
Abstract
Many proteins in public databases are recognized as proteins with domains of unknown function (DUF). DUF1191 is such a protein family that consists of plant proteins with unknown function. In this study, we analyzed the basic information of four OsDUF1191 members (OsDUF1191.1–OsDUF1191.4) in rice Nipponbare. OsDUF1191 family members can be divided into four categories by phylogenetic analysis. In order to find out whether these members have tissue-specific expression preference, we used real-time polymerase chain reaction to examine the expression in 15 distinct rice tissues. We found that OSDUF1191.1 was preferentially expressed in the roots. The expression of the four OsDUF1191 members under various abiotic stress and abscisic acid (ABA) conditions was also examined. The expression of OsDUF1191.4 was significantly (p < 0.01) upregulated under heat stress conditions. Overexpression of OsDUF1191.4 in Escherichia coli led to a significant enhancement of catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD) activities, and improved bacterial resistance to heat stress. The results can provide new insights into the function of the OsDUF1191 family in rice.
Keywords:
Introduction
Even though deep research has been done on organisms, many functions of proteins have not been marked and annotated. Many proteins in public databases are recognized as proteins with domains of unknown function (DUF) [Citation1]. The names of DUFs are given when protein families have no functional annotation in the Pfam database (http://pfam.xfam.org/) [Citation2]. In the Pfam database, there are about 3600 domains of unknown function that account for 20% of all domains [Citation3–4]. Many DUF structures are highly conserved and they play different roles in biology [Citation5–9].
Rice is an important crop in arable land worldwide. Some environmental factors, such as drought, salinity, nutrient imbalances and extreme temperatures, have important effects on the growth and final yield of crops [Citation10]. In order to make the growth, production and living capacity of crops suffer limited damage during the whole process of plant life, plants have evolved a reaction mechanism that is controlled by genes [Citation11]. It can be reflected in molecules, cells, tissues, organs or individual plants [Citation12–16]. At present, some genes have been used successfully to improve the tolerance of stresses in rice [Citation17–21].
The DUF1191 family consists of unknown functional plant proteins, with an average length of about 253 residues. So far, the function of OsDUF1191 family in rice has not been identified. In this study, we analyzed the OsDUF1191 family in rice and studied the expression profile of four OsDUF1191 members and their expression pattern under various stresses and abscisic acid (ABA) treatment. Furthermore, we overexpressed gene OsDUF1191.4 in Escherichia coli and the results showed that OsDUF1191.4 could improve the tolerance to heat in transgenic E. coli. The results may provide new insights into the function of the OsDUF1191 family in rice.
Materials and methods
Database searches
We obtained the sequence of assumed OsDUF1191 family proteins in rice by retrieving the rice genome annotation project(RGAP version 7)database (http://rice.plantbiology.msu.edu/) with domain number PF06697. Then downloaded all the related protein sequences of the OsDUF1191 family from RGAP and confirmed with the SMART database (http://smart.embl-heidelberg.de/smart/batch.pl) [Citation22]. If there were several gene models at one locus, only the complete gene model was selected for further sequence analysis.
Sequence analysis
RGAP and Knowledge-Based Oryza Molecular Biological Encyclopedia (KOME) (http://cdna01.dna.affrc.go.jp/cDNA) can be used to query the information about chromosomal localization, amino acid length, full-length cDNA accessions for each OsDUF1191 family member. The protein localization of OsDUF1191 members was predicted by online protein location prediction software TargetP 1.1 (http://www.cbs.dtu.dk/services/TargetP/) [Citation23]. Motifs of OsDUF1191 family members were predicted using MEME version 4.11.2 (http://meme.nbcr.net/meme/) [Citation24]. MEGA4 was used to construct the phylogenetic tree [Citation25], using the maximum-likelihood method.
