
ABSTRACT
Recovery methods and techniques for archaeological sampling can yield major differences in abundance and anatomo-taxonomical representation of animals, affecting past social and ecological reconstruction. Despite being a common organic material in archaeological sites, faunal remains typically exhibit differential preservation of species and skeletal elements due to pre- and post-depositional processes. This is particularly true for small-sized animals such as certain species of fish, whose often small and fragile fragments are difficult to recover and identify. Here, we present the results of a comparative analysis between two ichthyoarchaeological assemblages from Caleta Vitor 3 in northern Chile (CV3, 18°45′09″ S), an Early to Middle Holocene (9.2–7.6 ka cal BP) Chinchorro shell midden site. We compare samples obtained and processed, both in the field and the lab, using different recovery techniques. We developed a data standardisation procedure to compare and evaluate skeletal representation, taxa distribution and variations throughout the stratigraphic sequence. Our results show that mesh screen size affects not only the abundance and density of fish but also species representation. Moreover, the identification of small pelagic fish at CV3 sheds further light upon prehistoric fishing strategies and social organisation during the site’s early occupation.
Introduction
Reproducible sampling and recovery methods are still open questions in archaeology, as they depend on fieldwork limitations, methodological and theoretical backgrounds, which are often conditioned by the emphasis placed on spatial or temporal approaches (Mueller Citation1974; Schiffer, Sullivan, and Klinger Citation1978; Orton Citation2000; Peregrine Citation2002). Thus, differences in field recovery sampling and lab processing can be expected depending on the methodological and theoretical focus of the study, and its research questions. The aforementioned differences introduce biases that could affect the analysis and interpretations of archaeological evidence, particularly in the case of animal remains.
Recovery Methods in Zooarchaeology: The Case of Fish Remains
In zooarchaeology, experimental and fieldwork studies have demonstrated that field sampling recovery and lab-processing techniques may affect anatomical and taxonomic taxa representation, depending on specimen size, bone density and pre- and post-depositional processes (e.g. Payne Citation1972; Watson Citation1972; Meadow Citation1980; Shaffer Citation1992; Shaffer and Sanchez Citation1994). Aside from these biases, the cultural value given to each animal must be considered alongside socio-cultural interpretations (Russell Citation2011; Hill Citation2013). For hunter-gatherer societies, an overrepresentation of large animals has influenced the prevalence of a big-game hunting paradigm (Hildebrandt and McGuire Citation2003; Gurven and Hill Citation2009; Konner and Eaton Citation2010; Buckley and Buikstra Citation2019), although since the middle of the twentieth century other smaller animals have been considered when analysing the archaeological record (Struever Citation1968; Stahl Citation1996; Landon Citation2005). Thus, intensive analysis of fish remains only really began in the early 1970s (Casteel Citation1976b; Wheeler and Jones Citation1989; Colley Citation1990).
The morphology and preservation of specific fish bones such as vertebrae (sometimes even from particular taxa) may generate biases in the ichthyoarchaeological record as they are easily recognised even by non-specialists. Furthermore, several studies have also demonstrated how recovery methods, especially mesh screen size, can affect anatomo-taxonomical representation and abundance of fish remains from archaeological sites (Casteel Citation1972, Citation1976a; Garson Citation1980; Wheeler and Jones Citation1989; Butler Citation1993; Nagaoka Citation1994, Citation2005; Matsui Citation1996; Barrett Citation1997; Ross and Duffy Citation2000; McKechnie Citation2005; Zohar and Belmaker Citation2005; Partlow Citation2006). Problems associated with logistical fieldwork issues – such as time and resources available – can affect the methodological decisions taken when developing an excavation. Despite recent efforts by zooarchaeologists to standardise faunistic recovery methods (Reitz Citation2009; Albarella Citation2017), there are still important differences between these, especially those concerned with the recovery of ichthyoarchaeological material. In the case of the Atacama Desert, the early and permanent human occupations in the coastal area have been studied for many years by different research teams, resulting in a variety of recovery methods and analytical procedures of fish bones and otoliths.
Fishing Along the Atacama Desert Coast, Northern Chile: The Chinchorro Culture
Regarding the fisher-hunter-gatherer societies that lived in the Atacama Desert coast of northern Chile, the study of coastal resources has been mainly focused on mollusks, even though the contribution of fish species is recognised (Santoro et al. Citation2017). This is partly due to the visibility of malacological material in archaeological shell middens, and also because they are easier to identify at a taxonomic level. Fish remains are much less visible and in some cases highly fragmented, all of which can make taxonomic identification difficult. Paradoxically, fishing gear found at these sites are much more diverse and they have been largely studied in relation to the tools used to collect mollusks. Indeed, even the earliest technologies (11-9 ka cal BP) included fishhooks made of cactus spines and composite fishhooks made of bone (Standen and Santoro Citation2004; Arriaza et al. Citation2005).
