section (Lf. albidopectinatus) by the apricot colored pileus and the distinctly capitate marginal cells. This species is phylogenetically closely related to Lf. luteolus from North America (). Lactifluus pallidotestaceus has subglobose to broadly ellipsoid basidiospores and lacks cheilocystidia, while Lf. luteolus has ellipsoid basidiospores and cheilocystidia in the lamellar edge [Citation43]. Lactifluus luteolus also lacks the reddish colors in the pileus, which is off-white to buff.
4.4 Key to the species of Lactifluus subgenus Lactariopsis section Albati
- Firm and white basidiocarps; trichoderm or lamprotrichoderm as pileipellis; presence of macrocystidia
1. Lamellae grayish cream, distant; latex white, changing pale yellow when dryLf. pilosus
1. Lamellae white, crowded; latex white, unchanging2
2. Associated with Betula; pseudocystidia 4–8 µm broadLf. multiseparatus
2. Associated with Abies; pseudocystidia 2–4.5 µm broadLf. orientivellereus
4.5. Lactifluus subgenus Lactariopsis section Albati
Lactifluus multiseparatus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832064
Etymology: “multi” means many and “separatus” means furcate. The name refers to the multi-furcate lamellae.
Holotypus: SOUTH KOREA: Gangwon-do, Inje-gun, Wondae-ri birch forest, alt. 549 m, N37°59′31″ E128°12′45″, 2 Sep 2015, SFC20150902-104 (holotypus, SFC)
Pileus 50–110 mm, convex when young, later slightly depressed in the center; margin inrolled and entire; surface dry, slightly velvety, white (1 A-) with pale cream to brownish spots in age. Lamellae decurrent, crowded, up to 2 mm broad, multi-furcate, with lamellulae of different lengths, whitish to pale cream. Stipe 40–80 × 15–30 mm, cylindrical to slightly tapering downwards, often irregularly compressed, central, sometimes eccentric; surface dry, velvety, and concolorous with pileus. Latex transparent changing to white in a few second, changing pale yellow slowly, staining lamellae brownish.
Basidiospores (n = 60) (7.0–)7.1–8.0–8.1–9.1 × (5.6–)5.8–6.7–7.6 µm, Q = 1.04–1.19–1.30, subglobose to broadly ellipsoid; ornamentation amyloid, mostly up to 0.2 µm high, consisting of ridges forming an incomplete reticulum; some ridges isolated; isolated warts common. Basidia 53.5–68.5 × 8.5–12 µm, 4-spored, subclavate, filled with granular contents. Pleuromacrocystidia 66–96.5 × 6–10 µm, subfusiform, abundant; apex obtuse, rarely moniliform. Lamella edge sterile with cheilomacrocystidia and marginal cells; cheilomacrocystidia 56.5–73.5 × 6–10 µm, narrowly clavate, abundant, with apex obtuse to subacute; marginal cells 12.5–30.5 × 4–8 µm, cylindrical to subclavate, 1–2 septate. Pseudocystidia rather abundant, 4–8 µm diam., aseptate, indistinctly emergent beyond the hymenium, cylindrical, often branched, with refringent contents; apex rounded or inflated. Pileipellis a lamprotrichoderm, 120–280 µm thick; terminal elements 40–210 × 4–7 µm, parallel to erect, cylindrical, rarely 1–2 septate, thin to thick walled; subpellis composed of isodiametric cells, 4–22 µm in diam.
Habitat: Scattered to gregarious on soil in mixed forests dominated by Betula.
Additional studied material: SOUTH KOREA: Gangwon-do, Inje-gun, Wondae-ri birch forest, alt. 493 m, N37°59′28″ E128°13′26″, 2 Sep 2015, SFC20150902-83 (SFC); Gyeongsangbuk-do, Sangju-si, Seongjubong Natural Recreation Forest, alt. 264 m, N36°31′59″ E128°02′24″, 9 Aug 2013, SFC20130809-37 (SFC).
Comments: Lactifluus multiseparatus is characterized by whitish basidiomata with transparent latex that changes yellowish when isolated and has brownish discoloration on lamellae. Another white Korean species (Lf. orientivellereus) is phylogenetically closely related to Lf. multiseparatus (). However, the former species is distinguished from the latter species by unchanging white latex and less interconnection of spore ornamentation.
Lactifluus orientivellereus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832065
Etymology: “orienti” means eastern and ‘vellereus’ refers to the European Lactifluus vellereus. This name refers to the similar morphology with Lf. vellereus and its origin in East Asia (South Korea).
Holotypus: SOUTH KOREA: Gangwon-do, Hongcheon-gun, Mt. Baegam, alt. 937 m, N37°50′50″ E128°10′44″, 9 Sep 2015, TPML150909-054 (holotypus, TPML; isotypus, SFC)
Pileus 80–170 mm, convex when young, later infundibuliform and deeply depressed in the center; margin inrolled when young, expanding in age, sometimes wavy; surface dry, velvety, white (A1-) tinged with ocherous to yellowish brown. Lamellae slightly decurrent, very crowded, up to 3 mm broad, sometimes forked, with abundant lamellulae of different lengths, white turning grayish yellow when bruised. Stipe 10–45 × 10–25 mm, cylindrical to slightly tapering downwards, central, sometimes eccentric; surface dry, velvety, and concolorous with pileus. Latex white, unchanging.
Basidiospores (n = 40) (6.8–)7.2–8.2–8.4–9.6 × (5.8–)6.0–6.4–6.5–7.3(–7.5) µm, Q = 1.08–1.26–1.48, subglobose to ellipsoid; ornamentation amyloid, mostly up to 0.2 µm high, consisting of irregular or linear warts, often connected by an incomplete to almost complete reticulum; isolated warts sometimes present; plage non-amyloid. Basidia 46–61 × 8–11 µm, 4-spored, subcylindrical to subclavate, filled with guttulate and granular contents. Pleuromacrocystidia 76–110.5 × 8–13 µm, subfusiform to narrowly fusiform, abundant; apex obtuse, subacute to moniliform. Lamella edge sterile with cheilomacrocystidia and marginal cells; cheilomacrocystidia 67–87 × 9–11.5 µm, narrowly clavate, abundant, apex obtuse to moniliform; marginal cells 11–36 × 3.5–8 µm, cylindrical to subclavate, 1–3 septate. Pseudocystidia very abundant, 2–4.5 µm diam., aseptate, distinctly emergent beyond the hymenium, cylindrical, often branched, with refringent contents; apex rounded. Pileipellis a lamprotrichoderm, 180-300 µm thick; termenal hyphae 40–250 × 3–5 µm, mostly erect, cylindrical, 1–2 septate, thin to thick walled; subpellis composed of filamentous hyphae and isodiametric cells, 5–12 µm in diam.
Habitat: Solitary to scattered on soil in Abies holophylla forest.
Additional studied material: SOUTH KOREA: Gangwon-do, Pyeongchang-gun, Mt. Odae, alt. 916 m, N37°46′42″ E128°34′24″, 12 Aug 2013, TPML130812-058 (TPML); Chungcheongnam-do, Geumsan-gun, Mt. Seodae, alt. 352 m, N36°13′45″ E127°31′50″, 26 Sep 2012, KA12-1562 (KA).
Comments: Lactifluus orientivellereus is phylogenetically closely related to Lf. bertillonii, Lf. vellereus, and Lf. subvellereus (). These latter three species are not found in Asia and can be distinguished from Lf. orientivellereus by the more distant lamellae and smaller size of the pleurocystidia: Lf. bertillonii (50–75 × 6–12 µm), Lf. vellereus (50–85 × 6–12 µm), and Lf. subvellereus (45–60 × 5–7.5 µm) [Citation43,Citation44]. Lactifluus orientivellereus is also phylogenetically closely related to Lf. pilosus found in Asia (Thailand and Korea). However, Lf. pilosus interacts with Castanopsis or Lithocarpus [Citation8], while Lf. orientivellereus grows under Abies trees. Additionally, Lf. orientivellereus has more crowded lamellae compared to the aforementioned four species.
4.6. Key to the species of Lactifluus subgenus Lactifluus section Gerardii
- Distant lamellae; white spore print; reticulate spore ornamentation; palisade structure in the pileipellis; no macrocystidia
1. Latex yellowish brown to light brown, staining lamellae reddish brownLf. ochrogalactus
1. Latex white, unchanging2
2. Pileus pale yellow to grayish orange; pileipellis 45–80 µm thickLf. luminosus
2. Pileus brown to dark brown; pileipellis thicker than 70 µm3
3. Associated with Betula; pseudocystidia 4–8 µm broad Lf. betulicola
3. Associated with Pinus or Quercus; pseudocystidia 2–4 µm broadLf. stellatus
4.7. Lactifluus subgenus Lactifluus section Gerardii
Lactifluus betulicola H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832066
Etymology: “betuli” means birch tree and “cola” means dweller. The name refers to the presence of the species in Betula forest and the putative ectomycorrhizal association with Betula.
Holotypus: SOUTH KOREA: Gangwon-do, Inje-gun, Wondae-ri birch forest, alt. 571 m, N37°59′43″ E128°11′59″, 19 Jul 2013, SFC20150902-72 (holotypus, SFC)
Pileus 30–70 mm diam., planoconvex to concave, center depressed, sometimes papillate; margin irregularly wavy to slightly crenate; surface dry, velutinous, rugulose to rugose, grayish brown (6E5) to dark brown (7F6), sometimes with whitish spots. Lamellae deeply decurrent, distant, up to 7 mm broad, white to pale cream, pale brown in age, not staining when bruised; decurrent teeth concolorous with stipe apex. Stipe 25–50 × 5–10 mm, cylindrical, slightly tapering downwards; surface dry, velutinous, concolorous with pileus, sometimes with whitish spots. Latex abundant, watery white, unchanging.
Basidiospores (n = 20) 6.6–7.8–8.5(–8.9) × (6.0–)6.1–6.9–7.8 µm, Q = 1.00–1.13–1.23, globose to broadly ellipsoid; ornamentation amyloid, mostly up to 0.7 µm high, consisting of low and rounded ridges, completely reticulate; isolated warts rare; plage almost totally amyloid. Basidia 47–65.5 × 8.5–11 µm, subclavate to narrowly subclavate, 4-spored, filled with granular contents. Macrocystidia absent. Lamellar edge sterile, filled with marginal cells; marginal cells 10–40.5 × 3.5–6 µm, narrowly subcylindrical, sometimes slightly tortuous, with rounded apex. Pseudocystidia abundant, 4–8 µm diam., cylindrical to irregular, sometimes branching. Pileipellis a palisade, 85–160 µm thick; terminal elements 16.5–29 × 3.5–7 µm, subcylindrical, mostly erect, sometimes oblique; subpellis composed of globose to subglobose cells, 9–38 µm in diam.
Habitat: Solitary to gregarious on soil in deciduous forests dominated by Betula.
Comments: Lactifluus betulicola is phylogenetically closely related to typical sect. Gerardii species, Lf. gerardii from North America and Lf. leae from Thailand (). However, this species differs from Lf. gerardii by the size of basidiospores and the lack of macrocystidia. Lactifluus gerardii has macrocystidia and bigger basdiospores (8–10 × 7.5–9 µm) than Lf. betulicola [Citation43]. In addition, Lf. betulicola is associated with Betula trees while Lf. leae grows with Dipterocarpus, Castanopsis, and Lithocarpus [Citation59]. The almost totally amyloid plage is a rather unique character, most often encountered in the pleurotoid species of this section.
