Abstract
Social insects like ants and bees live in cooperative colonies containing up to millions of individuals. These colonies are sometimes termed ‘superorganisms’ and have evolved tightly integrated and sophisticated collective behaviours. Different species, however, often differ in the type and mechanisms of communication and collective organization employed. I show here how individual-based models can be used to identify the non-intuitive benefits of different mechanisms of communication and division of labour and how these benefits may depend on the external environment as well as traits of the society itself. This allows us to understand under what ecological conditions particular types of collective organization may have evolved, and thus can also help to explain variation among species.
1. Introduction
Social insects, such as ants, bees and termites, live in associations called ‘eusocial colonies’. These differ from other animal groups, such as flocks of birds or most mammal groups, in that they exhibit reproductive division of labour: only one individual, the so-called queen, usually reproduces. All other individuals, called ‘workers’, are essentially sterile. The consequence of this is that colony members do not experience conflicting interests, since both worker and queen behaviours evolve to maximize collective success, that is, the queen's reproduction. While there are exceptions and worker reproduction can, rarely, occur, on the whole this means that these colonies resemble multicellular organisms with soma and germ cells, where selection is also largely on the collective. This means that social insect colonies have evolved sophisticated collective behaviours that require the cooperation and interaction of many individuals [Citation1].
I study these collective, and distributed, problem-solving mechanisms and their benefits, that is, the reasons for their evolution, here. This type of research is called ‘behavioural ecology’. In particular, I am interested in investigating why different species use different solutions to apparently similar problems. Are these different solutions tailored to subtle differences among species, such as group size or the parameters of the problem as usually encountered by a particular type of insect? Here, I quantify the performance of different possible strategies under different conditions and group sizes. I show that some, but not all, distributed problem-solving strategies are group-size sensitive and that the detailed parameters of the problem to be solved may have a big effect on which strategy is most suitable to solving it.
The problems solved by social insect colonies using distributed strategies can be very complex. For example, bee colonies collect food from flowers, whose abundance varies strongly in time and space. Finding these resources can require considerable search effort: bees will search an area of several kilometres diameter, and each forager will visit hundreds of flowers per day. In addition to the search itself, effective foraging by the colony depends on an appropriate allocation of bee workers to exploration (search for new resources) and exploitation (collection of food from known resources), and an effective allocation of foragers among resource patches, which may vary in quality, distance from the nest, longevity and reliability, leading to complex trade-offs [Citation2]. The actual number of workers allocated to search and exploitation of resources is a result of the individual decisions made by these workers, which in turn are based on information available to these individuals through their own search or through communicated information from others [Citation3]. The collective pattern is therefore self-organized, without a central leader or template directing individuals to particular resources. The role of communication in the process of resource collection is investigated in the first modelling study presented here. Resource collection is, however, only one example of the many complex problems that social insect colonies solve with self-organized behaviours. Another example is task allocation or division of labour: individual workers in a colony have to be flexibly allocated to different tasks according to need. This problem is discussed in the second modelling study presented here.
2. Research questions and approach
To quantify the benefits of different strategies in different environments, modelling is often necessary. In studying self-organized collective behaviour, such as foraging with or without communication, individual-based models are ideal for several reasons. First, it is straightforward to implement the behaviours of and constraints on individuals as known from empirical research. Second, spatially explicit environments are used, and thus constraints on movement and spatial heterogeneity in resources and behavioural or information heterogeneity among workers are taken into account [Citation4–7]. However, because this usually implies that a (relatively) large number of parameters are used, whose numerical values may have an impact on the predictions produced by the model, this necessitates the use of similar techniques as empirical research, namely a strict hypothesis-testing approach and extensive sensitivity analyses [Citation3,Citation8]. I will present the results of two modelling efforts, the first concerned with different communication systems and the second focusing on different mechanisms of task allocation within the nest.
3. Communication in foraging: resource distribution matters!
The degree to which communication is used in resource collection varies between species. Honey bees (genus Apis) can directly communicate distance and direction and pass on information about the odour of rewarding food sources to their nestmates using the ‘waggle dance’ [Citation8]. In addition, high availability and quality of resources cause increased frequency and duration of waggle dances, thus increasing the number of workers at those particular resources. Bumble bees (genus Bombus), on the other hand, do not communicate location information. Only the presence, availability and type of rewarding resources are communicated. Recruitment to specific patches is thus not possible in bumble bees [Citation8,Citation9].
