Abstract
The construction of nests in insect societies requires building materials, pulp and water foragers, builders and also an organized workforce for effective construction. The central hypothesis of this study is that wasp societies developed a social crop, or common stomach, which stores water and provides a mechanism for worker connectivity, which in turn regulates construction behaviour. Inspired by the construction behaviour of social wasps, an agent-based model is presented to show that via the usage of the common stomach, larger colonies enjoy the benefit of having highly effective foragers, while most of the swarm stays on the nest and only a few engage in highly risky foraging trips. We also demonstrate how colony efficiency changes as a function of colony size and the constitution of the labour distribution, as well as how indirect interactions can increase efficiency of labour in wasp societies.
1. Introduction
In social insects, colony-level complexity emerges from simple individual-level behaviours and interactions. Insect societies can be conceived as superorganisms [Citation1] in which inter-individual conflict for reproductive privilege is largely reduced and the worker caste is selected to maximize colony efficiency [Citation2]. Emergent global properties such as colony size and the degree of division of labour, which can be viewed as a consequence of life history traits, may influence individual-level behaviours [Citation3]. Division of labour is one of the most interesting and studied features of colony-level functioning in insect societies. These studies are commonly concerned with the integration of individual worker behaviour into colony-level task organization and with the question of how regulation of division of labour may contribute to colony efficiency [Citation4,Citation5].
Colonies have to perform many tasks including foraging for resources, brood care, nest building and defence. However, because colonies and their environments are dynamic in nature, the labour requirements of the colony may change over time and the division of labour must accommodate to new demands. To meet new labour demands, efficient re-allocation of individuals to different tasks is required via continuous dynamic adjustments in response to these changes. These re-allocations may be manifested through recruiting and abandoning given tasks and/or changing the rate of work at the individual level. This colony-level flexibility in response to external and internal changes is an essential feature of division of labour [Citation6,Citation7]. Parallel processing by specialists in large colonies provides flexible and efficient colony-level functioning, while the individual behavioural flexibility of jack-of-all trades workers ensures success of small and early societies [Citation3].
Theoretical and empirical findings on a diverse array of social insect taxa show that interactions among workers (called worker connectivity) often play important roles in structuring division of labour [Citation8]. O'Donnell and Bulova [Citation9] propose several advantages of relying on shared and connected information rather than on the individual agent's own assessment. (1) Connectivity can allow the sharing of information among more workers, and across greater distances, than direct perception of task stimuli. (2) Connectivity may function to push workers into a different task or to overcome task inertia. (3) There is a possibility that better informed individuals can propagate the information through the connected colony faster. The possible mechanisms of worker connectivity range from simple encounters with nestmates [Citation10,Citation11] to specialized communicative displays [Citation12].
Besides the well-known connection networks, such as pheromones and dances, honeybee and ant workers are also connected through a wide variety of direct and indirect communication networks to regulate or fine-tune their division of labour [Citation13]. Cassill and Tschinkel [Citation14] found that the division of labour in Solenopsis invicta ants depends on worker age and size and is fine-tuned by ever-changing states of their crop volume and content. Food reserves maintained by honeybee colonies not only ensure homeostasis, but also regulate division of labour [Citation15]. In social wasps we found that construction behaviour is regulated by a special type of indirect worker connectivity. The colony forms a common stomach (CS) or social crop to store water temporally, and the pace of work and division of labour are regulated via the amount of water in the CS [Citation16]. We argued that the regulation of division of labour is based on a self-organized adaptive mechanism, which relies on simple interactions between the individuals through the CS and the use of simple rules of thumb for decisions and actions [Citation17].
Models on decision-making in social animal collectives are recently reviewed in [Citation18] and the role of nutritional interaction among individual agents is reviewed in detail in [Citation19]. Beshers and Fewell [Citation20] reviewed different approaches used to model division of labour in social insects. The Metapolybia wasp societies that we are using as a model system neither display morphologically distinct workers nor have a high degree of age polyethism. Therefore, while our model shares some similarities with other studies, especially those centred around network models, it has several unique features for modelling task partitioning. In our model we assume no intrinsic differences among the workers, and we focus on worker connectivity using simple cues. Water is not only a building material but also a regulator. The water is stored temporally in the CS which is used as an information centre allowing regulation of behaviour and collective information processing and also provides a short-term buffer against fluctuation in material inflow.