Figure 5. Maximum Likelihood (ML) phylogram of Korean Lactifluus inferred using RAxML based on a concatenated dataset of ITS, nrLSU, rpb2, and rpb1. ML bootstrap support values/Bayesian posterior probability greater than 70%/0.95 are indicated.
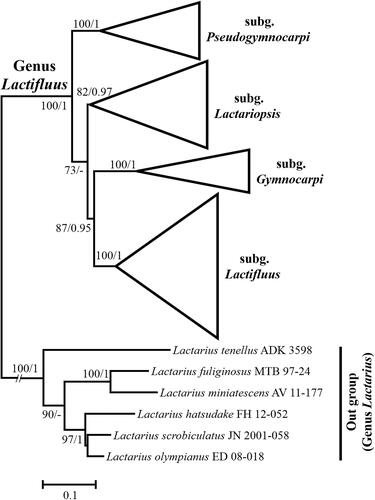
Figure 6. Microscopic features of Lactifluus albidopectinatus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
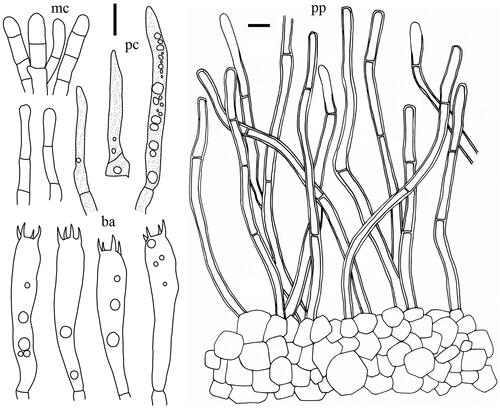
Figure 7. Microscopic features of Lactifluus pallidotestaceus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
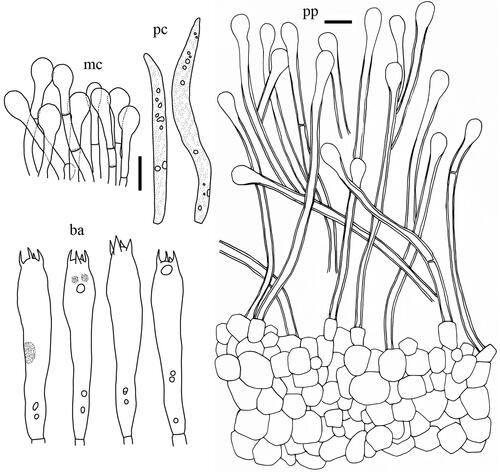
Figure 8. Microscopic features of Lactifluus multiseparatus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
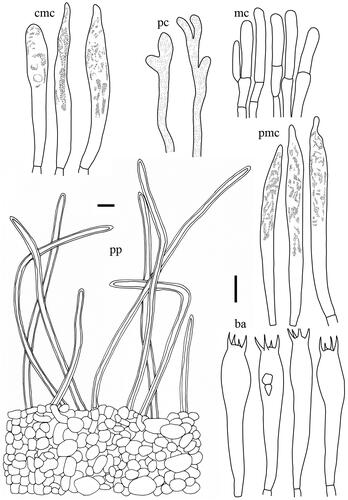
Lactifluus luminosus H. Lee & Y. W. Lim sp. nov. ()
Figure 25. Scanning electron micrographs of basidiospores from Korean species of Lactifluus subg. Pseudogymnocarpi (a∼c), subg. Lactariopsis (d∼f), subg. Gymnocarpi (g∼h). (a) Lf. holophyllus, (b) Lf. luteolamellatus, (c) Lf. pseudohygrophoroides, (d) Lf. multiseparatus, (e) Lf. orientivellereus, (f) Lf. pilosus, (g) Lf. albidopectinatus, (h) Lf. pallidotestaceus. Scale bar = 2 µm.
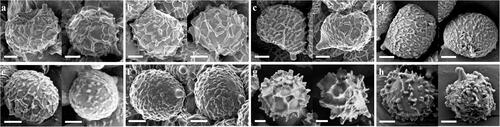
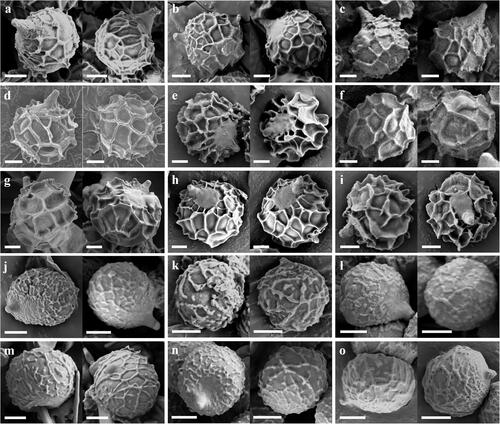
Figure 26. Fruiting bodies of Korean species of Lactifluus subg. Lactifluus sect. Gerardii (a∼c), sect. Lactifluus (d∼i), sect. Piperati (j∼o). (a) Lf. betulicola, (b) Lf. luminosus, (c) Lf. stellatus, (d) Lf. acicularis, (e) Lf. koreanus, (f) Lf. longistipes, (g) Lf. orientivolemus, (h) Lf. porphyreus, (i) Lf. rugiformis, (j) Lf. curvativus, (k) Lf. quercicola, (l) Lf. subquercicola, (m) Lf. subviridilacteus, (n) Lf. undulatus, (o) Lf. viridilacteus. The color of the two circles at the top-right of each species picture indicates the color (left circle) and color change (right circle) of the latex. Scale bar = 1 cm.

Figure 27. Scanning electron micrographs of basidiospores from Korean species of Lactifluus subg. Lactifluus sect. Gerardii (a∼c), sect. Lactifluus (d∼i), sect. Piperati (j∼o). (a) Lf. betulicola, (b) Lf. luminosus, (c) Lf. stellatus, (d) Lf. acicularis, (e) Lf. koreanus, (f) Lf. longistipes, (g) Lf. orientivolemus, (h) Lf. porphyreus, (i) Lf. rugiformis, (j) Lf. curvativus, (k) Lf. quercicola, (l) Lf. subquercicola, (m) Lf. subviridilacteus, (n) Lf. undulatus, (o) Lf. viridilacteus. The color of the two circles at the top-right of each species picture indicates the color (left circle) and color change (right circle) of the latex. Scale bar = 2 µm.
MycoBank: MB 832067
Etymology: “luminosus” means bright. The name refers to the bright and yellowish colored pileus.
Holotypus: SOUTH KOREA: Jeju-do, Jeju-si, Dongbaek-dongsan wetland, alt. 150 m, N33°30′58″ E126°43′23″, 1 Jul 2015, SFC20150701-53 (holotypus, SFC)
Pileus 10–45 mm diam., convex with depressed center to irregularly concave; center with small papilla; surface slightly rugulose when young, becoming strongly radially wrinkled in age; margin irregularly wavy to slightly crenate; surface, dry, velutinous, pale yellow (4A3) to grayish orange (5B4) with darker center, becoming paler toward margin with whitish spots. Lamellae deeply decurrent, distant, up to 8 mm broad, whitish to pale cream colored. Stipe 40–95 × 6–20 mm, cylindrical, often tapering downwards, sometimes slightly curved, sometimes subfasciculate to fasciculate with 2 basidiomata with connected stipes; surface dry, faintly longitudinally wrinkled, minutely velutinous, concolorous with pileus or paler, sometimes with whitish spots. Latex moderately abundant, white or watery white, unchanging.
Basidiospores (n = 20) 7.1–8.1–9.1(–9.2) × (6.1–)6.5–7.5–8.6 µm, Q = 1.01–1.08–1.15, globose to subglobose; ornamentation amyloid, consisting of low and straight ridges, mostly up to 0.5 µm high, completely reticulate, isolated warts very rare; plage totally amyloid. Basidia 45–60 × 8–10.5 µm, subclavate, 4-spored, filled with granular contents. Macrocystidia absent. Lamellar edge sterile, composed of marginal cells; marginal cells 17–25 × 3.5–5 µm, subcylindrical to narrowly subcylindrical, tapering toward apex. Pseudocystidia numerous, 1.5–3.5 µm diameter, cylindrical, sometimes branching; apex obtuse, sometimes irregular. Pileipellis a palisade, 45–80 µm thick; terminal elements 10–28 × 4.5–6.5 µm, subcylindrical, sometimes septate, mostly erect, sometimes oblique; subpellis composed of globose to subglobose cells, 7–22.5 in diam.
Habitat: Solitary to gregarious on soil in evergreen Quercus forests.
Comments: Lactifluus luminosus can be easily recognized by its strikingly bright yellow pileus. This species has the lightest basidiomata among the three Korean species (Lf. luminosus, Lf. betulicola, and Lf. stellatus) in the Lf. sect. Gerardii. In addition, the relatively low ornamentation of basidiospores and short margical cells are useful characters to identifiy this species.
Lactifluus stellatus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832068
Etymology: “stellatus” means star or star shaped. The name refers to numerous, tiny white spots on pileus, which is reminiscent of star clusters.
Holotypus: SOUTH KOREA: Jeollabuk-do, Jangsu-gun, Bangwhadong Natural Recreation Forest, alt. 441 m, N35°34′59″ E127°31′53″, 24 Jul 2014, SFC20140724-70 (holotypus, SFC)
Pileus 10–40 mm diam., convex to concave when young, irregularly infundibuliform in age, center depressed, rugulose with radical wrinkles; margin irregularly wavy, often crenate; surface dry, velutinous, light brown (6D4) to dark brown (9F5) with numerous white spots, scattered like a star cluster. Lamellae deeply decurrent, distant, up to 12 mm broad, white to pale cream, staining orange brown when bruised; decurrent teeth concolorous with stipe apex. Stipe 25–40 × 5–10 mm, subcylindrical, tapering downwards; surface dry, velutinous, concolorous with pileus. Latex abundant, watery white, unchanging.
Basidiospores (n = 20) 5.8–7.1–8.4(–8.5) × 5.4–6.5–7.7(–7.8) µm, Q = 1.02–1.09–1.15, globose to subglobose; ornamentation amyloid, up to 0.3 µm high, consisting of low and rounded ridges, completely reticulate; isolated warts rare; plage centrally amyloid. Basidia 52.5–65.5 × 7.5–11 µm, subclavate, 2-or 4-spored, filled with granular contents. Macrocystidia absent. Lamellar edge sterile, composed of marginal cells; marginal cells 7.5–31 × 4.5–8 µm, subcylindrical to subclavate, 1–2 septate. Pseudocystidia rare, 2–4 µm diam., cylindrical, sometimes branching; apex obtuse. Pileipellis a palisade, 70–110 µm thick; terminal elements 8.5–25 × 2–4.5 µm, subcylindrical, sometimes septate, mostly erect, sometimes oblique; subpellis composed of globose to subglobose cells, 5.5–23 µm in diam.
Habitat: Solitary to gregarious on soil in mixed forests dominated by Quercus and Pinus.
Comments: Lactifluus stellatus is a unique species in the sect. Gerardii, and this species can be characterized by the uneven dark brown color of pileus with striking whitish spots, the strong rugulose surface, and the low ornamentation of reticulate basidiospores. This species is phylogenetically closely related to Lf. gerardii, Lf. leae, and Lf. betulicola, but can be distinguished from the aforementioned three species by the height of spore ornamentation. The basidiospore ornamentation Lf. gerardii (0.5–0.8 µm), Lf. leae (0.5–1.0 µm) and Lf. betulicola (up to 0.7 µm) is distinctly higher than that of Lf. betulicola (maximum 0.3 µm) [Citation41].