Plant growth and various stress and ABA treatment
Rice (Oryza sativa L. subsp. japonica cv. Nipponbare) seeds were germinated for 3 days before they were transferred to a 30-L nutrient solution container for hydroponic growth [Citation26]. When the fourth leaf of rice seedlings appeared, they were subjected to various stresses and ABA treatment. For the salt treatment, high salinity stress was achieved by adding NaCl to the solution to a final concentration of about 200 mmol/L. For the drought tests, intact seedlings were put in the air without water supply. For the cold tests, the seedlings were transferred to a chamber at 4 °C (12-h-light/12-h-dark cycle). For ABA treatment, 100 μmol/L ABA was sprayed onto the leaves of seedlings. For heat treatment, the seedlings were exposed to 42 °C. Stress treatment continued until the seedlings were sampled. For salt stress and drought stress, seedlings were sampled at 0, 4, 8 and 16 h. For ABA, heat and cold stress, seedlings were sampled at 0, 1, 3, 8 h.
In order to further understand the expression patterns of OsDUF1191 family members, 15 different tissues of rice Nipponbare seedlings were sampled, as follows: Lb1, leaf blade at the four-leaf stage; Lb2, leaf blade from plants with four tillers; Lb3, leaf blade at the ripening stage; Ls1, leaf sheath at the four-leaf stage; Ls2, leaf sheath from plants with four tillers; Rt1, root at the four-leaf stage; Rt2, root from plants with four tillers; St1, stem from plants with four tillers; St2, stem at the ripening stage; An, 1.2–1.5 mm anther; Pi, pistil from 10–14 cm inflorescence; Em1, embryo at 7 days after flowering; Em2, embryo at 28 days after flowering; En1, endosperm at 7 days after flowering; En2, endosperm at 28 days after flowering.
RNA extraction and real-time polymerase chain reaction
The total RNA was extracted using Trizol reagent (GIBCO, Burlington, ON, USA) according to the instructions. In order to degrade any possible DNA contamination in total RNA, it was treated with RNase-free DNase for 15 min. Real-time PCR was performed as described before [Citation27]. The rice Actin1 gene (LOC_Os03g50885) was used as an internal control with primers 5′-TGGCATCTCTCAGCACATTC-3′ and 5′-TGCACAATGGATGGGCCAGA-3′. The primers used in the expression analysis of the OsDUF1191 gene family members are listed in Supplementary material Table S1. The 2-△△Ct method was used to analyze the relative expression data as mentioned previously [Citation28].
Assay for heat stress tolerance of E. coli transformants
In order to construct the expression vector pET32a-OsDUF1191.4, specific primers were designed as follows: sense, 5′-GGATCCATGGATTTCTTGGCCTGCGT-3′ (BamHI site underlined) and antisense, 5′-AAGCTTCTATGCAACATACTCGCTCT-3′ (HindIII site underlined). OsDUF1191.4 was transformed into E. coli according to the previous method [Citation29]. The transformed E. coli Rosetta cells (Sanborn, MN, USA) were grown in Luria-Bertani (LB) liquid medium including 100 mg/mL of ampicillin at 37 °C. When the optical density of bacteria at 600 nm reached 0.5 - 0.6, 1 mmol/L isopropyl-β-D-thiogalactoside (IPTG) was added to induce the expression of the target protein. In the heat tolerance test, at 0, 0.5, 1, 1.5, 2, 2.5 h after heat stress treatment (50 °C water bath), respectively, 100 μL of dilution (1:100) was spotted onto LB agar plates with 1 mmol/L IPTG.
Measurements of antioxidant enzyme activity
The bacteria were collected by centrifugation. After ultrasonic cell-breakage at 4 °C, the supernatant (as a crude enzyme solution) was collected by spinning at 12,000 rpm (13200 xg rcf) for 20 min. CAT, SOD and POD activities were determined as described previously [Citation30].
Data analysis
The experiment was arranged as a completely randomized design and all the experiments were repeated three times. Data were obtained from three independent experiments with standard error of the means. Statistical analysis was performed using the Student’s t-test by EXCEL version 2003.
