
ABSTRACT
According to a longstanding view, sex differences in cognitive abilities such as mental rotation or verbal memory arise from sex differences in hemispheric asymmetry: males are thought to be more lateralized than females which boosts their spatial but hampers their verbal skills. This idea sparked great interest and, even though it lost support in the 1990s, it is still put forward in contemporary (popular) scientific papers and textbooks. We aimed to provide a comprehensive review that summarizes the last 40 years of research. First, we confirm previous findings that the stronger hemispheric asymmetry in males is very small but robust. Second, we conclude that stronger hemispheric asymmetry, in general, does not enhance spatial and reduce verbal performance. Crucially, we carried out a systematic literature review showing that cognitive sex differences often emerge in the absence of sex differences in hemispheric asymmetry (and vice versa), implying the two phenomena are at least partly independent of each other. At present, there is insufficient data to conclude that sex differences in hemispheric asymmetry and cognitive performance are uncorrelated. However, we can conclude that sex differences in hemispheric asymmetry are certainly not the driving force behind sex differences in cognitive functioning.
A meta-synthesis (i.e., a meta-analysis of meta-analyses) based on 12 million participants revealed that the male advantage in mental rotation is the second largest sex difference in the psychological literature (Zell, Krizan, & Teeter, Citation2015) – only topped by males rating themselves more masculine than females. Other, smaller cognitive sex differences with – on average – better female or better male performance are also well documented (for review Halpern, Citation2012; Kimura, Citation2000). In general, these sex differences are the exception rather than the rule: In most cognitive tasks males and females perform equally well and when sex differences emerge there is substantial overlap in cognitive performance (Hyde, Citation2005, Citation2014). However, even small effects can be highly relevant if, for instance, sex-sensitive tasks are utilized in job recruitment (Rosenthal & Rubin, Citation1982). Moreover, cognitive sex differences have been suggested to be one of many reasons why females are underrepresented in science, technology, engineering, and mathematics (for a comprehensive review see Ceci, Williams, & Barnett, Citation2009) and why boys and girls should be taught in single-sex environments (but see the review by Halpern et al., Citation2011). The study of cognitive sex differences is thus highly relevant for both the individual and for the society at large.
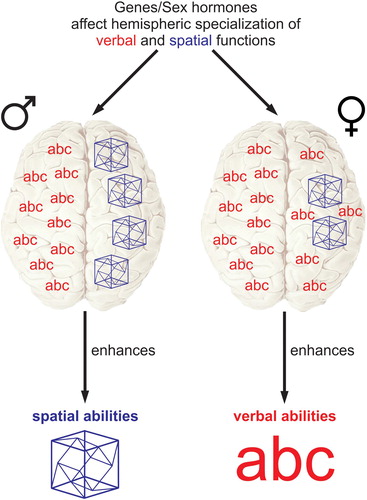
There is consensus that the origins of cognitive sex differences are a complex mixture of nature and nurture (Miller & Halpern, Citation2014), but the underlying neural mechanisms are still unknown. In the 1970s a popular idea was put forward according to which cognitive sex differences arise from a sex difference in brain asymmetry. Specifically, Jerre Levy (Citation1972, Citation1978) proposed that males have a more asymmetric brain organization where the left hemisphere would be clearly specialized for verbal processing and the right hemisphere for spatial processing. In females, the brain would be more “bilateral”, that is, both the left and the right hemisphere would be carrying out verbal processing. As a consequence, the more asymmetric, male brain would be superior for spatial skills and the more bilateral, female brain would be superior for verbal skills (see an illustration in ). Several similar theories have been put forward, all sharing the basic idea, namely that sex differences in cognitive abilities arise from sex differences in hemispheric asymmetry. This idea became very popular: Numerous studies examined the relationship between sex, hemispheric asymmetry, and cognitive abilities; popular science and textbooks frame it as a very likely explanation (e.g., Kimura, Citation2000; Springer & Deutsch, Citation1993) and although the enthusiasm has declined over the years, the idea features in contemporary articles about cognitive sex differences (Grabowska, Citation2017; Pletzer, Citation2014). We aim to provide a critical, comprehensive review that summarizes the empirical evidence. As a starting point, we take Levy’s original theory (henceforth referred to as Levy’s hypothesis), since it is by far the most popular variant, but as will hopefully become clear during this review our conclusions apply to the general idea that sex differences in cognitive abilities arise from hemispheric asymmetry.
Figure 1. Simplified model of Levy’s hypothesis. Males have stronger hemispheric specialization for verbal and spatial abilities than females. The stronger specialization enhances spatial abilities while the more bilateral organization enhances verbal abilities.
We think such a review is timely for two reasons. First, earlier critique has been insufficient to dismiss the idea. In the 1990s and 2000s behavioural and neuroimaging data accrued showing that the stronger male lateralization is very small, if any (for a more detailed discussion see section “Do Males Have More Asymmetric Brains?” below). This led some researchers to question Levy’s hypothesis because it seemed improbable that such a small sex difference could account for the sizeable male advantage in mental rotation (e.g., see p. 238 in Halpern, Citation2012). We share the critique but even though it seems unlikely, it is not enough to disprove Levy’s hypothesis. First, there are examples where small sex differences turn out to be a tip of the iceberg. For instance, Del Giudice, Booth, and Irwing (Citation2012) demonstrated how sex differences in personality that were believed to be small and insignificant are in fact quite large when appropriate methodology and statistical tools are used. Secondly, there are examples where a small sex difference in the brain can have far-reaching behavioural consequences. For example, the third interstitial nucleus of the anterior hypothalamus (INAH-3) is “a subdivision of a subdivision of the medial preoptic nucleus, itself a relatively small brain region” (McCarthy, Citation2016, p. 3) with an overall volume of approximately a tenth of a mm (Garcia-Falgueras & Swaab, Citation2008). It is, however, consistently larger in males than in females and by many believed to play a crucial role for sexual identity and, perhaps, also sexual orientation (for review Balthazart, Citation2016; McCarthy, Citation2016). Thus, in order to falsify Levy’s hypothesis, it is insufficient to look at the magnitude of the sex difference in hemispheric asymmetry. It is also necessary to review the empirical evidence for whether sex differences in cognitive performance depend on sex differences in hemispheric asymmetry and how hemispheric asymmetry and cognitive performance are associated in males and females, in general. This, to our knowledge, has not been done so far.
Secondly, the introduction of functional neuroimaging techniques, like positron emission tomography (PET) and especially functional magnetic resonance imaging (fMRI), allowed for more direct assessment of hemispheric asymmetry, putting us now in a better position to test Levy’s hypothesis directly. For example, a number of large-scale neuroimaging studies have addressed sex differences in hemispheric asymmetry (e.g., Guadalupe et al., Citation2015; Hirnstein, Westerhausen, Korsnes, & Hugdahl, Citation2013; Plessen, Hugdahl, Bansal, Hao, & Peterson, Citation2014; Sommer, Aleman, Somers, Boks, & Kahn, Citation2008) and how hemispheric asymmetry relates to cognitive performance (e.g., Gotts et al., Citation2013; Mellet, Zago, et al., Citation2014). In pre-neuroimaging times, Levy’s hypothesis was typically investigated by using handedness as an indicator for laterality (e.g., Harshman, Hampson, & Berenbaum, Citation1983; Sherman, Citation1979) but as we will see below this approach has limitations.
We have divided this review into six parts. First, we describe the origin and background of the Levy idea that sex differences in hemispheric asymmetry give rise to cognitive sex differences. Then, we examine the empirical evidence for four underlying assumptions: (1) Do males and females really have better spatial and verbal skills, respectively? (2) Do males have a more asymmetric brain organization than females? (3) Does stronger hemispheric asymmetry, in general, promote spatial but impede verbal performance? (4) Are cognitive sex differences dependent upon sex differences in hemispheric asymmetry? Finally, we provide suggestions for future research (a) how the links between hemispheric asymmetry, sex, and cognitive performance can be unravelled and (b) how better models that seek to explain the underlying neural mechanisms of cognitive sex differences can be developed.
Background and origins of Levy’s hypothesis
All major cognitive, sensory and motor functions, including language, attention, learning, memory, perception, emotional processing, tool use, and fine motor control involve the processing capacities of both hemispheres. To some extent, however, these functions are “lateralized”. That is, one hemisphere is specialized and plays a dominant role (for review Davidson & Hugdahl, Citation1995; Hellige, Citation1993; Ocklenburg & Güntürkün, Citation2018) relative to the other hemisphere. This is collectively called “hemispheric asymmetry”, “hemispheric specialization” or “lateralization”. Those functional hemispheric asymmetries can also be found on structural and neurochemical level, making lateralization a fundamental principle of human brain organization (Rogers, Citation2014).
The best documented asymmetry is the left-hemispheric specialization for language, first described by Dax (Citation1836), which was subsequently corroborated in numerous studies in both the intact and lesioned brain (Hellige, Citation1993). At around the same time, Jackson (Citation1864; as cited by Bogen & Gazzaniga, Citation1965) was among the first to suggest that the right hemisphere may be specialized for perception and, indeed, the experiments with split-brain patients led by Roger Sperry (Gazzaniga, Bogen, & Sperry, Citation1962; Sperry, Citation1968) confirmed a right-hemispheric dominance for visuospatial processing (Bogen & Gazzaniga, Citation1965). Jerre Levy then asked the question whether it would be detrimental to spatial abilities, if the right-hemisphere also hosted verbal functions. Assuming that left-handers would represent a group with such a bilateral language representation, she tested 15 right- and 10 left-handers (all were students at the California Institute of Technology) with the Wechsler Adult Intelligence Scale (Levy, Citation1969). The left-handers had a slightly higher verbal intelligence quotient (IQ) than right-handers (142 versus 138), while right-handers had a higher performance IQ (140 versus 117). Levy reasoned that because left-handers have verbal functions in both hemispheres there would be less neural space for spatial functions in the right-hemisphere. In other words, the verbal functions would “crowd” the right hemisphere – hence, her theory became later known as the “crowding hypothesis”. In right-handers, however, there would be a clear division of labour (i.e., left-hemisphere = verbal functions, right-hemisphere = spatial functions), and this more asymmetric organization would be superior for spatial abilities.
The final step was to apply the crowding hypothesis to males and females. Lansdell (Citation1961) had previously suggested that females had a more bilateral language representation than males, based on unilateral lesion studies. Moreover, Levy was aware of findings showing superior spatial abilities in males (MacFarlane Smith,, Citation1964). She concluded: “It is hard to reject the notion that a spatial-perceptive deficit in women is a sex-linked, genetically determined incapacity, an incapacity which possibly results from hemispheres less well laterally specialized than those of males” (Levy, Citation1972, p. 174). Similarly, Witelson (Citation1976) conducted a study where children aged 6-13 yrs were asked to identify shapes out of view with either their left or right hand. She found that boys showed a consistent left-hand advantage, which she interpreted as indicative of a right-hemispheric bias in spatial processing, while girls did not show any hand effect and suggested: “The superiority of males to females on many, although not all, spatial tests [ … ] may be related to the hypothesized neural dimorphism [in hemispheric asymmetry]” (Witelson, Citation1976, p. 426). A re-analysis of Levy’s (Citation1969) handedness data revealed significantly higher verbal IQ in left-handers as compared to right-handers (Levy, Citation1974), leading Levy (Citation1978) to conclude that the more bilaterally lateralized verbal functions in females would give rise to superior verbal abilities.
Levy’s hypothesis is the most popular but not the only attempt to explain cognitive sex differences with hemispheric asymmetry. For example, at around the same time Buffery and Gray (Citation1972) argued that females, in contrast to males, have a more asymmetric brain organization. Specifically, the left-hemispheric language specialization would develop earlier in females and facilitate verbal skills, while the more bilateral organization in males would be beneficial for spatial skills. The Buffery and Gray hypothesis was quickly dismissed on the grounds that, if a sex difference in hemispheric specialization emerged, it typically showed stronger lateralization in males (McGlone, Citation1980). Waber (Citation1976) also linked earlier maturation in girls to sex differences in hemispheric asymmetry and cognitive abilities: She reported that early maturing individuals performed better at verbal tasks while late maturing individuals performed better at spatial tasks. The late maturing individuals also had stronger hemispheric asymmetry, as assessed with dichotic listening, which is in line with Levy’s claim that stronger lateralization enhances spatial skills. These differences between early and late maturing individuals, however, were regardless of sex, leading Waber to conclude that the sex difference in cognitive tasks mediated by hemispheric asymmetry is not so much about sex but rather about differences in maturation. Finally, Levy’s hypothesis is often tied to the idea that females have larger corpora callosa than males – relative to body size – which would facilitate interhemispheric transfer and result in a more bilateral brain organization in females and a more asymmetric organization in males (e.g., Grabowska, Citation2017). Thus, Levy’s hypothesis is part of a larger framework that tied hemispheric asymmetry to sex differences in cognitive abilities.
This framework grew further in popularity when neurobiological theories emerged in the 80s that sought to explain what could cause sex differences in hemispheric asymmetry. For instance, it was suggested that testosterone has a masculinizing effect which “lateralizes” the brain (Hines & Gorski, Citation1985, 1985; see also Smith & Hines, Citation2000) and that testosterone reduces interhemispheric connectivity via the corpus callosum leading to increased lateralization (Witelson, Citation1991). The most influential theory at that time (Geschwind & Galaburda, Citation1985) posited that testosterone slows down the maturation of the left hemisphere leading to a relatively more developed right-hemisphere and, thus, superior spatial and inferior verbal abilities in males. Note that contrary to Levy’s hypothesis the Geschwind & Galaburda theory would also predict males to be less left-lateralized for language than females. None of the theories outlined above, however, has received sufficient empirical support (Bryden, McManus, & Bulman-Fleming, Citation1994; Pfannkuche, Bouma, & Groothuis, Citation2009). Several studies reported an association between sex hormones and hemispheric asymmetry (for review Hausmann, Citation2017; Hausmann & Bayer, Citation2010; Pfannkuche et al., Citation2009). However, as a consequence of inconsistent findings, we still do not know exactly, how sex hormones are involved in orchestrating the functional and anatomical specialization of the two hemispheres (for review Hausmann, Citation2017; Hausmann & Bayer, Citation2010; Pletzer, Citation2014).
