
ABSTRACT
Humans are good at recognizing familiar faces, but are more error-prone at recognizing an unfamiliar person across different images. It has been suggested that familiar and unfamiliar faces are processed qualitatively differently. But are unfamiliar faces at least processed differently from monkey faces? Here we tested 366 volunteers on a face matching test – two images presented side-by-side with participants judging whether the images show the same identity or two different identities – comparing performance with familiar and unfamiliar human faces, and monkey faces. The results showed that performance was most accurate for familiar faces, and was above chance for monkey faces. Although accuracy was higher for unfamiliar humans than monkeys on different identity trials, there was no unfamiliar human advantage over monkeys on same identity trials. The results give new insights into unfamiliar face processing, showing that in some ways unfamiliar faces might as well be another species.
KEYWORDS:
Introduction
It is often argued that as social animals, humans are “face experts” (see Diamond & Carey, Citation1986), but this statement ignores a basic distinction between faces – that is whether or not they are familiar to us (Young & Burton, Citation2018). While humans are good at recognizing familiar faces, we are much poorer at recognizing unfamiliar people across different images (e.g., Bruce et al., Citation2001). This phenomenon has been demonstrated using many different paradigms, including face matching tasks where participants are shown two images side-by-side and are asked to determine whether the images show the same person or two different people. Standard tests of face matching such as the Kent Face Matching Test (Fysh & Bindemann, Citation2018) show that although performance on this task with unfamiliar faces is above chance, people are much poorer at matching unfamiliar compared to familiar faces, even when the images have been taken only moments apart (as in the Glasgow Face Matching Test and its updated version, Burton et al., Citation2010; White et al., Citation2022). A recent study found typical observers’ face matching accuracy was over 90% for familiar faces (celebrities) and around 10% poorer for unfamiliar faces (Noyes et al., Citation2021). This is an important issue as we are frequently asked to prove our identity through the use of photo-ID which is checked by someone who is unfamiliar with us, and an increase in error rates with unfamiliar faces could create security risks. Studies have shown that live face-to-photo matching is no more accurate than matching two photographs (Davis & Valentine, Citation2009; Ritchie et al., Citation2020), and that security officials such as police (Burton et al., Citation1999) and border control officers (White et al., Citation2014) are no more accurate than untrained individuals. With familiar faces, however, performance is much more accurate (e.g., Ritchie et al., Citation2015), even when the faces are distorted (Burton et al., Citation1999) or occluded by sunglasses or face masks (Noyes et al., Citation2021).
It has been argued that we are good at recognizing familiar faces across different images because we have previous exposure to the way that person’s face can vary, whereas we have no prior knowledge with unfamiliar faces (Bruce, Citation1994; Burton, Citation2013). Evidence from the face learning literature corroborates this argument, as multiple studies have found that exposure to within-person variability helps with face learning (Longmore et al., Citation2008; Longmore et al., Citation2017; Matthews et al., Citation2018; Murphy et al., Citation2015; Ritchie & Burton, Citation2017). Face sorting tasks (e.g., Jenkins et al., Citation2011) present participants with a set of images typically depicting two similar-looking people. Participants are not told how many people are depicted in the set, and are asked to sort the images into piles with each pile denoting one identity. While familiar observers can do this task without error, creating two piles each containing images of only one identity, unfamiliar observers typically create around 7 piles or “identities’”. What is particularly interesting about unfamiliar observers’ performance is that they typically create multiple piles of each identity without confusing the identities within the same pile. That is, they can tell people apart by separating the images of person A from the images of person B, but they cannot “tell people together” by seeing that their three piles of person B images should be combined into one identity. This suggests that poor unfamiliar face recognition may be an error of cohering together multiple images of one identity rather than confusing images of different people.