4.8. Key to the species of Lactifluus subgenus Lactifluus section Lactifluus
- Grayish-yellow, orange to orange-brown or reddish-brown pileus; white latex discoloring brownish on lamellae; reticulate spore ornamentation; lampropalisade type pileipellis; presence of lamprocystidia
1. Pileus light yellow to light brown, spore ornamentation up to 2.0 µmLf. pinguis
1. Pileus darker colored, spore ornamentation never up to 2.0 µm2
2. Pileipellis terminal elements up to 100 µm or more3
2. Pileipellis terminal hyphae never up to 100 µm4
3. Spore ornamentation up to 1.4 µm, pileus brown to orange brownLf acicularis
3. Spore ornamentation up to 0.9 µm, pileus light orangeLf. orientivolemus
4. Subpellis thickness up to 140 µm. Lf. koreanus
4. Subpellis thickness never up to 140 µm5
5. Associated with Pinus treesLf. porphyreus
5. Associated with Quercus trees6
6. Ratio of stipe length/pileus diameter > 1.5, spore ornamentation up to 0.6 µmLf. longistipes
6. Ratio of stipe length/pileus diameter < 0.7, spore ornamentation up to 1.7 µmLf. rugiformis
4.9. Lactifluus subgenus Lactifluus section Lactifluus
Lactifluus koreanus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832069
Etymology: “koreanus” means Korean. The name refers to the distribution of this species. This species is very common in Korea, including Jeju Island.
Holotypus: SOUTH KOREA: Jeju-do, Seogwipo-si, Hannam Experimental Forest, alt. 451 m, N33°22′29″ E126°39′54″, 7 Aug 2015, SFC20120807-03 (holotypus, SFC)
Pileus 35–85 mm diam., convex when young, becoming applanate with slightly depressed center to infundibuliform when mature, rugulose when young; margin entire, sometimes wavy; surface dry, slightly velutinous, pruinose when young, light orange (6A4) to grayish red (7B6), getting paler toward margin, sometimes with orange brown tinges. Lamellae subdecurrent to decurrent, crowded, up to 6 mm broad, sometimes forked near the stipe, with numerous lamellulae of different length, cream colored, discoloring grayish brown to dark brown when bruised. Stipe 45–75 × 5–15 mm, cylindrical to slightly tapering upwards, centrally attached, surface dry, sometimes velutinous, pruinose when young, concolorous to pileus or darker tinged often with brownish. Latex abundant, sticky, white, slowly changing to brown.
Basidiospores (n = 40) 7.1–7.8–8.0–8.6 × 6.6–7.3–7.4–8.2(–8.3) µm, Q = 1.02–1.06–1.11, globose to subglobose; ornamentation amyloid, up to 1.3 µm high, forming a complete reticulum, with rather irregular edges; isolated warts rare; plage totally amyloid. Basidia 46.5–61 × 10–13 µm, subclavate to clavate, 4-spored, filled with guttulate and granular contents. Pleurolamprocystidia abundant, 50–95 × 6–10 µm, cell wall 1.5–3.5 µm thick, subfusiform to subcylindrical, with acute apex, emergent, arising from the subhymenium. Lamellar edge composed of basidia, cheilocystidia, and marginal cells; cheilocystidia 21.5–64.5 × 3.5–7.5 µm, mostly thick-walled, fusiform to subfusiform, with subacute to acute apex; marginal cells 7–26 × 2.5–7 µm, subcylindrical to clavate, 1–2 septate. Pseudocystidia rather abundant, 2–5.5 µm diam., emergent, subcylindrical, often branched; apex obtuse. Pileipellis a lampropalisade; terminal elements 14–42.5 × 2–4 µm, cell wall 0.5–1.0 µm thick, subcylindrical to subfusiform with tapering apices, acuminate or obtuse apex; subpellis 85–140 µm thick, composed of rounded to elongated, thick-walled cells, 6–20 µm in diam.
Habitat: Scattered on soil in Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Gangwon-do, Pyeongchang-gun, Mt. Odae, alt. 874 m, N37°46′51″ E128°34′41″, 12 Aug 2013, TPML130812-025 (TPML); Gangwon-do, Chuncheon-si, Mt. Daeryong, alt. 748 m, N37°50′47″ E127°48′59″, 17 Aug 2015, TPML150817-038 (TPML).
Comments: Lactifluus koreanus shares common characters of the Lf. volemus group such as an orange-colored pileus, brownish discoloring white latex, and pleurolamprocystidia. Lactifluus koreanus has the thickest subpellis (85–140 µm) among the Korean species in Lf. sect. Lactifluus. This species is phylogenetically closely related to Lf. oedematopus distributed in Europe (). However, Lf. oedematopus has larger basidiospores (7.5–8.4–8.9–10.1 × 6.7–7.5–8.2–9.2 µm) and broader terminal elements (3–10 µm) than those of Lf. koreanus [Citation26].
Lactifluus longistipes H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832070
Etymology: “longi” means long and “stipes” means stipe. The name refers to the long stipe relative to the pileus diameter.
Holotypus: SOUTH KOREA: Gangwon-do, Inje-gun, Mt. Jeombong, alt. 813 m, N38°02′35″ E128°27′40″, 22 Aug 2014, SFC20140822-50 (holotypus, SFC)
Pileus 35–70 mm diameter, convex when young, becoming applanate with depressed center when mature; margin entire; surface dry, velvety, smooth, dark brown to reddish brown (7E8–8E7) in the center, gradually getting paler toward margin; margin tinged with grayish brown. Lamellae subdecurrent to decurrent, crowded, up to 4 mm broad, with 3–4 series of lamellulae between lamellae, pale cream, discoloring ocher when bruised. Stipe 70–105 × 8–15 mm, cylindrical to slightly tapering upwards, centrally attached, surface dry, velvety, longitudinally rugose, yellowish orange, paler upward. Latex copious, sticky, white changing light brown.
Basidiospores (n = 40) 7.1–8.3–8.6–9.6 × 6.9–7.9–8.0–9.1 µm, Q = 1.00–1.05–1.06–1.10, globose to subglobose; ornamentation amyloid up to 0.6 µm high, almost completely reticulate, warts rare; plage distally to almost totally amyloid. Basidia 45–52 × 9–12.5 µm, clavate to subclavate, 4-spored, filled with granular contents. Pleurolamprocystidia 52–90.5 × 6.5–10 µm, thick-walled with cell wall 1.5–3.5 µm thick, fusiform to subfusiform, acuminate, sometimes obtuse, emergent, arising from the subhymenium, abundant. Lamellae-edge composed of basidia, cheilocystidia, and marginal cells; cheilocystidia 40–59.5 × 5.5–8.5 µm, mostly thick-walled (1–2.5 µm), fusiform to subfusiform, acuminate; marginal cells 10–35.5 × 3–9 µm, subcylindrical to clavate, often septate. Pseudocystidia abundant, 2.5–4.5 µm diameter, emergent, cylindrical; apex obtuse. Pileipellis a lampropalisade; terminal elements 8.5–58.5 × 2–4 µm, cell wall 0.5–1.5 µm thick, subcylindrical to subfusiform, slightly acuminate or obtuse apex; subpellis 65–110 µm thick, composed of rounded to elongated cells, thick-walled, 5.5–30 µm in diam.
Habitat: Solitary or scattered on soil in Quercus forests.
Additional studied material: SOUTH KOREA: Gyeongsangbuk-do, Uljin-gun, Mt. Tonggo, alt. 506 m, N36°55′17″ E129°11′34″, 30 Jul 2013, TPML130730-025 (TPML); Gangwon-do, Hongcheon-gun, Mt. Odae, alt. 876 m, N37°40′48″ E128°27′09″, 13 Aug 2013, TPML130813-004 (TPML).
Comments: Lactifluus longistipes can be easily distinguished from other species in sect. Lactifluus by its long stipe relative to the pileus size. The ratio of stipe length/pileus diameter is between 1.5 and 2. This species also has the lowest spore ornamentation among the Korean species of Lf. sect. Lactifluus. This species is phylogenetically closely related to Lf. volemus and Lf. subvolemus from Europe (). However, these two European species have larger basidiospores and higher spore ornamentation (up to about 1.5 µm in Lf. volemus and up to about 1.6 µm in Lf. subvolemus) than those of Lf. longistipes (up to 0.6 µm) [Citation26].
Lactifluus orientivolemus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832071
Etymology: “orienti” means eastern and “volemus” refers to the European Lactifluus volemus. This name refers to the similar morphology with Lf. volemus and its origin in East Asia (South Korea).
Holotypus: SOUTH KOREA: Gyeongsangbuk-do, Sangju-si, Mt. Songni, alt. 419 m, N36°28′58″ E127°56′23″, 3 Aug 2011, TPML110803-018 (holotypus, TPML; isotypus, SFC)
Pileus 40–80 mm diam., convex with slightly depressed center when young, becoming applanate to deeply depressed, smooth to slightly rugulose when young; margin entire, sometimes wavy in age; surface dry, pruinose, velutinous, very pale yellowish white or straw-colored, locally slightly browning, light orange (5A5) in center and light yellow (4A5) on margin. Lamellae decurrent with small tooth, crowded, up to 5 mm broad, with numerous lamellulae of different length, whitish to cream, discoloring pale brown when bruised. Stipe 40–80 × 10–18 mm, cylindrical to slightly tapering downwards, centrally attached, surface dry, yellowish brown tinged with orange brown. Latex abundant, sticky, white changing light brown.
Basidiospores (n = 20) 8.2–9.1–9.9 × 7.6–8.6–9.5 µm, Q = 1.03–1.06–1.09, globose to subglobose; ornamentation amyloid, up to 0.9 µm high, composed of ridges forming a complete reticulum; isolated warts rare; plage distally to totally amyloid. Basidia 56.5–69 × 9.5–13 µm, clavate to subclavate, 4-spored, filled with granular contents. Pleurolamprocystidia 65–149.5 × 6.5–11 µm, cell wall 2.5–4 µm thick, subfusiform to subclavate, acuminate, strongly emergent, arising from the subhymenium, abundant. Lamellar edge composed of basidia, cheilocystidia, and marginal cells; cheilocystidia 30.5–71.5 × 3–6.5 µm, mostly thick-walled (0.5–3 µm), subfusiform, acuminate; marginal cells 10–32 × 3.5–6 µm, subcylindrical to cylindrical, 1–3 septate. Pseudocystidia 2.5–4.5 µm diameter, slightly emergent, cylindrical to conspicuously tortuous, often irregularly branched; apex obtuse. Pileipellis a lampropalisade; terminal elements 26.5–138 × 2–5.5 µm, cell wall 0.5–2 µm thick, subcylindrical to subfusiform, with acuminate apex; subpellis 55–110 µm thick, composed of rounded to elongated cells, thick-walled, 4.5–24 µm in diam.
Habitat: Scattered on soil in the mixed forests dominated by Quercus and Pinus.