A first critical look
Before we turn to the empirical support for Levy’s hypothesis, already at this stage a few critical issues can be raised. First of all, it is important to bear in mind that Levy was mostly interested in the evolution of hemispheric asymmetry, not the origins of cognitive sex differences. The idea that the two phenomena could be linked is mentioned “en passant” in a book chapter on what advantages an asymmetric brain might have bestowed on humans, seen in an evolutionary perspective. Secondly, she did not directly test the association between sex differences in hemispheric asymmetry and cognitive performance since all participants were male (Levy, Citation1969), but given the scientific knowledge and zeitgeist at that time her conclusions were plausible and logically sound. From what we know today, the data in the Levy (Citation1969) paper have been over-interpreted: Levy took the higher verbal and lower spatial performance of left-handers as evidence for the notion that a bilateral brain enhances verbal skills (and hampers spatial skills). This rests on the idea that left-handers are more bilaterally organized than right-handers. However, this is only partly true. Left-handers are more likely to have a bilateral language representation (ca. 15%) than right-handers (ca. 4%), but the majority of both left- (ca. 70%) and right-handers (ca. 96%) are left lateralized – the remaining 15% left-handers have rightward language lateralization (Rasmussen & Milner, Citation1977; for review Carey & Johnstone, Citation2014). If one applies those numbers to Levy’s sample (Levy, Citation1969), then only one or two left-handers (i.e., 15%, n = 10) should have had bilaterally organized brains. Could this explain a significant mean difference of 23 IQ points between left- and right-handers? Obviously, bilateral brain organization is far less common in left-handers than Levy assumed; suggesting factors other than hemispheric asymmetry must have been behind the observed handedness effect. Another important lesson from the distribution of language lateralization in left- and right-handers is that handedness is only a very coarse proxy for hemispheric asymmetry (for review Ocklenburg, Beste, Arning, Peterburs, & Güntürkün, Citation2014). For example, Van der Haegen, Westerhausen, Hugdahl, and Brysbaert (Citation2013) found that on group level language dominance (as assessed with fMRI) is more reliably predicted by dichotic listening than by handedness. Thus, studies aiming to verify or falsify Levy’s hypothesis should employ more direct lateralization assessment methods.
Nevertheless, these shortcomings do not directly contradict the possibility that a more bilateral female brain organization enhances verbal performance, and a more asymmetric male brain organization enhances spatial performance.
Do cognitive sex differences exist?
Before discussing the role of hemispheric asymmetry, it needs to be established first whether males actually outperform females and females outperform males in spatial and verbal tasks, respectively. As pointed out above, the largest and most researched cognitive sex difference is observed in mental rotation. Myriads of studies and meta-analyses unanimously reveal a male advantage typically between Cohen’s d = 0.56 (Voyer, Voyer, & Bryden, Citation1995; Zell et al., Citation2015) and d = 0.73 (Linn & Petersen, Citation1985). The most widespread mental rotation test was developed by Vandenberg and Kuse (Citation1978; for a revised version Peters et al., Citation1995) and comprises 3-dimensional cube figures designed by Shepard and Metzler (Citation1971). Better male performance has also been documented in a group of tasks that assesses “understanding abstract spatial principles, such as horizontal invariance or verticality” (Uttal et al., Citation2013, p. 355) or in the terminology of Linn and Petersen (Citation1985) spatial perception. Typical spatial perception tasks are the Water Level Test in which participants draw a line in tilted bottles to indicate the (horizontal) water orientation (Vasta & Liben, Citation1996) or the Benton Line Orientation task in which participants have to match lines with different angles (Benton, Varney, & Hamsher, Citation1978). The male advantage lies between d = 0.48 (Voyer et al., Citation1995) and d = 0.64 (Linn & Petersen, Citation1985). Males also fare slightly better on tasks that require “perceiving objects, paths, or spatial configurations amid distracting background information” (Uttal et al., Citation2013, p. 355). For instance, in the Embedded Figures Test an object like a triangle needs to be found within a larger, complex figure (Witkin, Oltman, Raskin, & Karp, Citation1971). Sometimes, objects also need to be mentally transformed like in the Paper Folding Test, where participants are asked to imagine what cube figures that are flattened out would look like if they were folded (e.g., Harshman et al., Citation1983). However, the male advantage in this group of tasks, sometimes labelled spatial visualization (Linn & Petersen, Citation1985), is small with d = 0.13 to d = 0.19 (Linn & Petersen, Citation1985; Voyer et al., Citation1995). Finally, males appear to perform better in mechanical reasoning, which typically involves predicting the behaviour of pulleys or gears (d = 0.76; Feingold, Citation1988). However, at present there is too little data available and this group of tasks might simply represent a special case of spatial visualization. For an illustration of some of the tasks that yield cognitive sex differences, see .
Figure 2. Overview of cognitive sex differences. Tasks favouring males (A) Redrawn Vandenberg and Kuse Mental Rotation Test: Which of the four stimuli are identical but rotated versions of the target figure? (with permission from Peters et al., Citation1995) (B) Paper folding test: Do the two arrows meet when the cube is folded? (adapted from Shepard & Feng, Citation1972) (C) Water level test: A line representing the water level needs to be drawn into the empty jar. Tasks favouring females (D) Object location memory test: Which items have changed its position? (adapted from Silverman & Eals, Citation1992) (E) Verbal fluency: Generate as many words as possible that start with the letter “F” or “A”. (F) Verbal memory: How many words from a list can be recalled?
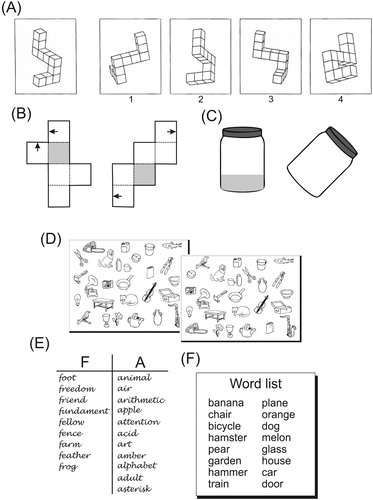
Although, in general, males appear to have better spatial abilities, at least one task that is often deemed to be spatial reveals a female advantage: object location memory. In a typical object location memory task participants are asked to remember the identity and/or position of multiple objects on a sheet of paper (e.g., Silverman & Eals, Citation1992) or computer display. According to a meta-analysis by Voyer, Postma, Brake, and Imperato-McGinley (Citation2007), there is a small but consistent female advantage (d = 0.27), which is dependent, however, on a number of factors including the participants’ age and the type of object to be memorized. Moreover, there are claims that females excel in this task because they encode the objects (and their position) verbally. That is, the female advantage may not necessarily reflect better spatial but rather better verbal memory (Choi & L’Hirondelle, Citation2005; Eals & Silverman, Citation1994).
Indeed, a female advantage in verbal memory is well-documented. For instance, when asked to retrieve a list of words, females outperform males by typically about d = 0.30 (e.g., Bleecker, Bolla-Wilson, Agnew, & Meyers, Citation1988; Herlitz, Reuterskiöld, Lovén, Thilers, & Rehnman, Citation2013; Kramer, Delis, Kaplan, Odonnell, & Prifitera, Citation1997; Lowe, Mayfield, & Reynolds, Citation2003). A similar female advantage in terms of magnitude (d = 0.33; Hyde & Linn, Citation1988) has been reported for verbal fluency – typically tested by asking participants to generate as many words as possible that fulfil a certain criterion (e.g., words starting with the letter “P”). Finally, females appear to excel in tasks related to reading achievement. Although previous meta-analyses found rather small female advantages (Hedges & Nowell, Citation1995; Hyde, Citation2005), the Programme for International Student Assessment (PISA) revealed that 15 year old girls consistently outperformed their male counterparts across 65 nations by an overall d = 0.44 (Reilly, Citation2012).
A huge and controversial topic is cognitive sex differences in mathematical abilities. For a comprehensive review we refer to the article by Ceci et al. (Citation2009). Our very general conclusion from several meta-analyses in this field is that (a) in school, females obtain better marks (Voyer & Voyer, Citation2014), (b) in aptitude tests, there are minor male advantages but sex differences are fairly small on average (Else-Quest, Hyde, & Linn, Citation2010; Hyde, Lindberg, Linn, Ellis, & Williams, Citation2008), (c) at the high end of mathematical performance, males are consistently overrepresented (Stoet & Geary, Citation2013). It has been suggested that the male advantage at the higher end of mathematical abilities are based on better spatial skills in males (Casey, Nuttall, & Pezaris, Citation1997; Geary, Citation1996).
So far, we focused on cognitive areas in which males and females differ and all those sex differences have been replicated repeatedly and are based on data from meta-analyses and large-scale studies (for more comprehensive reviews Halpern, Citation2012; Hyde, Citation2014; Kimura, Citation2000; Jäncke, Citation2018). Yet, the majority of cognitive skills, including spatial and verbal skills, do not show significant sex differences (Hyde, Citation2005, Citation2014) and we would like to emphasize that there is large consensus that there are no sex differences with respect to general intelligence (Halpern, Citation2012). The magnitude and direction of sex differences in those specific, cognitive tasks are also highly malleable: Changes over time (Wai, Cacchio, Putallaz, & Makel, Citation2010), training effects (e.g., Uttal et al., Citation2013), culture, and contextual factors such as solution strategies, task format, confidence, and time restrictions are some of the many variables that can alter the male or female advantage (for review Halpern, Citation2012; Jäncke, Citation2018). For example, several studies found that playing computer games more frequently is related to the male advantage in mental rotation (Feng, Spence, & Pratt, Citation2007; Terlecki & Newcombe, Citation2005). However, such sociocultural factors cannot explain all findings in the literature as a study by Lippa, Collaer, and Peters (Citation2010) illustrates: Males had on average higher mental rotation scores than females in all of the 53 countries where participants were tested – this is difficult to explain purely with socio-cultural factors. On the other hand, the magnitude of the male advantage varied substantially across those countries – this is difficult to explain purely with biological factors. According to Halpern’s “psychobiosocial model” (Citation2012; Halpern & Tan, Citation2001), a complex interplay of psychological, biological, and social factors gives rise to cognitive sex differences (see also Hausmann, Schoofs, Rosenthal, & Jordan, Citation2009).
Finally, one should also point out that, in general, males tend to show higher variability in performance than females both within a single cognitive task and across several cognitive tasks. That is, males are more likely to be found at the higher and lower end of performance in a single task and they are more likely to perform well in one cognitive task but poorly in another whereas the pattern is more consistent in females (e.g., Roalf et al., Citation2014). Despite all these caveats, it seems fair to conclude that – as Levy (Citation1972) did – if cognitive sex differences arise, they tend to be such that males excel in certain spatial and females in certain verbal tasks. The fact that many spatial and verbal tasks do not show any sex difference, however, illustrates that a clear male/spatial and female/verbal divide is an oversimplification.
Do males have more asymmetric brains?
The idea that females have a more bilateral brain organization than males dates back at least to 1879 when James Crichton-Browne weighed the brains of deceased asylum inmates and found that “[…] the tendency to symmetry in the two halves of the cerebrum is stronger in women than in men” (Crichton-Browne, Citation1879, p. 65). The aforementioned lesion study by Lansdell (Citation1961) sparked a great interest in sex differences in hemispheric asymmetry and 20 years later McGlone (Citation1980) concluded in an often-cited review article: “Nevertheless, there is an impressive accummulation of evidence suggesting that the male brain may be more asymmetrically organized than the female brain” (p. 215). More recent evidence, particularly from neuroimaging, however, is far more nuanced than textbooks (“Females are less lateralized than males”, Springer & Deutsch, Citation1993, p. 212) and popular science media (“Women use their whole brain, men just half of it”, Anitei, Citation2007) suggest.
Myriads of studies investigated sex differences in hemispheric asymmetry and numerous studies revealed stronger asymmetry in males (Chance, Casanova, Switala, & Crow, Citation2006; Hausmann et al., Citation1998; Proverbio, Riva, Martin, & Zani, Citation2010; Shaywitz et al., Citation1995), although null-findings (Boles, Citation2005; Frost et al., Citation1999; Knecht et al., Citation2000), and even stronger asymmetry in females were also reported (Kaiser, Kuenzli, Zappatore, & Nitsch, Citation2007; Ladavas, Umiltà, & Ricci-Bitti, Citation1980). A first attempt to quantitatively review the empirical evidence was undertaken by Merrill Hiscock and colleagues in a series of studies from the 1990s. They counted how many studies showed stronger male, stronger female, or no asymmetry difference in the auditory (Hiscock, Inch, Jacek, Hiscockkalil, & Kalil, Citation1994), visual (Hiscock, Israelian, Inch, Jacek, & Hiscockkalil, Citation1995), and tactile modality (Hiscock, Inch, Hawryluk, Lyon, & Perachio, Citation1999) as well as dual task interference (Hiscock, Perachio, & Inch, Citation2001). Hemispheric asymmetry was mostly assessed behaviourally, that is, reaction times and accuracy rates derived from visual half-field tasks and dichotic as well as dichaptic stimulation (i.e., unilateral presentation of tactile stimuli). The authors concluded that there is “a small but reliable population-level sex difference [favoring males]” (Hiscock et al., Citation2001, p. 137). The first meta-analysis utilizing a more contemporary approach with effect sizes was carried out by Voyer (Citation1996). He too found stronger hemispheric asymmetry in males across a range of visual, auditory, and tactile laterality tasks that were assessed behaviourally. Again, while statistically significant the magnitude of this sex difference was very small, explaining less than 0.1% of variance. These findings were corroborated by a follow-up meta-analysis that only examined dichotic listening (Voyer, Citation2011).
As their numbers increased, neuroimaging findings were also included in meta-analyses. Vogel, Bowers, and Vogel (Citation2003), for example, specifically investigated the lateralization of spatial tasks based on both behavioural and neuroimaging findings. They found a significant right-hemispheric bias for males and no clear hemispheric specialization in females. Iris Sommer and colleagues (Sommer, Aleman, Bouma, & Kahn, Citation2004; Sommer et al., Citation2008) then further focused on neuroimaging findings in two meta-analyses on language lateralization: The first study examined functional neuroimaging methods, that is, functional transcranial Doppler Sonography (fTCD), fMRI, and PET (Sommer et al., Citation2004); the second study explored structural asymmetry of the planum temporale area and functional language asymmetry with dichotic listening (Sommer et al., Citation2008). Except for stronger male asymmetry in consonant vowel dichotic listening tasks, neither study yielded any significant sex differences. Sommer (Citation2010) concluded that sex differences in hemispheric asymmetry do not exist. She further noted that the small male bias reported by her and other meta-analyses (Sommer et al., Citation2008; Vogel et al., Citation2003; Voyer, Citation1996, Citation2011) could be the result of publication bias and that large-scale studies with sufficient power to detect subtle effects were missing (Sommer, Citation2010).
Such large-scale studies have been published in the meantime. More than 900 participants completed a smartphone application with a dichotic listening test (Bless et al., Citation2015). They were presented with two different consonant–vowel syllables, one in the right ear and the other simultaneously in the left ear (e.g., /ba/ to the left and /ta/ to the right ear). When instructed to report the sound they heard best or most clearly, participants typically reported the syllable from the right ear, reflecting left-hemispheric language dominance (for review Hugdahl, Citation1995). This right ear advantage was stronger in males than females (Bless et al., Citation2015). Kenneth Hugdahl at the University of Bergen set up a database with the same dichotic listening test, applied under more controlled experimental conditions in several laboratories around the world for over more than 20 years (Hugdahl, Citation2003). In a subsample of this database, comprising behavioural data from 1782 participants and 100 participants with behavioural and fMRI data, it was found that, behaviourally, there were no significant differences between males and females (d = 0.02). When broken down by age, males were more lateralized than females in children <10 yrs (d = 0.16), young adults between 16–50 yrs (d = 0.13,) and older adults > 50 yrs (d = 0.07). However, this difference was only significant in young adults, the largest sub sample (n = 839, one-tailed, 0.6% explained variance). In young adolescents (10–15 yrs), females were in fact more lateralized than males (d = −0.23, 1% explained variance). There were no sex differences in the fMRI data (Hirnstein et al., Citation2013). The stronger lateralization is likely based on an earlier onset of puberty and thus brain maturation in female adolescents. In general, language lateralization has been found to change with age: lateralization gradually increases from early childhood till adolescence, appears to peek at young adulthood and declines in older adults (for review Rosselli, Ardila, Matute, & Velez-Uribe, Citation2014). The fact that language lateralization is most pronounced in young adults might also explain why the sex difference in behavioural language lateralization was strongest in that age group.