Despite its importance, we know little about the mechanisms underlying the differences between familiar and unfamiliar face processing, and how we represent different faces. In fact one study suggested in its title that “unfamiliar faces are not [even] faces” (Megreya & Burton, Citation2006) citing evidence of a qualitative difference between familiar and unfamiliar face processing. But what about faces from a different species? Are unfamiliar faces at least processed differently from monkey faces? The distinction between familiar and unfamiliar faces has not received attention in literature surrounding inter-species face perception. It has been suggested that species-specific face recognition whereby babies show preferential recognition for human as opposed to monkey faces appears at around 9 months (Pascalis et al., Citation2002; Pascalis & Kelly, Citation2009), and this preference for human faces can be reversed if babies are exposed to monkey faces (Pascalis et al., Citation2005). This narrowing of face perception to specialize in only our own species has been shown across childhood (Pascalis et al., Citation2005) and into adulthood (Dufour et al., Citation2004; Phelps & Roberts, Citation1994). It was also reported that training and/or experience with a species increases recognition performance for non-human animals (Dufour & Petit, Citation2009; Ueno et al., Citation2021), but mixed evidence was found for short exposure prior to testing (Dufour et al., Citation2004). However, these studies do not distinguish between familiar and unfamiliar human faces, and the majority have used memory or categorization paradigms. It is therefore unknown whether unfamiliar faces are processed differently to faces of a different species in a task which requires identity processing without memory, such as face matching.
Here we pitted human familiar and unfamiliar faces against monkey unfamiliar faces in a face matching task. Each trial showed two different images of either a familiar person, an unfamiliar person, or a monkey side-by-side, and participants were asked to judge whether the images show the same person (or monkey) or two different people (or monkeys).
Method
Participants
Three hundred and sixty-six participants from the UK (258 female, 107 male, 1 other; mean age: 30 years; age range: 18–74 years; 351 self-reported as White) took part in an online face matching task. Sample size exceeded previous tests of face matching (e.g., 60 in experiment 1 and 50 in experiment 2 of Fysh & Bindemann, Citation2018). All gave informed consent and ethics was granted from the University of Lincoln Research Ethics Committee (UoL2020_3814).
Stimuli and procedure
The task consisted of 28 pairs of faces for each face type: familiar humans, unfamiliar humans, and monkeys. Half of the identity pairs for each face type depicted female faces, half male. All human stimuli depicted faces of different White ethnic backgrounds. Familiar faces showed celebrities chosen to be familiar in the UK (participants’ mean familiarity M = 25 identities), and unfamiliar faces showed local celebrities from other countries, selected to be unfamiliar to participants in the UK (participants’ mean familiarity M = 1 identity). The monkey images showed Barbary macaques (Macaca sylvanus) from author LM used in previous research (Clark et al., Citation2020). For each face type (familiar, unfamiliar, monkey) images were ambient images showing natural variability, cropped to show just the head (see ). Ambient images better represent how we encounter faces in real life than posed images, and have been used in previous face matching studies (e.g., Noyes et al., Citation2021; Ritchie et al., Citation2015, Citation2020). Half the trials for each face type showed the same identity (match) and half showed two different identities (mismatch). Foil identities for mismatch trials were selected to fit the same verbal description e.g., “woman, dark hair”. All monkey foils came from the same image set as the target monkey faces, and all human foils pictured an unfamiliar celebrity (so as to keep image quality consistent between match and mismatch trials).
Figure 1. Example match trials from the human (left) and monkey (right) face matching tasks.
Note. Copyright restrictions prevent publication of the human face images used in the experiment. Human face images are illustrative of the experimental stimuli and depict someone who did not appear in the experiment but has given permission for the images to be reproduced here.

Each identity was seen once always in the same condition (match or mismatch), therefore there were 28 trials per face type, with a total of 84 trials. The experiment was blocked by face type, with the order of identities within each block randomized, and the order of blocks counterbalanced between participants. There was no time limit for responses on each trial, and response time was not recorded.