In the northernmost area of the Chilean coast, between what is now Arica (18°S) and Patillo (20°S), Chinchorro communities developed a predominantly maritime economy (Standen and Santoro Citation2004; Arriaza et al. Citation2008; Santoro et al. Citation2012, Citation2020; Carter Citation2016). During the Early to Middle Holocene (9.2-4 ka cal BP), these groups used a number of fishing technologies, including a variety of fishhooks made from shell, bone, rocks and plants; along with harpoons and spear throwers (Arriaza Citation1995). Recent stable isotope studies (Roberts et al. Citation2013; Standen et al. Citation2018) provide further evidence of the important contribution of coastal resources in the diet of these populations. Finally, it is interesting to highlight the apparent absence of watercraft, which in areas slightly further south (26°S) allowed local populations access to pelagic fish (Llagostera Citation1989; Olguín, Salazar, and Jackson Citation2014; Béarez et al. Citation2016; Salazar et al. Citation2018, Citation2020; Santoro et al. Citation2020).
Further evidence for the exploitation of fish species comes from studies on vegetal and animal fibres found at the Caleta Vitor site. In this regard, Martens and Cameron (Citation2019) suggest its use, among other activities, as fishing nets. Even though the authors and other researchers (Carter Citation2016; Disspain et al. Citation2016b) have suggested the use of these fishing strategies for small (sardines and anchovetas) and middle size fish species (Sciaenidae), there is no in-depth discussion about taxonomical distribution and variation through time. Furthermore, discussion of methodologies for field and lab recovery of fish remains as well as their analytical procedures is also lacking.
Recovery Methods of Fish Remains in Caleta Vitor
Caleta Vitor (18°45′09″ S) is situated in the mouth of Quebrada Vitor, 30 km south of the city of Arica. A key wetland environment occurs as water from the Cordillera de los Andes meets the Pacific Ocean, generating a phreatic prism. Coastal resources are further enriched by upwelling of subsurface nutrients; a combination that creates an unusually high marine biomass. This highly productive ecosystem, coupled with terrestrial resources yielded by the wetland provides an abundance and variety of terrestrial and marine fauna and flora, all within the context of an extreme hyperarid environment (Herreros Citation2011; Fariña and Camaño Citation2012; Hernández et al. Citation2014; González and Molina Citation2017; Latorre et al. Citation2017). Interdisciplinary analyses of Caleta Vitor shell middens show long-term subsistence systems and socio-cultural structures based on terrestrial and maritime resources since the beginning of human occupation early in the Holocene (Roberts et al. Citation2013; Carter Citation2016; Disspain et al. Citation2016b; Santoro et al. Citation2017; Martens and Cameron Citation2019; Santoro et al. Citation2020).
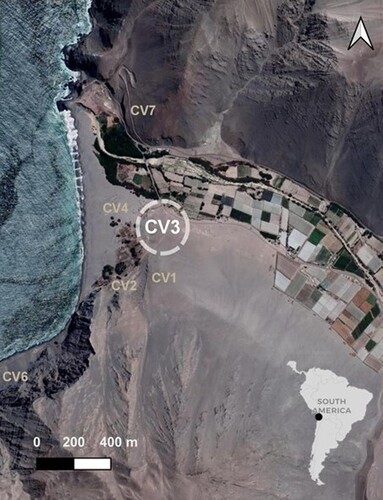
Archaeological surveys in the Caleta Vitor archaeological complex started in the early 2000s and these quickly divided the site into seven areas, named CV1–CV7 (Carter Citation2016; Santoro et al. Citation2017) (). While some areas were synchronically occupied, other areas correspond to different time periods. The earliest occupational sequence corresponds to Caleta Vitor 3 (CV3) (18°45′09.94″ S, 70°20′08.65″ W), which is located at ∼25 m above sea level and 0.3–0.4 km from the present-day shoreline. It comprises an area of 9000 m2 and is composed of three anthropogenic shell middens associated with human burials (Carter Citation2016). Radiocarbon dates established its stratigraphic sequence between 9.2 and 7.6 ka cal BP, during the Early and Middle Holocene (Carter Citation2016; Santoro et al. Citation2017). Fish data in CV3 were first obtained as part of a larger economic study of the site (Carter Citation2016). Later, CV3 was analysed for paleoenvironmental data. In both cases, neither of these studies performed an exhaustive ichthyoarchaeological analysis.
