
ABSTRACT
The plastidic thioredoxin F-type (TrxF) protein plays a key role in plant carbohydrate metabolism. In this work, a gene encoding the TrxF protein, named StTrxF, was successfully isolated from potato and transformed into Arabidopsis thaliana to obtain transgenic plants. Constitutive expression of StTrxF significantly increased the starch accumulation in the transgenic plants. Real-time quantitative polymerase chain reaction analysis showed that constitutive expression of StTrxF up-regulated the expression of the adenosine diphosphate (ADP)-glucose pyrophosphorylase (AGPase) small subunit (AtAGPase-S1 and AtAGPase-S2), AGPase large subunit (AtAGPase-L1 and AtAGPase-L2) and soluble starch synthases (AtSSS I, AtSSS II, AtSSS III and AtSSS IV) genes involved in starch biosynthesis in the transgenic A. thaliana plants. Meanwhile, enzymatic analyses indicated that the major enzymes (AGPase and SSS) involved in the starch biosynthesis exhibited higher activities in the transgenic plants compared to the wild-type. These results suggest that StTrxF may improve the starch content of A. thaliana by up-regulating the expression of the related genes and increasing the activities of the major enzymes involved in starch biosynthesis. The StTrxF gene may be applied for increasing starch accumulation of plants in the future.
Introduction
Starch, as a main source of nutrition in human and animal diet, is a major storage carbohydrate in plants, which plays a fundamentally important role in plant survival and adaptation to varying environmental conditions [Citation1,Citation2]. In plants, starch is an insoluble glucan composed of two glucose polymers: amylose and amylopectin. Amylose mainly comprises linear chains that are linked by α-1,4 O-glycosidic bonds, whereas amylopectin is highly branched and contains 5%–6% α-1,6 O-glycosidic bonds to generate glucan branches of various lengths [Citation3]. Four major enzymes are involved in starch biosynthesis: adenosine diphosphate (ADP)-glucose pyrophosphorylase (AGPase), starch synthase, starch branching enzyme and starch debranching enzyme [Citation4,Citation5].
Starch metabolism is regulated at many levels, including allosteric regulation by redox regulation, protein–protein interactions and protein phosphorylation [Citation2,Citation6]. Redox regulation, which has been widely researched in plant starch biosynthesis, is also related to the coordination of enzymes involved in starch degradation [Citation6,Citation7]. The redox regulation of the AGPase [Citation8] and β-amylase 1 (BAM1) activity [Citation9,Citation10] are all widely known.
Thioredoxins (Trxs) are ubiquitous small proteins (ca. 12 kDa), members of the oxidoreductase family of enzymes. Trxs are essential players in the regulation of multiple cellular processes [Citation11,Citation12]. The active site of Trxs contains a conserved CXXC motif with two reactive cysteine (Cys) residues, which is located within the characteristic thioredoxin fold [Citation2]. Since the plastidic Trxs F-type proteins act in vitro as reductants for a number of the redox-regulated enzymes involved in starch metabolism, it has been suggested that the redox regulation that occurs in response to light signals is similar to that controlling other photosynthetic enzymes [Citation7,Citation13]. The TrxF gene has been shown to be involved in starch metabolism. Overexpression of NtTrxF increases the starch accumulation in tobacco leaves [Citation14] and knockout of AtTrxF1 decreases the starch content in the leaves of Arabidopsis thaliana mutants [Citation15].
There are several reports on increasing the starch content by genetic engineering in plants. For example, constitutive expression of the A. thaliana AtAATP1 gene can increase the starch content in potato tubers [Citation16]. Heterologous expression of SnRK1 from apple/sweet potato increases the starch content in transgenic tomato/tobacco plants [Citation17,Citation18]. Arabidopsis thaliana plants expressing AATP from tomato/potato exhibit higher starch content [Citation19,Citation20]. Overexpression of IbAATP increases the starch and amylose content in transgenic sweet potato [Citation21].
Although the function of Trxs from other plant species has been well studied, the regulatory role of StTrxF (Genbank accession No. XM_006345 713) from potato in increasing starch accumulation still remains unknown. In this work, the coding region of StTrxF from potato was cloned into a binary vector under the control of 35S promoter and constitutively expressed in A. thaliana to explore its role in starch accumulation.
Materials and methods
Plant materials
Potato cultivar Zhongshu No. 5 was employed for StTrxF gene cloning in this study. A. thaliana [ecotype Columbia-0, wild-type (WT)] was used as a model plant to investigate the functions of StTrxF.