Results and discussion
Identification of OsDUF1191 family members in rice
In order to identify four members of the OsDUF1191 family, we searched the rice genome annotation project (RGAP) using domain number PF06697. The SMART database was then used to confirm the presence of these OsDUF1191 family members in the rice genome. These genes are named OsDUF1191.1 to OsDUF1191.4 according to their position on the pseudomolecules (). Four members belonging to the rice OsDUF1191 gene family are distributed on chromosome 4 (OsDUF1191.1 and OsDUF1191.2), chromosome 6 (OsDUF1191.3) and chromosome 11 (OsDUF1191.4) ().
Table 1. OsDUF1191 genes and the properties of the putative proteins they encode.
As the rice genome includes 10 duplicated blocks accounting for 45% of its total size [Citation31], it is possible that the distribution of OsDUF1191 family members to different rice chromosomes could be explained by these large segment duplications, similar to that previously suggested for OsDUF866, OsDUF946, OsDUF936 and OsDUF829 family members [Citation27,Citation29,Citation32,Citation33].
Sequence analysis of OsDUF1191 family proteins
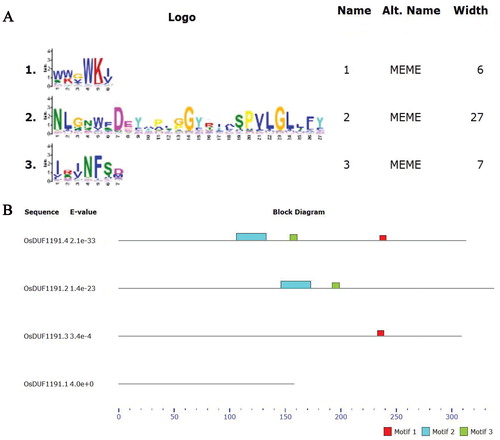
Information about the OsDUF1191 family is listed in . The predicted length of the OsDUF1191 proteins varies from 158 (OsDUF1191.1) to 338 (OsDUF1191.2) amino acids (). Interestingly, OsDUF1191.3 contains one intron, whereas the other three OsDUF1191 members do not contain any introns. The predicted protein of OsDUF1191.1 is located in the nucleus and OsDUF1191.2 is located in chloroplasts, whereas OsDUF1191.3 and OsDUF1191.4 are located in the secretory pathway. Three distinct motifs in the four OsDUF1191 members were identified using the MEME motif search tool (). In OsDUF1191.4, motif 1, motif 2 and motif 3 were found, whereas motif 2 and motif 3 were found in OsDUF1191.2. Interestingly, OsDUF1191.3 contains only motif 1, whereas OsDUF1191.1 contains none.
Figure 1. Conservative structure analysis of rice OsDUF 1191 family. (A) Motif 1, motif 2 and motif 3 are conserved motifs of rice OsDUF1191 family obtained by MEME. (B) The distribution of conserved motifs identified by MEME software in OsDUF1191 members.
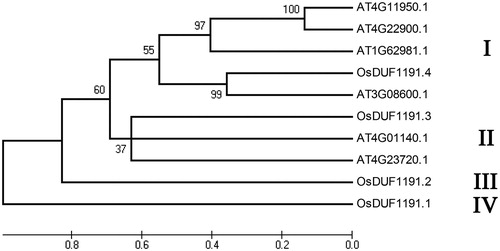
The multiple sequence alignment analysis of the OsDUF1191 members was performed using Clustal Omega (Supplementary material Figure S1). The percent identity was 20.37% for OsDUF1191.1 and OsDUF1191.2, 34.15% for OsDUF1191.1 and OsDUF1191.3, 29.27% for OsDUF1191.1 and OsDUF1191.4, 26.21% for OsDUF1191.2 and OsDUF1191.3, 31.91% for OsDUF1191.2 and OsDUF1191.4 and 31.51 for OsDUF1191.3 and OsDUF1191.4. The phylogenetic analysis was carried out using MEGA4 in order to examine the evolutionary relationship among DUF1191 family members in rice and Arabidopsis. Here, the reference sequence includes six Arabidopsis DUF119 members. The results showed that the members of the DUF1191 family can be divided into four major groups (I, II, III, IV) (). Group I consists of 5 members (AT4G11950.1, AT4G22900.1, AT1G62981.1, OsDUF1191.4 and AT3G08600.1), and group II contains 3 members (OsDUF1191.3, AT4G01140.1 and AT4G23720.1). Groups III and IV include only one member each, OsDUF1191.2 and OsDUF1191.1, respectively.