Results
We analysed match and mismatch trials separately (see Megreya & Burton, Citation2006). In addition to traditional frequentist hypothesis testing, we included Bayes factors using JASP (JASP Team, Citation2020), which allowed us to quantify the extent to which the data support the alternative hypothesis (BF10). A one way analysis of variance (ANOVA) on match trials showed a significant main effect of face type (p < 0.001, BF10 too large to report (infinity symbol reported in JASP)) with Bonferroni corrected pairwise comparisons showing that performance was better with familiar humans (M = 93.75%, SD = 7.81%) than both unfamiliar humans (M = 58.70%, SD = 17.52%) and monkeys (M = 56.85%, SD = 16.01%, both p < 0.001, both BF10 > 1 × 10125), but the comparison between unfamiliar humans and monkeys was non-significant (p > 0.999, BF10 = 0.26, see ). A Bayes factor of 1/3 (0.333) can be interpreted as anecdotal evidence for the null hypothesis (Andraszewicz et al., Citation2015). A one-sample t-test comparing monkey matching performance to chance (50%) showed that participants could perform the task with above chance accuracy t(365) = 8.18, p < 0.001, BF10 = 1.060 × 1012.
Figure 2. Data for match and mismatch trials.
Note. Error bars show standard error of the mean (SEM). *** p < 0.001.
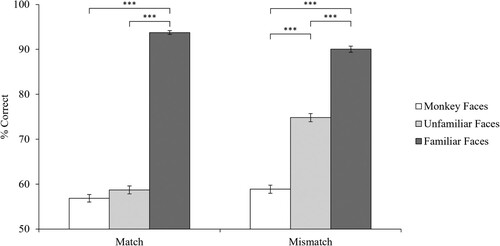
The ANOVA for mismatch trials showed a main effect of face type (p < 0.001, BF10 too large to report (infinity symbol reported in JASP)) with Bonferroni corrected pairwise comparisons showing that performance was best with familiar humans (M = 90.05%, SD = 13.16%), followed by unfamiliar humans (M = 74.82%, SD = 17.45%), and poorest with monkeys (M = 58.88%, SD = 17.01%, all p < 0.001, all BF10 > 1 × 1032).
Discussion
Our data show that for match trials, people were no better at matching two images of an unfamiliar person than two images of an unfamiliar monkey. People were more accurate on mismatch trials for unfamiliar humans than monkeys, and most accurate on both trial types with familiar human faces, as would be expected (see e.g., Bruce et al., Citation2001; Ritchie et al., Citation2015). We know from the face sorting literature (e.g., Jenkins et al., Citation2011) that people are reasonably good at telling unfamiliar people apart, but have trouble cohering multiple images of one person together as one identity (“telling people together”). Our results show that people can tell other humans apart better than they can monkeys, but that when it comes to telling people together, the key aspect of identity processing that differs for familiar and unfamiliar faces, they are just as poor with unfamiliar faces as they are with monkeys. Our results therefore argue against general face expertise, and suggest that unfamiliar faces might as well be a different species, at least in terms of determining whether two images depict the same identity. Our data show that humans were able to perform the monkey face matching test with above chance-level accuracy, perhaps not surprising since previous research has shown that humans can remember monkey faces with above chance accuracy (Phelps & Roberts, Citation1994), and can even recognize kin (relatedness) in several non-human primate species (Alvergne et al., Citation2009).
Accuracy on the matching trials of our unfamiliar face matching task was lower (M = 59%) than in some standard tests of unfamiliar face matching (GFMT2 Form A M = 74%, Form B M = 74%; KFMT M = 65%; Fysh & Bindemann, Citation2018; White et al., Citation2022). This could potentially be due to our use of ambient images which may produce more difficult matching tests. For example some of our previous studies using ambient images have produced similarly low mean match performance of 62% (experiment 3 younger participants, Ritchie et al., Citation2015) 63% (incongruent eyewear conditions, Graham & Ritchie, Citation2019). Taking both match and mismatch accuracy, however, mean accuracy on our unfamiliar face matching test here (M = 66%) is no different to that of the KFMT (M = 66%, Fysh & Bindemann, Citation2018). Future research should seek to replicate the effects reported here using different stimulus sets.