Comments: Lactifluus orientivolemus has the longest terminal elements in the pileipellis among the Korean species of this section. This species is macromorphologically very similar to Lf. rugiformis, which can cause misidentifications in the field. However, microscopic chracters of Lf. orientivolemus, such as larger pleurolamprocystidia and basidia, distinguish the species from Lf. rugiformis. Lactifluus orietivolemus is phylogenetically closely related to Lf. dissitus from India (). However, Lf. dissitus has shorter terminal elements in the pileipellis (11–55 × 3–6 µm) [Citation25] than Lf. orientivolemus (26.5–138 × 2–5.5 µm).
Lactifluus porphyreus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832072
Etymology: “porphyreus” means red brown. The name refers to the reddish-brown colored pileus of this species.
Holotypus: SOUTH KOREA: Incheon-si, Ongjin-gun, Jangbong-do Island, alt. 71 m, N37°32′09″, E126°20′53″, 26 Jul 2016, SFC20160726-60 (holotypus, SFC)
Pileus 45–100 mm diam., convex when immature, becoming applanate, subumbilicate or infundibuliform when mature; margin regular or slightly wavy; surface dry, velutinous, pruinose when young, rugose at margin, ochraceous orange (5A8–6B7) with reddish brown (5A6) tinges, getting paler toward the margin. Lamellae subdecurrent to decurrent, rather crowded, up to 5 mm, with numerous lamellulae of different length, whitish cream, discoloring brown when bruised. Stipe 45–90 × 8–20 mm, cylindrical to slightly tapering downwards, centrally attached, surface dry, slightly velutinous, concolorous with pileus, often getting paler toward the top. Latex abundant, sticky, white turning light brown slowly.
Basidiospores (n = 40) 7.8–8.6–8.7–9.3(–9.5) × 7.0–7.9–8.1–8.9(–9.0) µm, Q = 1.02–1.07–1.08–1.14, globose to subglobose, sometimes broadly ellipsoid; ornamentation amyloid; forming complete reticulum, ridges up to 0.8 µm high, isolated warts very scarce; plage distally to almost complete amyloid. Basidia 47–61.5 × 11–12.5 µm, clavate to subclavate, 4-spored, filled with guttulate and granular contents. Pleurolamprocystidia 60–115.5 × 7.5–13 µm, cell wall 2.5–6 µm thick, fusiform, subfusiform to subclavate, acuminate or with obtuse apex, sometimes with wavy outline, distinctly emergent, arising from the subhymenium, very abundant. Lamellae-edge composed of basidia, cheilocystidia, and marginal cells; cheilocystidia 24–61 × 3.5–9.5 µm, mostly thick-walled, fusiform to subfusiform, acuminate or furcate apex, sometimes with wavy outline; marginal cells 8.5–30.5 × 2–8 µm, subcylindrical to cylindrical, 1–2 septate. Pseudocystidia numerous, 2.5–4.5 µm diameter, slightly to distinctly emergent, cylindrical to tortuous; apex obtuse. Pileipellis a lampropalisade; terminal elements 10.5–63.5 × 3–5 µm, cell wall 0.5–1.5 µm thick, subcylindrical to subfusiform, with acuminate apex; subpellis 60–100 µm thick, composed of rounded to elongated cells, with thick-wall, 5–22 µm in diam.
Habitat: Scattered on soil in Pinus forest or mixed forests dominated by Quercus and Pinus.
Additional studied material: SOUTH KOREA: Gyeongsangbuk-do, Yeongcheon-si, Mt. Bohyeon, alt. 659 m, N37°50′34″ E128°59′01″, 29 Jul 2013, TPML130729-005 (TPML); Gangwon-do, Chuncheon-si, Mt. Daeryong, alt. 591 m, N37°50′34″ E127°48′35″, 17 Aug 2015, TPML150817-059 (TPML).
Comments: Lactifluus porphyreus can be easily distinguished from other Korean Lactifluus species belonging this section by its pale yellowish-orange pileus and ectomycorrhizal host (Pinus). This species is phylogenetically closely related to Lf. volemus and Lf. subvolemus (). However, the latter two species have larger basidiospores and higher spore ornamentations than Lf. porphyreus [Citation26].
Lactifluus rugiformis H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832073
Etymology: “rugiformis” means rugulose or wrinkled. The name refers to the distinctly rugulose pileus margin of this species.
Holotypus: SOUTH KOREA: Incheon-si, Ganghwa-gun, Mt. Mani, alt. 149 m, N37°36′29″ E126°26′02″, 18 Aug 2015, SFC20150818-14 (holotypus, SFC)
Pileus 50–110 mm diameter, convex with depressed center; margin decurved, smooth in age, distinctly rugulose toward the margin; surface dry, velutinous, rusty orange (6C8–7C8) tinged with a more brownish, darker in the center, slightly paler toward the margin. Lamellae decurrent, rather crowded, up to 3 mm broad, rarely furcate, with numerous lamellulae of different length, cream to pale orange. Stipe 30–70 × 15–20 mm, subcylindrical, tapering downwards, centrally attached, surface dry, velutinous, longitudinally rugulose, concolorous with pileus. Latex abundant, sticky, white turning dark brown.
Basidiospores (n = 20) 7.1–8.4–9.6 × 6.7–7.9–9.2 µm, Q = 1.01–1.05–1.09, globose to subglobose; ornamentation amyloid, composed of ridges up to 1.7 µm high, forming complete reticulum, occasionally isolated warts present; plage totally amyloid. Basidia 49.5–60 × 9–12.5 µm, clavate to subclavate, 4-spored, filled with granular contents. Pleurolamprocystidia 41–108 × 4–9.5 µm, cell wall 1.5–3 µm, subcylindrical to subfusiform, with acuminate apex, slightly to distinctly emergent, arising from the subhymenium, very abundant. Lamellae edge composed of basidia, cheilocystidia, and marginal cells; cheilocystidia 17–70.5 × 2.5–7 µm, mostly thick-walled (0.5–2 µm), subcylindrical to subfusiform, with acuminate apex; marginal cells 9.5–36.5 × 3–13 µm, subcylindrical to subclavate, 1–3 septate. Pseudocystidia 2–4 µm in diameter, slightly emergent, cylindrical to slightly tortuose. Pileipellis a lampropalisade; terminal elements 20–68 × 2.5–4 µm, cell wall 0.5–1.5 µm thick, erect, subcylindrical to cylindrical with obtuse apex; subpellis 65–100 µm thick, composed of rounded to elongated cells, thick-walled, 5–22 µm in diameter.
Habitat: Scattered on soil in mixed forests dominated by Quercus.
Comments: Lactifluus rugiformis can be recognized by the overall reddish-brown colors, the strongly rugulose pileus margin, the cream to pale orange lamellae, and the short stipe relative to the pileus size. The ratio of stipe length/pileus diameter is between 0.4 and 0.7. This species is phylogenetically closely reated to Lf. longipilus from Thailand (). However, the terminal elements in the pileipellis (10–130 µm) of Lf. longipilus are longer than Lf. rugiformis [Citation10].
4.10. Key to the species of Lactifluus subgenus Lactifluus section Piperati
- Firm and whitish pileus; hyphoepithelium type pileipellis; presence of dermatocystidia in the pileipellis
1. Latex white, turning grayish green2
1. Latex white, unchanging3
2. Lamellae rather distant, suprapellis 30–70 µm thickLf. subviridilacteus
2. Lamellae very crowded, suprapellis 50–130 µm thickLf. viridilacteus
3. Associated with Pinus treesLf. undulatus
3. Associated with Quercus trees4
4. Lamellar edge sterile, marginal cells up to about 15 µm longLf. subquercicola
4. Lamellar edge with basidia, marginal cells up to about 25 µm long5
5. Spore ornamentation dense, almost completely reticulateLf. curvativus
5. Spore ornamentation loose, incompletely reticulateLf. quercicola
4.11. Lactifluus subgenus Lactifluus section Piperati
Lactifluus curvativus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832074
Etymology: “curvativus” means slightly curved margin. This name refers to the in-rolled to decurved margin of this species.
Holotypus: SOUTH KOREA: Gyeonggi-do, Goyang-si, Seo-o-reung royal tombs, alt. 62 m, N37°37′48″ E126°53′54″, 26 Jul 2016, SFC20160726-95 (holotypus, SFC)
Pileus 50–120 mm, at first convex with depressed center; margin entire, decurved, often crenate in age; surface smooth, dry, concentrically rugulose toward margin, whitish with ochraceous spots. Lamellae subdecurrent, very crowded, up to 1.5 mm broad, often furcate, white to pale cream, sometimes tinged with a pale ocherous. Stipe 45–85 × 20–35 mm, subcylindric, often tapering downwards, smooth, often irregularly compressed, dry, white tinged sometimes with ochraceous tinges. Latex rather abundant, white, unchanging.
Basidiospores (n = 40) (5.4–)5.5–6.1–6.3–6.8 × 5.1–5.4–5.5–5.9 µm, Q = 1.03–1.11–1.21, globose to broadly ellipsoid; ornamentation amyloid, up to 0.2 µm high, consisting of irregularly rounded to elongate warts connected by lower lines, forming almost complete reticulum; plage inamyloid. Basidia 36–49.5 × 7.5–9 µm, subcylindric to subclavate, 4-spored, mostly filled with guttulate and granular contents. Pleuromacrocystidia abundant, 38–74 × 4.5–6.5 µm, subcylindrical to narrowly subfusiform, with obtuse apex. Lamellae edge composed of basidia, cheilomacrocystidia, and marginal cells; cheilomacrocystidia 18.5–43 × 4–7 µm, subcylindrical, with obtuse apex; marginal cells 12–28 × 3–6 µm, cylindrical to subclavate, 1–2 septate. Pseudocystidia 2–4 µm in diam., slightly emergent, cylindrical to subcylindrical. Pileipellis a hyphoepithelium; suprapellis 30–50 µm thick, consisting of thin, hyaline hyphae, 2–4 µm broad; subpellis almost completely cellular, consisting of globose to subglobose cells, 5–22 µm in diam.; dermatocystidia rather abundant in suprapellis, 3.5–6.5 µm broad, subcylindrical to cylindrical.
Habitat: Solitary or scattered on soil in the Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Gyeonggi-do, Incheon-si, Mt. Mani, alt. 326 m, N37°36′42″ E126°26′05″, 18 Aug 2015, SFC20150818-15 (SFC).
Comments: Lactifluus curvativus is characterized by white unchanging latex, very crowded lamellae, and small basidiospores. Lactifluus dwaliensis distributed in India is phylogenetically closely related to Lf. curvativus (), however, Lf. dwaliensis can be distinguished from Lf. curvativus by its big basidiospores (8–10.5 × 7.2–8.6 µm) and sterile lamellar edge [Citation60].
Lactifluus quercicola H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832075
Etymology: “querci” means oak tree and “cola” means dweller. The name refers to the presence of the species in Quercus forests and the putative ectomycorrhizal association with Quercus.
Holotypus: SOUTH KOREA: Gyeongsangbuk-do, Sangju-si, Mt. Noeum, alt. 163 m, N36°25′35″ E128°06′24″, 19 Jul 2013, SFC20130719-29 (holotypus, SFC)
Pileus 40–150 mm, at first convex with depressed center; margin decurved, crenulate or wavy in age; surface smooth, dry, matt or slightly shiny, whitish, sometimes with buff-colored spots. Lamellae adnate to decurrent, very crowded, up to 1.5 mm broad, often furcate, never multi-furcate, whitish to pale cream tinged with buff. Stipe 40–95 × 12–30 mm, cylindrical, often tapering downwards, smooth or uneven, dry, white tinged with pale cream, becoming buff or brownish from base. Latex rather abundant, white, unchanging.