Figure 2. Phylogenetic relationship of OsDUF1191 members in rice and four DUF1191 members in Arabidopsis.
Note: The unrooted tree was generated using MEGA4.0 program by the maximum-likelihood method.
Expression patterns of OsDUF1191 family members
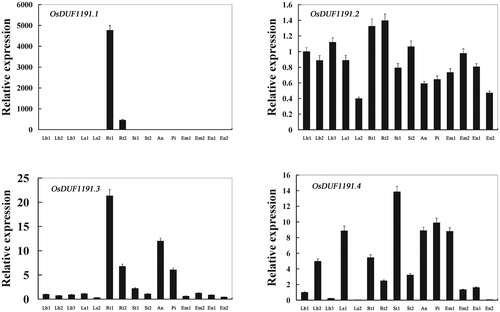
So far, few studies have been done about the gene expression patterns of OsDUF1191 genes. Therefore, we used real-time PCR to investigate the expression patterns of OsDUF1191 family genes in 15 samples from different tissues or stages. The results showed that the expression patterns of every member of the OsDUF1191 family had their own characteristics (). OsDUF1191.1 was specifically expressed in the Rt1 (root at the four-leaf stage) and Rt2 (root from plants with four tillers), and its expression was very poor or not detected in other tissues. Therefore, it is speculated that OsDUF1191.1 plays a crucial role in rice root development. OsDUF1191.2 was always highly expressed in almost all 15 samples. For OsDUF1191.3, the highest expression was in the Rt1 (root at the four-leaf stage), however the lowest expression was in the Ls2 (leaf sheath from plants with four tillers). For OsDUF1191.4, the expression level in the St1 (stem from plants with four tillers) was the highest, whereas the expression level in the Ls2 (leaf sheath from plants with four tillers) was extremely low. According to the different expression patterns of OsDUF1191 family members, we speculated that they may play distinct roles in different tissues in rice.
Figure 3. Real-time PCR analysis of OsDUF1191 genes in different tissues of Nipponbare rice. Fifteen representative tissues are as follows: Lb1, leaf blade at four-leaf stage; Lb2, leaf blade from plants with four tillers; Lb3, leaf blade at ripening stage; Ls1, leaf sheath at four-leaf stage; Ls2, leaf sheath from plants with four tillers; Rt1, root at four-leaf stage; Rt2, root from plants with four tillers; St1, stem from plants with four tillers; St2, stem at ripening stage; An, 1.2–1.5 mm anther; Pi, pistil from 10–14 cm inflorescence; Em1, embryo at 7 days after flowering; Em2, embryo at 28 days after flowering; En1, endosperm at 7 days after flowering; En2, endosperm at 28 days after flowering.
Note: The rice Actin1 transcript levels were used as internal controls. Error bars indicate standard error of the means based on three biological replicates.
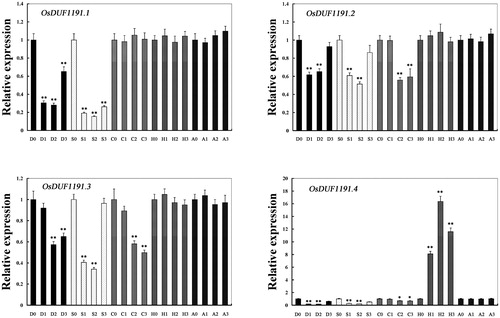
Real-time PCR was performed to examine the expression of OsDUF1191 family genes under various stress conditions and ABA treatment (). Under drought and salinity conditions, the expression of four OsDUF1191 family members was significantly (p < 0.01) downregulated. Under cold treatment, the expression of OsDUF1191.2, OsDUF1191.3 and OsDUF1191.4 was significantly (p < 0.01) downregulated, whereas the expression of OsDUF1191.1 remained unchanged. Under heat conditions, only the expression of OsDUF1191.4 increased significantly (p < 0.01). The results indicated that different members in OsDUF1191 family play different roles in response to various abiotic stress.