Two earlier fMRI studies with samples between ca. 100 and 300 participants also did not detect sex differences in hemispheric asymmetry (Frost et al., Citation1999; Knecht et al., Citation2000). However, a structural MRI study with three datasets comprising 2337, 935, and 888 participants found a stronger leftward asymmetry of the planum temporale in males (Guadalupe et al., Citation2015) with effect sizes between d = 0.04–0.38. In addition, the authors carried out a genome wide association study and were able to link planum temporale asymmetry to genes involved in steroid hormone receptor activity and steroid metabolic processes corroborating that sex hormones may play a crucial role for the development of sex differences in hemispheric asymmetry.
Taken together, the findings from meta-analyses (for an overview, see ) as well as large scale behavioural and functional neuroimaging studies paint a very coherent picture across different forms of lateralization assessment and sensory modalities: In our view, stronger hemispheric asymmetry in males is not the result of a publication bias – it truly exists. However, regardless of how hemispheric asymmetry is assessed – whether behaviourally or with neuroimaging – the effect sizes are very small, on the order of d = 0.05–0.15, which can be reliably detected only with meta-analyses and large-scale studies.
Table 1. Overview of meta-analyses on sex differences in hemispheric asymmetry, pointing to an overall slightly stronger lateralization in males.
Does a bilateral brain generally enhance verbal performance?
If males perform better on spatial tasks because they have a more asymmetric brain, and if females perform better in verbal tasks because they have a more bilateral brain, then the underlying assumption is that, in general (i.e., in both males and females), stronger lateralization is associated with superior spatial and inferior verbal processing. We now evaluate the evidence for this assumption, presented according to how lateralization was determined.
Handedness data
As pointed out above, Levy (Citation1969) found better spatial performance in right-handers and better verbal performance in left-handers, which she attributed to a more bilateral brain organization in left-handers. Apart from the fact that she probably overestimated the impact of bilateral left-handers in her sample, most replication attempts were unsuccessful. Already in 1979, Sherman dismissed claims by Levy (Citation1974) that her findings were replicated and listed several studies, alongside her own (Sherman, Citation1979) that yielded different results.
In subsequent years, several studies addressed the relationship between cognitive abilities and handedness. According to Mellet, Jobard, et al. (Citation2014), these studies can be broadly divided into three categories: The first shows that right-handers consistently obtain higher scores on cognitive tasks than left-handers (Johnston, Nicholls, Shah, & Shields, Citation2009; Nicholls, Chapman, Loetscher, & Grimshaw, Citation2010). The second line of findings did not find differences between left- and right-handers but found differences between participants with weak and strong hand preferences. Thus, handedness strength rather than handedness direction seems to matter. For example, two large-scale studies reported generally lower cognitive performance in mixed-handers compared to participants with consistent left- or right-hand preference (Corballis, Hattie, & Fletcher, Citation2008; Crow, Crow, Done, & Leask, Citation1998; Leask & Crow, Citation2006). Annett (Citation1992, Citation1995), on the other hand, argued that mixed-handers would have inferior verbal skills while individuals with strong hand preferences would have inferior spatial skills (but see McManus, Shergill, & Bryden, Citation1993). The third line of findings, based on the “BBC internet study” comprising ca. 250.000 participants, revealed an “M”- shaped relationship between handedness and mental rotation scores: That is, on a continuum from strong left- to strong right-handedness those individuals that were strong left-handers, had no hand preference at all, or were strong right-handers obtained rather low scores, while participants with intermediate left and right hand preferences got relatively high scores (Peters, Reimers, & Manning, Citation2006). Mellet, Jobard, et al. (Citation2014) found that familial left-handedness together with mixed hand preference was related to poorer verbal and spatial abilities. The most recent study on handedness and intelligence, a meta-analysis comprising data from more than 60.000 participants (Ntolka & Papadatou-Pastou, Citation2018), revealed a mildly higher IQs for right- as compared to left-handers with d = 0.07.
Taken together, the literature is quite inconsistent regarding handedness and cognitive performance and it is difficult to ascertain how handedness and cognitive abilities are associated. This may be the consequence of large methodological variability in handedness assessment. Handedness can be defined on the basis of hand preference, hand skill or physical strength and several different self-report and behavioural tools are in use (Beaton, Citation1985). However, none of the major trends in the findings described above is consistent with Levy’s (Citation1969) results.
Behavioural laterality data
A comprehensive analysis of how behaviourally assessed hemispheric asymmetry and cognitive performance are associated was provided by Boles, Barth, and Merrill (Citation2008). They examined several visual half-field and dichotic listening tasks assessing various verbal and spatial abilities and correlated the degree of asymmetry in those tasks with the accuracy and reaction times. Here too the findings were heterogeneous. Some verbal tasks, particularly those that were assessed with dichotic listening, showed that performance increased as the left-hemispheric language bias became stronger (for replication see Barth, Boles, Giattina, & Penn, Citation2012; Hirnstein, Hugdahl, & Hausmann, Citation2014). However, verbal tasks that were assessed with the visual half-field paradigm (e.g., lexical decisions) were better performed by participants with a more symmetric language representation (for replication Hirnstein et al., Citation2014; but see Chiarello, Welcome, Halderman, & Leonard, Citation2009). Similarly, tasks that tapped into what were named “spatial quantitative” functions yielded positive asymmetry-performance correlations, whereas negative correlations emerged in “spatial positional” tasks (Boles et al., Citation2008) – both assessed with the visual half-field paradigm. A neurodevelopmental model was put forward according to which functions that lateralize very early (until 5 years of age) and very late in the ontogenetic development (after 11 years of age) yield positive asymmetry-performance correlations. Functions that lateralize at intermediate stages on the other hand would show negative correlations (Boles et al., Citation2008).
Neuroimaging data
Consistent with Levy’s idea, a smaller fMRI study (N = 14) found that stronger right-hemispheric activation during a dichotic listening task – and therefore a more bilateral language representation – correlated positively with performance in a set of verbal tasks (van Ettinger-Veenstra et al., Citation2010). In contrast, Everts et al. (Citation2009) found that stronger language lateralization (also assessed with fMRI) was associated with a higher verbal IQ in 20 adolescents. Similarly, another fMRI study (N = 78) revealed that stronger leftward asymmetry in the pre-central gyrus and stronger rightward asymmetry in the cerebellum during a verbal fluency task was correlated with a higher number of generated words (Razafimandimby, Tzourio-Mazoyer, Mazoyer, Maiza, & Dollfus, Citation2011). In accordance with these findings, stronger structural asymmetry in cortical thickness in the inferior and posterior temporal cortices has been associated with better performance in a vocabulary task in 215 children and adults (Plessen et al., Citation2014).
Two further fMRI studies found – inconsistent with Levy’s hypothesis – that stronger lateralization is related to both better verbal and spatial performance. Specifically, Mellet, Zago, et al. (Citation2014) tested 297 participants on various verbal and spatial cognitive tasks including, among others, verbal fluency, verbal memory, and mental rotation. The composite scores of all verbal and spatial tasks correlated positively with the degree of language lateralization. However, sex had only been included as a covariate, leaving open whether this pattern was the same for males and females. Using fMRI, Gotts et al. (Citation2013) identified a network lateralized to the left hemisphere, comprising language areas, and a network lateralized to the right hemisphere, comprising areas involved in visuospatial and attentional processes. The stronger both systems were lateralized, the higher were participants’ verbal and performance IQ. Only males (N = 62) were tested, however.
Groen, Whitehouse, Badcock, and Bishop (Citation2012) examined the relationship between hemispheric asymmetry and cognitive performance in a sample of 60 typically developing children (aged 6-16). Lateralization of verbal and spatial functions was determined with a language production and visuospatial working memory task, respectively, using fTCD. As predicted, the verbal task was lateralized to the left hemisphere and the spatial task to the right hemisphere. Male children showed slightly stronger rightward lateralization of the spatial task than female children but there was no sex difference in performance. This is inconsistent with Levy’s hypothesis. Groen et al. also specifically tested the “crowding hypothesis” and found that children with verbal and spatial functions lateralized to the same hemisphere performed similarly on a number of cognitive and language tasks as compared to children who had both functions lateralized to different hemispheres. This is inconsistent with the “crowding hypothesis” according to which the two functions should “crowd” the one hemisphere they are lateralized to and result in poorer performance. In line with the “crowding hypothesis”, however, Powell, Kemp, and García-Finaña (Citation2012) found that when verbal and spatial functions were lateralized to different hemispheres, performance in verbal and spatial tasks in the Wechsler Adult Intelligence Scale was higher. Lateralization of verbal and spatial functions was determined with a verbal fluency and line bisection fMRI paradigm, respectively. The sample size was N = 82.
Summary
This section examined a key assumption of Levy’s hypothesis, namely that hemispheric asymmetry promotes spatial but impedes verbal performance. The majority of tasks discussed here thus did not reveal cognitive sex differences and rather probed verbal or spatial abilities, in general. The findings across different forms of laterality assessment are very heterogeneous but they show a few clear trends: Firstly, Levy’s finding of superior spatial and verbal performance by right- and left-handers, respectively – the basis of her hypothesis regarding cognitive sex difference – is seldom replicated. Secondly, the work by Boles et al. (Citation2008) emphasizes that even within the verbal and spatial domain the relationship between hemispheric asymmetry and cognitive performance is task-dependent. This clearly contradicts the assumption of a generally beneficial or detrimental effect of laterality for cognitive performance (Levy, Citation1972). Thirdly, although contradictory findings exist (van Ettinger-Veenstra et al., Citation2010), the bulk of neuroimaging studies rather suggests that stronger lateralization enhances both spatial (Gotts et al., Citation2013; Mellet, Zago, et al., Citation2014) and verbal performance (Everts et al., Citation2009; Gotts et al., Citation2013; Mellet, Zago, et al., Citation2014; Plessen et al., Citation2014; Razafimandimby et al., Citation2011). The enhanced spatial performance would be consistent with Levy’s hypothesis; the enhanced verbal performance is not: females are thought to outperform males because they have a bilateral, less lateralized language representation. Finally, there is also no clear empirical support for the “crowding hypothesis”, the antecedent of the idea that a more asymmetrical and bilateral brain organization in males and females, respectively, would lead to better spatial and verbal performance.
In general, the relationship between hemispheric asymmetry and cognitive performance is very complex and appears to depend on which cognitive function or task is assessed, which groups are compared, and how lateralization is determined. One may argue, perhaps, in Levy’s favour that a strongly lateralized brain seems to enhance spatial performance but a bilateral brain does certainly not enhance verbal performance per se.
Are cognitive sex differences directly dependent upon sex differences in hemispheric asymmetry?
Since the relationship between hemispheric asymmetry and performance appears to be task-dependent, this section specifically deals with tasks that reliably yield a male or female advantage. If Levy’s hypothesis is correct, then we would expect a dependency between the sex differences in cognitive abilities and hemispheric asymmetries. That is, studies that assessed both performance in sex-sensitive cognitive tasks and hemispheric asymmetry should yield: (1) the typical male/female advantage in a sex-sensitive spatial/verbal task (= “cognitive criterion”), and (2) stronger hemispheric asymmetry in males (= “hemispheric asymmetry criterion”). A conservative approach would additionally require that cognitive performance and hemispheric asymmetry are correlated but as we will see most studies already fail to confirm the first two criteria.
To verify whether cognitive sex differences depend upon the sex difference in hemispheric asymmetry, we carried out a systematic literature review. To this end, we identified studies that included both findings on males’ and females’ cognitive performance in sex-sensitive, cognitive tasks and males’ and females’ degree of hemispheric asymmetry.
Systematic review
We followed the PRISMA guidelines for systematic reviews (Moher, Liberati, Tetzlaff, & Altman, Citation2009). ISI Web of Science was searched by combining each of the following sex-sensitive tasks “mental rotation”, “paper folding”, “line orientation task”, “embedded figures”, “water level task”, “verbal fluency”, “verbal memory”, and “reading performance” with each of the expressions “asymmetry” and “laterality”. The search was conducted on 6th March 2017 and yielded 1687 records. In addition, 31 records were identified by searching reference lists of relevant articles. Studies that fulfilled the following criteria were included: (1) Participants completed a sex-sensitive, cognitive task and performance for both males and females was provided, (2) information on hemispheric asymmetry is provided for both males and females, (3) hemispheric asymmetry was assessed with neuroimaging, electroencephalography (EEG), dichotic listening, or visual-half field methods. For reasons stated above (Ocklenburg et al., Citation2014), handedness was considered a too coarse proxy for hemispheric asymmetry. Questions have also been raised regarding the reliability and validity for dichotic listening and visual-half field techniques (for review Voyer, Citation1998). Indeed, there are indications that reliability in these measures can be low (Voyer, Citation1998). However, there are also findings showing that if the right statistical measures are used and if the studies are designed properly (Brysbaert & D’Ydewalle, Citation1990; Hunter & Brysbaert, Citation2008), reliability and validity are satisfactory. Therefore, studies employing these methods were included. Studies using other questionable laterality assessment methods, however, such as asking participants to turn their heads or eyes to the left or right to activate the right or left hemisphere, were omitted. Further exclusion criteria were (a) irrelevant field (e.g., “asymmetry” research in geology, palaeontology, physics); (b) irrelevant cognitive task; (c) clinical, homosexual, or transsexual samples; (d) no presentation of additional emotional arousal/stimuli to prevent confounding effects of emotional lateralization (e.g., studies that induce sadness); (e) studies published in languages other than English, German or any Scandinavian language; (f) case studies; (g) review articles. An overview of the article selection process is provided in .
Figure 3. PRISMA flow diagram showing the study selection process for the systematic review.
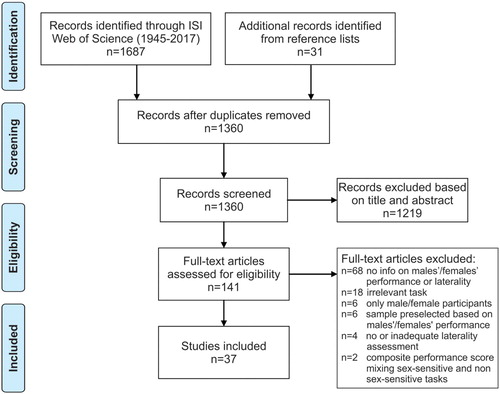
In total, 37 studies were included in the systematic review, comprising data from 2623 participants (1265 males and 1358 females). Hemispheric asymmetries were examined with a wide range of methods including behavioural assessment (using dichotic listening and the visual-half field technique), as well as EEG and various neuroimaging procedures. The vast majority of studies examined spatial abilities (n = 31) with a focus on mental rotation (n = 25). This is hardly surprising as mental rotation is the largest (Zell et al., Citation2015) and arguably the most researched cognitive sex difference. However, other abilities/tasks such as line orientation (n = 3), paper folding (n = 3), spatial memory tasks (n = 3), the Water level task (n = 2), and embedded figures (n = 2) are also included. In addition, nine studies examined verbal abilities, with six providing data on verbal fluency, three on verbal memory, and one on reading skills. Thus, all major sex-sensitive tasks/abilities are included. We feel that this represents a solid sample upon which conclusions can be based.