Basidiospores (n = 60) 4.7–5.5–5.7–6.4 × (4.1–)4.2–4.9–5.0–5.6 µm, Q = 1.04–1.11–1.12–1.22, globose to broadly ellipsoid; ornamentation amyloid, up to 0.3 µm high, consisting of irregularly rounded to elongate warts connected by low ridges, forming an incomplete reticulum; plage centrally amyloid. Basidia 29.5–42 × 5–9 µm, subcylindrical to subclavate, 2- or 4-spored, filled with granular contents. Pleuromacrocystidia abundant, 35.5–60 × 6.5–8 µm, subcylindrical to subclavate, sometimes with wavy out-line, with obtuse apex. Lamellae edge composed of basidia, cheilomacocystidia, and marginal cells; cheilomacrocystidia 24–40 × 4.5–7 µm, subcylindrical to cylindrical, with obtuse apex; marginal cells 13–35.5 × 4–5.5 µm, cylindrical to subcylindrical, 1–2 septate. Pseudocystidia rare, 2–4 µm in diam., not emergent, cylindrical to slightly tortuose. Pileipellis a hyphoepithelium; suprapellis 20–50 µm thick, consisting of thin, hyaline hyphae, 2–3.5 µm broad; subpellis almost completely cellular, consisting of globose to subglobose cells, 6–34 µm in diam.; dermatocystidia rather abundant in suprapellis, 4.5–7 µm broad, subcylindrical to cylindrical.
Habitat: Solitary or gregarious on soil in Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Gyeongsangbuk-do, Yeongcheon-si, Mt. Bohyeon, alt. 695 m, N36°10′30″ E128°59′07″, 29 Jul 2013, TPML130729-023 (TPML); ibid., Yeongdeok-gun, Mt. Chilbo, alt. 360 m, N36°37′26″ E128°21′60″, 29 Jul 2013, TPML130729-035 (TPML).
Comments: Lactifluus quercicola is phylogenetically closely related to Lf. piperatus (). Lactifluus piperatus is distinguished from Lf. quercicola by bigger basidiospore size (7.0–10.4 × 5.2–7.5) [Citation42]. Smaller marginal cells of Lf. quercicola (13–35.5 × 4–5.5 µm) and its association with Quercus are distinguishing freatures separating it from morphologically similar species, Lf. subquercicola (5–15 × 2–5.5 µm) and Lf. undulatus (association with Pine), respectively.
Lactifluus subquercicola H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832076
Etymology: ‘sub’ means almost and ‘quercicola’ refers Lactifluus quercicola. This name refers to the fact that this species is reminiscent of Lf. quercicola.
Holotypus: SOUTH KOREA: Gangwon-do, Inje-gun, Mt. Jeombong, alt. 864 m, N38°04′06″ E128°23′43″, 30 Jul 2012, TPML120730-006 (holotypus, TPML; isotypus, SFC)
Pileus 50–120 mm, planoconvex with decurved margin when young, with age expanding and becoming depressed in the center; margin crenulate and wavy in age; surface smooth, dry, slightly shiny, whitish, sometimes with brownish-orange-colored spots. Lamellae adnate to decurrent, very crowded, up to 2 mm broad, furcate, often multi-furcate, sometimes furcate toward stipe, pale cream to cream tinged with pale orange. Stipe 50–110 × 10–30 mm, cylindrical, often tapering downwards, smooth, dry, white tinged with whitish chrome or pale cream, becoming buff or brownish from base. Latex rather abundant, white, unchanging.
Basidiospores (n = 60) 5.2–6.2–6.3–7.2(–7.4) × 4.5–5.2–5.4–6.3(–6.4) µm, Q = 1.03–1.11–1.14–1.22, globose to broadly ellipsoid; ornamentation amyloid, up to 0.2 µm high, consisting of irregularly rounded warts connected by fine lines, forming an incomplete reticulum; plage inamyloid. Basidia 33–45 × 6.5–10.5 µm, subcylindric to subclavate, 4-spored, filled with granular contents. Pleuromacrocystidia abundant, 49.5–63.5 × 7–9.5 µm, subclavate to subfusiform, with obtuse apex. Lamellae edge sterile, composed of cheilomacrocystidia and marginal cells; cheilomacrocystidia 17.5–45 × 5.5–7.5 µm, subcylindrical, with obtuse apex; marginal cells 5–15 × 2–5.5 µm, subclavate to clavate, 1–2 septate. Pseudocystidia rare 1.5–3.5 µm in diam., not emergent, cylindrical to slightly tortuous, often branching. Pileipellis a hyphoepithelium; suprapellis 20–60 µm thick, consisting of thin, hyaline hyphae, 1.5–3.5 µm broad; subpellis cellular, consisting of globose to subglobose cells, 5–27 µm in diameter; dermatocystidia rather abundant in suprapellis, 4–6.5 µm broad, subcylindrical to cylindrical.
Habitat: Solitary or gregarious on soil in Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Gangwon-do, Goseong-gun, Mt. Hyangrobong, alt. 516 m, N38°18′51″ E128°20′35″, 18 Aug 2011, TPML110818-058 (TPML); Jeollabuk-do, Muju-gun, Mt. Deogyu, alt. 932 m, N35°51′31″ E127°46′06″, 17 Aug 2002, HCCN10587 (HCCN).
Comments: Lactifluus subquercicola has similar macromorphology with Lf. quercicola and Lf. undulatus, and species may be easily misidentified in the field. However, Lf. subquercicola can be distinguished from these two species based on the sterile lamellar edge.
Lactifluus subviridilacteus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832077
Etymology: ‘sub’ means almost and ‘viridilacteus’ refers Lactifluus viridilacteus. This name refers to the fact that this species is reminiscent of Lf. viridilacteus.
Holotypus: SOUTH KOREA: Chungcheongnam-do, Buyeo-gun, Muryangsa Temple, alt. 163 m, N36°19′02″ E126°41′36″, 28 Aug 2014, SFC20140828-19 (holotypus, SFC)
Pileus 50–120 mm, convex to applanate with a depressed center; margin decurved; surface smooth, dry, slightly velutinous, rugulose, white to pale cream often with irregular brownish spots. Lamellae slightly decurrent, up to 8 mm broad, rather distant, never furcate, with 1–2 series of lamellulae between lamellae, whitish to pale cream tinged often with yellowish brown. Stipe 40–85 × 10–30 mm, cylindrical; surface smooth, dry, white to pale cream tinged often with yellowish brown. Latex rather abundant, white, turning grayish green.
Basidiospores (n = 40) 6.5–7.5–7.6–8.6(–8.7) × 5.5–6.4–6.6–7.4(–7.7) µm, Q = 1.11–1.17–1.24, subglobose to broadly ellipsoid; ornamentation amyloid, up to 0.2 µm high, composed of irregular isolated warts, connected by low lines, forming an incomplete reticulum; plage mostly inamyloid, sometimes slightly amyloid at the center. Basidia 49.5–56 × 7.5–11 µm, subcylindric to subclavate, 4-spored, filled with granular contents. Pleuromacrocystidia abundant 71.5–113.5 × 7.5–10.5 µm, subcylindrical to subfusiform, with obtuse or inflated and rounded apex, originating deep in the trama, mostly emergent. Lamella edge composed of basidia, strongly emergent cheilomacrocystidia, and marginal cells; cheilomacrocystidia 39.5–80.5 × 6–10 µm, subclavate, with obtuse apex; marginal cells 15–32 × 3–6 µm, cylindrical to subclavate, 1–2 septate. Pseudocystidia 3.5–5 µm in diam., slightly emergent, cylindrical to slightly irregular. Pileipellis a hyphoepithelium; suprapellis 30–70 µm thick, consisting of thin, hyaline hyphae, 2.5–4 µm broad; subpellis cellular, consisting of globose to subglobose cells, 9–28 µm in diameter; dermatocystidia very abundant in suprapellis, 6–10 µm broad, subcylindrical to subclavate.
Habitat: Solitary or gregarious on soil in Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Jeollabuk-do, Jangsu-gun, Bangwhadong Natural Recreation Forest, alt. 477 m, N35°35′48″ E127°31′45″, 27 Aug 2014, SFC20140827-34 (SFC).
Comments: Lactifluus subviridilacteus is characterized by white latex with grayish-green discoloration, rather distant and non-furcate lamellae. This species is sometimes confused with Lf. subpiperatus by relatively distant lamellae. However, they can be distinguished from each other as the former has pleuromacrocystidia, while the latter does not [Citation43]. In addition, Lf. subpiperatus has smaller basidia (34–40 × 7–8 µm) and cheilomacrocystidia (20–34 × 3–6 µm) than Lf. subviridilacteus [Citation43].
Lactifluus undulatus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832078
Etymology: ‘undulatus’ means wavy. This name refers to the wavy margin of this species.
Holotypus: SOUTH KOREA: Gyeongsangbuk-do, Yeongdeok-gun, Mt. Chilbo, alt. 435 m, N36°37′27″ E129°21′50″, 29 Jul 2013, TPML130729-041 (holotypus, TPML; isotypus, SFC)
Pileus 60–130 mm, convex with slightly depressed center and decurved margin when young, later expanding and infundibuliform; margin uplifted and wavy in age; surface smooth, dry, slightly shiny, whitish, sometimes with ocher colored spots. Lamellae adnate, up to 3 mm broad, very crowded, often furcate, sometimes multi-furcate, whitish to pale cream tinged often with pale cream. Stipe 45–90 × 10–20 mm, cylindrical, often tapering downwards; surface smooth, dry, white tinged often with pale cream, pale ocher at the base. Latex rather abundant, white, unchanging.
Basidiospores (n = 40) 5.1–5.7–5.9–6.4 × 4.6–5.2–5.3–5.9 µm, Q = 1.03–1.09–1.11–1.15, globose to subglobose; ornamentation amyloid, up to 0.2 µm high, composed of irregular isolated warts, occasionally connected by low lines, forming an incomplete reticulum; plage inamyloid. Basidia 28.5–35.5 × 5.5–8 µm, subcylindrical, 4-spored, mostly filled with guttulate and granular contents. Pleuromacrocystidia abundant, 41–77.5 × 6–8.5 µm, subcylindrical to subclavate, with obtuse or rounded apex, sometimes with irregularly wavy outline, originating deep in the trama. Lamella edge composed of basidia, cheilomacrocystidia, and marginal cells; cheilomacrocystidia 24–47 × 5.5–8 µm, subcylindric to subclavate, with obtuse or rounded apex; marginal cells 8–21 × 3–6 µm, cylindrical to subcylindrical, 1–2 septate. Pseudocystidia rare, 2.5–3.5 µm in diameter, not emergent, cylindrical to subcylindrical. Pileipellis a hyphoepithelium; suprapellis 20–50 µm thick, consisting of thin, hyaline hyphae, 2–3.5 µm broad; subpellis almost completely cellular, consisting of globose to subglobose cells, 7.5–40 µm in diam.; dermatocystidia scarce in suprapellis, 3.5–6.5 µm broad, subcylindrical to cylindrical.
Habitat: Solitary or scattered on soil in Pinus forests.
Additional studied material: SOUTH KOREA: Chungcheongbuk-do, Boeun-gun, Mt. Songni, alt. 454 m, N36°32′37″ E127°50′22″, 3 Aug 2011, TPML110803-086 (TPML).