Why are there these differences? Do different communication systems work equally well or is each system adaptive only under particular ecological conditions? Previous empirical research has primarily investigated the mechanistic details of such communication systems (e.g. [Citation9,Citation10]), but little is known about the benefits and costs of using one strategy over another and how these depend on ecological conditions or internal constraints [Citation11–13]. Here, I use an individual-based modelling approach to investigate the benefits of particular collective strategies under different environmental and social conditions.
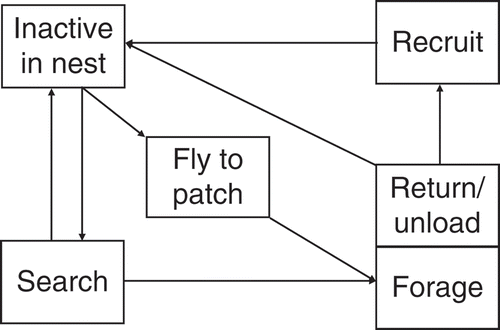
A model of honey bee foraging was used that contained six behavioural states and corresponding transition rules () and was parameterized using the extensive literature on bee foraging behaviour [Citation14]. Bees start in the hive in an ‘inactive’ state and leave with a certain probability per time step to search for food sources. Once a bee has left the hive to search, it performs a random walk until a resource patch is encountered; there, nectar is collected according to the quality of the resource, and the bee then returns to the hive in a straight path. At the hive, bees add the collected nectar to the hive's honey stores. Depending on the quality of the resource in comparison with other resources that foragers currently collect from, the bee may then recruit another bee by transmitting information on the location of the found resource patch. Only ‘inactive’ bees can be recruited in this way. Any recruited bee, as well as any bee that has just returned from a food patch, will then again leave the hive with a certain probability per time step to fly to the known resource in a straight line. With some probability, bees ‘forget’ the resource location information and return to the pool of potential searchers or recruits. These behavioural rules and the necessary parameterizations are derived from empirical results [Citation14]. In the patchiness simulations, 1000 bee foragers were simulated, foraging on between 4096 and 64 resource patches in a simulated coordinate space of 8 km × 8 km, with the bee colony located in the middle. Each bee could collect between 1.25 and 80 μl nectar per trip depending on the ‘resource quality’ of the patch visited. Colony foraging success was measured as the net energy collected, that is, the nectar collected by all bees in foraging trips minus the energy used by all bees over a simulated time of 50 h. Ten simulation runs were performed for every parameter combination tested. For more extensive sensitivity analysis of this model, see [Citation14]. The model was implemented using a graphical user interface in SeSam (http://www.simsesam.de).
Figure 1. Schematic representation of the behavioural model used in the individual-based simulations. Arrows correspond to conditional/probabilistic transitions between behavioural states. For example, ‘Forage’ immediately leads to subsequent ‘Return’, but to initiate ‘Search’, bees have a certain probability to leave the inactive state in each time step. This leads to varying durations in each state. The transition from ‘Search’ to ‘Forage’ depends on how quickly the bee actually finds a food source in the spatially explicit simulation.
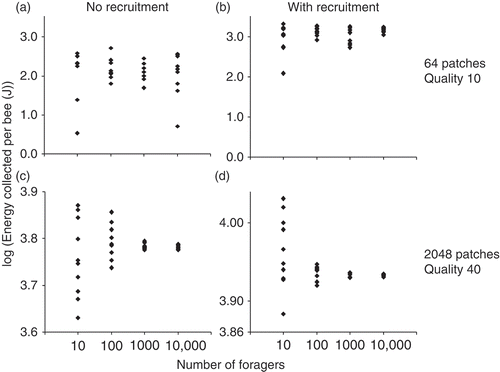
Using these methods, I quantified the benefits of a honey bee-style recruitment system in different situations. In the test for an effect of colony size, I simulated colony sizes of 10–10,000 forager bees. There was higher variance in foraging success (energy collected by the group) among simulation runs for smaller colonies. However, median foraging success was unaffected by colony size, and thus median benefits of recruitment (foraging success with/foraging success without communication) were also unaffected by colony size: Kruskal–Wallis tests show no significant effect of colony size for the 4 environments: 64/10 (64 patches of quality 10), 64/40, 2048/10 and 2048/40 without recruitment (N = 40, df = 3 for each, p = 0.63, 0.88, 0.10, 0.77, respectively) and with recruitment (N = 40, df = 3 for each, p = 0.32, 0.27, 0.28, 0.12, respectively) ().