Rationale of the systematic review
To evaluate Levy’s hypothesis, we classified every study as “in line”, “not in line”, or ”partly in line” with Levy’s hypothesis. “Partly in line” refers to studies where one finding supports while another does not support Levy’s hypothesis. For example, Catani et al. (Citation2007) found that greater bilateral language organization in females was associated with better verbal memory performance – in line with Levy’s hypothesis. However, in the same study females were not better in verbal fluency – not in line with Levy’s hypothesis. A fourth category was “inconclusive”. This refers to studies that neither found a sex difference in cognitive tasks nor a sex difference in hemispheric asymmetry.
We opted for such a classification approach, because one could induce unwanted bias by lumping together all the studies in a traditional, quantitative meta-analysis. For example, studies employing the visual-half field technique or dichotic listening tend to have larger samples than neuroimaging studies. This would give them more weight in a meta-analysis even though neuroimaging certainly allows for more direct assessment of hemispheric asymmetry. For the same reason different neuroimaging laterality measures such as DTI, fMRI, SPECT, PET, and fTCD felt too diverse to be lumped together. Nevertheless, to provide a better overview and to disentangle whether different methods yield different outcomes, the results are presented in three separate sections: Behavioural assessment, EEG, and neuroimaging (see below). A summary of the results can be found in .
Table 2. Overview of studies that examined sex differences in sex-sensitive cognitive tasks and hemispheric asymmetry.
Behavioural assessment
Of the twelve included studies, only one was consistent with Levy’s hypothesis: S. D. Voyer and Voyer (Citation2015) found the typical male advantage in mental rotation and a stronger right ear advantage in a dichotic listening task, reflecting a stronger left-hemispheric specialization for language. Four studies revealed partial support (Kail & Siegel, Citation1978; Rilea, Citation2008a; van der Ham & Borst, Citation2011; Wegesin, Citation1998). For example, Kail and Siegel (Citation1978) asked participants to memorize either digits (interpreted as a verbal memory task) or digit positions (interpreted as a spatial memory task), presented with the visual half-field technique. In line with Levy’s hypothesis, females were better in the verbal condition and males showed stronger asymmetry. However, males were better in the spatial condition even though there was no sex difference in hemispheric asymmetry. It is also debatable whether both cognitive tasks are sex-sensitive. In another visual half-field study van der Ham and Borst (Citation2011) presented spatial stimuli and found the expected stronger right-hemispheric involvement in males as compared to females. In line with Levy’s hypothesis, there was also a trend towards better male performance in mental rotation, but no sex difference emerged in verbal fluency and the paper folding task.
Two studies were inconsistent with Levy’s hypothesis: Both employed the visual half-field technique and found a male advantage in mental rotation but no sex difference in hemispheric asymmetry (Dollinger, Citation1995; Rilea, Citation2008b). Two studies were inconclusive yielding neither a sex difference in mental rotation nor in hemispheric asymmetry (Jones & Anuza, Citation1982; Uecker & Obrzut, Citation1993) and three studies had elements of both inconclusive and incongruent findings (Chiarello, Welcome, Halderman, Towler, et al., Citation2009; Rilea et al., Citation2004; Voyer, Citation1995). For example, Chiarello, Welcome, Halderman, Towler, et al. (Citation2009) assessed hemispheric asymmetry by presenting verbal stimuli in a series of visual half-field tasks and administered standardized reading tests. In two out of eight visual half-field tasks, males yielded a stronger left-hemispheric advantage than females. However, this did not translate into better reading skills in females: Most subtests did not reveal sex differences, except one where, in fact, males performed better. The other six visual half-field tasks did not show any sex difference in hemispheric asymmetry. Similarly, Rilea et al. (Citation2004) presented mental rotation stimuli with the visual half-field technique and found stronger asymmetry in males, but no male advantage in mental rotation; in the Water level test males actually performed better but did not show any stronger asymmetry. Both findings are inconsistent with Levy’s hypothesis. In the Paper Folding task there was neither a sex difference in performance nor in hemispheric asymmetry (= inconclusive).
EEG
Of the ten included EEG studies, none fully supported Levy’s hypothesis. Two studies yielded at least partial support (Gootjes et al., Citation2008; Wegesin, Citation1998). For example, Gootjes et al. (Citation2008) reported better male than female performance in mental rotation regardless of whether Flanker stimuli were presented together with the mental rotation stimuli or not. However, a stronger right-hemispheric asymmetry in males, as assessed with EEG, only emerged when the Flanker stimuli were presented (in line with Levy’s hypothesis). When Flanker stimuli were absent, there was no sex difference in hemispheric asymmetry (not in line with Levy’s hypothesis).
Four studies were inconsistent with Levy’s hypothesis. Either males showed the expected stronger hemispheric asymmetry but no superior mental rotation performance (Hahn et al., Citation2010), or males showed the expected superior spatial performance but there was no sex difference in hemispheric asymmetry (Roberts & Bell, Citation2003) – or in two cases females even had stronger hemispheric asymmetry (Pellkofer et al., Citation2014; Yu et al., Citation2009). Three studies failed to find sex differences in both hemispheric asymmetry and cognitive performance (Beste et al., Citation2010; Johnson et al., Citation2002; Papousek et al., Citation2011), while Jonathan E. Roberts and Bell (Citation2002) had elements of both inconsistent and inconclusive findings: Their participants completed two mental rotation tasks, a 2-dimensional and a 3-dimensional one. In the 3-dimensional task, there was neither a sex difference in performance nor in hemispheric asymmetry (inconclusive), but in the 2-dimensional task females had an unexpectedly stronger left hemispheric asymmetry than males – and there was no sex difference in performance (not in line with Levy’s hypothesis).
Neuroimaging
Of the seventeen studies that assessed hemispheric asymmetry with neuroimaging one is in line with Levy’s hypothesis: R. C. Gur et al. (Citation2000) found that after a median-split males were overrepresented among the group of “good performers” in a line orientation task. Males also showed stronger right-hemispheric activations as compared to females during this task as assessed with fMRI. Three studies provided partial support (Catani et al., Citation2007; Deutsch et al., Citation1988; Hattemer et al., Citation2011). For example, Catani et al. (Citation2007) reconstructed a direct neural pathway between Wernicke’s and Broca’s area through the arcuate fasciculus using MR based diffusion tensor imaging (DTI). The less pronounced this direct pathway was in the left hemisphere (i.e., the more bilaterally language was organized), the better participants performed on a verbal memory task. Females were overrepresented among those with a more bilateral or weaker language lateralization and, accordingly, outperformed males (in line with Levy’s hypothesis). However, the same males and females performed equally well on a verbal fluency test.
On the other hand, seven studies yielded results that were inconsistent with Levy’s hypothesis: In a number of spatial tasks comprising mental rotation, spatial memory, and line orientation males either showed the expected stronger right-hemispheric asymmetry but no superior spatial performance (Frings et al., Citation2006), or showed the expected superior spatial performance but there was no sex difference in hemispheric asymmetry (Persson et al., Citation2013) – or hemispheric asymmetry was in fact stronger in females (Clements et al., Citation2006; Walter et al., Citation2000). When verbal abilities are considered, one fMRI study found that males were slightly more lateralized during verbal fluency than females, but both sexes produced a similar number of words (Badzakova-Trajkov et al., Citation2010). In turn, Ragland et al. (Citation2000) found that females remembered more words in a verbal memory task, but no sex differences emerged in hemispheric asymmetry as assessed with PET. Finally, Halari et al. (Citation2006) found both the typical male advantage in mental rotation and the typical female advantage in verbal fluency. However, there was no sex difference in hemispheric asymmetry when males and females were directly compared using fMRI. None of these findings is in line with Levy’s hypothesis.
Six studies using fMRI, DTI, structural MRI, SPECT, and PET in mental rotation, verbal fluency, reading skills, and the Embedded Figures task found neither a sex difference in cognitive performance nor in hemispheric asymmetry (Allendorfer et al., Citation2016; Chiarello, Welcome, Halderman, Towler, et al., Citation2009; Hao et al., Citation2013; Hugdahl et al., Citation2006; Unterrainer et al., Citation2000; Wendt & Risberg, Citation1994). Finally, an fMRI study by O’Boyle et al. (Citation2005) was not included in the systematic review, because only males were tested. Nevertheless, we found the results of note: Better mental rotation performance was accomplished with a more bilaterally activated cortical network comprising parietal and frontal areas – which is inconsistent with Levy’s idea that greater right-hemispheric asymmetry would facilitate spatial abilities.
Summary
Regardless of whether the findings are grouped by method (behavioural, EEG, or neuroimaging assessment) or domain (spatial/verbal abilities), the empirical evidence for Levy’s hypothesis is very slim. Only two studies fully supported it. Eight studies yielded partial support. Leaving aside the inconclusive studies, thirteen studies showed a pattern that was inconsistent with Levy’s hypothesis. In many of them there was either cognitive sex differences or stronger asymmetry in males (Badzakova-Trajkov et al., Citation2010; Halari et al., Citation2006; Rilea, Citation2008b; Roberts & Bell, Citation2003). This would suggest that both phenomena are at least to some extent independent of each other.
A general major concern is low statistical power. To support Levy’s hypothesis, we expected two sex differences, one in cognitive abilities and one in hemispheric asymmetry. To find a significant sex differences in mental rotation with 80% power, a sample of 48 participants (24 males and 24 females) is sufficient – based on a one-tailed, independent t-test with d = 0.73 (Linn & Petersen, Citation1985) and α = .05 as calculated with G*Power 3.1 (Faul, Erdfelder, Buchner, & Lang, Citation2009). To find the female advantage in verbal fluency or verbal memory (ca. d = 0.30) with the same parameters, one would need already 278 participants in total. The sex difference in hemispheric asymmetry ranges between d = 0.05 and 0.15 as outlined above. In the more optimistic scenario (d = 0.15), one would need roughly 1100 participants in total; with the more conservative d = 0.05 one would need roughly 10000 participants in total. None of the studies that were included in our systematic review came close to such numbers. Running a design analysis (Gelman & Carlin, Citation2014) on Levy’s original dataset from 1969 further illustrates the problems that arise from drawing conclusions based on small, noisy samples. As pointed out above a recent meta-analysis with more than 60.000 participants found IQ differences between left- and right-handers on the order of d = 0.07 (Ntolka & Papadatou-Pastou, Citation2018). This translates roughly into a difference of 1.5 IQ points between left- and right-handers - in contrast to the 13 IQ point difference reported by Levy (Citation1969). If we take these 1.5 IQ points as a reasonable estimate for the true effect size – ignoring that factors other than hemispheric asymmetry might contribute to this difference – and enter it together with 4.26 as the standard error (based on Levy’s reported mean difference and p-value) in the “retrodesign” function by (Gelman & Carlin, Citation2014), we find that there is only 6% power to replicate Levy’s finding. If a replication study finds a significant effect, there is a 16% probability that it is in the wrong direction (i.e., showing higher IQs for left-handers) and the estimated effect will be 7 times too high. Based on those numbers from the power and design analysis, it is clear that many of the studies that are included in this systematic review are underpowered and likely to produce chance findings that can go in either direction.
Given the large number of participants that are necessary to reliably detect a stronger lateralization in males, it is conceivable that studies such as Roberts and Bell (Citation2003) could be classified “inconclusive” instead of “not in line with Levy’s hypothesis”, because the sample size (N = 32) may have been sufficient to find a significant male advantage in mental rotation but not in hemispheric asymmetry. On the other hand, one would expect that a study that was large or lucky enough to find the subtle sex difference in hemispheric asymmetry should also yield the larger sex differences in cognitive abilities. However, several studies that had picked up stronger hemispheric asymmetry in males did not find cognitive sex differences (Badzakova-Trajkov et al., Citation2010; Chiarello, Welcome, Halderman, Towler, et al., Citation2009; Hahn et al., Citation2010) or found sex differences in only one task but not another (Catani et al., Citation2007; Deutsch et al., Citation1988; Rilea et al., Citation2004; van der Ham & Borst, Citation2011). In any case, it is striking that out of 37 studies only two produce findings that are fully in line with Levy’s hypothesis.
In summary, the systematic review comprises data from more than two thousand participants on a wide array of behavioural and neuroimaging methods. This provides a comprehensive fundament for drawing conclusions, but it also makes it challenging to synthesize the data. We opted for a qualitative approach to avoid unwanted biases and because it seems sufficient to test whether sex differences in cognitive abilities are contingent on sex differences in hemispheric asymmetry. We found a pervasive lack of support for Levy’s hypothesis, either because the results were not statistically significant or inconsistent with her idea. Nevertheless, this approach has limitations: The major downside is that it does not allow quantifying the association between hemispheric asymmetry and cognitive performance. Due to the general lack of power, we cannot conclude that there is no association between sex differences in cognitive tasks and hemispheric asymmetry; they may well be correlated (see also the general discussion below). However, Levy’s hypothesis was not that the two phenomena are correlated but that one causes the other. As we have demonstrated, the presence of cognitive sex differences does not necessitate the presence of sex differences in hemispheric asymmetry and, vice versa, the presence of sex differences in hemispheric asymmetry does not necessitate the presence of cognitive sex differences.
General discussion
Jerre Levy deserves credit for putting forward an idea that was well-conceived and novel at the time and that has been stimulating numerous studies for more than 40 years. Her hypothesis was the starting point for similar theories that all argue that cognitive sex differences arise from a sex difference in hemispheric asymmetry. The popularity of this idea was fading with evidence in the late 1990s that sex differences in hemispheric asymmetries are very small. Nevertheless, a comprehensive review that specifically investigated whether stronger lateralization in males gives rise to cognitive sex differences has been missing. The current review aimed to address this issue and the bottom line is: In light of the empirical evidence, the idea that males perform better in spatial tasks and females in verbal tasks because males have a more lateralized brain is no longer tenable.
Although oversimplified, it is fair to conclude that if sex differences emerge, they tend to be such that males excel in certain spatial and females in certain verbal tasks. Levy was also correct to assume that males have a more asymmetric brain organization than females. Converging empirical evidence reviewed here suggests, however, that the effect is very small with, d = 0.05–0.15 (Guadalupe et al., Citation2015; Hirnstein et al., Citation2013; Sommer et al., Citation2004; Sommer et al., Citation2008; Voyer, Citation1996, Citation2011). As was pointed out previously (e.g., Halpern, Citation2012; Voyer, Citation1996), it is hard to see how such a small effect could underlie the male advantage in mental rotation of d = 1.0 and above in some studies (e.g., Jansen & Heil, Citation2010; Peters, Citation2005). On the other hand, stronger male asymmetry emerges with such a consistency across different methods that it cannot be dismissed. It should also be noted that the vast majority of studies that investigated sex differences in hemispheric asymmetry did not account for hormonal fluctuations (e.g., across the menstrual cycle), which have been shown to affect lateralization (for review Hausmann, Citation2017; Hausmann & Bayer, Citation2010; Weis & Hausmann, Citation2010). This would add noise to the data and the fact that the stronger lateralization pattern in males still consistently emerges is further testimony to the robustness of the effect.