Comments: Lactifluus undulatus is macromorphologically very similar to Lf. quercicola and Lf. subquercicola. However, their habitats are different. While Lf. quercicola and Lf. subquercicola grow in oak forests, Lf. undulatus grows in pine forests.
Lactifluus viridilacteus H. Lee & Y. W. Lim sp. nov. ()
MycoBank: MB 832079
Etymology: “viridi” means greenish and “lacteus” means milky. The name refers to the grayish-green latex discoloration of this species.
Holotypus: SOUTH KOREA: Gyeonggi-do, Gwangju-si, Mt. Taewha, alt. 285 m, N37°18′29″ E127°18′01″, 19 Aug 2015, SFC20150819-08 (holotypus, SFC)
Pileus 50–150 mm, convex to applanate with a depressed center; surface smooth, dry, slightly velutinous, sometimes slightly rugose, white to pale cream with darker spots. Lamellae decurrent, very crowded, up to 2 mm broad, whitish, turning greenish by the latex and finally becoming brownish. Stipe 30–90 × 10–40 mm; surface smooth, dry, white to pale cream. Latex rather abundant, white becoming grayish green.
Basidiospores (n = 40) 5.2–5.9–6.0–6.6 × 4.5–5.1–5.2–5.7(–5.8) µm, Q = 1.07–1.15–1.18–1.27, subglobose to broadly ellipsoid; ornamentation amyloid, up to 0.3 µm high, composed of irregular isolated warts, aligned or connected by lower lines, but never forming a reticulum; plage inamyloid, sometimes with a slightly amyloid spot. Basidia 38–45 × 7.5–9 µm, subclavate, 4-spored, filled with guttulate and granular contents. Pleuromacrocystidia abundant, 45–80 × 7.5–10.5 µm, subcylindrical to subclavate, originating deep in the trama, mostly strongly emergent; apex obtuse. Lamella edge composed of basidia, strongly emergent cheilomacrocystidia, and marginal cells; cheilomacrocystidia abundant, 32.5–71.5 × 7–9.5 µm, cylindric to subclavate, with obtuse apex; marginal cells 6–20 × 2–4.5 µm, subcylindrical to clavate, 1–3 septate. Pseudocystidia 2–3.5 µm in diam., emergent, cylindrical to slightly tortuose, often branching. Pileipellis a hyphoepithelium; suprapellis 50–130 µm thick, consisting of thin, hyaline hyphae, 1.5–4 µm broad; subpellis cellular, consisting of globose to subglobose cells, 6–23.5 µm in diameter; dermatocystidia rather abundant in suprapellis, 4.5–6.5 µm broad, subcylindrical to cylindrical.
Habitat: Solitary or gregarious on soil in the Quercus forests or mixed forests dominated by Quercus.
Additional studied material: SOUTH KOREA: Gyeongsangbuk-do, Sangju-si, Mt. Noem, alt. 203 m, N36°26′39″ E128°06′50″, 8 Aug 2013, SFC20130808-37 (SFC); Jeollanam-do, Haenam-gun, Mt. Duryun, alt. 364 m, N34°28′59″ E126°37′30″, 1 Aug 2005, HCCN12866 (HCCN).
Comments: Lactifluus viridilacteus and Lf. subviridilacteus are the two Korean species in this section with greenish discoloring latex. Lactifluus viridilacteus furthermore characterized by the very crowded lamellae. Lactifluus glaucescens, distributed in Europe, is phylogenetically closely related to this species (). However, the basidiospores of Lf. glaucescens (6.5–9.3 × 5.3–6.9) are larger than those of Lf. viridilacteus (38–45 × 7.5–9 µm) [Citation42].
5. Discussion
Through a comprehensive morphological and multilocus phylogenetic analysis of 199 Lactifluus specimens collected over 37 years, a total of 24 Lactifluus species were identified. Our study shows that Lactifluus is not only well distributed in tropical ecosystems and the Southern Hemisphere, but can also be diverse in temperate forest in the Northern Hemisphere. Surprisingly, only five species were previously described species, while at least 18 others were confirmed as new species (one putatively new species requires additional data). The five previously recognized species were all described from Asia: two species (Lf. acicularis and Lf. pilosus) were originally reported from Thailand [Citation8,Citation10] and three (Lf. holophyllus, Lf. luteolamellatus and Lf. pseudohygrophoroides) were described from South Korea [Citation23]. As expected, many specimens were previously misidentified, being assigned to morphologically similar species from Europe and North America. Our results strongly support that most Asian Lactifluus species are not conspecific with morphologically similar taxa from other continents [Citation5,Citation8]. Such was also the case for Lactarius species in Korea [Citation47]. Therefore, European and North American species are not likely to be distributed in Asia, and most of the recorded species based on morphological surveys in Korea (and other parts of Asia) are likely incorrect. For example, Lf. piperatus, a European species was reported in Korea without detailed morphological descriptions [Citation61], and in our study, sequence data of a European specimen (voucher 2001 08 19 68, France) did not match data from any Korean specimens (). Specimens labeled as Lf. piperatus in Korea have been identified as one of three new species: Lf. quercicola, Lf. subquercicola, and Lf. undulatus.
Previously, 21 Lactifluus species were reported in Korea [Citation22–24,Citation62], yet only five were found in this study. Of the 16 remaining recorded species, only three of them were originally reported from Asia, while seven were described from Europe, and six from North America. The three Asian species recorded in Korea are Lf. ochrogalactus (Hashiya) X.H. Wang, Lf. pinguis (Van de Putte & Verbeken) Van de Putte, and Lf. subpiperatus Hongo. Lactifluus ochrogalactus and Lf. pinguis were recently reported in Korea [Citation24,Citation62], while the specimens labeled as Lf. subpiperatus turn out to be a new species in this study, Lf. subviridilacteus. Although we did not identify Lf. subpiperatus in our study, the existence of this species in Korea cannot be ruled out as it is present in Japan.
For infrageneric classification of Korean Lactifluus, color and color change of latex, pileipellis type, and cystidium type are useful characters. Lactifluus species show various latex discoloration patterns when exposed to air, and this informative to distinguish some groups. The morphological characters of the new species discovered in Korea correspond well with the species in the known clades of Lactifluus [Citation5]. Korean species belonging to Lf. sect. Lactifluus and Lf. sect. Gymnocarpi have brownish discoloration of latex without exception. On the other hand, some groups have diverse patterns of latex discoloration. Most species in the Lf. sect. Piperati have greenish discoloration of latex, which is true for two Korean species, but the other four show no discoloration. Korean species of Lf. sect. Pseudogymnocarpi exhibit mixed patterns—two species with brownish discoloration of white latex and one species with unchanging white latex.
In previous studies, the type of pileipellis was noted as an important character to delimit sections or subgenera within Lactifluus [Citation7,Citation44]. Lactifluus species in sect. Lactifluus and sect. Pseudogymnocarpi share the same type of pileipellis. However, various pileipellis types are observed in species of sect. Albati and sect. Gerardii. For species native to Korea, we found only one type of pileipellis for each section; lamprotrichoderm type of sect. Albati and palisade type of sect. Gerardii. The type of cystidia and presence/absence of cystidia is also useful to delimit some sections such as sect. Lactifluus (pleurolamprocyatidia) and sect. Luteoli (no true cystidia). The presence of only one type of cystidia for each section of Korean species highlights its informativeness for identification. Further work is needed to determine if cystidia characters are useful for species identification in other parts of the world. The characters of the context (color, dicolouration, smell, and taste) are largely unknown in the Korean species, although in some groups they might reveal additional relevant differences. They certainly need more attention in future.
In addition to the three characters mentioned above (latex color and color change, pileipellis type, cystidia type), the shape of basidiomata also is an important character of Lactifluus species. All species of Lactifluus investigated in this study are agaricoid. So far, pleurotoid Lactifluus species are known in three Lactifluus subgenera: Lactariopsis, Gymnocarpi, and Lactifluus. While pleurotoid species are reported from all continents except Europe [Citation11,Citation41,Citation63–68], they have thus far not been observed in Korea. Previous studies show that the pleurotoid basidiomata has multiple evolutionary origins [Citation65,Citation66]. As pluerotoid species are reported from Japan and China [Citation11,Citation69], whether they exist in Korea requires further investigation.
Ecological information of ecotomycorrhizal fungi can be useful for their identification and forest management. Sequence data in GenBank may include information on species distribution and host. From the same sequence data with the new species found in this study, we could confirm that many new species are also distributed in other countries of Asia: Lf. orientivellereus from Japan [Citation70], Lf. betulicola from Japan [Citation9], Lf. longistipes from China and Far East Russia (GenBank), Lf. quercicola from China (GenBank) and Japan [Citation71], Lf. subquercicola from China and Japan (GenBank), and Lf. undulata from Thailand and Vietnam [Citation42]. Hosts that interacted with these species were similar to those studied in Korea (). An ITS sequence (AB568438) consistent with Lf. quercicola was obtained from the root of orchid Cephalanthera longibracteata, Japan [Citation72]. Thus, Lf. quercicola may be one of the mixotrophic hosts of Cephalanthera longibracteata.
In conclusion, we confirm the presence of 24 Lactifluus species in Korea, including at least 18 species new to science, based on morphological and multilocus phylogenetic approaches. Our results show that identification of Lactifluus species using morphological data alone can be unreliable due to considerable overlap of characters among species. In addition to morphological observation of defining characters, a molecular approach is necessary for infrageneric classification and species identification in Korean Lactifluus. Since many specimens used in this study were missing information regarding host plants, the ecological relevance of all Korean Lactifluus species could not be examined. Further investigation is necessary to better understand the general ecology of Lactifluus species.