Figure 4. Relative expression levels of OsDUF1191 genes in Nipponbare rice seedlings at the emergence of the fourth leaf under various stress conditions and ABA treatment detected by real time PCR. D, drought; S, salt; C; cold; H, heat; A, ABA.
Note: For salt stress and drought stress, seedlings were sampled at 0, 4, 8 and 16 h. For ABA, heat and cold stress, seedlings were sampled at 0, 1, 3, 8 h. The rice Actin1 transcript levels were used as internal controls. Error bars indicate standard error of the means based on three biological replicates. **, P < 0.01 (Student’s t-test).
OsDUF1191.4 improved heat stress resistance in transgenic E. coli
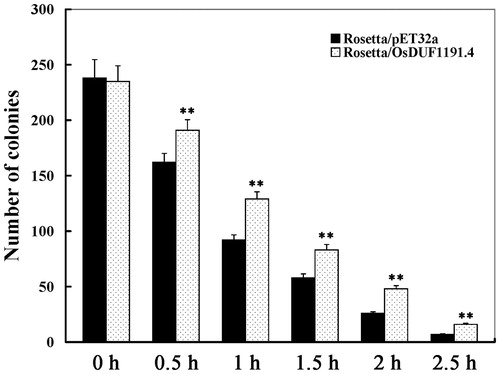
As mentioned above, the expression of OsDUF1191.4 increased significantly under heat conditions, so it is speculated that the gene may enhance the heat tolerance of plants. To test this hypothesis, we overexpressed the gene OsDUF1191.4 in E. coli. Under normal conditions, the difference in the growth rate between transgenic lines (E. coli transformed with pET32a-OsDUF1191.4) and the control (E. coli transformed with pET-32a) was not evident. However, under heat conditions, transgenic colonies transformed with pET32a-OsDUF1191.4 outnumbered the control (Rosetta/pET-32a) at 0.5, 1, 1.5, 2, 2.5 h after heat treatment, respectively (). This result shows that the overexpression of OsDUF1191.4 in E. coli may significantly improve the resistance to heat.
Figure 5. Growth of E. coli recombinants overexpresing OsDUF1191.4 under heat stress.
Note: 100-μL dilutions were spotted onto LB agar plates with 1 mmol/L IPTG at 0–2.5 h after heat shock (50 °C water bath), and then the number of the control (Rosetta/pET-32a) and recombinant (Rosetta/OsDUF1191.4) colonies was counted. Error bars indicate standard error of the means based on three biological replicates. **, P < 0.01 (Student’s t-test).
OsDUF1191.4 improves the antioxidant defenses of E. coli transformants grown under heat conditions
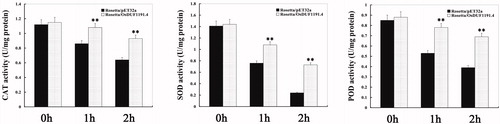
In order to investigate the effects of OsDUF1191.4 on the intracellular antioxidant ability, we determined catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD) activities of E. coli transformants and control bacteria. Under normal conditions, there was no evident difference in CAT, SOD and POD activities between transgenic lines and the control (). Under heat conditions, CAT, SOD and POD activities of E. coli transformed with pET32a-OsDUF1191.4 were significantly higher than those of the control from 1 to 2 h (). These results indicated that the improvement in the tolerance to stress in E. coli OsDUF1191.4 transformants may be related to the increase in the activities of CAT, SOD and POD under heat conditions.