Another strong argument against Levy’s hypothesis is that one of the key assumptions – namely that stronger lateralization generally enhances spatial and reduces verbal performance – does not hold. Crucially, many studies find either cognitive sex differences or stronger asymmetry in males (e.g., Badzakova-Trajkov et al., Citation2010; Chiarello, Welcome, Halderman, Towler, et al., Citation2009; Halari et al., Citation2006; Rilea, Citation2008b; Roberts & Bell, Citation2003), suggesting the two phenomena are at least to some extent independent of each other. Taken together, sex differences in hemispheric asymmetry and cognitive sex differences may be related, but sex differences in hemispheric asymmetry are certainly not the driving force for cognitive sex differences.
Towards better models
Given the apparent lack of empirical support, why is the idea that hemispheric asymmetry gives rise to cognitive sex differences still marketed as a viable biological explanation these days? As outlined above, one reason could be that a comprehensive review was missing so far. Our hope is that the present paper represents the metaphorical final nail in the coffin. Another reason might be the high intuitive appeal of the model. At first glance, it may sound pervasive that if more neural space is devoted to a function, then this function will yield better results. That is, by “giving up” computational power for spatial tasks in the right hemisphere females gained extra space for verbal processing and hence better verbal performance. This may have sound convincing in the pre-neuroimaging era but it has repeatedly been shown that as task performance improves, it becomes more automatized and, in fact, fewer brain areas are involved or those areas involved are less activated (e.g., Haier, Karama, Leyba, & Jung, Citation2009).
A further reason for the continuous popularity of Levy’s hypothesis is the lack of alternatives: With all its empirical shortcomings it is at least a testable model. Over the last decades, a wealth of studies examined in which areas cognitive sex differences exist, under which circumstances they emerge, and which biological, psychological, and socio-cultural factors affect them. At the same time huge progress in neuroscientific methods allowed for in vivo examination of the brain with high temporal and spatial resolution. Nevertheless, a better model that could explain the underlying neural mechanism of the male advantage in certain spatial and the female advantage in certain verbal tasks is not in sight. What is needed to develop such model(s)? We believe that more studies are required that specifically try to tie neuroscientific to behavioural outcomes. On the one hand, the behavioural aspects of cognitive sex differences, particularly for the male advantage in mental rotation, have been extensively researched. On the other hand, there is an ever growing literature on sex differences in the brain on all macroscopic and microscopic levels including, for example, structure, function, connectivity, and neurochemistry (for review Cahill, Citation2006). What is largely missing are the associations between behaviour and its underlying neural correlates (see also McCarthy, Citation2016). There are, of course, a number of neuroimaging studies that examined sex differences in tasks such as mental rotation (e.g., Halari et al., Citation2006; Hugdahl et al., Citation2006; Jordan, Wüstenberg, Heinze, Peters, & Jancke, Citation2002; e.g., Semrud-Clikeman, Fine, Bledsoe, & Zhu, Citation2012), verbal fluency (Catani et al., Citation2007; Halari et al., Citation2006), or across other verbal and spatial tasks (Gur et al., Citation2000; Gur et al., Citation1999; Gur & Gur, Citation2016; Satterthwaite et al., Citation2015; Tunç et al., Citation2016). However, brain-behaviour associations that could account for cognitive sex differences are rare. Structural asymmetry (Catani et al., Citation2007) and overall intracranial volumes (Gur et al., Citation1999) have been linked to verbal memory as well as spatial and verbal performance, respectively. In the latter case, spatial and verbal performance was based on composite scores of different verbal and spatial tasks. Likewise, data from a large-scale neuroimaging cohort revealed different connectivity patterns in males and females that are associated with their performance in a number of cognitive tasks comprising verbal memory and line orientation (Satterthwaite et al., Citation2015; Tunç et al., Citation2016; for review Gur & Gur, Citation2016). These findings are promising but need corroboration and do not address, for instance, the well-established sex differences in mental rotation or verbal fluency. The neural mechanisms that underlie cognitive sex differences remain elusive for the moment.
The relationship between hemispheric asymmetry and cognitive performance
One could argue that at its core, Levy’s hypothesis is primarily about the general relationship between lateralization and cognitive performance, rather than about males versus females who served as proxies for asymmetric versus bilateral individuals, respectively. Based on existing literature and this review, it seems that neither the assumption that males/females are good proxies for an asymmetric/bilateral brain organization, respectively, nor that bilateral brains promote verbal skills is correct. There is ample evidence, however, that lateralization and cognitive performance are associated. Yet, current findings on the lateralization/cognitive performance association are very inconsistent and we only have a vague understanding of how exactly lateralization and cognitive performance are linked. For instance, which tasks (or group of tasks) are enhanced by an asymmetric or bilateral brain and what is the underlying mechanism? A fair number of dichotic listening and visual half-field studies have been carried out for which a neurodevelopmental model has been proposed (Boles et al., Citation2008), but this work requires corroboration – especially from neuroimaging which has now begun to explore these associations (Gotts et al., Citation2013; Mellet, Zago, et al., Citation2014; Plessen et al., Citation2014; van Ettinger-Veenstra et al., Citation2010).
Another issue regarding the lateralization/cognitive performance relationship that needs clarification is causality. Does the lateralization pattern cause differences in cognitive performance or does better or worse cognitive performance lead to different laterality patterns? Are both cognitive performance and lateralization the consequence of other processes? Bishop (Citation2013) raised a similar issue for the relationship between anomalous language lateralization and language disorders. Individuals with language disorders such as dyslexia or selective language impairment frequently show abnormal lateralization patterns but it is unclear whether lateralization is the “cause, correlate, or consequence” (Bishop, Citation2013, p. 1302) of language disorders. Bishop (Citation2013) then provides a number of useful suggestions how the nature of the relationship can be uncovered, for example, by using longitudinal twin and family studies. A similar approach would seem fruitful to disentangle the relationship between asymmetry and cognitive performance.
A third issue is whether the lateralization/cognitive performance relationship is the same for males and females. Levy implicitly assumed this would be the case: both females and males would show better verbal skills if they had bilateral language lateralization. Waber (Citation1976) explicitly stated that sex is not the important variable but the developmental trajectory of lateralization. While there is some support for this notion (Boles, Citation2005; Hirnstein et al., Citation2014; Mellet, Zago, et al., Citation2014), there are also hints that the relationship might be sex-dependent (e.g., Frings et al., Citation2006; Hirnstein, Leask, Rose, & Hausmann, Citation2010). So far, there is no clear consensus.
A final issue pertains to the question what is the impact of the small (but robust) stronger lateralization in males? As pointed out above, even if hemispheric asymmetries are not the driving force, they may still contribute to cognitive sex differences. In fact, if there are small but robust sex differences in hemispheric asymmetry, and if performance in certain cognitive tasks correlates with hemispheric asymmetry, then those cognitive tasks should also show at least subtle sex differences. To further elucidate the impact of stronger lateralization in males, large-scale studies with sufficient statistical power are needed. Studies with small samples produce chance findings that can go in either direction and overestimate the true effect size. The problem with underpowered studies in psychology and neuroscience has been recognized for a while (e.g., Button et al., Citation2013) and several remedies have been put forward (e.g., Brysbaert & Stevens, Citation2018; Button et al., Citation2013; Giner-Sorolla, Citation2018). Secondly, the dynamic nature of lateralization needs to be taken more into account. This does not only pertain to studying the impact of stronger asymmetry in males but, in general, the relationship between lateralization and cognitive performance. Functional hemispheric asymmetries are not invariant but change with age (e.g., Cabeza, Citation2002) and emotional states (e.g., Davidson, Citation1995; Papousek & Schulter, Citation2002). As pointed out above, hemispheric asymmetry has also been found to fluctuate with changes in sex hormone levels (for review Hausmann, Citation2017; Hausmann & Bayer, Citation2010; Weis & Hausmann, Citation2010). Several studies already investigated the relationship between hemispheric asymmetry and cognition, for instance, by testing women in different phases of the menstrual cycle (e.g., Hausmann, Becker, Gather, & Güntürkün, Citation2002; Mead & Hampson, Citation1996; Sanders & Wenmoth, Citation1998; Weis et al., Citation2008), but to date there is no clear pattern with respect to which hormones are crucial and how lateralization changes. This is in part the result of different methodological approaches but also because sample sizes are often small to produce stable results. In general, dynamic changes in hemispheric asymmetry are often overlooked or deliberately ignored as including them would make study designs much more complex and costly. However, in order to get a comprehensive picture of the relationship between lateralization and cognitive performance in general, and the effect of sex differences in lateralization on cognitive performance in particular, dynamic changes need to be taken into account more routinely. Third, when carrying out neuroimaging studies with a focus on sex differences several pitfalls need to be avoided (Kaiser, Haller, Schmitz, & Nitsch, Citation2009; Rippon, Jordan-Young, Kaiser, & Fine, Citation2014). For example, sex differences in the brain do not automatically imply that those sex differences were inborn. They may have simply arisen from different experiences. Further, cognitive performance needs to be measured directly and cannot simply be inferred from brain activations.
What other behavioural consequences could arise from the small but robust sex difference in hemispheric asymmetry? Another popular theory, originally proposed by Lansdell (Citation1961, Citation1962), argued that the more bilateral language lateralization would make females more resilient to unilateral stroke. That is, if the left hemisphere is lesioned, the right hemisphere can take over language functions more easily. However, after reviewing the literature Sommer et al. (Citation2004) dismissed the idea: “[…] a sex difference after left-hemisphere lesions has not been consistently reported and differences in aphasia after right-hemisphere lesions have never been demonstrated.” (p. 1849). Finally, one could speculate that the slightly more asymmetric brain organization in males might facilitate the ability to simultaneously handle two tasks that are lateralized to opposite hemispheres. This idea is based on animal research showing that strongly lateralized chicks (Rogers, Zucca, & Vallortigara, Citation2004) and fish (Dadda & Bisazza, Citation2006) are better at concurrently performing a foraging task (left-lateralized) and a vigilance task (right-lateralized) than less lateralized chicks and fish, respectively. If applied to humans this would suggest that the slightly stronger brain organisation in males might be advantageous when processing a verbal and a spatial task simultaneously. However, the attempt to replicate a parallel processing advantage of left- (verbal) and right- (spatial) hemispheric tasks in males and females has yielded only mixed results so far (Hirnstein, Hausmann, & Güntürkün, Citation2008; Lust, Geuze, Groothuis, & Bouma, Citation2011; Lust, Geuze, Groothuis, van der Zwan, et al., Citation2011).
Conclusion
Levy’s hypothesis clearly left its mark on the literature but when looking at the empirical evidence it is time now to abandon the idea and develop new and better models for the underlying neural mechanisms of cognitive sex differences. To be able to do so we need more well-designed studies with sufficient statistical power that try to tie behavioural sex differences to their underlying neural mechanisms. Moreover, we need to move beyond cognitive sex differences and gain a better understanding of the general relationship between hemispheric asymmetry and cognitive performance. This review includes a few suggestions of what and how future studies could explore.
Acknowledgments
This work was supported by the Bergen Research Foundation (grant BFS2016REK03) to M. H.
Disclosure Statement
No potential conflict of interest was reported by the authors.
Data availability statement
Data sharing is not applicable to this article as no new data were created or analysed in this study.
Additional information
Funding
References
- *Allendorfer, J. B., Hernando, K. A., Hossain, S., Nenert, R., Holland, S. K., & Szaflarski, J. P. (2016). Arcuate fasciculus asymmetry has a hand in language function but not handedness. Human Brain Mapping, 37(9), 3297–3309. doi: 10.1002/hbm.23241
- Anitei, S. (2007). Women use their whole brain, men just half of it. Retrieved from http://archive.news.softpedia.com/news/Women-Use-Their-Whole-Brain-Men-Just-Half-of-It-51982.shtml
- Annett, M. (1992). Spatial ability in subgroups of left-and right-handers. British Journal of Psychology, 83(4), 493–515. doi: 10.1111/j.2044-8295.1992.tb02455.x
- Annett, M. (1995). The right shift theory of a genetic balanced polymorphism for cerebral-dominance and cognitive processing. Cahiers De Psychologie Cognitive-Current Psychology of Cognition, 14(5), 427–480.
- *Badzakova-Trajkov, G., Häberling, I. S., Roberts, R. P., & Corballis, M. C. (2010). Cerebral asymmetries: Complementary and independent processes. PLoS ONE, 5(3), e9682. doi: 10.1371/journal.pone.0009682
- Balthazart, J. (2016). Sex differences in partner preferences in humans and animals. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1688), 20150118. doi: 10.1098/rstb.2015.0118
- Barth, J. M., Boles, D. B., Giattina, A. A., & Penn, C. E. (2012). Preschool child and adult lateralisation and performance in emotion and language tasks. Laterality: Asymmetries of Body, Brain and Cognition, 76(4), 1–16. doi: 10.1080/1357650x.2011.626435
- Beaton, A. A. (1985). Left side/right side: A review of laterality. London: Batsford Academic and Educational.
- Benton, A. L., Varney, N. R., & Hamsher, K. S. (1978). Visuospatial judgment: A clinical test. Archives of Neurology, 35(6), 364–367. doi: 10.1001/archneur.1978.00500300038006
- *Beste, C., Heil, M., & Konrad, C. (2010). Individual differences in ERPs during mental rotation of characters: Lateralization, and performance level. Brain and Cognition, 72(2), 238–243.
- Bishop, D. V. M. (2013). Cerebral asymmetry and language development: Cause, correlate, or consequence? Science, 340(6138), 1230531–1230531. doi: 10.1126/science.1230531
- Bleecker, M. L., Bolla-Wilson, K., Agnew, J., & Meyers, D. A. (1988). Age-related sex differences in verbal memory. Journal of Clinical Psychology, 44(3), 403–411. doi:10.1002/1097-4679(198805)44:3<403::AID-JCLP2270440315>3.0.CO;2-0
- Bless, J. J., Westerhausen, R., Torkildsen, J. v. K., Gudmundsen, M., Kompus, K., & Hugdahl, K. (2015). Laterality across languages: Results from a global dichotic listening study using a smartphone application. Laterality: Asymmetries of Body, Brain and Cognition, 20(4), 434–452. doi: 10.1080/1357650X.2014.997245
- Bogen, J. E., & Gazzaniga, M. S. (1965). Cerebral commissurotomy in man - minor hemisphere dominance for certain visuospatial functions. Journal of Neurosurgery, 23(4), 394–399. doi: 10.3171/jns.1965.23.4.0394
- Boles, D. B. (2005). A large-sample study of sex differences in functional cerebral lateralization. Journal of Clinical and Experimental Neuropsychology, 27(6), 759–768.