Acknowledgements
The authors thank Dr. Soon Ja Seok, Prof. Jong Kyu Lee, and Dr. Sang Kuk Han for providing specimens. We also greatly appreciate Dr. Xiang-Hua Wang to improve our manuscript through detailed comments.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Figure 9. Microscopic features of Lactifluus orientivellereus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
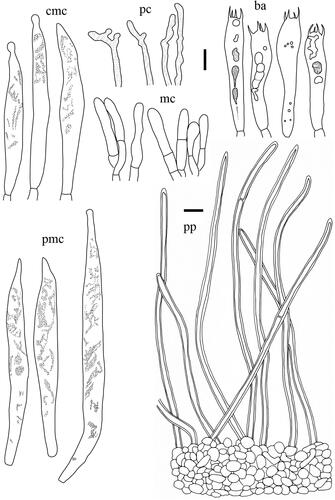
Figure 10. Microscopic features of Lactifluus betulicola sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
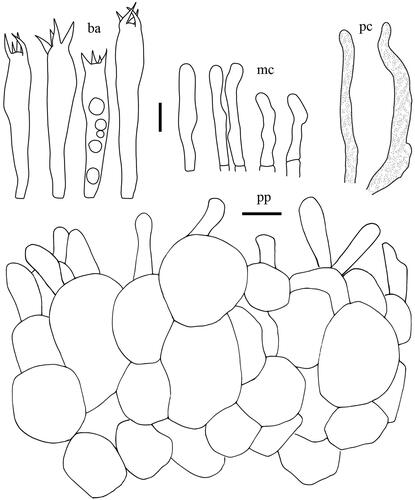
Figure 11. Microscopic features of Lactifluus luminosus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
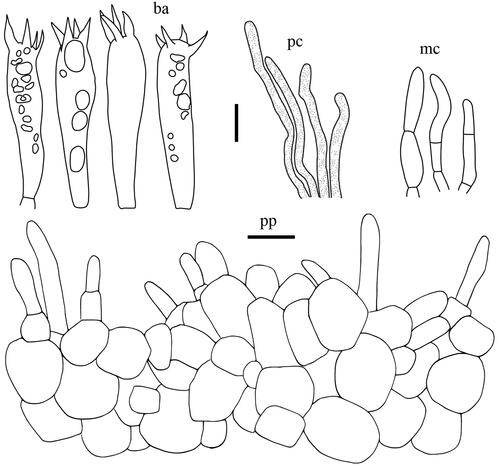
Figure 12. Microscopic features of Lactifluus stellatus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
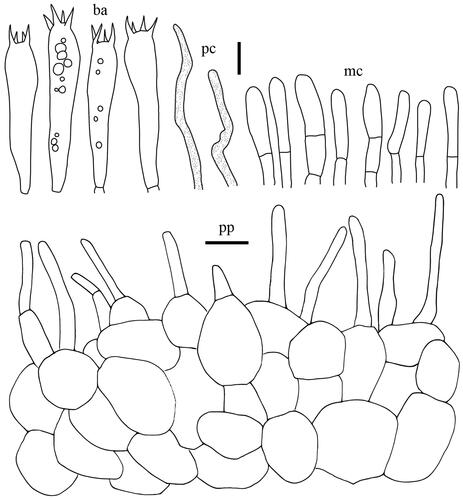
Figure 13. Microscopic features of Lactifluus koreanus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
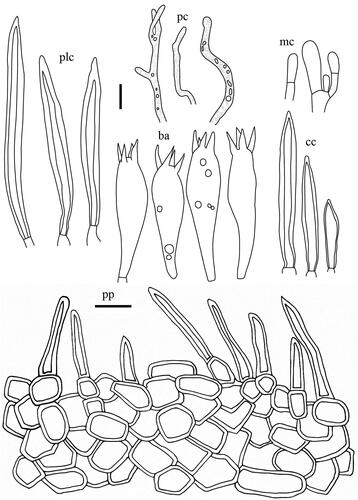
Figure 14. Microscopic features of Lactifluus longistipes sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
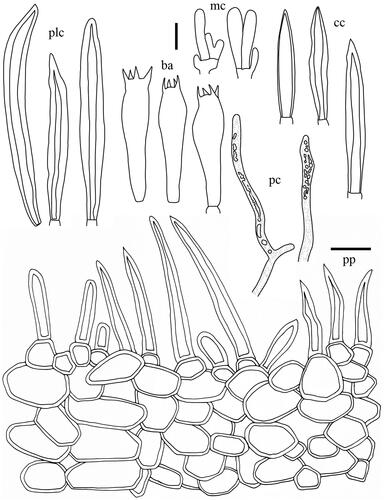
Figure 15. Microscopic features of Lactifluus orientivolemus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
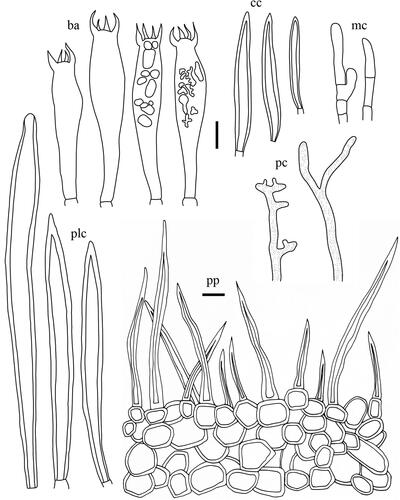
Figure 16. Microscopic features of Lactifluus porphyreus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
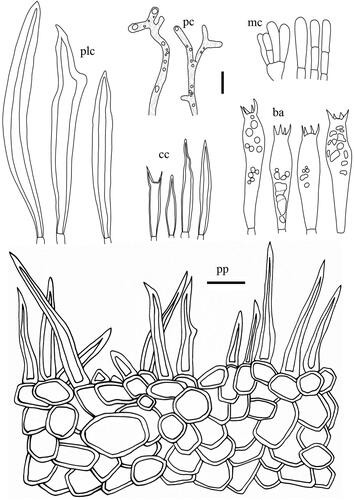
Figure 17. Microscopic features of Lactifluus rugiformis sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
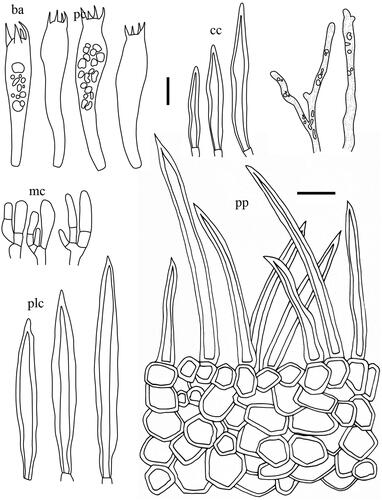
Figure 18. Microscopic features of Lactifluus curvativus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
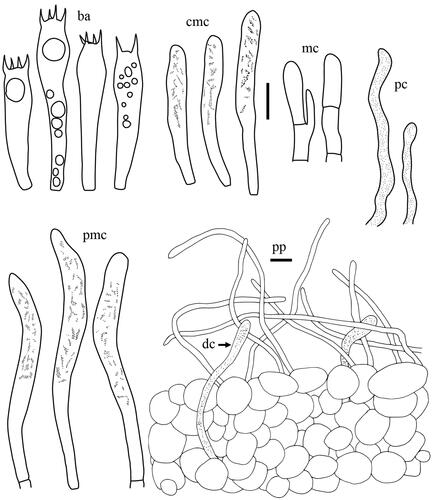
Figure 19. Microscopic features of Lactifluus quercicola sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
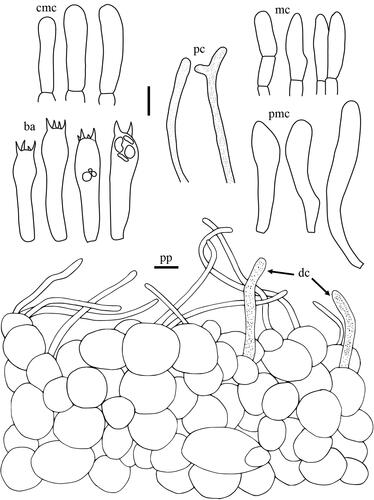
Figure 20. Microscopic features of Lactifluus subquercicola sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
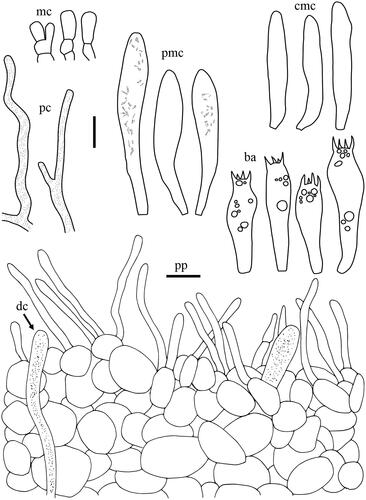
Figure 21. Microscopic features of Lactifluus subviridilacteus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
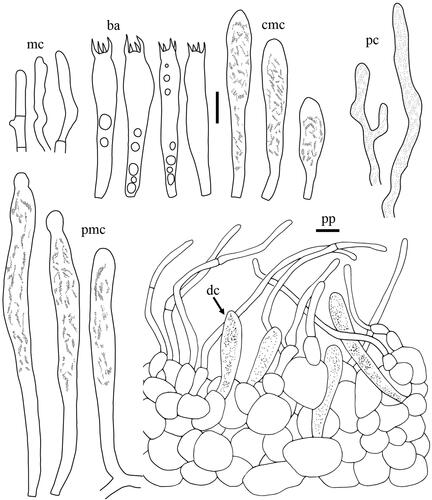
Figure 22. Microscopic features of Lactifluus undulatus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
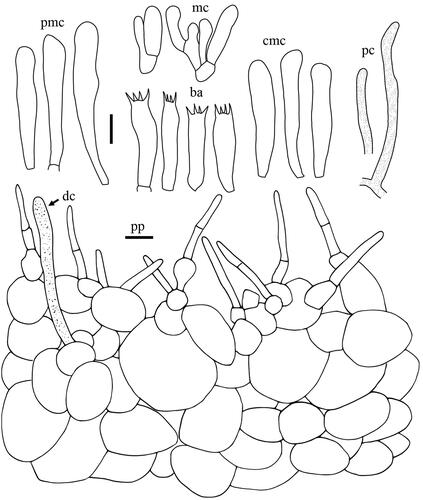
Figure 23. Microscopic features of Lactifluus viridilacteus sp. nov. All scale bars = 10 µm. Horizontal scale bars are for pileipellis and vertical scale bars are for other microscopic features.
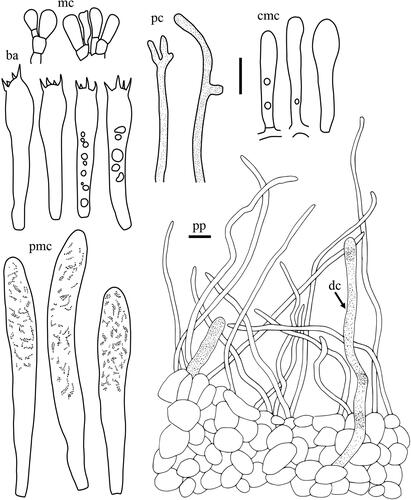
Figure 24. Fruiting bodies of Korean species of Lactifluus subg. Pseudogymnocarpi (a∼c), subg. Lactariopsis (d∼f), subg. Gymnocarpi (g∼h). (a) Lf. holophyllus, (b) Lf. luteolamellatus, (c) Lf. pseudohygrophoroides, (d) Lf. multiseparatus, (e) Lf. orientivellereus, (f) Lf. pilosus, (g) Lf. albidopectinatus, (h) Lf. pallidotestaceus. The color of the two circles at the top-right of each species picture indicates the color (left circle) and color change (right circle) of the latex. Scale bar = 1 cm.

Additional information
Funding
References
- Buyck B, Hofstetter V, Eberhardt U, et al. Walking the thin line between Russula and Lactarius: the dilemma of Russula subsect. Ochricompactae. Fungal Divers. 2008;28:15–40.
- Buyck B, Hofstetter V, Verbeken A, et al. Proposal 1919: To conserve Lactarius nom. Cons. (Basidiomycota) with avconserved type. Taxon. 2010;59(1):295–296.
- McNeill J, Turland NJ, Monro AM, et al. XVIII International Botanical Congress: Preliminary mail vote and report of Congress action on nomenclature proposals. Taxon. 2011;60(5):1507–1520.
- Verbeken A, Nuytinck J. Not every milkcap is a Lactarius. Scr Bot Bel. 2013;51:162–168.
- De Crop E, Nuytinck J, Van de Putte K, et al. A multi-gene phylogeny of Lactifluus (Basidiomycota, Russulales) translated into a new infrageneric classification of the genus. Persoonia. 2017;38:58–80.
- De Crop E, Delgat L, Nuytinck J, et al. A short story of nearly everything in Lactifluus (Russulaceae). Fungal Syst Evol. 2021;7(1):133–164.