In this article our goal is not to build a model of the exact behaviour of wasp societies, but rather to investigate in a more abstract way some of the features of the CS. While our model is inspired by the colony regulation of social wasps, the presented model is intentionally minimalistic. Our agents are much less sophisticated than the wasps, and the interactions among the individuals are simplified. In this present model we focus on the function of the CS rather than on the dynamically evolving agents or seeking optimal colony allocation schemas. Specifically, we will investigate why the CS provides benefits contrary to the extra interactions involved and how this depends on colony size, resource availability and the constitution of the task force.
2. The model
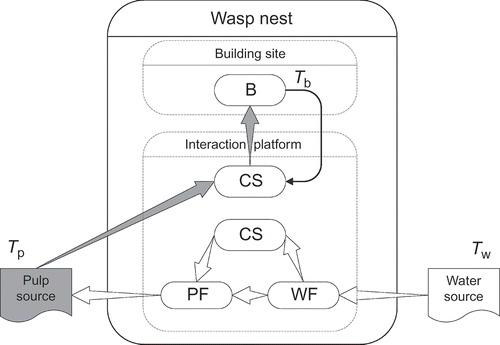
The multi-agent simulation is written in Java to model the construction behaviour of a swarm. Nest building requires pulp and builders; for the pulp collection the colony needs water foragers (WF) and pulp foragers (PF), and for the water the colony needs WF. For simplicity, we assumed that each agent belongs to a given task group and this will not change during the simulation:
WF: These wasps are specialists; they deliver water to the nest from an outside source. | |||||
PF: These wasps are specialists; they use the water they receive in the nest and fly out to an external source to collect pulp while using up the water in the process. | |||||
CS wasps: These are generalist wasps; they do not leave a nest and can accept water from the WF or give water to PF. They also accept pulp and they build this pulp into the nest, acting as builders. | |||||
Collection of pulp and water happens outside the nest at the pulp and water sources, respectively (). The time required for collecting these materials is parameterized with T w (water) and T p (pulp) collecting times, while the given wasp is outside the nest collecting materials (). One time unit corresponds to 10 seconds of real time as measured in wasp colonies [Citation16]. To simplify the construction of the nest in our model we assumed that there is no variation in collection times or the amount of water and pulp collected. The PF gives away all her pulp the next turn after her arrival if there is at least one CS wasp in its Moore neighbourhood. If there is no receptive wasp, the PF offers her pulp load until it is taken in the consecutive turns, and then in the following turn she starts to collect water from the CS wasps for the next pulp collection trip. The CS wasp that received the pulp load shares this pulp with B – 1 other CS wasps (this way the pulp is shared among B wasps) currently unoccupied on the active platform. These wasps then leave the interaction platform for T b (building) time to build the pulp into the nest. If there are not enough free wasps, the excess pulp is discarded.
Table 1. Parameters of the model
Figure 1. Schematic representation of the wasp nest. Wasp types: WF, water forager; PF, pulp forager; CS, common stomach wasp; B, builder. The flow of the water is shown by white arrows. Pulp is transported from pulp source to the nest (grey arrow) and it is given to builders. Builders build the pulp into the nest on the building site. Solid arrow shows the transition of behaviour of builders who finished their job. T p, T w and T b: time needed for pulp, water collection and building, respectively.