Cloning and sequence analysis of the potato StTrxF gene
Total RNA was extracted from 0.5 g of fresh leaves of four-week-old in vitro-grown plants of Zhongshu No. 5 with the RNAprep Pure Kit (Tiangen Biotech, Beijing, China). RNA samples were reverse-transcribed according to the instructions of Quantscript Reverse Transcriptase Kit (Tiangen Biotech, Beijing, China). Based on the sequence of StTrxF (Genbank accession No. XM_006345 713), we designed a gene-specific primer for reverse transcription polymerase chain reaction (RT-PCR) () to obtain its full-length cDNA sequence. PCR was performed with an initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min and final extension at 72 °C for 10 min in a TaKaRa PCR Thermal Cycler Dice (TaKaRa, Beijing, China). PCR products were separated in a 1.0% (w/v) agarose gel. Target DNA bands were recovered by gel extraction, then cloned into PMD19-T (TaKaRa, Beijing, China), and finally transformed into competent cells of Escherichia coli strain DH5α. White colonies were checked by PCR and the positive colonies were sequenced (Invitrogen, Beijing, China).
Table 1. Primers used in this study.
The full-length cDNA of StTrxF was analysed by BLAST (Basic Local Alignment Search Tool) on the National Center for Biotechnology Information (NCBI) website (http://www.ncbi.nlm.nih.gov/). For multiple sequence alignment analysis, the amino-acid sequence of StTrxF and A. thaliana AtTrxF homologues retrieved from NCBI were aligned using the DNAMAN software (Lynnon Biosoft, Quebec, Canada). The theoretical molecular weight and isoelectronic point (pI) were calculated using the ProtParam tool (http://web.expasy.org/protparam/). The conserved domain of StTrxF was scanned by the InterProScan program (http://www.ebi.ac.uk/Tools/pfa/iprscan/).
Generation of transgenic A. thaliana plants
The coding region of StTrxF was cut out from the PMD19-T vector with the restriction enzymes BamHI and SacI, and then inserted into the same enzyme sites in pCAMBIA1301 to create the plant expression vector pCAMBIA1301-StTrxF, under the control of the cauliflower mosaic virus (CaMV) 35S promoter and the nopaline synthase (NOS) terminator. This vector also contains β-glucuronidase (gusA) and hygromycin resistance (hptII) genes driven by the CaMV 35S promoter. Both pCAMBIA1301-StTrxF and the control vector (VC) pCAMBIA1301 were transformed into Agrobacterium tumefaciens strain LBA4404 cells by the electroporation method for A. thaliana transformation [Citation22]. Transgenic plants were produced according to methods described previously [Citation23]. Transformants were selected based on their resistance to hygromycin (Hyg). Putative transformant seeds were germinated on agar-solidified Murashige and Skoog (MS) medium [Citation24] containing 25 mg/L Hyg. Positive transgenic seedlings were grown in pots containing a mixture of soil, vermiculite and humus (1:1:1, v/v/v) for T2 and T3 seed selection. The incubation and growth conditions of A. thaliana were the same as previously described [Citation23].
PCR analysis of transgenic plants
The presence of StTrxF in hygromycin-resistant plants was assessed by PCR analysis using specific primers () to amplify fragments of the hptII coding sequence. DNA was first extracted from A. thaliana leaves according to the instructions of EasyPure Plant Genomic DNA Kit (Transgen, Beijing, China). PCR amplifications were performed with an initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min and final extension at 72 °C for 10 min. PCR products were separated by electrophoresis in a 1.0% (w/v) agarose gel.
Starch content assays
Starch extraction and quantification were performed as described previously [Citation25]. Seeds were grown on MS medium for two weeks and transferred to pots containing a mixture of soil, vermiculite and humus (1:1:1, v/v/v). Plants were grown in a growth chamber for four weeks at 22 °C under standard long day conditions (14 h light and 10 h dark). Leaves of four-week-old plants were harvested, and then fresh weight was measured to determine starch content in light at 10–11 am. All treatments were performed in triplicate.
Expression analysis of the related genes
The expression of StTrxF and starch biosynthesis-related genes was analysed by real-time quantitative PCR (qRT-PCR). Transgenic, VC and WT plants were grown in pots for four weeks under standard/physiological conditions. Total RNA was extracted from the leaves of these plants, using the RNAprep Pure Plant Kit (Tiangen Biotech, Beijing, China). RNA samples were reverse-transcribed using Quantscript Reverse Transcriptase Kit (Tiangen Biotech, Beijing, China). The cDNA solution was used as templates for PCR amplification with gene-specific primers (). The A. thaliana Atactin gene (Genbank accession No. NM112764) was used as an internal control () [Citation26].