- Boles, D. B., Barth, J. M., & Merrill, E. C. (2008). Asymmetry and performance: Toward a neurodevelopmental theory. Brain and Cognition, 66(2), 124–139. doi:10.1016/j.bandc.2007.06.002|ISSN 0278-2626
- Bryden, M. P., McManus, I. C., & Bulman-Fleming, M. B. (1994). Evaluating the empirical support for the Geschwind-Behan-Galaburda model of cerebral lateralization. Brain and Cognition, 26(2), 103–167. doi:10.1006/brcg.1994.1045
- Brysbaert, M., & D’Ydewalle, G. (1990). Tachistoscopic presentation of verbal stimuli for assessing cerebral dominance: Reliability data and some practical recommendations. Neuropsychologia, 28(5), 443–455. doi:https://doi.org/10.1016/0028-3932(90)90071-U
- Brysbaert, M., & Stevens, M. (2018). Power analysis and effect size in mixed effects models: A tutorial. Journal of Cognition, 1(1), 9. doi:10.5334/joc.10
- Buffery, A., & Gray, J. (1972). Sex differences in the development of spatial and linguistic skills. In C. Ounsted, & D. Taylor (Eds.), Gender differences, their ontogeny and significance (pp. 123–158). London: Churchill Livingstone.
- Button, K. S., Ioannidis, J. P. A., Mokrysz, C., Nosek, B. A., Flint, J., Robinson, E. S. J., & Munafo, M. R. (2013). Power failure: Why small sample size undermines the reliability of neuroscience. Nature Reviews Neuroscience, 14(5), 365–376. doi: 10.1038/nrn3475
- Cabeza, R. (2002). Hemispheric asymmetry reduction in older adults: The HAROLD model. Psychology and Aging, 17(1), 85–100. doi: 10.1037//0882-7974.17.1.85
- Cahill, L. (2006). Why sex matters for neuroscience. Nature Reviews Neuroscience, 7(6), 477–484.
- Carey, D. P., & Johnstone, L. T. (2014). Quantifying cerebral asymmetries for language in dextrals and adextrals with random-effects meta analysis. Frontiers in Psychology, 5, 1446. doi: 10.3389/fpsyg.2014.01128
- Casey, M. B., Nuttall, R. L., & Pezaris, E. (1997). Mediators of gender differences in mathematics college entrance test scores: A comparison of spatial skills with internalized beliefs and anxieties. Developmental Psychology, 33(4), 669–680. doi: 10.1037/0012-1649.33.4.669
- *Catani, M., Allin, M. P. G., Husain, M., Pugliese, L., Mesulam, M. M., Murray, R. M., & Jones, D. K. (2007). Symmetries in human brain language pathways correlate with verbal recall. Proceedings of the National Academy of Sciences of the United States of America, 104(43), 17163–17168.
- Ceci, S. J., Williams, W. M., & Barnett, S. M. (2009). Women’s underrepresentation in science: Sociocultural and biological considerations. Psychological Bulletin, 135(2), 218–261.
- Chance, S. A., Casanova, M. F., Switala, A. E., & Crow, T. J. (2006). Minicolumnar structure in Heschl’s gyrus and planum temporale: Asymmetries in relation to sex and callosal fiber number. Neuroscience, 143(4), 1041–1050. doi: 10.1016/j.neuroscience.2006.08.057
- Chiarello, C., Welcome, S. E., Halderman, L. K., Towler, S., Julagay, J., Otto, R., & Leonard, C. M. (2009). A large-scale investigation of lateralization in cortical anatomy and word reading: Are there Sex differences? Neuropsychology, 23(2), 210-222. doi: 10.1037/a0014265
- *Chiarello, C., Welcome, S. E., Halderman, L. K., & Leonard, C. M. (2009). Does degree of asymmetry relate to performance? An investigation of word recognition and reading in consistent and mixed handers. Brain and Cognition, 69(3), 521–530.
- Choi, J., & L’Hirondelle, N. (2005). Object location memory: A direct test of the verbal memory hypothesis. Learning and Individual Differences, 15(3), 237–245. doi:10.1016/j.lindif.2005.02.001
- *Clements, A. M., Rimrodt, S. L., Abel, J. R., Blankner, J. G., Mostofsky, S. H., Pekar, J. J., & Cutting, L. E. (2006). Sex differences in cerebral laterality of language and visuospatial processing. Brain and Language, 98(2), 150–158. doi:10.1016/j.bandl.2006.04.007
- Corballis, M. C., Hattie, J., & Fletcher, R. (2008). Handedness and intellectual achievement: An even-handed look. Neuropsychologia, 46(1), 374–378. doi:10.1016/j.neuropsychologia.2007.09.009
- Crichton-Browne, J. (1879). On the weight of the brain and its component parts in the insane. Brain, 2(1), 42–67. doi: 10.1093/brain/2.1.42
- Crow, T. J., Crow, L. R., Done, D. J., & Leask, S. (1998). Relative hand skill predicts academic ability: Global deficits at the point of hemispheric indecision. Neuropsychologia, 36(12), 1275–1282.
- Dadda, M., & Bisazza, A. (2006). Does brain asymmetry allow efficient performance of simultaneous tasks? Animal Behaviour, 72, 523–529. doi: 10.1016/j.anbehav.2005.10.019
- Davidson, R. J. (1995). Cerebral asymmetry, emotion, and affective style. In R. J. Davidson, & K. Hugdahl (Eds.), Brain asymmetry (pp. 361–387). Cambridge, MA: MIT Press.
- Davidson, R. J., & Hugdahl, K. (1995). Brain asymmetry. Cambridge, MA: MIT Press.
- Dax, M. (1836). Lesions de la moitie gauche de l’encephale soincidant avec l’oubli des signes de la pensee. Paper presented at the Congres meridional tenu a Montpellier, Montpellier.
- Del Giudice, M., Booth, T., & Irwing, P. (2012). The distance between Mars and Venus: Measuring global Sex differences in personality. PLoS ONE, 7(1), e29265. doi: 10.1371/journal.pone.0029265
- *Deutsch, G., Bourbon, W. T., Papanicolaou, A. C., & Eisenberg, H. M. (1988). Visuospatial tasks compared via activation of regional cerebral blood flow. Neuropsychologia, 26(3), 445–452. doi:10.1016/0028-3932(88)90097-8
- *Dollinger, S. M. C. (1995). Mental rotation performance - age, sex, and visual-field differences. Developmental Neuropsychology, 11(2), 215–222.
- Eals, M., & Silverman, I. (1994). The hunter-gatherer theory of spatial sex-differences - proximate factors mediating the female advantage in recall of object arrays. Ethology and Sociobiology, 15(2), 95–105.
- Else-Quest, N. M., Hyde, J. S., & Linn, M. C. (2010). Cross-National patterns of gender differences in mathematics: A meta-analysis. Psychological Bulletin, 136(1), 103–127. doi: 10.1037/a0018053
- Everts, R., Lidzba, K., Wilke, M., Kiefer, C., Mordasini, M., Schroth, G., & Steinlin, M. (2009). Strengthening of laterality of verbal and visuospatial functions during childhood and adolescence. Human Brain Mapping, 30(2), 473–483. doi: 10.1002/hbm.20523
- Faul, F., Erdfelder, E., Buchner, A., & Lang, A.-G. (2009). Statistical power analyses using G*power 3.1: Tests for correlation and regression analyses. Behavior Research Methods, 41(4), 1149–1160. doi: 10.3758/brm.41.4.1149
- Feingold, A. (1988). Cognitive gender differences are disappearing. American Psychologist, 43(2), 95–103. doi: 10.1037/0003-066X.43.2.95
- Feng, J., Spence, I., & Pratt, J. (2007). Playing an action video game reduces gender differences in spatial cognition. Psychological Science, 18(10), 850–855. doi: 10.1111/j.1467-9280.2007.01990.x
- *Frings, L., Wagner, K., Unterrainer, J., Spreer, J., Halsband, U., & Schulze-Bonhage, A. (2006). Gender-related differences in lateralization of hippocampal activation and cognitive strategy. Neuroreport, 17(4), 417–421. doi: 10.1097/01.wnr.0000203623.02082.e3
- Frost, J. A., Binder, J. R., Springer, J. A., Hammeke, T. A., Bellgowan, P. S. F., Rao, S. M., & Cox, R. W. (1999). Language processing is strongly left lateralized in both sexes: Evidence from functional MRI. Brain, 122, 199–208. doi: 10.1093/brain/122.2.199
- Garcia-Falgueras, A., & Swaab, D. F. (2008). A sex difference in the hypothalamic uncinate nucleus: Relationship to gender identity. Brain, 131(12), 3132–3146. doi: 10.1093/brain/awn276
- Gazzaniga, M. S., Bogen, J. E., & Sperry, R. W. (1962). Some functional effects of sectioning the cerebral commissures in man. Proceedings of the National Academy of Sciences, 48(10), 1765–1769.
- Geary, D. C. (1996). Sexual selection and sex differences in mathematical abilities. Behavioral and Brain Sciences, 19(2), 229–284.
- Gelman, A., & Carlin, J. (2014). Beyond power calculations: Assessing type S (sign) and type M (magnitude) errors. Perspectives on Psychological Science, 9(6), 641–651. doi: 10.1177/1745691614551642
- Geschwind, N., & Galaburda, A. M. (1985). Cerebral lateralization: Biological mechanisms, associations, and pathology: I. A hypothesis and a program for research. Archives of Neurology, 42(5), 428–459. doi: 10.1001/archneur.1985.04060050026008
- Giner-Sorolla, R. (2018). Powering your interaction. Retrieved from https://approachingblog.wordpress.com/2018/01/24/powering-your-interaction-2/
- *Gootjes, L., Bruggeling, E. C., Magnee, T., & Van Strien, J. W. (2008). Sex differences in the latency of the late event-related potential mental rotation effect. Neuroreport, 19(3), 349–353.
- Gotts, S. J., Jo, H. J., Wallace, G. L., Saad, Z. S., Cox, R. W., & Martin, A. (2013). Two distinct forms of functional lateralization in the human brain. Proceedings of the National Academy of Sciences, 110(36), E3435–E3444. doi: 10.1073/pnas.1302581110
- Grabowska, A. (2017). Sex on the brain: Are gender-dependent structural and functional differences associated with behavior? Journal of Neuroscience Research, 95(1-2), 200-212. doi: 10.1002/jnr.23953
- Groen, M. A., Whitehouse, A. J. O., Badcock, N. A., & Bishop, D. V. M. (2012). Does cerebral lateralization develop? A study using functional transcranial Doppler ultrasound assessing lateralization for language production and visuospatial memory. Brain and Behavior, 2(3), 256–269. doi: 10.1002/brb3.56
- Guadalupe, T., Zwiers, M. P., Wittfeld, K., Teumer, A., Vasquez, A. A., Hoogman, M., & Francks, C. (2015). Asymmetry within and around the human planum temporale is sexually dimorphic and influenced by genes involved in steroid hormone receptor activity. Cortex, 62(0), 41–55. doi:10.1016/j.cortex.2014.07.015
- Gur, R. E., & Gur, R. C. (2016). Sex differences in brain and behavior in adolescence: Findings from the Philadelphia neurodevelopmental cohort. Neuroscience & Biobehavioral Reviews, 70, 159–170. doi:10.1016/j.neubiorev.2016.07.035
- *Gur, R. C., Alsop, D., Glahn, D., Petty, R., Swanson, C. L., Maldjian, J. A., & Gur, R. E. (2000). An fMRI study of sex differences in regional activation to a verbal and a spatial task. Brain and Language, 74(2), 157–170. doi: 10.1006/brln.2000.2325
- Gur, R. C., Turetsky, B. I., Matsui, M., Yan, M., Bilker, W., Hughett, P., & Gur, R. E. (1999). Sex differences in brain gray and white matter in healthy young adults: Correlations with cognitive performance. Journal of Neuroscience, 19(10), 4065–4072.
- *Hahn, N., Jansen, P., & Heil, M. (2010). Preschoolers’ mental rotation of letters: Sex differences in hemispheric asymmetry. Cognitive Neuroscience, 1(4), 261–267. doi: 10.1080/17588928.2010.485248
- Haier, R. J., Karama, S., Leyba, L., & Jung, R. E. (2009). MRI assessment of cortical thickness and functional activity changes in adolescent girls following three months of practice on a visual-spatial task. BMC Research Notes, 2(1), 174. doi: 10.1186/1756-0500-2-174
- *Halari, R., Sharma, T., Hines, M., Andrew, C., Simmons, A., & Kumari, V. (2006). Comparable fMRI activity with differential behavioural performance on mental rotation and overt verbal fluency tasks in healthy men and women. Experimental Brain Research, 169(1), 1–14. doi: 10.1007/s00221-005-0118-7
- Halpern, D. F. (2012). Sex differences in cognitive abilities (4th ed.). New York, NY: Psychology Press, Taylor and Francis Group.
- Halpern, D. F., Eliot, L., Bigler, R. S., Fabes, R. A., Hanish, L. D., Hyde, J. S., & Martin, C. L. (2011). The pseudoscience of single-sex schooling. Science, 333(6050), 1706–1707. doi: 10.1126/science.1205031
- Halpern, D. F., & Tan, U. (2001). Stereotypes and steroids: Using a psychobiosocial model to understand cognitive sex differences. Brain and Cognition, 45(3), 392–414.
- *Hao, X., Wang, K. C., Li, W. F., Yang, W. J., Wei, D. T., Qiu, J., & Zhang, Q. L. (2013). Individual differences in brain structure and resting brain function underlie cognitive styles: Evidence from the embedded figures test. PLoS ONE, 8(12), doi: 10.1371/journal.pone.0078089
- Harshman, R. A., Hampson, E., & Berenbaum, S. A. (1983). Individual-differences in cognitive-abilities and brain organization .1. Sex and handedness differences in ability. Canadian Journal of Psychology/Revue Canadienne de Psychologie, 37(1), 144–192. doi: 10.1037/h0080690
- *Hattemer, K., Plate, A., Heverhagen, J. T., Haag, A., Keil, B., Klein, K. M., & Knake, S. (2011). Determination of hemispheric dominance with mental rotation using functional transcranial Doppler sonography and FMRI. Journal of Neuroimaging, 21(1), 16–23. doi: 10.1111/j.1552-6569.2009.00402.x
- Hausmann, M. (2017). Why sex hormones matter for neuroscience: A very short review on sex, sex hormones, and functional brain asymmetries. Journal of Neuroscience Research, 95(1-2), 40–49. doi: 10.1002/jnr.23857
- Hausmann, M., & Bayer, U. (2010). Sex hormonal effects on hemispheric asymmetry and interhemispheric interaction. In K. Hugdahl, & R. Westerhausen (Eds.), The Two halves of the brain. Information processing in the cerebral hemispheres (pp. 253–286). Cambridge, MA: MIT press.
- Hausmann, M., Becker, C., Gather, U., & Güntürkün, O. (2002). Functional cerebral asymmetries during the menstrual cycle: A cross-sectional and longitudinal analysis. Neuropsychologia, 40(7), 808–816.
- Hausmann, M., Behrendt-Korbitz, S., Kautz, H., Lamm, C., Radelt, F., & Gunturkun, O. (1998). Sex differences in oral asymmetries during wordrepetition. Neuropsychologia, 36(12), 1397–1402. doi: 10.1016/s0028-3932(98)00027-x
- Hausmann, M., Schoofs, D., Rosenthal, H. E., & Jordan, K. (2009). Interactive effects of sex hormones and gender stereotypes on cognitive sex differences-A psychobiosocial approach. Psychoneuroendocrinology, 34(3), 389–401.