Research using different methods such as memory paradigms (Phelps & Roberts, Citation1994) and fixation duration measurements (Pascalis & Bachevalier, Citation1998) have shown that humans have better memory and longer fixation durations for human faces compared to monkey faces. Our results are partially consistent with this species-specific bias as we found higher accuracy on mismatch trials with unfamiliar human faces than monkey faces. Interestingly, Phelps and Roberts (Citation1994) also found that monkeys had better memory for human faces than monkey faces (no own-species bias), whereas Pascalis and Bachevalier (Citation1998) found that monkeys had longer fixation durations for monkey faces (own-species bias). Future research ought to test monkeys on a face matching task like ours in order to establish whether monkeys show an own-species bias on this task.
There is evidence of group biases in face memory with images of human faces whereby members of one’s own group are remembered more accurately than members of other groups (Bernstein et al., Citation2007; Malpass & Kravitz, Citation1969; c.f. Fuller et al., Citation2021). There is, however, mixed evidence of group biases in face matching tasks. Studies which have found group differences typically report performance with other-group faces of 60–86% (Robertson et al., Citation2020; Susa et al., Citation2019). Here performance with our monkey faces was 58% (match and mismatch trials combined) which is at the lower end of the human other-group face matching performance.
Previous research on inter-species face recognition has suggested that our face perception abilities narrow during development to specialize in only our own species (Dufour et al., Citation2004; Phelps & Roberts, Citation1994; Pascalis et al., Citation2001). These studies, however, do not distinguish between familiar and unfamiliar human faces, a distinction which we know from the face perception literature to be important (e.g., Bruce et al., Citation2001; Ritchie et al., Citation2015). Here, we once again highlight the importance of considering familiar and unfamiliar face perception separately, as unfamiliar faces are matched only as well as faces of another species. One previous paper found that monkey experts were no more accurate than non-experts at recognizing the faces of unfamiliar monkeys (Dufour & Petit, Citation2009). Future research, therefore, ought to include familiar monkeys to explore the role of familiarity in other species.
Open practices statement
The experiment was not preregistered. Data are available in the Supplemental Online Material.
Acknowledgements
The authors would like to thank students in the Research Skills III groups for data collection.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplementary material is available via the ‘Supplementary’ tab on the article’s online page (http://dx.doi.org/10.1080/13506285.2023.2184894).
References
- Alvergne, A., Huchard, E., Caillaud, D., Charpentier, M. J., Setchell, J. M., Ruppli, C., Raymond, M. (2009). Human ability to recognize kin visually within primates. International Journal of Primatology, 30(1), 199–210. https://doi.org/10.1007/s10764-009-9339-0
- Andraszewicz, S., Scheibehenne, B., Rieskamp, J., Grasman, R., Verhagen, J., & Wagenmakers, E. J. (2015). An introduction to Bayesian hypothesis testing for management research. Journal of Management, 41(2), 521–543. https://doi.org/10.1177/0149206314560412
- Bernstein, M. J., Young, S. G., & Hugenberg, K. (2007). The cross-category effect: Mere social categorization is sufficient to elicit an own-group bias in face recognition. Psychological Science, 18(8), 706–712. https://doi.org/10.1111/j.1467-9280.2007.01964.x
- Bruce, V. (1994). Stability from variation: The case of face recognition the M.D. Vernon memorial lecture. Quarterly Journal of Experimental Psychology, 47(1), 5–28. https://doi.org/10.1080/14640749408401141
- Bruce, V., Henderson, Z., Newman, C., & Burton, A. M. (2001). Matching identities of familiar and unfamiliar faces caught on CCTV images. Journal of Experimental Psychology: Applied, 7(3), 207–218. https://doi.org/10.1037/1076-898X.7.3.207
- Burton, A. M. (2013). Why has research in face recognition progressed so slowly? The importance of variability. The Quarterly Journal of Experimental Psychology, 66(8), 1467–1485. https://doi.org/10.1080/17470218.2013.800125
- Burton, A. M., White, D., & McNeill, A. (2010). The Glasgow face matching test. Behavior Research Methods, 42(1), 286–291. https://doi.org/10.3758/BRM.42.1.286
- Burton, A. M., Wilson, S., Cowan, M., & Bruce, V. (1999). Face recognition in poor-quality video: Evidence from security surveillance. Psychological Science, 10(3), 243–248. https://doi.org/10.1111/1467-9280.00144
- Clark, L., Butler, K., Ritchie, K. L., & Maréchal, L. (2020). The importance of first impression judgements in interspecies interactions. Scientific Reports, 10(1), 1–10. https://doi.org/10.1038/s41598-019-56847-4
- Davis, J. P., & Valentine, T. (2009). CCTV on trial: Matching video images with the defendant in the dock. Applied Cognitive Psychology, 23(4), 482–505. https://doi.org/10.1002/acp.1490
- Diamond, R., & Carey, S. (1986). Why faces are and are not special: An effect of expertise. Journal of Experimental Psyhcology General, 115(2), 107–117. https://doi.org/10.1037/0096-3445.115.2.107
- Dufour, V., Coleman, M., Campbell, R., Petit, O., & Pascalis, O. (2004). On the species-specificity of face recognition in human adults. Current Psychology of Cognition, 22, 315–333.