PCR amplifications were conducted by ABI PRISM 7500 (Software for 7500 and 7500 Fast Real-Time PCR Systems, V2.0.1, USA) using SYBR Green PCR Master Mix (Tiangen Biotech, Beijing, China). The amplifications were performed with an initial denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min. Quantification of gene expression was done with the comparative CT method [Citation27]. All experiments were repeated three times and each value represents the average of three experiments.
AGPase and SSS activity analyses
The activity of starch biosynthetic enzymes (AGPase and SSS) in the leaves of four-week-old transgenic, VC and WT plants was determined according to the method described by Nakamura et al. [Citation28] One unit of activity (AGPase and SSS) was defined as the formation of 1 nmol ADP per minute at 30 °C.
Statistical analysis
All experiments were repeated three times and the data are presented as mean values with standard error of the means (±SEM). Where applicable, data were analysed by Student's t-test in a two-tailed analysis. Values of P < 0.05 or P < 0.01 were considered to indicate statistically significant differences.
Results and discussion
Cloning and sequence analysis of StTrxF
The StTrxF (Genbank accession No. XM_006345 713) gene was cloned by RT-PCR. StTrxF contains a 549 bp open reading frame (ORF), encoding a polypeptide of 182 amino-acids, with a predicted molecular weight of 19.81 kDa and a theoretical isoelectric point (pI) of 9.15. Sequence analysis via InterProScan showed that the StTrxF protein contains a thioredoxin F-type domain ((a)). A BLASTP search indicated that the amino-acid sequence of StTrxF shares 58.92% and 54.59% amino-acid identity with AtTrxF1 (A. thaliana, AT3G02730) and AtTrxF2 (AT5G16400), respectively ((b)).
Figure 1. Gene structure and protein alignment analyses. (a) Structure of StTrxF and StTrxF. (b) Amino-acid sequence alignment of StTrxF with Arabidopsis AtTrxF1 and AtTrxF2.
Increased starch content in Arabidopsis thaliana expressing StTrxF
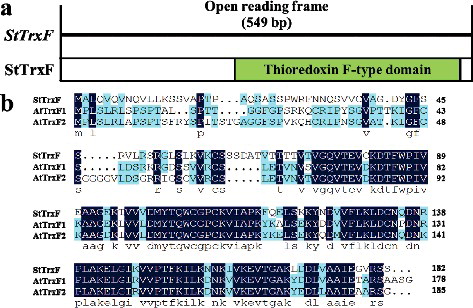
The ORF of StTrxF was ectopically expressed in A. thaliana (Col-0, WT) using the binary vector pCAMBIA1301-StTrxF ((a)). Six independent transgenic lines constitutively expressing StTrxF (T1 generation) were obtained by Hyg resistance selection, named #1–#6, respectively, and their progenies (T3 generation) were generated. PCR analysis of genomic DNA confirmed the successful integration of transgene ((b)). qRT-PCR analysis showed that the highest expression levels of StTrxF were observed in transgenic lines #2 and #3, while no transgene expression was observed in VC and WT ((c)). Therefore, transgenic lines #2 and #3 were selected for further analyses.
Figure 2. Molecular confirmation of transgenic plants. (a) Schematic diagram of the T-DNA region of binary plasmid pCAMBIA1301-StTrxF. LB, left border; RB, right border; hptII, hygromycin phosphotransferase II gene; StTrxF, potato thioredoxin type-F gene; gusA, β-glucuronidase gene; 35S, cauliflower mosaic virus (CaMV) 35S promoter; 35S T, CaMV 35S terminator; NOS T, nopaline synthase terminator. (b) PCR analysis of StTrxF expressing Arabidopsis plants. Lane M: DL2000 DNA marker (Transgen, Beijing, China); Lane W: water as a negative control; Lane P: plasmid pCAMBIA1301-StTrxF as a positive control; Lane WT: wild type; VC, control vector; Lanes #1–#6: different transgenic lines. (c) Expression levels of StTrxF in different transgenic lines. The A. thaliana Atactin gene was used as an internal control.