Figure 2. Total energy collected by the simulated bees as it depends on group size. Each data point is a simulation run. The ‘quality’ of patches corresponds to the volume of nectar available per trip (in μl). No effect of group size on energy collected per bee was found. (a) and (b) show foraging success for a poor habitat; and (c) and (d) show foraging success for a rich habitat. (a) and (c) are simulations without recruitment; (b) and (d) are simulations with recruitment.
To test for an effect of resource distribution, I simulated environments with many small resources and ones with few large ones. Bees foraged more successfully in the latter ((a)). This indicates that with our choice of parameters, searching for resource patches played a smaller role than effectively collecting nectar once patches are found: with fewer resources search is harder; when resources deliver little reward per trip, collection is less efficient. However, the benefit of recruitment (ratio of foraging success with and without recruitment) showed no increasing or decreasing trend with increasing patchiness (in simulations with recruitment, bees always collected on average about 2–3 times as much energy as without recruitment: regression p = 0.08, R 2 = 0.03) ((b)).
Figure 3. (a) In our model, bees had a higher net energy gain in patchy environments (ones with few, rich resources) compared with ones with many small resources. Boxes show median and interquartile interval, whiskers show range; grey: with recruitment; white: without recruitment. (b) In the same environments, the benefits of recruitment, however, were always the same (no relationship with patchiness). Each data point is the ratio of energy collected in a simulation run with recruitment over that collected in a simulation run without recruitment.
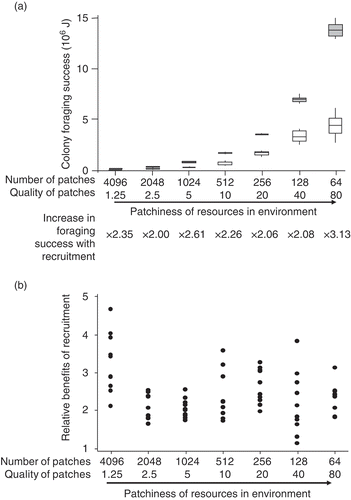
Thus, I found that neither colony size nor patchiness had a significant effect on benefits of recruitment, meaning that differences in colony size or resource patchiness cannot explain why honey bees evolved a recruitment system in foraging and bumble bees did not. However, I show that communication is particularly useful in resource-poor habitats: benefits of recruitment are highest with few, low-quality patches ((a); for full analysis see [Citation14]). Both number and quality of patches by themselves affect benefits of recruitment, but if patchiness is increased as in , the effects of patch density and quality counteract each other. At low densities and low patch qualities, foraging success is sometimes negative (bees use more energy than they are able to collect); in these cases, no relative benefit was calculated (in (a)). However, I could calculate what proportion of the energy collected was dependent on recruitment being present [(E recr–E norec)/E recr] ((b); ANOVA df = 41, R 2 = 0.92, p < 0.001 for both number and quality of resources; ANOVA was used because the data distributions were not significantly different from normal distributions). It seems that benefits of recruitment are low if resources are of high quality, even if they are at low density (and thus hard to find). This is likely the case because with rich resources, the cost of search time becomes negligible in relation to the gain from discovering a patch, whereas with poor patches it is not (and thus search time affects overall foraging success much more strongly).
Figure 4. (a) Relative benefits of recruitment (foraging success with/foraging success without communication) decrease with number of patches in the environment and with quality of patches (amount of nectar available per trip). Benefits of recruitment are thus highest with few, low-quality patches (for full analysis see [Citation14]). (b) The proportion of the energy collected with recruitment that was dependent on recruitment being present [(E recr–E norec)/E recr] also decreases with increasing number or quality of resources.