- Dufour, V., & Petit, O. (2009). Recognition of monkey faces by monkey experts. Journal of Ethnology, 28, 231–238. https://doi.org/10.1007/s10164-009-0174-8
- Fuller, E. A., Majolo, B., Flack, T. R., & Ritchie, K. L. (2021). The importance of out-group characteristics for the own-group face memory bias. Visual Cognition, 29(4), 263–276. https://doi.org/10.1080/13506285.2021.1905125
- Fysh, M. C., & Bindemann, M. (2018). The Kent face matching test. British Journal of Psychology, 109(2), 219–231. https://doi.org/10.1111/bjop.12260
- Graham, D. L., & Ritchie, K. L. (2019). Making a spectacle of yourself: The effect of glasses and sunglasses on face perception. Perception, 48(6), 461–470. https://doi.org/10.1177/0301006619844680
- JASP Team. (2020). JASP (Version 0.14) [Computer software].
- Jenkins, R., White, D., Van Montfort, X., & Burton, A. M. (2011). Variability in photos of the same face. Cognition, 121(3), 313–323. https://doi.org/10.1016/j.cognition.2011.08.001
- Longmore, C. A., Liu, C. H., & Young, A. W. (2008). Learning faces from photographs. Journal of Experimental Psychology: Human Perception and Performance, 34(1), 77–100. https://doi.org/10.1037/0096-1523.34.1.77
- Longmore, C. A., Santos, I. M., Silva, C. F., Hall, A., Faloyin, D., & Little, E. (2017). Image dependency in the recognition of newly learnt faces. Quarterly Journal of Experimental Psychology, 70(5), 863–873. https://doi.org/10.1080/17470218.2016.1236825
- Malpass, R. S., & Kravitz, J. (1969). Recognition for faces of own and other race. Journal of Personality and Social Psychology, 13(4), 330–334. https://doi.org/10.1037/h0028434
- Matthews, C. M., Davis, E. E., & Mondloch, C. J. (2018). Getting to know you: The development of mechanisms underlying face learning. Journal of Experimental Child Psychology, 167, 295–313. https://doi.org/10.1016/j.jecp.2017.10.012
- Megreya, A. M., & Burton, A. M. (2006). Unfamiliar faces are not faces: Evidence from a matching task. Memory & Cognition, 34(4), 865–876. https://doi.org/10.3758/BF03193433
- Murphy, J., Ipser, A., Gaigg, S., & Cook, R. (2015). Exemplar variance supports robust learning of facial identity. Journal of Experimental Psychology: Human Perception and Performance, 41(3), 577–581. https://doi.org/10.1037/xhp0000049
- Noyes, E., Davis, J. P., Petrov, N., Gray, K. L., & Ritchie, K. L. (2021). The effect of face masks and sunglasses on identity and expression recognition with super-recognizers and typical observers. Royal Society Open Science, 8(3), 201169. https://doi.org/10.1098/rsos.201169
- Pascalis, O., & Bachevalier, J. (1998). Face recognition in primates: A cross-species study. Behavioural Processes, 43(1), 87–96. https://doi.org/10.1016/S0376-6357(97)00090-9
- Pascalis, O., de Haan, M., & Nelson, C. A. (2002). Is face processing species-specific during the first year of life? Science, 296(5571), 1321–1323. https://doi.org/10.1126/science.1070223