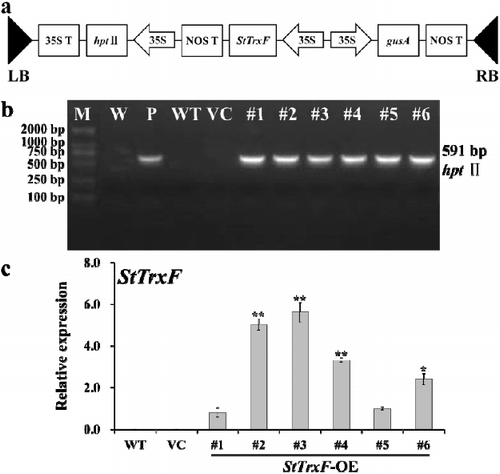
The TrxF gene has been shown to be involved in starch accumulation. The study by Thormählen et al. [Citation15] revealed that knockout of AtTrxF1 resulted in a significantly decreased starch accumulation in the leaves of A. thaliana mutants. The leaves of NtTrxF-overexpressing tobacco exhibited increased starch content [Citation14]. Here, two-week-old WT, VC and transgenic plants (lines #2 and #3) were grown in pots under standard/physiological conditions for four weeks. The expression of StTrxF did not change the growth of transgenic plants, since no significant differences in growth were observed between WT, VC and the transgenic plants under standard/physiological conditions. However, the starch content in the leaves of these plants was different. The starch content in the StTrxF-expressing plants increased 66%–87% compared to that in the WT plants (), whereas no significant difference was observed between VC and WT plants ().
Figure 3. Starch content analysis in the leaves of four-week-old WT, VC and StTrxF expressing plants grown under standard/physiological conditions.
Up-regulated starch biosynthetic genes and increased enzyme activities in A. thaliana expressing StTrxF
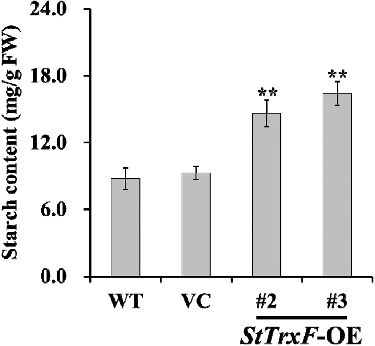
To dissect how expression of StTrxF increased the starch content in transgenic plants, the transcript levels of nine starch biosynthetic genes in WT, VC and transgenic plants (lines #2 and #3) were examined by qRT-PCR (). The activity of major enzymes (AGPase and SSS) involved in starch biosynthesis was also investigated in the leaves of WT, VC and transgenic plants (lines #2 and #3) ().
Figure 4. Gene expression analysis: transcription levels of starch biosynthesis genes in the leaves of four-week-old WT, VC and StTrxF expressing plants grown under standard/physiological conditions.
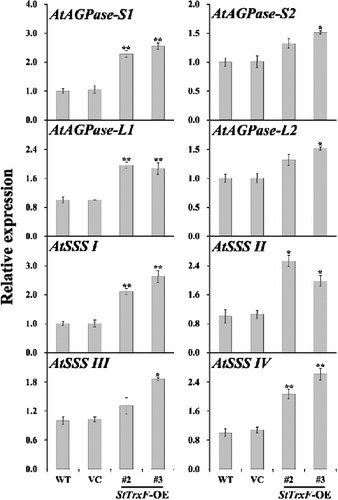
Figure 5. Enzyme activity assays: activities of AGPase and SSS in the leaves of four-week-old WT, VC and StTrxF expressing plants grown under standard/physiological conditions.
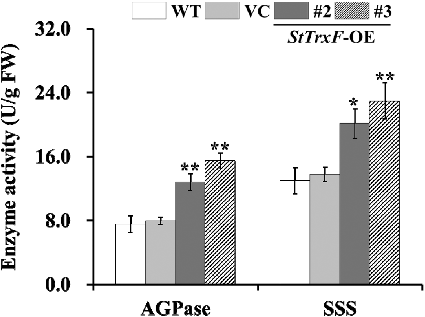
The TrxF gene is related to carbohydrate metabolism, functions in post-translational redox activation of AGPase and starch biosynthesis [Citation14,Citation15,Citation29,Citation30]. AGPase has been found to be subject to post-translational redox modification [Citation8]. Further study with a knockout mutant of AtTrxF1 also showed that loss of function of AtTrxF1 resulted in decreased redox activation of AGPase and starch accumulation in the mutant [Citation15]. Sanz-Barrio et al. [Citation14] reported that overexpression of NtTrxF increased the AGPase activity, leading to increased starch content in transgenic tobacco. Consistently, AGPase activity was also significantly increased in the StTrxF expressing A. thaliana plants (). In addition, the expression of AtAGPase-S1 and AtAGPase-S2, encoding the AGPase small subunit, and AtAGPase-L1 and AtAGPase-L2, encoding the AGPase large subunit, was up-regulated in transgenic plants (). Thus, constitutive expression of StTrxF up-regulated the expression of AtAGPase and increased the activity of AGPase, which further modulated starch metabolism ().