![Figure 4. (a) Relative benefits of recruitment (foraging success with/foraging success without communication) decrease with number of patches in the environment and with quality of patches (amount of nectar available per trip). Benefits of recruitment are thus highest with few, low-quality patches (for full analysis see [Citation14]). (b) The proportion of the energy collected with recruitment that was dependent on recruitment being present [(E recr–E norec)/E recr] also decreases with increasing number or quality of resources.](/cms/asset/3c2e14fc-7b1e-4741-8497-e9b5357f90bc/nmcm_a_601422_o_f0004g.gif)
There are of course many previous modelling studies that have investigated optimal foraging strategies, and many have studied recruitment behaviours in social insects (e.g. [Citation15–18]). However, only a few studies have looked at the benefits of differing recruitment strategies, and thus implicitly or explicitly at the evolution of such behaviours [Citation19–23]. These studies have typically also investigated the role of resource characteristics and colony size for social insect recruitment. For example, one previous study found a strong role for colony size in the effectiveness of forager communication [Citation24]. However, this study assumed that group size directly relates to the number of individuals recruited per forager who is recruiting: an entirely reasonable assumption to make in ants, where a pheromonal foraging trail may attract twice as many recruits in a colony that is twice the size, because there is essentially no limit to how many foragers can walk on one trail. However, this is not the case in the honey bee waggle dance: only about four bees can follow a dancer at any one time, regardless of how many bees are available to be recruited. Since a recruit has to follow on average several dances before she arrives at the indicated food source, a single forager can essentially, on average, only recruit one other individual per trip, independent of colony size [Citation25,Citation26]. It is likely that this limitation, reflected in my model, is the reason that larger colonies do not show increased advantages of recruitment.
4. Optimal levels of variation in a group: What are the benefits of division of labour?
Similar to the problem of optimally allocating foragers among resources, it is interesting to look at how insect colonies allocate workers to different tasks needed in the nest, such as brood care, defence, nest repair and cleaning. Such allocation can be achieved in principle in two ways: by inherently identical individuals reacting to a heterogeneous environment or by heterogeneous individuals who differ in their likelihood of tackling different tasks. The former strategy allows greater flexibility in re-allocating workers to different tasks if the demand for work in these tasks changes [Citation27–29]; the latter allows permanent specialization with possible benefits in that individuals may be built or adapted to particular tasks, with resulting increases in individual work efficiency ([Citation30,Citation31], although see [Citation32,Citation33]). In many social insect species, the allocation of workers to tasks is affected by worker age, genetic background or body size, that is, division of labour results from heterogeneous individuals [Citation1]. All these traits are fixed in any particular worker, and thus independent of the demand for particular tasks or the condition of the colony. In other species, task allocation is the result of worker interactions or individual experience [Citation34,Citation35]. Some species seem to employ individuals who frequently switch tasks [Citation36,Citation37]. However, the apparent diversity of task allocation mechanisms among social insects remains unexplained. What is the benefit of one or the other mechanism, and under what conditions is it adaptive to start with a heterogeneous group, rather than identical workers that react only to local cues? So far there is a lack of empirical studies on this [Citation32]; here I demonstrate that individual-based models can answer some of these questions.
Previous modelling studies which have assumed that workers differ in their propensity to engage in different tasks have often modelled this as a difference in ‘response thresholds’ [Citation38]. These are thought of as task-specific thresholds for the perception of task-relevant stimuli. Empirical evidence for this has been found in bumble bees: some bees may be more sensitive to heat and therefore start cooling (fanning) the nest at a lower temperature than other bees. If they are successful in this, bees with high-temperature thresholds would never engage in this task (fanning) [Citation39]. These thresholds are also sometimes modelled as different slopes in the increasing probability of task performance with stimulus intensity – in other words, ‘threshold’ is not always meant to imply a strict step function of response. Threshold-based mechanisms can indeed lead to individual specialization, and thus division of labour [Citation40].