Figure 6. Antioxidant ability of E. coli transformants overexpressing OsDUF1191.4 under heat stress.
Note: After E. coli transformants were exposed to heat shock conditions (50 °C water bath) for 0, 1, and 2 h, respectively, catalase (CAT), superoxide dismutase (SOD) and peroxidase (POD) activities were determined. Error bars indicate standard error of the means based on three biological replicates. **, P < 0.01 (Student’s t test).
Production of reactive oxygen species (ROS) is a common event in organisms subjected to abiotic stresses, such as heat, and ROS production can cause oxidative damage to membrane lipids and perturbation of cell membrane function [Citation34]. Maintaining the balance between ROS generation and ROS scavenging in cells requires CAT, SOD and POD ROS scavenging enzymes. In this study, we observed an increase in activities of CAT, SOD and POD in transgenic E. coli. This increase possibly indicates that the extent of damage to the E. coli transformed with pET32a-OsDUF1191.4 exposed to heat stress is lower than that in the control bacteria exposed to the same amounts of stress.
So far, the specific mechanism by which heat tolerance is enhanced in E. coli recombinants transformed with OsDUF1191.4 remains unclear. Under heat conditions, it is possible that OsDUF1191.4 interacts with some heat shock protein, thus better initiating the heat shock response in transgenic E. coli. Another possibility is that the heat stress signal was strengthened in E. coli recombinants transformed with OsDUF1191.4, and the enhanced heat stress signal promotes the increased expression of some heat stress-related genes, including genes encoding ROS-scavenging enzymes, leading to better improvement in heat tolerance in transgenic E. coli.
Conclusions
This study provides important sequence information regarding the OsDUF1191 family members and the expression profile of OsDUF1191 family members. Our results suggest that OsDUF1191.4 may improve the resistance to heat stresses in transgenic E. coli. These data can provide important reference for further research on the function of OsDUF1191 family.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Bateman A, Coggill P, Finn RD. DUFs: families in search of function. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2010;66(10):1148–1152.
- Mudgal R, Sandhya S, Chandra N, et al. De-DUFing the DUFs: Deciphering distant evolutionary relationships of Domains of Unknown Function using sensitive homology detection methods. Biol Direct. 2015;10(1):38.
- Goodacre NF, Gerloff DL, Uetz P. Protein domains of unknown function are essential in bacteria. MBio [Internet]. 2014;5:e00744–00713. Available from: http://mbio.asm.org/content/5/1/e00744-13.full.
- Finn RD, Coggill P, Eberhardt RY, et al. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 2016;44(D1):D279–285.
- He X, Hou X, Shen Y, et al. TaSRG, a wheat transcription factor, significantly affects salt tolerance in transgenic rice and Arabidopsis. FEBS Lett. 2011;585(8):1231–1237.
- Kim SJ, Ryu MY, Kim WT. Suppression of Arabidopsis RING-DUF1117 E3 ubiquitin ligases, AtRDUF1 and AtRDUF2,reduces tolerance to ABA-mediated drought stress. Bio-chem Biophys Res Commun. 2012;420(1):141–147.
- Luo C, Guo C, Wang W, et al. Overexpression of a new stress-repressive gene OsDSR2 encoding a protein with a DUF966 domain increases salt and simulated drought stress sensitivities and reduces ABA sensitivity in rice. Plant Cell Rep. 2014;33(2):323–336.
- Wang L, Shen R, Chen LT, et al. Characterization of a novel DUF1618 gene family in rice. J Integr Plant Biol. 2014;56(2):151–158.
- Guo C, Luo C, Guo L, et al. OsSIDP366, a DUF1644 gene, positively regulates responses to drought and salt stresses in rice. J Integr Plant Biol. 2016;58(5):492–502.
- Boyer JS. Plant productivity and environment. Science. 1982;218(4571):443–448.
- Mittler R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006;11(1):15–19.