- Hedges, L. V., & Nowell, A. (1995). Sex-differences in mental test-scores, variability, and numbers of high-scoring individuals. Science, 269(5220), 41–45. doi: 10.1126/science.7604277
- Hellige, J. B. (1993). Hemispheric asymmetry: What’s right and what’s left. Cambridge, MA: Harvard University Press.
- Herlitz, A., Reuterskiöld, L., Lovén, J., Thilers, P. P., & Rehnman, J. (2013). Cognitive Sex differences Are Not magnified as a function of age, sex hormones, or puberty development during early adolescence. Developmental Neuropsychology, 38(3), 167–179. doi: 10.1080/87565641.2012.759580
- Hines, M., & Gorski, B. (1985). Hormonal influences on the development of neural asymmetries. In D. F. Benson, & E. Zaidel (Eds.), The dual brain: Hemispheric specialization in humans (pp. 75–96). New York, NY: Guildford.
- Hirnstein, M., Hausmann, M., & Güntürkün, O. (2008). The evolutionary origins of functional cerebral asymmetries in humans: Does lateralization enhance parallel processing? Behavioural Brain Research, 187(2), 297-303.
- Hirnstein, M., Hugdahl, K., & Hausmann, M. (2014). How brain asymmetry relates to performance - a large-scale dichotic listening study. Frontiers in Psychology, 4. doi: 10.3389/fpsyg.2013.00997
- Hirnstein, M., Leask, S., Rose, J., & Hausmann, M. (2010). Disentangling the relationship between hemispheric asymmetry and cognitive performance. Brain and Cognition, 73(2), 119–127.
- Hirnstein, M., Westerhausen, R., Korsnes, M. S., & Hugdahl, K. (2013). Sex differences in language asymmetry are age-dependent and small: A large-scale, consonant-vowel dichotic listening study with behavioral and fMRI data. Cortex, 49(7), 1910–1921. doi: 10.1016/j.cortex.2012.08.002
- Hiscock, M., Inch, R., Hawryluk, J., Lyon, P. J., & Perachio, N. (1999). Is there a sex difference in human laterality? III. An exhaustive survey of tactile laterality studies from six neuropsychology journals. Journal of Clinical and Experimental Neuropsychology (Neuropsychology, Development and Cognition: Section A), 21(1), 17–28. doi: 10.1076/jcen.21.1.17.944
- Hiscock, M., Inch, R., Jacek, C., Hiscockkalil, C., & Kalil, K. M. (1994). Is there a sex difference in human laterality? I. An exhaustive survey of auditory laterality studies from 6 neuropsychology journals. Journal of Clinical and Experimental Neuropsychology, 16(3), 423–435. doi: 10.1080/01688639408402653
- Hiscock, M., Israelian, M., Inch, R., Jacek, C., & Hiscockkalil, C. (1995). Is there a sex difference in human laterality? II. An exhaustive survey of visual laterality studies from 6 neuropsychology journals. Journal of Clinical and Experimental Neuropsychology, 17(4), 590–610. doi: 10.1080/01688639508405148
- Hiscock, M., Perachio, N., & Inch, R. (2001). Is there a sex difference in human laterality? IV. An exhaustive survey of dual-task interference studies from six neuropsychology journals. Journal of Clinical and Experimental Neuropsychology (Neuropsychology, Development and Cognition: Section A), 23(2), 137–148. doi: 10.1076/jcen.23.2.137.1206
- Hugdahl, K. (1995). Dichotic listening: Probing temporal lobe functional integrity. In R. J. Davidson, & K. Hugdahl (Eds.), Brain asymmetry (pp. 123–156). Cambridge, MA: MIT Press.
- Hugdahl, K. (2003). Dichotic listening in the study of auditory laterality. In K. Hugdahl, & R. J. Davidson (Eds.), The asymmetrical brain (pp. 441–478). Cambridge, MA: MIT Press.
- *Hugdahl, K., Thomsen, T., & Ersland, L. (2006). Sex differences in visuo-spatial processing: An fMRI study of mental rotation. Neuropsychologia, 44(9), 1575–1583. doi: 10.1016/j.neuropsychologia.2006.01.026
- Hunter, Z. R., & Brysbaert, M. (2008). Visual half-field experiments are a good measure of cerebral language dominance if used properly: Evidence from fMRI. Neuropsychologia, 46(1), 316–325.
- Hyde, J. S. (2005). The gender similarities hypothesis. American Psychologist, 60(6), 581–592.
- Hyde, J. S. (2014). Gender similarities and differences. Annual Review of Psychology, 65(1), 373–398. doi: 10.1146/annurev-psych-010213-115057
- Hyde, J. S., Lindberg, S. M., Linn, M. C., Ellis, A. B., & Williams, C. C. (2008). Diversity - gender similarities characterize math performance. Science, 321(5888), 494–495. doi: 10.1126/science.1160364
- Hyde, J. S., & Linn, M. C. (1988). Gender differences in verbal-ability - a meta-analysis. Psychological Bulletin, 104(1), 53–69.
- Jackson, J. H. (1864). Notes on the physiology and pathology of the nervous system. “coarse” disease of the brain and optic neuritis. In J. Taylor (Ed.), Selected writings og John Hughlings Jackson (pp. 215–237). New York, NY: Basic Books Inc.
- Jäncke, Lutz. (2018). Sex/gender differences in cognition, neurophysiology, and neuroanatomy. F1000Research, 7, 805. doi: 10.12688/f1000research
- Jansen, P., & Heil, M. (2010). Gender differences in mental rotation across adulthood. Experimental Aging Research, 36(1), 94–104. doi: 10.1080/03610730903422762
- *Johnson, B. W., McKenzie, K. J., & Hamm, J. P. (2002). Cerebral asymmetry for mental rotation: Effects of response hand, handedness and gender. Neuroreport, 13(15), 1929–1932. doi: 10.1097/00001756-200210280-00020
- Johnston, D. W., Nicholls, M. E., Shah, M., & Shields, M. A. (2009). Nature’s experiment? Handedness and early childhood development. Demography, 46(2), 281–301. doi: 10.1353/dem.0.0053
- *Jones, B., & Anuza, T. (1982). Effects of sex, handedness, stimulus and visual field on “mental rotation”. Cortex, 18(4), 501–514.
- Jordan, K., Wüstenberg, T., Heinze, H. J., Peters, M., & Jancke, L. (2002). Women and men exhibit different cortical activation patterns during mental rotation tasks. Neuropsychologia, 40(13), 2397–2408.
- *Kail, R. V., Jr., & Siegel, A. W. (1978). Sex and hemispheric differences in the recall of verbal and spatial information. Cortex, 14(4), 557–563.
- Kaiser, A., Haller, S., Schmitz, S., & Nitsch, C. (2009). On sex/gender related similarities and differences in fMRI language research. Brain Research Reviews, 61(2), 49–59. doi: 10.1016/j.brainresrev.2009.03.005
- Kaiser, A., Kuenzli, E., Zappatore, D., & Nitsch, C. (2007). On females’ lateral and males’ bilateral activation during language production: A fMRI study. International Journal of Psychophysiology, 63(2), 192–198. doi: 10.1016/j.ijpsycho.2006.03.008
- Kimura, D. (2000). Sex and cognition. Cambridge, MA: MIT Press.
- Knecht, S., Drager, B., Deppe, M., Bobe, L., Lohmann, H., Floel, A., & Henningsen, H. (2000). Handedness and hemispheric language dominance in healthy humans. Brain, 123, 2512–2518.
- Kramer, J. H., Delis, D. C., Kaplan, E., Odonnell, L., & Prifitera, A. (1997). Developmental sex differences in verbal learning. Neuropsychology, 11(4), 577–584. doi: 10.1037/0894-4105.11.4.577
- Ladavas, E., Umiltà, C., & Ricci-Bitti, P. E. (1980). Evidence for sex differences in right-hemisphere dominance for emotions. Neuropsychologia, 18(3), 361–366. doi:10.1016/0028-3932(80)90132-3
- Lansdell, H. (1961). The effect of neurosurgery on a test of proverbs. American Psychologist, 16(7), 448.
- Lansdell, H. (1962). A sex difference in effect of temporal-lobe neurosurgery on design preference. Nature, 194(4831), 852–854. doi: 10.1038/194852a0
- Leask, S. J., & Crow, T. J. (2006). A single optimum degree of hemispheric specialisation in two tasks, in two UK national birth cohorts. Brain and Cognition, 62(3), 221–227.
- Levy, J. (1969). Possible basis for the evolution of lateral specialization of the human brain. Nature, 224(5219), 614–615.
- Levy, J. (1972). Lateral specialization of the human brain: Behavioral manifestations and possible evolutionary basis. In J. A. J. Kiger (Ed.), The biology of behavior (pp. 159–180). Corvallis, OR: Oregon State University Press.
- Levy, J. (1974). Psychobiological implications of bilateral asymmetry. In S. Dimond, & J. G. Beaumont (Eds.), Hemisphere function in the human brain (pp. 121–183). London: Paul Elek.
- Levy, J. (1978). Lateral differences in the human brain in cognition and behavioral control. In P. Buser, & A. Rougeul-Buser (Eds.), Cerebral correlates of conscious experience (pp. 285–298). New York, NY: North Holland Publishin Co.
- Linn, M. C., & Petersen, A. C. (1985). Emergence and characterization of sex-differences in spatial ability - a meta-analysis. Child Development, 56(6), 1479–1498.
- Lippa, A., Collaer, M. L., & Peters, M. (2010). Sex differences in mental rotation and line angle judgments Are positively associated with gender equality and economic development across 53 nations. Archives of Sexual Behavior, 39(4), 990–997. doi: 10.1007/s10508-008-9460-8
- Lowe, P. A., Mayfield, J. W., & Reynolds, C. R. (2003). Gender differences in memory test performance among children and adolescents. Archives of Clinical Neuropsychology, 18(8), 865–878. doi:10.1016/S0887-6177(02)00162-2
- Lust, J. M., Geuze, R. H., Groothuis, A. G. G., & Bouma, A. (2011). Functional cerebral lateralization and dual-task efficiency--testing the function of human brain lateralization using fTCD. Behavioural Brain Research, 217(2), 293–301.
- Lust, J. M., Geuze, R. H., Groothuis, A. G. G., van der Zwan, J. E., Brouwer, W. H., van Wolffelaar, P. C., & Bouma, A. (2011). Driving performance during word generation--testing the function of human brain lateralization using fTCD in an ecologically relevant context. Neuropsychologia, 49(9), 2375–2383.
- MacFarlane Smith, I. (1964). Spatial ability. Its educational and social significance. London: University of London Press.
- McCarthy, M. M. (2016). Multifaceted origins of sex differences in the brain. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1688), 20150106. doi: 10.1098/rstb.2015.0106
- McGlone, J. (1980). Sex-differences in human-brain asymmetry - a critical survey. Behavioral and Brain Sciences, 3(2), 215–227.
- McManus, I. C., Shergill, S., & Bryden, M. P. (1993). Annett’s theory that individuals heterozygous for the right shift gene are intellectually advantaged: Theoretical and empirical problems. British Journal of Psychology, 84(Pt 4), 517–537.
- Mead, L. A., & Hampson, E. (1996). Asymmetric effects of ovarian hormones on hemispheric activity: Evidence from dichotic and tachistoscopic tests. Neuropsychology, 10(4), 578–587.
- Mellet, E., Jobard, G., Zago, L., Crivello, F., Petit, L., Joliot, M., & Tzourio-Mazoyer, N. (2014). Relationships between hand laterality and verbal and spatial skills in 436 healthy adults balanced for handedness. Laterality: Asymmetries of Body, Brain and Cognition, 19(4), 383–404. doi: 10.1080/1357650x.2013.796965
- Mellet, E., Zago, L., Jobard, G., Crivello, F., Petit, L., Joliot, M., & Tzourio-Mazoyer, N. (2014). Weak language lateralization affects both verbal and spatial skills: An fMRI study in 297 subjects. Neuropsychologia, 65(0), 56–62. doi:10.1016/j.neuropsychologia.2014.10.010
- Miller, D. I., & Halpern, D. F. (2014). The new science of cognitive sex differences. Trends in Cognitive Sciences, 18(1), 37–45. doi:10.1016/j.tics.2013.10.011
- Moher, D., Liberati, A., Tetzlaff, J., Altman, D. G., & The Prisma Group. (2009). Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLOS Medicine, 6(7), e1000097. doi: 10.1371/journal.pmed.1000097
- Nicholls, M. E., Chapman, H. L., Loetscher, T., & Grimshaw, G. M. (2010). The relationship between hand preference, hand performance, and general cognitive ability. Journal of the International Neuropsychological Society, 16(04), 585–592. doi: 10.1017/S1355617710000184
- Ntolka, E., & Papadatou-Pastou, M. (2018). Right-handers have negligibly higher IQ scores than left-handers: Systematic review and meta-analyses. Neuroscience & Biobehavioral Reviews, 84, 376–393. doi: 10.1016/j.neubiorev.2017.08.007
- O’Boyle, M. W., Cunnington, R., Silk, T. J., Vaughan, D., Jackson, G., Syngeniotis, A., & Egan, G. F. (2005). Mathematically gifted male adolescents activate a unique brain network during mental rotation. Cognitive Brain Research, 25(2), 583–587.
- Ocklenburg, S., Beste, C., Arning, L., Peterburs, J., & Güntürkün, O. (2014). The ontogenesis of language lateralization and its relation to handedness. Neuroscience & Biobehavioral Reviews, 43(0), 191–198. doi:10.1016/j.neubiorev.2014.04.008
- Ocklenburg, S., & Güntürkün, O. (2018). The lateralized brain - The neuroscience and evolution of hemispheric asymmetries. London, UK: Academic Press.
- Papousek, I., & Schulter, G. (2002). Covariations of EEG asymmetries and emotional states indicate that activity at frontopolar locations is particularly affected by state factors. Psychophysiology, 39(3), 350–360. doi: 10.1017/s0048577201393083
- *Papousek, I., Murhammer, D., & Schulter, G. (2011). Intra- and interindividual differences in lateralized cognitive performance and asymmetrical EEG activity in the frontal cortex. Brain and Cognition, 75(3), 225–231. doi: 10.1016/j.bandc.2010.11.013
- *Pellkofer, J., Jansen, P., & Heil, M. (2014). Sex-specific lateralization of event-related potential effects during mental rotation of polygons. Neuroreport, 25(11), 848–853. doi: 10.1097/wnr.0000000000000198
- *Persson, J., Herlitz, A., Engman, J., Morell, A., Sjolie, D., Wikstrom, J., & Soderlund, H. (2013). Remembering our origin: Gender differences in spatial memory are reflected in gender differences in hippocampal lateralization. Behavioural Brain Research, 256, 219–228. doi: 10.1016/j.bbr.2013.07.050
- Peters, M. (2005). Sex differences and the factor of time in solving Vandenberg and Kuse mental rotation problems. Brain and Cognition, 57(2), 176–184.
- Peters, M., Laeng, B., Latham, K., Jackson, M., Zaiyouna, R., & Richardson, C. (1995). A redrawn Vandenberg and Kuse mental rotations test: Different versions and factors that affect performance. Brain and Cognition, 28(1), 39–58.
- Peters, M., Reimers, S., & Manning, J. T. (2006). Hand preference for writing and associations with selected demographic and behavioral variables in 255,100 subjects: The BBC internet study. Brain and Cognition, 62(2), 177–189.