- Verbeken A, Walleyn R. Monograph of Lactarius in tropical Africa. Fungus Flora of Tropical Africa. National Botanic Garden, Belgium. 2010;2.
- Le HT, Verbeken A, Nuytinck J, et al. Lactarius in Northern Thailand: 3. Lactarius Subgenus Lactoriopsis. Mycotaxon. 2007;102:281–291.
- Stubbe D, Nuytinck J, Verbeken A. Critical assessment of the Lactarius gerardii species complex (Russulales). Fungal Biol. 2010;114(2-3):271–283.
- Van de Putte K, Nuytinck J, Stubbe D, et al. Lactarius volemus sensu lato (Russulales) from northern Thailand: morphological and phylogenetic species concepts explored. Fungal Divers. 2010;45(1):99–130.
- Wang XH, Stubbe D, Verbeken A. Lactifluus parvigerardii sp. nov., a new link towards the pleurotoid habit in Lactifluus subgen. Gerardii (Russulaceae, Russulales). Cryptogam Mycol. 2012;33(2):181–190.
- Wang X-H, Buyck B, Verbeken A, et al. Revisiting the morphology and phylogeny of Lactifluus with three new lineages from southern China. Mycologia. 2015;107(5):941–958.
- Henkel TW, Aime MC, Miller SL. Systematics of pleurotoid Russulaceae from Guyana and Japan, with notes on their ectomycorrhizal status. Mycologia. 2000;92(6):1119–1132.
- Miller SL, Aime MC, Henkel TW. Russulaceae of the Pakaraima Mountains of Guyana. I. New species of pleurotoid Lactarius. Mycologia. 2002;94(3):545–553.
- Smith ME, Henkel TW, Aime MC, et al. Ectomycorrhizal fungal diversity and community structure on three co-occurring leguminous canopy tree species in a Neotropical rainforest. New Phytol. 2011;192(3):699–712.
- Sá MCA, Baseia IG, Wartchow F. Lactifluus dunensis, a new species from Rio Grande do Norte. Brazil. Mycosphere. 2013;4(2):261–265.
- Sá MCA, Wartchow F. Lactifluus aurantiorugosus (Russulaceae), a new species from Southern Brazil. Darwiniana, Nueva Serie. 2013;1(1):54–60.
- Crous PW, Wingfield MJ, Burgess TI, et al. Fungal Planet description sheets: 625-715. Persoonia. 2017;39(715):270–467.
- Sá MCA, Delgat L, Verbeken A, et al. A new species of Lactifluus (Russulales, Agaricomycetes) from the Brazilian caatinga semiarid region. N Z J Botan. 2019;57(3):169–110.
- Delgat L, Dierickx G, De Wilde S, et al. Looks can be deceiving: the deceptive milkcaps (Lactifluus, Russulaceae) exhibit low morphological variance but harbour high genetic diversity. IMA Fungus. 2019;10(1):14.
- Bok JD, Shin GC. Taxonomic Studies on the Genus Lactarius of Korea (I). Kor J Mycol. 1985;13(4):249–262.
- Lee YS, Lim YW, Kim JJ, et al. 2015. National list of species of Korea: Basidiomycota. National Institute of Biological Resources, Korea. 364 pp.
- Hyde KD, Norphanphoun C, Abreu VP, et al. Fungal divers notes 603–708: taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017;87(1):1–235.
- Lee H, Park JY, Wisitrassameewong K, et al. First report of eight milkcap species belonging to Lactarius and Lactifluus in Korea. Mycobiology. 2018;46(1):1–12.
- Van de Putte K, Nuytinck J, Das K, et al. Exposing hidden diversity by concordant genealogies and morphology - a study of the Lactifluus volemus (Russulales) species complex in Sikkim Himalaya (India). Fungal Divers. 2012;55(1):171–194.
- Van de Putte K, Nuytinck J, De Crop E, et al. Lactifluus volemus in Europe: three species in one-Revealed by a multilocus genealogical approach, Bayesian species delimitation and morphology. Fungal Biol. 2016;120(1):1–25.
- Boa E. 2004. Wild Edible Fungi. A Global Overview of Their Use and Importance to People. Non-Wood Forest Products 17. FAO, Rome.
- Colak A, Faiz O, Sesli E. Nutritional composition of some wild edible mushrooms. Turk J Biochem. 2009;34(1):25–31.
- Garibay-Orijel R, Cifuentes J, Estrada-Toress A, et al. People using macro-fungal diversity in Oaxaca. Mexico. Fungal Divers. 2006;21:41–67.
- Garibay-Orijel R, Caballero J, Estrada-Toress A, et al. Understanding cultural significance, the edible mushrooms case. J Ethnobiol Ethnomed. 2007;3:4.
- Christensen M, Bhattarai S, Devkota S, et al. Collection and use of wild edible fungi in Nepal. Econ Bot. 2008;62(1):12–23.
- Dell B, Sanmee R, Lumyong P, et al. 2005. Ectomycorrhizal fungi in dry and wet dipterocarp forests in northern Thailand — diversity and use as food. Asia Pacific Association of Forestry Research Institutions (APAFRI). 8th Round-Table Conference on Dipterocarps. Ho Chi Minh, Vietnam
- Wang XH, Liu P, Yu F. 2004. Color atlas of wild commercial mushrooms in Yunnan (in Chinese). Yunnan Science and Technology Press, Yunnan.
- Daniewski WM, Gumułka M, Przesmycka D, et al. Sesquiterpenes of Lactarius origin, antifeedant structure-activity relationships. Phytochemistry. 1995;38(5):1161–1168.
- Mooibroek H, Cornish K. Alternative sources of natural rubber. Appl Microbiol Biotechnol. 2000;53(4):355–365.
- Tanaka Y. Structural characterization of natural polyisoprenes: solve the mystery of natural rubber based on structural study. Rubber Chemistry and Technology. 2001;74(3):355–375.
- Eberhardt U. Molecular kinship analyses of the agaricoid Russulaceae: correspondence with mycorrhizal anatomy and sporocarp features in the genus Russula. Mycol Prog. 2002;1(2):201–223.
- Miller SL, Buyck B. Molecular phylogeny of the genus Russula in Europe with a comparison of modern infrageneric classifications. Mycol Res. 2002;106(3):259–276.
- Matheny PB, Liu YJ, Ammirati JF, et al. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am J Bot. 2002;89(4):688–698.
- Matheny PB. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol Phylogenet Evol. 2005;35(1):1–20.
- Stubbe D, Le HT, Wang XH, et al. The Australasian species of Lactarius subgenus Gerardii (Russulales). Fungal Divers. 2012;52(1):141–167.
- De Crop E, Nuytinck J, Van de Putte K, et al. Lactifluus piperatus (Russulales, Basidiomycota) and allied species in Western Europe and a preliminary overview of the group worldwide. Mycol Prog. 2014;13(3):493–511.
- Hesler LR, Smith AH. 1979. North American species of Lactarius. University of Michigan Press, Ann Arbor, 841. pp.
- Heilmann-Clausen J, Verbeken A, Vesterholt J. The Genus Lactarius: Fungi of Northern Europe. Svampetryk, Mundelstrup, DK. 1998;2
- Pegler DN, Fiard JP. Taxonomy and ecology of Lactarius (Agaricales) in the Lesser Antilles. Kew Bull . 1979;33(4):601–628.
- Singer R, Araujo I, Ivory MH. The ectotropically mycorrhizal fungi of the neotropical lowlands, especially Central Amazonia. (Litter decomposition and ectomycorrhiza in Amazonian forests 2.) Beihefte zur. Nova Hedwigia. 1983;77:1–352.
- Lee H, Wissitrassameewong K, Park MS, et al. Taxonomic revision of the genus Lactarius (Russulales, Basidiomycota) in Korea. Fungal Divers. 2019;95(1):275–335.
- Kornerup A, Wanscher JH. 1978. Methuen Handbook of colour., 3rd ed. Eyre Methuen Ltd., London.
- Rogers SO, Bendich AJ. 1994. Extraction of total cellular DNA from plants, algae and fungi. In: Gelvin SB, Schilperoort RA (eds), Plant molecular biology manual. Kluwer Academic Publisher, Boston. pp 183–190.
- Rozen S, Skaletsky HJ. Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol. 2000;132:365–386.
- Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729.
- Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30(4):772–780.
- Miller MA, Pfeiffer W, Schwartz T. 2010. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. SC10 Workshop on Gateway Computing Environments (GCE10).
- Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30(9):1312–1313.
- Darriba D, Taboada GL, Doallo R, et al. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods. 2012;9(8):772.
- Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19(12):1572–1574.
- Rambaut A, Suchard M, Xie W, et al. 2014. Tracer v. 1.6. Institute of Evolutionary Biology. Edinburgh. Available from: http://tree.bio.ed.ac.uk/software/tracer/.
- Bon M. Un nouveau lactaire de la section Plinthogali (Burl. Sing. – Lactarius Brunneo-Violascens Bon n. sp. Documents Mycologiques (in French. 1971;1(2):45–48.
- Stubbe D, Verbeken A, Wang XH. New combinations in Lactifluus. 2. Lf. subgenus Gerardii. Mycotaxon. 2012;119(1):483–485.
- Das K, Sharma JR, Verbeken A. New species of Lactarius from Kumaon Himalaya. India. Mycotaxon. 2003;88:333–342.
- Lee YW. Higher fungi of Dagelet Island (forest experiment station). Korean Journal of Botany. 1959;2:22–24.
- Ko PY, Lee HB, Lee SH, et al. New records of fungi growing on Jejudo. Korean Journal of Mycology. 2015;43(3):142–148.
- Verbeken A. New Taxa of Lactarius (Russulaceae) in Tropical Africa. Bull Jard Bot Natl Belg. 1996;65(1/2):197–213.
- Verbeken A, Stubbe D, Nuytinck J. Two new Lactarius species from Cameroon. Cryptogamie Mycologie. 2008;29:137–143.
- Miller SL, Aime MC, Henkel TW. Russulaceae of the Pakaraima Mountains of Guyana 2. New species of Russula and Lactifluus. Mycotaxon. 2013;121(1):233–253.
- Morozova OV, Popov ES, Kovalenko AE. Studies on mycobiota of Vietnam II. Two Species of Lactifluus (Russulaceae) with Pleurotoid Basidiomata. Mikol Fitopatol. 2013;47(2):92–102.
- Latha KPD, Raj KNA, Farook VA, et al. Three new species of Russulaceae from India based on morphology and molecular phylogeny. Phytotaxa. 2016;246(1):61–77.
- De Crop E, Hampe F, Wisitrassameewong K, et al. Novel diversity in Lactifluus section Gerardii from Asia: five new species with pleurotoid or small agaricoid basidiocarps. Mycologia. 2018;110(5):962–984.
- Singer R. Tropical Russulaceae. II. Lactarius section Panuoidei. Nova Hedwigia. 1984;40(1–4):435–447.
- Murata M, Kanetani S, Nara K. Ectomycorrhizal fungal communities in endangered Pinus amamiana forests. PloS One. 2017;12(12):e0189957.
- Murata M, Kinoshita A, Nara K. Revisiting the host effect on ectomycorrhizal fungal communities: implications from host-fungal associations in relict Pseudotsuga japonica forests. Mycorrhiza. 2013;23(8):641–653.
- Sakamoto Y, Yokoyama J, Maki M. Mycorrhizal diversity of the orchid Cephalanthera longibracteata in Japan. Mycoscience. 2015;56(2):183–189.