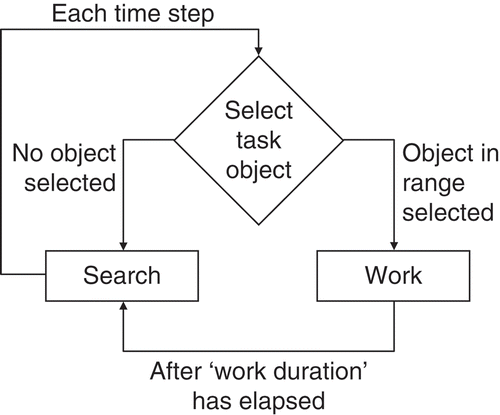
Here I describe a model in which I used a simplified representation of a social insect nest, containing workers and nest positions that needed work (). I used a simple model of workers looking for work in a two-dimensional nest. Each position in the nest had two different variables for ‘work demand’ associated with it, reflecting two different types of tasks. For example, one can imagine that each position in the nest needs to be regularly cleaned, repaired, the brood residing there cared for and so on. Here I only simulate two task types for simplicity. Workers in the nest move randomly while searching for work and will check every time step whether there is a need for work within their perception range. Depending on the task allocation mechanism used, they will then attend to this work, thus bringing local demand for work in this task down. After this, workers resume searching. The model was implemented in ObjC using the SWARM library (http://www.swarm.org).
Figure 5. Schematic representation of the task allocation model. Different task allocation rules are compared by changing the ‘Select task object’ routine. The simulation is spatially explicit with demand for each task continuously increasing at every grid cell and workers moving around on the grid randomly to search for work.
I compared how much of the needed work was performed depending on which behavioural rules, that is, what task allocation strategy the workers were using. I compared the following rules: global task allocation (a searching worker moves to the task that has the highest demand globally), random task choice (a worker will work at each time step in a randomly selected local task), local task choice based on demand (a worker will work in the locally perceptible task that has the highest demand associated with it) and various threshold-based rules (each worker has a task-specific threshold, such that it only considers working in tasks whose demand exceeds this threshold). In the threshold-based strategies, workers have inherent, fixed differences in their likelihood of working in any particular task; in all other strategies, all workers are identical. The three threshold-based strategies differ in that the first implies workers choose a task randomly as long as its demand exceeds their task-specific threshold; the second implies that workers can only test one position for its work demand at a time and then choose to work or to continue searching depending on their threshold; and the third implies that a worker will work on the task that has the highest demand divided by that worker's task-specific threshold. Thus the last rule effectively lets workers choose the task that has the highest demand, but modulated by their task threshold.
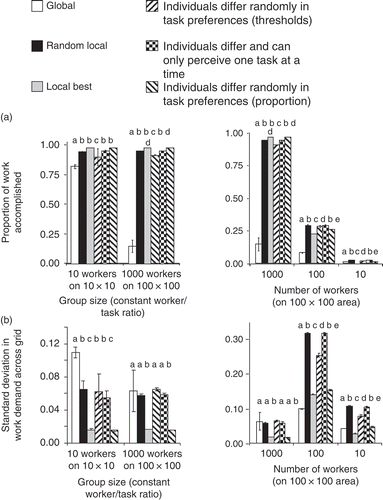
I measured group-level task performance for these different task allocation strategies, and these strategies differed in their success. (a) shows the proportion of work accomplished [between 0 and 1, defined as (1 – average remaining demand level across all tasks and grid positions) at the end of the simulation] as a measure of success for the different strategies. The task allocation strategy used has a significant effect on group performance no matter what group size and what ratio of workers to work are simulated (ANOVA, all p < 0.05). It seems that a task allocation strategy in which workers are identical and select work based on highest demand (Local best) outperforms all other strategies under many circumstances.
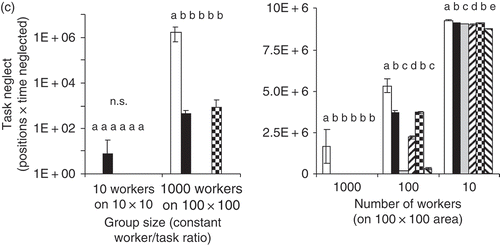
Figure 6. Three different measures of group-level performance all show that strategies differ significantly in the level of task completion achieved. In addition, not only group size but also the density of workers in a given work area affect which strategy performs optimally. (a) ‘Task demand’, the simulation parameter that shows whether workers have attended to a particular task type in a particular position, is averaged over all positions and task types at the end of the simulation. ‘Proportion of work accomplished’, shown here, is defined as (1 – average remaining demand level) (shown are averages of 10 simulation runs with standard deviations). Different letters above bars indicate significant differences in Tukey's post hoc test (for each condition separately). (b) Another measure is the evenness with which all positions and task types are attended to. Here, the amount of variation produced by the different task allocation strategies is shown as the standard deviation in task demand level at the end of the simulation. (c) The third measure shown here reflects how many positions received extremely low levels of work across the simulation. n.s. denotes no significant difference among strategies.