- Wang W, Vinocur B, Altman A. Plant responses to drought,salinity and extreme temperatures: towards genetic engieering for stress tolerance. Planta. 2003;218(1):1–14.
- Herms DA, Mattson WJ. The dilemma of plants: to grow ordefend. Q Rev Biol. 1992;67(3):283–335.
- Soltis DE, Soltis PS. The role of phylogenetics in comparative genetics. Plant Physiol. 2003;132(4):1790–1800.
- Xiang Y, Tang N, Du H, et al. Characterization of OsbZIP23as a key player of the basic leucine zipper transcription factor fmaily or conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008;148(4):1938–1952.
- Sato H, Todaka D, Kudo M, et al. The Arabidopsis transcriptional regulator DPB3-1 enhances heat stress tolerance without growth retardation in rice. Plant Biotechnol J. 2016;14(8):1756–1767.
- Liu J, Zhang C, Wei C, et al. The RING finger ubiquitin E3 ligase OsHTAS enhances heat tolerance by promoting H2O2-induced stomatal closure in rice. Plant Physiol. 2016;170(1):429–443.
- Hong Y, Zhang H, Huang L, et al. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front Plant Sci [Internet]. 2016. [cited 2016 Jul2];7:4. Available from: http://journal.frontiersin.org/article/10.3389/fpls.2016.00004/full.
- Yoon DH, Lee SS, Park HJ, et al. Overexpression of OsCYP19-4 increases tolerance to cold stress and enhances grain yield in rice (Oryza sativa). J. Exp. BOT. 2016;67(1):69–82.
- Min HJ, Jung YJ, Kang BG, et al. CaPUB1, a hot pepper U- box E3 ubiquitin ligase, confers enhanced cold stress tolerance and decreased drought stress tolerance in trans-genic rice (Oryza sativa L.). Mol Cells 2016;39:250–257.
- Shinozaki K, Yamaguchi-Shinozaki K, Seki M. Regulatory network of gene expression in the drought and cold stressm responses. Curr Opin Plant Biol. 2003;6(5):410–417. 5:5857–5864.
- Letunic I, Doerks T, Bork P. SMART: recent updates, new developments and status in 2015. Nucleic Acids Res. 2014;43:257–260.
- Emanuelsson O, Brunak S, von Heijne G, et al. Locating proteins in the cell using TargetP, SignalP and related tools. Nat Protoc. 2007;2(4):953–971.
- Bailey TL, Boden M, Buske FA, et al. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37:202–208.
- Tamura K, Dudley J, Nei M, et al. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol. 2007;24(8):1596–1599.
- Li L, Liu C, Lian X. Gene expression profiles in rice roots under low phosphorus stress. Plant Mol Biol. 2010;72(4-5):423–432.
- Li L, Xie C, Ye T, et al. Molecular characterization, expression pattern, and function analysis of the rice OsDUF866 family. Biotechnol Biotechnol Equip. 2017;31(2):243–249.
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-DDC(T)) method. Methods 2001;25(4):402–408.
- Li L, Ye T, Xu J, et al. Molecular characterization and function analysis of the rice OsDUF946 family. Biotechnol Biotechnol Equip. 2017;31(3):477–485.
- Liang Y, Chen Q, Liu Q, et al. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J Plant Physiol. 2003;160(10):1157–1164.
- Wang X, Shi X, Hao B, et al. Duplication and DNA segmental loss in the rice genome: implications for diploidization. New Phytol. 2005;165(3):937–946.
- Li L, Ye T, Guan Y, et al. Genome-wide identification and analyses of the rice OsDUF936 family. Biotechnol Biotechnol Equip. 2018;32(2):309–315.
- Li L, Lv M, Zhao L, et al. Molecular characterization and function analysis of the rice OsDUF829 family. Biotechnol Biotechnol Equip. 2018;32(3):550–557.
- Miller G, Shulaev V, Mittler R. Reactive oxygen signaling and abiotic stress. Physiol Plant. 2008;133(3):481–489.