The interaction platform of the wasp nest, where water exchange occurs, was represented by a 10 × 10 cell grid, which is able to accommodate the whole wasp colony. To make interaction simple and independent of walking patterns, in each time step, we reshuffle all wasps that are supposed to be on the active platform and place them randomly on the grid. After the wasps are positioned, each wasp attempts to interact with a single wasp in its Moore neighbourhood (). The agent in focus examines how many potential cooperative agents are in the neighbouring cells and randomly chooses one to interact with. We allowed both direct and indirect interactions between water providers and consumers. Direct interaction entails water transfer between a WF and a PF, while indirect interaction involves CS wasps that store the water temporally. The rules of interaction are described as simple material transfer: if the states of the interaction are matching (one giver and one receiver) then material transfer can happen. No interaction happens if their states are not matching (e.g. two foragers of the same type interact). If no interaction is possible, then the agent retains its behavioural state and makes a random landing again on the interaction platform in the next turn. This simplified routine is close to what we can observe in real wasp colonies during a 10-second time interval: the wasp either makes an interaction with a neighbour and material transfer happens or she moves around on the interaction platform [Citation16].
Figure 2. Interactions of workers on the interaction platform. Workers: WF wasp with dark grey colour and drop shape: WF; PF wasp with medium grey colour and hexagonal shape: PF; CS wasp with grey body and white abdomen: CS wasp. CS wasps listed as builders are on the building site constructing the nest. The bar next to the wasp indicates the fullness of stomach with water. The large bar next to the active platform shows the relative fullness of the CS.

The simplifications above allowed us to concentrate on the water exchange among individuals which is the main focus of this study. Based on our field study [Citation16], we assumed that the water is exchanged in units of volume. A forager needs to visit minimum of W (6 × V) wasps to fully unload its water or fully fill up her stomach. This partial water exchange and the need for several interactions to achieve the job might be the consequence of an optimal information acquisition system which was described in other insect societies [Citation21]. We studied the following three different water exchange strategies:
| 1. | No-resistance strategy: In this case when a giver and a receiver meet, they always exchange V quantity of water. | ||||
| 2. | Linear resistance strategy: The water exchange is probabilistic and it is the linear function of the CS. An empty CS wasp always accepts water from a WF, but if the CS wasp has 5 units of water in their stomach, only 20% of interactions result in water exchange. | ||||
| 3. | Sigmoid resistance strategy: This is similar to the linear strategy, but the shape of the function is sigmoid. The probability of water exchange is | ||||
where x is the amount of water in the CS. In this strategy, for example, water unloading to CS wasps happens with higher probability when the CS is less than half full and with low probability when the CS is above 50%.
The number of delivered pulp, used pulp (the pulp that was built into the nest), used pulp/foragers (WF + PF) and wasted pulp (all in pulp unit: 1 pulp load has 6 pulp units) were used as a measure of efficiency in a given colony. Each simulation started with empty CS wasps and all foragers landed on the interaction platform with a full load. To avoid the effect of this colony initiation biasing the results, the first 100 time steps (about 20 complete foraging cycles) were discarded and only the pulp arrival of the next 150 time steps was measured. Twenty parallel runs were made for each colony combination and the average values are presented. Non-parametric statistics (Mann–Whitney U-test) were used to compare different simulations using SPSS version 17 (IBM SPSS Inc., Armonk, NY, USA).
3. Results
At the first step we compared the performance of two very different colony set-ups: colonies with a low number of CS wasps but with many foragers (ratio: CS:WF:PF = 2:4:4) versus colonies where more than half of the colony members are CS wasps and have fewer foragers (ratio: CS:WF:PF = 6:2:2). In the former colonies, direct interactions between PF and WF are very common and in the latter colonies, most interaction occurs between the foragers and the CS wasps. We also studied the effect of resource availability in terms of collection time: in one setup, collecting both resources took the same amount of time (T w = T p = 4) versus when pulp foraging required more time than water foraging (T w = 2, T p = 8).
With the increasing swarm size, the quantity of delivered pulp significantly increased (Mann–Whitney U-test p < 0.05, N = 40) (). In general, swarms that had fewer CS wasps delivered significantly more pulp than those with many CS wasps, simply because these colonies had more PF. However, at the largest swarm size, this trend reversed and colonies with a smaller number of pulp foragers delivered more pulp when the cost of pulp foraging was the same as the cost of water foraging (Mann–Whitney U-test p < 0.05, N = 40) (). When pulp foraging was more time-consuming, there was no significant difference between the amount of delivered pulp of the two largest colonies (Mann–Whitney U-test p > 0.05, N = 40) ().