Strategies also differed in how accurately workers chose the tasks that needed most work. (b) shows the amount of variation, that is, spatial heterogeneity, produced by the different task allocation strategies. A high amount of variation implies that many grid positions did not receive ‘work’, such that the tasks accumulated a locally very high demand, whereas other positions were worked on much more, creating locally low demand for work.
A third measure of how well a task allocation strategy performs is how much work was neglected over the course of the simulation ((c)). High values indicate bad performance; in a social insect nest, such locally extreme neglect may lead to starvation of larvae, overheating of nest or failure to repair the nest (leading to flooding or invasion of predators). Locally extreme neglect of any task could thus have nonlinear, irreversible consequences. With variation among workers in task preferences, fewer positions in the nest were neglected compared with random task choice. It is somewhat surprising that the global task allocation had so low performance; this may be caused by its tendency to send several workers to the same position, creating competition.
Overall, I found that the worker/work ratio, that is, whether there is always more work available than workers can complete or whether there are more workers than needed, strongly affects which task allocation mechanism is optimal (). With more workers (but constant work, i.e. less work per worker), choosing based on local highest demand does best. With fewer workers (and more work per worker), completely random selection becomes better (even the best strategy). This sounds plausible, as when there is much more work than can ever be accomplished, just working anyplace randomly is as good as anything and saves travel time. However, although random selection can produce the best result in terms of work accomplished, it also causes much variation and neglect of individual positions, whereas task selection based on locally highest demand (with or without variation among workers) keeps neglect to a minimum. With more work per worker, the threshold rules, that is, those that imply variation among workers in task preferences, generally do better compared with other strategies. Thus specialization (=differences between individuals through response thresholds) may be beneficial to prevent crowding at high-demand tasks or to spread individuals across the two task types. Worker crowding and worker/work ratio are factors that have not received much attention in empirical studies; it would be interesting to see what the worker/work ratio is in social insect colonies. Evolutionary considerations indicate that more work completed should always increase fitness, so in a sense one may argue that there is always more work available than workers can complete. However, it is likely that the benefits of more nest defence, more brood care and so on level off eventually in real social insect colonies, because reproductive rates, nest size and even food collection may be constrained by factors outside worker control. Thus, whether real colonies operate near this work saturation point would be an interesting question, which may affect the optimal behavioural strategy in division of labour for different species.
5. Conclusion
In summary, individual-based models are useful for testing hypotheses about the benefits of different collective strategies under varying ecological conditions [Citation14,Citation19,Citation23,Citation41–43]. Considering spatial heterogeneity and heterogeneity in individual behaviour, information available to individuals can be crucial to understanding how behavioural rules at the individual level can lead to complex adaptive group behaviour. They can also be used to understand the evolution of particular collective strategies, if the performance of different strategies under different environmental conditions is quantified, as demonstrated here with communication and division of labour.
Acknowledgement
I thank the NSF (grant nos. IOS-0921280, IOS-0841756 and IOS-1045239) for funding.
References
- Hölldobler , B. and Wilson , E.O. 2008 . The Superorganism: The Beauty, Elegance, and Strangeness of Insect Societies , New York : W.W. Norton .
- Dechaume-Moncharmont , F.-X. , Dornhaus , A. , Houston , A.I. , McNamara , J.M. , Collins , E.J. and Franks , N.R. 2005 . The hidden cost of information in collective foraging . Proc. R. Soc. , B272 : 1689 – 1695 .
- Raine , N.E. , Ings , T.C. , Dornhaus , A. , Saleh , N. and Chittka , L. 2006 . Adaptation, genetic drift, pleiotropy, and history in the evolution of bee foraging behavior . Adv. Study Behav. , 36 : 305 – 354 .
- Grimm , V. , Revilla , E. , Berger , U. , Jeltsch , F. , Mooij , W.M. , Railsback , S.F. , Thulke , H.H. , Weiner , J. , Wiegand , T. and DeAngelis , D.L. 2005 . Pattern-oriented modeling of agent-based complex systems: Lessons from ecology . Science , 310 : 987 – 991 .