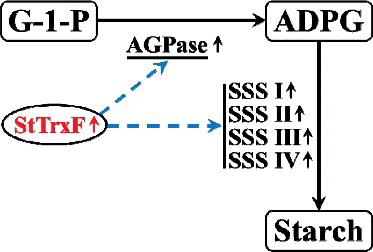
Figure 6. A proposed model of the regulatory network of StTrxF in starch accumulation. Biosynthesis pathway (solid arrows); regulatory interactions (broken arrows); up-regulation (↑) of the relative enzyme-encoding genes.
The higher level of starch content is related to the increased expression of starch biosynthesis genes [Citation3]. SSS can be grouped into four types, SSS I, SSS II, SSS III and SSS IV [Citation3–5]. A large body of evidence has illustrated that up-regulation of these genes could increase starch accumulation in plants [Citation5,Citation18–21,Citation31–35]. In our study, a significant increase of SSS activity was also observed in transgenic plants (). And consistently, the genes (AtSSS I, AtSSS II, AtSSS III and AtSSS IV) involved in starch biosynthesis were systematically up-regulated in transgenic plants ().
Taken together, constitutive expression of StTrxF up-regulated the expression of AtAGPase and increased the activity of AGPase, which further increased the expression of the genes and the activity of the major enzymes involved in starch biosynthesis, leading to increased starch accumulation in transgenic plants.
Conclusions
The StTrxF gene was successfully isolated from potato. Constitutive expression of StTrxF significantly increased the starch accumulation in transgenic A. thaliana plants by up-regulating the expression of the related genes and increasing the activities of the major enzymes involved in starch biosynthesis. Therefore, our results suggest that StTrxF plays a crucial role in starch metabolism, and has great potential in the engineering of alternative energy crop plants with improved starch accumulation.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Blennow A, Jensen SL, Shaik SS, et al. Future cereal starch bioengineering: cereal ancestors encounter gene technology and designer enzymes. Cereal Chem. 2013;90:274–287.
- Skryhan K, Cuesta-Seijo JA, Nielsen MM, et al. The role of cysteine residues in redox regulation and protein stability of Arabidopsis thaliana starch synthase 1. PLoS One [Internet]. 2015[ cited 2016 Jul 30];10:e0136997. Available from: http://journals.plos.org/plosone/article?idD10.1371/journal.pone.0136997.
- Delvallé D, Dumez S, Wattebled F, et al. Soluble starch synthase I: a major determinant for the synthesis of amylopectin in Arabidopsis thaliana leaves. Plant J. 2005;43:398–412.
- Fujita N, Yoshida M, Asakura N, et al. Function and characterization of starch synthase I using mutants in rice. Plant Physiol. 2006;140:1070–1084.
- Szydlowski N, Ragel P, Raynaud S, et al. Starch granule initiation in Arabidopsis requires the presence of either class IV or class III starch synthase. Plant Cell. 2009;21:2443–2457.
- Kötting O, Kossmann J, Zeeman SC, et al. Regulation of starch metabolism: the age of enlightenment? Curr Opin Plant Biol. 2010;13:321–329.
- Sanz-Barrio R, Corral-Martinez P, Ancin M, et al. Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnol J. 2013;11:618–627.
- Geigenberger P, Stamme C, Tjaden J, et al. Tuber physiology and properties of starch from tubers of transgenic potato plants with altered plastidic adenylate transporter activity. Plant Physiol. 2001;125:1667–1678.
- Sparla F, Costa A, Lo Schiavo F, et al. Redox regulation of a novel plastid-targeted beta-amylase of Arabidopsis. Plant Physiol. 2006;141:840–850.
- Valerio C, Costa A, Marri L, et al. Thioredoxin-regulated beta-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress. J Exp Bot. 2010;62:545–555.
- Gelhaye E, Rouhier N, Navrot N, et al. The plant thioredoxin system. Cell Mol Life Sci. 2005;62:24–35.