Figure 3. Function of building efficiency (pulp input) as the function of swarm size in case of no-resistance water exchange strategy. Black columns: 60% of the swarms are CS wasps and 20–20% are PF and WF, respectively; grey columns: 20% of the swarms are CS wasps and 40–40% are PF and WF, respectively. Panels (a) and (b): total pulp unit delivered; panels (c) and (d): pulp used for construction; panels (e) and (f): pulp used for construction/(WF + PF); panels (a), (c) and (e): T w = T p = 4; panels (b), (d) and (f): T w = 2, T p = 8.
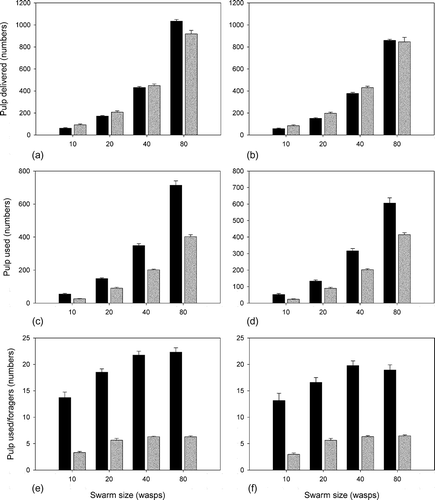
Evaluating the amount of pulp that was actually used for construction (delivered – discarded) and the amount of pulp used divided by the number of foragers showed a general tendency that was independent of the cost of pulp foraging and colony size. Colonies with more CS wasps in their workforce used more pulp and their foragers were more effective (Mann–Whitney U-test p > 0.05, N = 40) (–(f), black vs. grey columns), contrary to the fact that these colonies had far fewer foragers and the foragers are interacting primarily in indirect way through CS wasps. The amount of used pulp increased with the colony size, but the efficiency of the foraging (used pulp/foragers) levelled off. The differences in foraging efficiency decreased and finally became insignificant (between CS:WF:PF = 48:16:16 and CS:WF:PF = 16:32:32 (Mann–Whitney U-test p > 0.1, N = 40)) or even decreased (CS:WF:PF = 24:8:8 and CS:WF:PF = 48:16:16 (Mann–Whitney U-test p < 0.05, N = 40)) between the swarm of 40 and 80 wasps ( and (f)).
When pulp foraging is more costly than water foraging, the quantity of delivered and used pulp as well as the efficiency of the foragers is significantly smaller (Mann–Whitney U-test p < 0.05, N = 40); however, there are some exceptions. Increased pulp foraging cost did not result in smaller amount of used pulp or decreased forager efficiency in colonies (CS:WF:PF = 6:2:2, CS:WF:PF = 4:8:8, CS:WF:PF = 8:16:16 (Mann–Whitney U-test p > 0.1, N = 40)) and it had an increasing effect in colonies (CS:WF:PF = 16:32:32 (Mann–Whitney U-test p < 0.05, N = 40)).
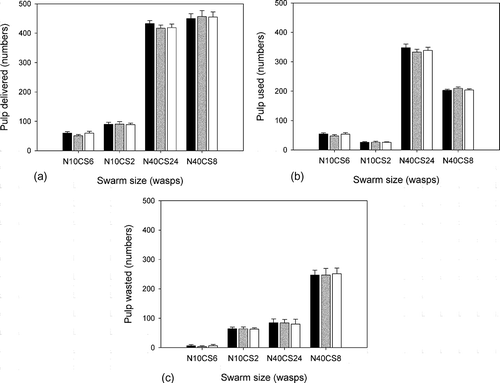
On average the different water exchange strategies had little effect on the efficiency of the construction (). This is especially true for comparing the amount of wasted pulp, which showed no significant difference when comparing colonies that used different strategies (the only exception is the marginally significant difference between CS:WF:PF = 6:2:2 colonies using linear vs. sigmoid function (Mann–Whitney U-test p = 0.043, N = 40)) (). In colonies where CS wasps were in the minority, the amounts of delivered and used pulp were the same, except that a slightly larger number of used pulp was detected in CS:WF:PF = 8:16:16 colonies that used linear strategy (Mann–Whitney U-test p < 0.05, N = 40). Due to the increased importance of indirect interaction in colonies with large CS wasp population, the effects of the types of water exchange strategies were somewhat more pronounced. In general, no resistance resulted in slightly more delivered and used pulp than linear resistance (Mann–Whitney U-test p < 0.05, N = 40), and sigmoid resistance resulted in slightly more delivered and used pulp in CS:WF:PF = 6:2:2 colonies (Mann–Whitney U-test p < 0.05, N = 40), but not in the larger CS:WF:PF = 24:8:8 colonies ().