- Judson , J. 1994 . The rise of individual-based model on ecology . TREE , 9 : 9 – 14 .
- Grimm , V. and Railsback , S.F. 2007 . Individual-Based Modeling and Ecology , Princeton , NJ : Princeton University Press .
- Grimm , V. , Berger , U. , Bastiansen , F. , Eliassen , S. , Ginot , V. , Giske , J. , Goss-Custard , J. , Grand , T. , Heinz , S.K. , Huse , G. , Huth , A. , Jepsen , J.U. , Jorgensen , C. , Mooij , W.M. , Muller , B. , Pe'er , G. , Piou , C. , Railsback , S.F. , Robbins , A.M. , Robbins , M.M. , Rossmanith , E. , Ruger , N. , Strand , E. , Souissi , S. , Stillman , R.A. , Vabo , R. , Visser , U. and DeAngelis , D.L. 2006 . A standard protocol for describing individual-based and agent-based models . Ecol. Modell. , 198 : 115 – 126 .
- Dornhaus , A. , Klugl , F. , Oechslein , C. , Puppe , F. and Chittka , L. 2006 . Benefits of recruitment in honey bees: Effects of ecology and colony size in an individual-based model . Behav. Ecol. , 17 : 336 – 344 .
- Dornhaus , A. and Chittka , L. 2004 . Information flow and regulation of foraging activity in bumble bees . Apidologie , 35 : 183 – 192 .
- Granero , A.M. , Sanz , J.M.G. , Gonzalez , F.J.E. , Vidal , J.L.M. , Dornhaus , A. , Ghani , J. , Serrano , A.R. and Chittka , L. 2005 . Chemical compounds of the foraging recruitment pheromone in bumblebees . Naturwissenschaften , 92 : 371 – 374 .
- Sherman , G. and Visscher , P.K. 2002 . Honeybee colonies achieve fitness through dancing . Nature , 419 : 920 – 922 .
- Dornhaus , A. 2002 . When do dances make a difference? Significance of honey bee recruitment depending on foraging distance . Entomol. Gen. , 26 : 93 – 100 .
- Dornhaus , A. and Chittka , L. 2004 . Why do honey bees dance? . Behav. Ecol. Sociobiol. , 55 : 395 – 401 .
- Dornhaus , A. , Klügl , F. , Oechslein , C. , Puppe , F. and Chittka , L. 2006 . Benefits of recruitment in honey bees: Effects of ecology and colony size in an individual-based model . Behav. Ecol. , 17 : 336 – 344 .
- Schmickl , T. and Crailsheim , K. 2004 . Costs of environmental fluctuations and benefits of dynamic decentralized foraging decisions in honey bees . Adapt. Behav. , 12 : 263 – 277 .
- Sumpter , D.J.T. and Pratt , S.C. 2003 . A modelling framework for understanding social insect foraging . Behav. Ecol. Sociobiol. , 53 : 131 – 144 .
- De Vries , H. and Biesmeijer , J.C. 2002 . Self-organization in collective honeybee foraging: Emergence of symmetry breaking, cross inhibition and equal harvest-rate distribution . Behav. Ecol. Sociobiol. , 51 : 557 – 569 .
- De Vries , H. and Biesmeijer , J.C. 1998 . Modelling collective foraging by means of individual behaviour rules in honey-bees . Behav. Ecol. Sociobiol. , 44 : 109 – 124 .
- Cox , M.D. and Myerscough , M.R. 2003 . A flexible model of foraging by a honey bee colony: The effects of individual behaviour on foraging success . J. Theor. Biol. , 223 : 179 – 197 .
- Haefner , J.W. and Crist , T.O. 1994 . Spatial model of movement and foraging in harvester ants(Pogonomyrmex) (I): The roles of memory and communication . J. Theor. Biol. , 166 : 299 – 313 .
- Crist , T.O. and Haefner , J.W. 1994 . Spatial model of movement and foraging in harvester ants(Pogonomyrmex) (II): The roles of environment and seed dispersion . J. Theor. Biol. , 166 : 315 – 323 .
- Anderson , C. 2001 . The adaptive value of inactive foragers and the scout-recruit system in honey bee(Apis mellifera) colonies . Behav. Ecol. , 12 : 111 – 119 .