- Pascalis, O., Demont, E., de Haan, M., & Campbell, R. (2001). Recognition of faces of different species: A developmental study between 5 and 8 years of age. Infant and Child Development: An International Journal of Research and Practice, 10(1-2), 39–45. https://doi.org/10.1002/icd.245
- Pascalis, O., & Kelly, D. J. (2009). The origins of face processing in humans: Phylogeny and ontogeny. Perspectives on Psychological Science, 4(2), 200–209. https://doi.org/10.1111/j.1745-6924.2009.01119.x
- Pascalis, O., Scott, L. S., Kelly, D. J., Shannon, R. W., Nicholson, E., Coleman, M., & Nelson, C. A. (2005). Plasticity of face processing in infancy. Proceedings of the National Academy of Sciences, 102(14), 5297–5300. https://doi.org/10.1073/pnas.0406627102
- Phelps, M. T., & Roberts, W. A. (1994). Memory for pictures of upright and inverted primate faces in humans (Homo sapiens), squirrel monkeys (Saimiri sciureus), and pigeons (Columba livia). Journal of Comparative Psychology, 108(2), 114–125. https://doi.org/10.1037/0735-7036.108.2.114
- Ritchie, K. L., & Burton, A. M. (2017). Learning faces from variability. Quarterly Journal of Experimental Psychology, 70(5), 897–905. https://doi.org/10.1080/17470218.2015.1136656
- Ritchie, K. L., Mireku, M. O., & Kramer, R. S. S. (2020). Face averages and multiple images in a live matching task. British Journal of Psychology, 111(1), 92–102. https://doi.org/10.1111/bjop.12388
- Ritchie, K. L., Smith, F. G., Jenkins, R., Bindemann, M., White, D., & Burton, A. M. (2015). Viewers base estimates of face matching accuracy on their own familiarity: Explaining the photo-ID paradox. Cognition, 141, 161–169. https://doi.org/10.1016/j.cognition.2015.05.002
- Robertson, D. J., Black, J., Chamberlain, B., Megreya, A. M., & Davis, J. P. (2020). Super-recognisers show an advantage for other race face identification. Applied Cognitive Psychology, 34(1), 205–216. https://doi.org/10.1002/acp.3608
- Susa, K. J., Gause, C. A., & Dessenberger, S. J. (2019). Matching faces to ID photos: The influence of motivation on cross-race identification. Applied Psychology in Criminal Justice, 15(1), 86–96. http://www.apcj.org/journal/index.php?mode=view&item=151
- Ueno, M., Yamamoto, H., Yamada, K., & Itakura, S. (2021). Individual recognition of monkey (Macaca fuscata) and human (Homo sapiens) images in primatologists. Journal of Comparative Psychology, 135(3), 394–405. https://doi.org/10.1037/com0000285
- White, D., Guilbert, D., Varela, V. P., Jenkins, R., & Burton, A. M. (2022). GFMT2: a psychometric measure of face matching ability. Behavior Research Methods, 54(1), 252–260. https://doi.org/10.3758/s13428-021-01638-x
- White, D., Kemp, R. I., Jenkins, R., Matheson, M., & Burton, A. M. (2014). Passport officers’ errors in face matching. PLoS ONE, 9(8), e103510. https://doi.org/10.1371/journal.pone.0103510
- Young, A. W., & Burton, A. M. (2018). Are we face experts? Trends in Cognitive Sciences, 22(2), 100–110. https://doi.org/10.1016/j.tics.2017.11.007