- Meyer Y, Reichheld JP, Vignols F. Thioredoxins in Arabidopsis and other plants. Photosynth Res. 2005;86:419–433.
- Schürmann P, Buchanan BB. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid Redox Signal. 2008;10:1235–1274.
- Sanz-Barrio R, Corral-Martinez P, Ancin M, et al. Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnol J. 2013;11:618–627.
- Thormählen I, Ruber J, von Roepenack-Lahaye E, et al. Inactivation of thioredoxin f1 leads to decreased light activation of ADP-glucose pyrophosphorylase and altered diurnal starch turnover in leaves of Arabidopsis plants. Plant Cell Environ. 2013;36:16–29.
- Tjaden J, Möhlmann T, Kampfenkel K, et al. Altered plastidic ATP/ADP-transporter activity influences potato (Solanum tuberosum L.) tuber morphology, yield and composition of tuber starch. Plant J. 1998;16:531–540.
- Wang X, Peng F, Li M, et al. Expression of a heterologous SnRK1 in tomato increases carbon assimilation, nitrogen uptake and modifies fruit development. J Plant Physiol. 2012;169:1173–1182.
- Jiang T, Zhai H, Wang FB, et al. Cloning and characterization of a carbohydrate metabolism-associated gene IbSnRK1 from sweetpotato. Sci Hortic. 2013;158:22–32.
- Wang FB, Ye YX, Niu Y, et al. A tomato plastidic ATP/ADP transporter gene SlAATP increases starch content in transgenic Arabidopsis. Physiol Mol Biol Plants. 2016;22:497–506.
- Wang FB, Fu LF, Kong WL, et al. Constitutive expression of StAATP, a potato plastidic ATP/ADP transporter gene, increases starch content in transgenic Arabidopsis. Biotechnol Biotechnol Equip. 2017 [cited 2017 Feb 24]. DOI: 10.1080/13102818.2017.1282837
- Wang YN, Li Y, Zhang H, et al. A plastidic ATP/ADP transporter gene, IbAATP, increases starch and amylose content and alters starch structure in transgenic sweetpotato. J Integr Agr. 2016;15:1968–1982.
- Lou XM, Yao QH, Zhang Z, et al. Expression of human hepatitis B virus large surface antigen gene in transgenic tomato. Clin Vaccine Immunol. 2007;14:464–469.
- Zhang X, Henriques R, Lin SS. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat Protoc. 2006;1:641–646.
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962;15:473–497.
- Smith AM, Zeeman SC. Quantification of starch in plant tissues. Nat Protoc. 2006;1:1342–1345.
- Li X, Ma H, Huang H, et al. Natural anthocyanins from phytoresources and their chemical researches. Nat Prod Res. 2013;27:456–469.
- Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative CT method. Nat Protoc. 2008;3:1101–1108.
- Nakamura Y, Yuki K, Park SY, et al. Carbohydrate metabolism in the developing endosperm of rice grains. Plant Cell Physiol. 1989;30:833–839.
- Ballicora MA, Frueauf JB, Fu Y, et al. Activation of the potato tuber ADP-glucose pyrophosphorylase by thioredoxin. J Biol Chem. 2000;275:1315–1320.
- Geigenberger P, Kolbe A, Tiessen A. Redox regulation of carbon storage and partitioning in response to light and sugars. J Exp Bot. 2005;56:1469–1479.
- Burton RA, Jenner H, Carrangis L, et al. Starch granule initiation and growth are altered in barley mutants that lack isoamylase activity. Plant J. 2002;31:97–112.
- Bustos R, Fahy B, Hylton CM, et al. Starch granule initiation is controlled by a heteromultimeric isoamylase in potato tubers. PNAS. 2004;101:2215–2220.
- Roldan I, Wattebled F, Lucas MM, et al. The phenotype of soluble starch synthase IV defective mutants of Arabidopsis thaliana suggests a novel function of elongation enzymes in the control of starch granule formation. Plant J. 2007;49:492–504.
- Wang FB, Guo XT, Qiao XQ, et al. The maize plastidic thioredoxin F-type gene ZmTrxF increases starch accumulation in transgenic Arabidopsis. Sci Hortic. 2016;210:205–212.
- Wang FB, Kong WL, Fu YR, et al. Constitutive expression of SlTrxF increases starch content in transgenic Arabidopsis. Biol Plantarum. 2016 [ cited 2017 Feb 24]. DOI:10.1007/s10535-016-0675-6