Figure 4. Comparison of the effectiveness of colonies using different water exchange strategies. Colony composition is given in the x-axis as NxxCSxx where Nxx is the colony size and CSxx is the number of CS wasps in the colony. The number of PF and WF in these colonies is (N – CS)/2). Black columns: no resistance; grey columns: linear resistance; white columns: sigmoid resistance.
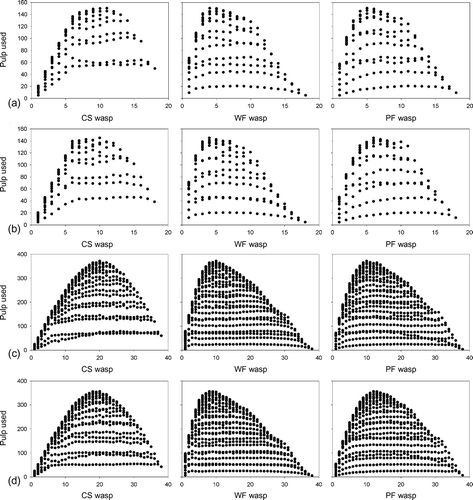
Examining all viable forager versus CS workforce combinations revealed that these combinations are very different in efficiency (). The most pulp was used in construction (most efficient colonies), where about half of the colony consisted of CS wasps and the other half comprised PF and WF close to equal numbers, but with slight PF dominance. Increasing the time cost of pulp foraging shifted this distribution towards having a greater proportion of PF. Also due to the increased cost of pulp foraging, the efficiency of these colonies is lower. These average patterns are consistent in different colony sizes and clearly show the importance of a large number of CS wasps for efficient colonies.
Figure 5. Average efficiency (dots) of different workforce combinations in small (N = 20) (a, b) and larger (N = 40) (c, d) swarms. Efficiency (pulp used for construction) is calculated from 20 parallel runs of the same type of colony mix and represented as a single dot at each panel that describes the number of wasps in a given task for that colony; (a, c): T p = T w = 4; (b, d): T w = 2, T p = 8.
4. Discussion
Our model predicted that the effective and low-risk use of worker force via worker connectivity (CS) is affected by both colony size and the time required for retrieving the resources. Those colonies that have fewer CS wasps, and therefore more PF, delivered more pulp to the colony, but this trend reversed at the highest colony sizes. However, the colonies with more CS wasps used the pulp more effectively. They wasted less pulp and their foragers were 3–4 times more effective than those from colonies where the CS wasps comprised only 20% of the swarm. Generating colonies with all possible workforce combinations showed that the highest pulp use is possible when about 50% of the swarm is comprised of CS wasps and the foragers share the other half of the colony in a balanced manner. When pulp and water collection needs the same effort, colonies with slightly biased PF workforce are the most effective. As pulp foraging becomes more costly, more PF are needed, and giving up WF for these extra PF ensured maximal effectiveness. Of course, the extra time required for collecting pulp decreases the amount of construction achieved, but the swarm seems able to adapt to these changes and still provide a very good construction performance.