- Beekman , M. and Lew , J.B. 2007 . Foraging in honeybees – When does it pay to dance? . Behav. Ecol. , 19 : 255 – 261 .
- Beekman , M. , Sumpter , D. and Ratnieks , F. 2001 . Phase transition between disordered and ordered foraging in Pharaoh's ants . Proc. Natl. Acad. Sci. U.S.A. , 98 : 9703 – 9704 .
- Esch , H. and Bastian , J.A. 1970 . How do newly recruited honey bees approach a food site? . Z. Vergl. Physiol. , 68 : 175 – 181 .
- Seeley , T.D. and Visscher , P.K. 1988 . Assessing the benefits of cooperation in honeybee foraging: Search costs, forage quality, and competitive ability . Behav. Ecol. Sociobiol. , 22 : 229 – 237 .
- Franks , N.R. and Tofts , C. 1994 . Foraging for work: How tasks allocate workers . Anim. Behav. , 48 : 470 – 472 .
- Pereira , H.M. and Gordon , D. 2001 . A trade-off in task allocation between sensitivity to the environment and response time . J. Theor. Biol. , 208 : 165 – 184 .
- Dornhaus , A. , Holley , J.-A. , Pook , V.G. , Worswick , G. and Franks , N.R. 2008 . Why don't all workers work? Colony size and workload during emigrations in the antTemnothorax albipennis . Behav. Ecol. Sociobiol. , 63 : 43 – 51 .
- Smith , A. 1776 . An Inquiry into the Nature and Causes of the Wealth of Nations Edited by: Strahan , W. and Cadell , T. London
- Wilson , E.O. 1980 . Caste and division of labor in leaf-cutter ants (Hymenoptera: Formicidae:Atta) II. The ergonomic optimization of leaf cutting . Behav. Ecol. Sociobiol. , 7 : 157 – 165 .
- Dornhaus , A. 2008 . Specialization does not predict individual performance in an ant . PLoS Biol. , 6 : 285
- Muscedere , M.L. , Willey , T.A. and Traniello , J.F.A. 2009 . Age and task efficiency in the antPheidole dentata: Young minor workers are not specialist nurses . Anim. Behav. , 77 : 911 – 918 .
- Gordon , D.M. 1996 . The organization of work in social insect colonies . Nature , 380 : 121 – 124 .
- Beshers , S. and Fewell , J.H. 2001 . Models of division of labor in social insects . Annu. Rev. Entomol. , 46 : 413 – 440 .
- Seid , M.A. and Traniello , J.F.A. 2006 . Age-related repertoire expansion and division of labor inPheidole dentata (Hymenoptera : Formicidae): A new perspective on temporal polyethism and behavioral plasticity in ants . Behav. Ecol. Sociobiol. , 60 : 631 – 644 .
- Jandt , J. , Huang , E. and Dornhaus , A. 2009 . Weak specialization of workers inside a bumble bee nest . Behav. Ecol. Sociobiol. , 63 : 1829 – 1836 .
- Beshers , S.N. and Fewell , J.H. 2001 . Models of division of labor in social insects . Annu. Rev. Entomol. , 46 : 413 – 440 .
- Weidenmüller , A. 2004 . The control of nest climate in bumblebee(Bombus terrestris) colonies: Interindividual variability and self reinforcement in fanning response . Behav. Ecol. , 15 : 120 – 128 .
- Bonabeau , E. , Theraulaz , G. and Deneubourg , J.-L. 1998 . Fixed response thresholds and the regulation of division of labor in insect societies . Bull. Math. Biol. , 60 : 753 – 807 .
- Marshall , J.A.R. , Kovacs , T. , Dornhaus , A. and Franks , N.R. 2003 . Simulating the evolution of ant behaviour in evaluating new nest sites . Lect. Notes Artif. Intell. , 2801 : 643 – 650 .
- Planque , R. , Dornhaus , A. , Franks , N.R. , Kovacs , T. and Marshall , J.A.R. 2007 . Weighting waiting in collective decision-making . Behav. Ecol. Sociobiol , 61 : 347 – 356 .
- Marshall , J.A.R. , Dornhaus , A. , Franks , N.R. and Kovacs , T. 2006 . Noise, cost, and speed-accuracy trade-offs: Decision making in decentralized systems . J. R. Soc. Interface , 3 : 243 – 254 .