- Pfannkuche, K. A., Bouma, A., & Groothuis, T. G. (2009). Does testosterone affect lateralization of brain and behaviour? A meta-analysis in humans and other animal species. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 364(1519), 929–942. doi:364/1519/929 [pii] 10.1098/rstb.2008.0282
- Plessen, K. J., Hugdahl, K., Bansal, R., Hao, X., & Peterson, B. S. (2014). Sex, Age, and cognitive correlates of asymmetries in thickness of the cortical mantle across the life span. Journal of Neuroscience, 34(18), 6294–6302. doi: 10.1523/jneurosci.3692-13.2014
- Pletzer, B. (2014). Sex-specific strategy use and global-local processing: A perspective toward integrating sex differences in cognition. Frontiers in Neuroscience, 8, 81. doi: 10.3389/fnins.2014.00425
- Powell, J. L., Kemp, G. J., & García-Finaña, M. (2012). Association between language and spatial laterality and cognitive ability: An fMRI study. Neuroimage, 59(2), 1818–1829. doi:10.1016/j.neuroimage.2011.08.040
- Proverbio, A. M., Riva, F., Martin, E., & Zani, A. (2010). Face coding is bilateral in the female brain. PLoS ONE, 5(6), e11242.
- *Ragland, J. D., Coleman, A. R., Gur, R. C., Glahn, D. C., & Gur, R. E. (2000). Sex differences in brain-behavior relationships between verbal episodic memory and resting regional cerebral blood flow. Neuropsychologia, 38(4), 451–461. doi: 10.1016/s0028-3932(99)00086-x
- Rasmussen, T., & Milner, B. (1977). The role of early left-brain injury in determining lateralization of cerebral speech functions. Annals of the New York Academy of Sciences, 299, 355–369.
- Razafimandimby, A., Tzourio-Mazoyer, N., Mazoyer, B., Maiza, O., & Dollfus, S. (2011). Language lateralization in left-handed patients with schizophrenia. Neuropsychologia, 49(3), 313–319. doi: 10.1016/j.neuropsychologia.2010.11.025
- References marked with an asterisk (*) are included in the systematic review.
- Reilly, D. (2012). Gender, culture, and Sex-typed cognitive abilities. PLoS ONE, 7(7), e39904. doi: 10.1371/journal.pone.0039904
- Rilea, S. L. (2008a). A lateralization of function approach to sex differences in spatial ability: A reexamination. Brain and Cognition, 67(2), 168–182.
- *Rilea, S. L. (2008b). Sex and hemisphere differences when mentally rotating meaningful and meaningless stimuli. Laterality, 13(3), 217–233.
- *Rilea, S. L., Roskos-Ewoldsen, B., & Boles, D. B. (2004). Sex differences in spatial ability: A lateralization of function approach. Brain and Cognition, 56(3), 332–343. doi:http://doi.org/10.1016/j.bandc.2004.09.002
- Rippon, G., Jordan-Young, R., Kaiser, A., & Fine, C. (2014). Recommendations for sex/gender neuroimaging research: Key principles and implications for research design, analysis and interpretation. Frontiers in Human Neuroscience, 8, 1244. doi: 10.3389/fnhum.2014.00650
- Roalf, D. R., Gur, R. E., Ruparel, K., Calkins, M. E., Satterthwaite, T. D., Bilker, W. B., & Gur, R. C. (2014). Within-individual variability in neurocognitive performance: Age- and sex-related differences in children and youths from ages 8 to 21. Neuropsychology, 28(4), 506–518. doi: 10.1037/neu0000067
- *Roberts, J. E., & Bell, M. A. (2002). The effects of age and sex on mental rotation performance, verbal performance, and brain electrical activity. Developmental Psychobiology, 40(4), 391–407. doi: 10.1002/dev.10039
- *Roberts, J. E., & Bell, M. A. (2003). Two- and three-dimensional mental rotation tasks lead to different parietal laterality for men and women. International Journal of Psychophysiology, 50(3), 235–246. doi: 10.1016/s0167-8760(03)00195-8
- Rogers, L. J. (2014). Asymmetry of brain and behavior in animals: Its development, function, and human relevance. Genesis, 52(6), 555–571. doi: 10.1002/dvg.22741
- Rogers, L. J., Zucca, P., & Vallortigara, G. (2004). Advantages of having a lateralized brain. Proceedings of the Royal Society of London Series B-Biological Sciences, 271, S420–S422.
- Rosenthal, R., & Rubin, D. B. (1982). Further meta-analytical procedures for assessing cognitive gender differences. Journal of Educational Psychology, 74(5), 708–712. doi: 10.1037//0022-0663.74.5.708
- Rosselli, M., Ardila, A., Matute, E., & Velez-Uribe, I. (2014). Language development across the life span: A neuropsychological/neuroimaging perspective. Neuroscience Journal, 2014, 1–21. doi: 10.1155/2014/585237
- Sanders, G., & Wenmoth, D. (1998). Verbal and music dichotic listening tasks reveal variations in functional cerebral asymmetry across the menstrual cycle that are phase and task dependent. Neuropsychologia, 36(9), 869–874. doi: 10.1016/s0028-3932(98)00022-0
- Satterthwaite, T. D., Wolf, D. H., Roalf, D. R., Ruparel, K., Erus, G., Vandekar, S., & Gur, R. C. (2015). Linked Sex differences in cognition and functional connectivity in youth. Cerebral Cortex, 25(9), 2383–2394. doi: 10.1093/cercor/bhu036
- Semrud-Clikeman, M., Fine, J. G., Bledsoe, J., & Zhu, D. C. (2012). Gender differences in brain activation on a mental rotation task. International Journal of Neuroscience, 122(10), 590–597. doi: 10.3109/00207454.2012.693999
- Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Constable, R. T., Skudlarski, P., Fulbright, R. K., & Gore, J. C. (1995). Sex-Differences in the functional-organization of the brain for language. Nature, 373(6515), 607–609.
- Shepard, R. N., & Feng, C. (1972). A chronometric study of mental paper folding. Cognitive Psychology, 3(2), 228–243. doi:10.1016/0010-0285(72)90005-9
- Shepard, R. N., & Metzler, J. (1971). Mental rotation of 3-dimensional objects. Science, 171(3972), 701–703. doi: 10.1126/science.171.3972.701
- Sherman, J. (1979). Cognitive performance as a function of Sex and handedness: An evaluation of the levy hypothesis. Psychology of Women Quarterly, 3(4), 378–390. doi: 10.1111/j.1471-6402.1979.tb00687.x
- Silverman, A. J., & Eals, M. (1992). Sex differences in spatial abilities: Evolutionary theory and data. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 533–549). New York, NY: Oxford University Press.
- Smith, L. L., & Hines, M. (2000). Language lateralization and handedness in women prenatally exposed to diethylstilbestrol (DES). Psychoneuroendocrinology, 25(5), 497–512. doi:10.1016/S0306-4530(00)00005-6
- Sommer, I. E. C. (2010). Sex differences in handedness, brain asymmetry, and language lateralization. In The Two halves of the brain: Information processing in the cerebral hemispheres (pp. 287–312). Cambridge, MA: MIT Press.
- Sommer, I. E. C., Aleman, A., Bouma, A., & Kahn, R. S. (2004). Do women really have more bilateral language representation than men? A meta-analysis of functional imaging studies. Brain, 127, 1845–1852. doi: 10.1093/brain/awh207
- Sommer, I. E. C., Aleman, A., Somers, M., Boks, M. P., & Kahn, R. S. (2008). Sex differences in handedness, asymmetry of the Planum Temporale and functional language lateralization. Brain Research, 1206, 76–88.
- Sperry, R. W. (1968). Hemisphere deconnection and unity in conscious awareness. American Psychologist, 23(10), 723–733. doi: 10.1037/h0026839
- Springer, S. P., & Deutsch, G. (1993). Left brain, right brain. (4th ed.). New York, NY: W. H. Freeman and Company.
- Stoet, G., & Geary, D. C. (2013). Sex differences in mathematics and reading achievement Are inversely related: Within- and across-nation assessment of 10 years of PISA data. PLoS ONE, 8(3), e57988. doi: 10.1371/journal.pone.0057988
- Terlecki, M. S., & Newcombe, N. S. (2005). How important Is the digital divide? The relation of computer and videogame usage to gender differences in mental rotation ability. Sex Roles, 53(5), 433–441. doi: 10.1007/s11199-005-6765-0
- Tunç, B., Solmaz, B., Parker, D., Satterthwaite, T. D., Elliott, M. A., Calkins, M. E., & Verma, R. (2016). Establishing a link between sex-related differences in the structural connectome and behaviour. Philosophical Transactions of the Royal Society B: Biological Sciences, 371(1688), 20150111. doi: 10.1098/rstb.2015.0111
- *Uecker, A., & Obrzut, J. E. (1993). Hemisphere and gender differences in mental rotation. Brain and Cognition, 22(1), 42–50. doi:10.1006/brcg.1993.1023
- *Unterrainer, J., Wranek, U., Staffen, W., Gruber, T., & Ladurner, G. (2000). Lateralized cognitive visuospatial processing: Is it primarily gender-related or due to quality of performance? A HMPAO-SPECT study. Neuropsychobiology, 41(2), 95–101. doi: 26639
- Uttal, D. H., Meadow, N. G., Tipton, E., Hand, L. L., Alden, A. R., Warren, C., & Newcombe, N. S. (2013). The malleability of spatial skills: A meta-analysis of training studies. Psychological Bulletin, 139(2), 352–402. doi: 10.1037/a0028446
- Vandenberg, S. G., & Kuse, A. R. (1978). Mental rotations, a group test of 3-dimensional spatial visualization. Perceptual and Motor Skills, 47(2), 599–604.
- Van der Haegen, L., Westerhausen, R., Hugdahl, K., & Brysbaert, M. (2013). Speech dominance is a better predictor of functional brain asymmetry than handedness: A combined fMRI word generation and behavioral dichotic listening study. Neuropsychologia, 51(1), 91–97. doi:10.1016/j.neuropsychologia.2012.11.002
- *van der Ham, I. J. M., & Borst, G. (2011). Individual differences in spatial relation processing: Effects of strategy, ability, and gender. Brain and Cognition, 76(1), 184–190. doi: 10.1016/j.bandc.2011.01.008
- van Ettinger-Veenstra, H. M., Ragnehed, M., Hallgren, M., Karlsson, T., Landtblom, A. M., Lundberg, P., & Engstrom, M. (2010). Right-hemispheric brain activation correlates to language performance. Neuroimage, 49(4), 3481–3488. doi: 10.1016/j.neuroimage.2009.10.041
- Vasta, R., & Liben, L. S. (1996). The water-level task: An intriguing puzzle. Current Directions in Psychological Science, 5(6), 171–177. doi: 10.1111/1467-8721.ep11512379
- Vogel, J. J., Bowers, C. A., & Vogel, D. S. (2003). Cerebral lateralization of spatial abilities: A meta-analysis. Brain and Cognition, 52(2), 197–204.
- Voyer, D. (1996). On the magnitude of laterality effects and Sex differences in functional lateralities. Laterality, 1(1), 51–84.
- Voyer, D. (1998). On the reliability and validity of noninvasive laterality measures. Brain and Cognition, 36(2), 209–236.
- Voyer, D. (2011). Sex differences in dichotic listening. Brain and Cognition, 76, 245–255.
- *Voyer, D. (1995). Effect of practice on laterality in a mental rotation task. Brain and Cognition, 29(3), 326–335. doi: 10.1006/brcg.1995.1285
- Voyer, D., Postma, A., Brake, B., & Imperato-McGinley, J. (2007). Gender differences in object location memory: A meta-analysis. Psychonomic Bulletin & Review, 14(1), 23–38.
- Voyer, S. D., & Voyer, D. (2015). Laterality, spatial abilities, and accident proneness. Journal of Clinical and Experimental Neuropsychology, 37(1), 27–36. doi: 10.1080/13803395.2014.985191
- Voyer, D., Voyer, S., & Bryden, M. P. (1995). Magnitude of sex-differences in spatial abilities - a metaanalysis and consideration of critical variables. Psychological Bulletin, 117(2), 250–270.
- *Voyer, D., & Voyer, S. D. (2014). Gender differences in scholastic achievement: A meta-analysis. Psychological Bulletin, 140(4), 1174–1204. doi: 10.1037/a0036620
- Waber, D. P. (1976). Sex differences in cognition: A function of maturation rate? Science, 192(4239), 572–574. doi: 10.1126/science.1257795
- Wai, J., Cacchio, M., Putallaz, M., & Makel, M. C. (2010). Sex differences in the right tail of cognitive abilities: A 30 year examination. Intelligence, 38(4), 412–423. doi: 10.1016/j.intell.2010.04.006
- *Walter, K. D., Roberts, A. E., & Brownlow, S. (2000). Spatial perception and mental rotation produce gender differences in cerebral hemovelocity: A TCD study. Journal of Psychophysiology, 14(1), 37–45. doi: 10.1027//0269-8803.14.1.37
- *Wegesin, D. J. (1998). Event-related potentials in homosexual and heterosexual men and women: Sex-dimorphic patterns in verbal asymmetries and mental rotation. Brain and Cognition, 36(1), 73–92. doi: 10.1006/brcg.1997.0964
- Weis, S., & Hausmann, M. (2010). Sex hormones: Modulators of interhemispheric inhibition in the human brain. The Neuroscientist, 16(2), 132–138. doi: 10.1177/1073858409341481
- Weis, S., Hausmann, M., Stoffers, B., Vohn, R., Kellermann, T., & Sturm, W. (2008). Estradiol modulates functional brain organization during the menstrual cycle: An analysis of interhemispheric inhibition. Journal of Neuroscience, 28(50), 13401–13410. doi: 10.1523/jneurosci.4392-08.2008
- *Wendt, P. E., & Risberg, J. (1994). Cortical activation during visual spatial processing: Relation between hemispheric asymmetry of blood flow and performance. Brain and Cognition, 24(1), 87–103. doi:10.1006/brcg.1994.1005
- Witelson, S. F. (1976). Sex and the single hemisphere: Specialization of the right hemisphere for spatial processing. Science (New York, N.Y.), 193(4251), 425–427. doi: 10.1126/science.935879
- Witelson, S. F. (1991). Neural sexual mosaicism: Sexual differentiation of the human temporo-parietal region for functional asymmetry. Psychoneuroendocrinology, 16(1-3), 131–153. doi: 10.1016/0306-4530(91)90075-5
- Witkin, H. A., Oltman, P. K., Raskin, E., & Karp, S. A. (1971). A manual for the embedded figures test. Palo Alto, CA: Consulting Psychologists Press.
- *Yu, Q. B., Tang, Y. Y., Li, J., Lu, Q. L., Wang, H. L., Sui, D., & Heil, M. (2009). Sex differences of event-related potential effects during three-dimensional mental rotation. Neuroreport, 20(1), 43–47. doi: 10.1097/WNR.0b013e32831c50f4
- *Zell, E., Krizan, Z., & Teeter, S. R. (2015). Evaluating gender similarities and differences using metasynthesis. American Psychologist, 70(1), 10–20. doi: 10.1037/a0038208