Our model predicted less wasted pulp as an important benefit of having a large fraction of workers participating to form a CS. This benefit stems from the complex role of the CS wasps. Obviously CS wasps form the group from which builders are recruited; therefore if there are not enough available wasps for construction, the excess pulp will be discarded. Due to this bottleneck effect, small colonies of wasps in nature operate with a different strategy than what we used in our model in order to avoid wasting pulp. Namely the pulp they collect is not shared or distributed, but the PF becomes a builder and builds the pulp into the nest herself [Citation3]. In larger colonies, sharing pulp and bringing larger load to the nest is important to decrease the number of foraging trips.
The other benefit the CS wasps provide is to make indirect interactions possible between water providers (WF) and water users (PF). These foragers spent considerable time out of the nest while collecting, and after arriving on the active platform they should be able to exchange water quickly. However, their meeting and interaction requires that both foragers wait on the nest and search each other actively. The CS wasps staying on the nest could provide a convenient recipient of water from the WF, and therefore the WF does not need to wait or meet with the water users directly. Increased water availability is beneficial to PF as well. These CS wasps provide convenient interaction partners and also the water they store provides a buffer against fluctuations in the material flows. These factors will lead to a colony composition where most wasps stay on the nest and only a few very effective foragers need to leave the nest.
Sakagami and Fukuda [Citation22] showed that honeybee workers carrying out foraging have high mortality. Keeping the number of foragers low in a much smaller wasp society is beneficial, because it decreases the quick loss of foragers, and the low number of specialized foragers will further increase their effectiveness due to experience gained by the frequent trips [Citation23].
Our findings imply that the effective and low-risk use of worker force via worker connectivity (CS) is reliant upon colony sizes. The benefit of organizing colony-level performance through worker connectivity may function as an important evolutionary pressure for increasing colony size for insect societies. The size of the interaction platform could be a consequence of evolutionary pressures that prefer to keep most wasps on the nest. Adjusting the size of the interaction platform and the number of CS wasps ensures an ideal density for quick material exchange. Low density on the interaction platform results in many turns without useful interaction, and very high density will commonly provide no benefit or can even be detrimental to the efficiency. We also showed that different water exchange strategies did not result in large differences in the rate of construction. Obviously if there is no resistance in the water exchange, that is, if the CS wasps always accept the material exchange request, this will result in the quickest water downloads and uploads. Having a resistance will require more interaction to download or upload the full load of water from and to the foragers. This might have a small detrimental effect in some colony combination as we presented, but it can also have some benefit. Having a resistance will decrease the variation of the amount of water in the individual wasps. Visiting more wasps, the forager also could gather more accurate information on the fullness of the CS and this in turn could be important in adjusting division of labour or speed of work.
Our goal with the current model was to study the effect of swarm size and the mix of workforce on the efficiency of construction. Further studies are required to build more elaborated agent-based models for the wasp societies and understanding the role of the CS in the regulation of division of labour. In our current model, the tasks of the agents did not change. While this is a realistic assumption in short term, the wasps do change behavioural profiles or adapt to the colony needs as has been shown [Citation16,Citation24]. Our preliminary studies show that these adjustments also depend on the state of the CS [Citation3,Citation16,Citation17], and we intend to investigate this further. Self-organization is a powerful theory to explain how minimal complexity at the individual level can generate much greater complexity at the collective level. Recently, we have more and more evidence that although genetic, physiological and other aspects must be taken into account, division of labour is an emergent property of the society [Citation1,Citation20]. The CS or social crop seems to be an important information centre that plays a crucial role in the regulation of the behaviour of the swarm.
Acknowledgements
The authors are grateful to T. Schmickl and the anonymous reviewers for their valuable comments. This work was supported by the Department of Biological Sciences and the Honors College of ETSU.
References
- Holldobler , B. and Wilson , E.O. 2008 . The Superorganism: The Beauty, Elegance and Strangeness of Insect Societies , New York : W. W. Norton & Company .
- Ratnieks , F.L.W. and Helantera , H. 2009 . The evolution of extreme altruism and inequality in insect societies . Philos. Trans. R. Soc. Lond. [Biol.] , 364 : 3169 – 3179 .
- Karsai , I. and Wenzel , J.W. 1998 . Productivity, individual-level and colony-level flexibility, and organization of work as consequences of colony size . Proc. Natl. Acad. Sci. U.S.A , 95 : 8665 – 8669 .
- Oster , G.F. and Wilson , E.O. 1979 . Caste and Ecology in the Social Insects , Princeton , NJ : Princeton University Press .
- Jeanson , R. , Fewell , J.H. , Gorelick , R. and Bertram , S.M. 2007 . Emergence of increased division of labor as a function of group size . Behav. Ecol. Sociobiol , 62 : 289 – 298 .
- Calabi , P. 1988 . “ Behavioral flexibility in Hymenoptera: A re-examination of the concept of caste ” . In Advances in Myrmecology , Edited by: Trager , J.C. 237 – 258 . Leiden : Brill Press .
- Robinson , G.E. 1992 . Regulation of division of labor in insect societies . Annu. Rev. Entomol , 37 : 637 – 665 .
- O'Donnell , S. 2006 . Polybia wasp biting interactions recruit foragers following experimental worker removals . Anim. Behav , 71 : 709 – 715 .
- O'Donnell , S. and Bulova , S.J. 2007 . Worker connectivity: A review of the design of worker communication systems and their effects on task performance in insect societies . Insect. Soc , 54 : 203 – 210 .
- Gordon , D.M. , Paul , R.E.H. and Thorpe , K. 1993 . What is the function of encounter patterns in ant colonies? . Anim. Behav , 45 : 83 – 100 .
- Pratt , S.C. 2005 . Quorum sensing by encounter rates in the ant Temnothorax albipennis . Behav. Ecol , 16 : 488 – 496 .
- Sherman , G. and Visscher , P.K. 2002 . Honeybee colonies achieve fitness through dancing . Nature , 419 : 920 – 922 .
- Dornhaus , A. Finding optimal collective strategies using individual-bases simulations: Colony organization in social insects . Proceedings MATHMOD 09 Vienna Full Papers CD Volume, 6th Vienna Conference on Mathematical Modelling . Vienna . Edited by: Troch , I. and Breitenecker , F. pp. 888 – 894 . ARGESIM Publishing House .
- Cassill , D.L. and Tschinkel , W.R. 1999 . Task selection by workers of the fire ant Solenopsis invicta . Behav. Ecol. Sociobiol , 45 : 301 – 310 .
- Schmickl , T. and Crailsheim , K. 2004 . Inner nest homeostasis in a changing environment with special emphasis on honey bee brood nursing and pollen supply . Apidologie , 35 : 249 – 263 .
- Karsai , I. and Wenzel , J.W. 2000 . Organization and regulation of nest construction behavior inMetapolybia wasps . J. Insect Behav , 13 : 111 – 140 .
- Karsai , I. and Balazsi , G. 2002 . Organization of work via a natural substance: Regulation of nest construction in social wasps . J. Theor. Biol , 218 : 549 – 565 .
- Couzin , I.D. 2009 . Collective cognition in animal groups . Trends Cogn. Sci , 13 : 36 – 43 .
- Simpson , S.J. , Raubenheimer , D. , Charleston , M.A. , Clissold , F.J. , Couzin , I.D. and Clements , K.D. 2010 . Modelling nutritional interactions: From individuals to communities . Trends Ecol. Evol , 25 : 53 – 60 .
- Beshers , S.N. and Fewell , J.H. 2001 . Models of division of labor in social insects . Annu. Rev. Entomol , 46 : 413 – 440 .
- Ratnieks , F.L.W. and Anderson , C. 1999 . Task partitioning in insect societies . Insect. Soc , 46 : 95 – 108 .
- Sakagami , S.F. and Fukuda , H. 1968 . Life tables for worker honeybees . Res. Popul. Ecol , 10 : 127 – 139 .
- Jeanne , R.L. 1986 . The organization of work in Polybia occidentalis: Costs and benefits of specialisation in a social wasp . Behav. Ecol. Sociobiol , 191 : 333 – 341 .
- Jeanne , R.L. 1996 . Regulation of nest construction behaviour in Polybia occidentalis . Anim. Behav , 52 : 473 – 488 .