Figure 1. Caleta Vitor archaeological shell midden sites located at the mouth of Quebrada Vitor.
The present study consists in a comparative analysis of two ichthyoarchaeological assemblages recovered using different retrieval methods in CV3. We aimed to contribute to the recovery techniques of fish remains, as well as enabling discussion on archaeological interpretations derived from the ichthyoarchaeological analysis, exemplified in this case study focused on Early to Mid-Holocene human occupations at Caleta Vitor archaeological complex. We also aimed to provide new insights regarding fish and the role of fishing within the Chinchorro communities at the local scale, while also considering regional patterns and the implications for the coastal archaeology of the Atacama Desert.
Materials and Methods
The ichthyoarchaeological assemblages from CV3 come from a trench and two column samples. The first assemblage, here defined as Sample 1, was collected from a trench excavated during the Anillo SOC 1405 project. The second one, corresponding to Sample 2, was collected from two column-samples (CV3/P1 and CV3/P2) as part of the FONDECYT 1150763 project.
The following paragraphs describe the recovery objectives, techniques and analysis of fish remains in each assemblage, establishing comparable categories between both samples. summarises the recovery techniques and analysis methods considered in this comparative analysis.
Table 1. Comparison of recovery techniques and analyses for Samples 1 and 2.
Sample 1
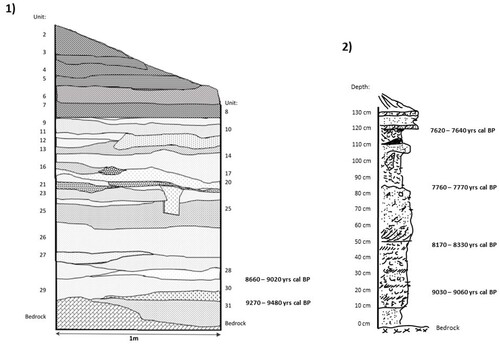
The first survey of CV3 aimed to contribute to the interpretation of the broad temporal range of the Caleta Vitor archaeological complex, identifying its stratigraphic position and cultural materials (Carter Citation2016). Excavation trench CV3/1 covered an area of 100 × 50 cm, with a depth of 180 cm, divided into 31 archaeological stratigraphic levels (Carter Citation2016; Disspain et al. Citation2016b) (). Archaeological remains were sieved using a 1.7 mm mesh screen, collected by hand and bagged according to type. Fish remains were weighed and separated according to the following skeletal elements: jaws (dentary, premaxilla and maxilla), vertebrae, otoliths, shark bones and general items (indeterminate) (Carter Citation2016). Otoliths and fish teeth were considered for taxonomic identification, using published comparative material and ichthyological reference collections (Carter Citation2016).
Figure 2. Trench profile of CV3/1 (Sample 1) (from Carter Citation2016) and CV3/P2 (Sample 2).
Sample 2
Once the CV3 chronological sequence was established, new surveys aimed to obtain more detailed information about the relationship between local cultural and environmental variations. For these, sediment was recovered from two different sections of 20 × 15 cm and 25 × 15 cm (P1 and P2, respectively), with a depth of 130 cm, divided by 5 cm artificial levels (). These sections are located c. 10 m apart, with one profile (P1) done on the south-facing slope of the north mound, whereas P2 was on the north-facing slope of the south mound. Intact sediment sample bags were shipped to the Paleoecology and Paleoenvironmental Laboratory of the Pontificia Universidad Católica de Chile and sieved using different mesh screen sizes (2, 1 mm, <1 mm). All fish remains were recovered, weighed and counted to distinguish the skeletal elements of cranial and postcranial skeletons alongside a reference collection of known taxa (Rebolledo Citation2019). In contrast with the previous analysis, taxonomic identification of the column samples considered all specimens with diagnostic features, and bones of <5 mm were observed using a Zeiss Stemi-C binocular microscope. Finally, considering that taphonomic and anatomo-taxonomic information did not reveal significant differences between P1 and P2 (Rebolledo Citation2019), we considered assemblages from both sections as part of Sample 2.
Comparative Analysis
The comparison between ichthyoarchaeological assemblages comprised a quantitative approach that considered common variables of Samples 1 and 2, along with information relating to the specific characteristics of each assemblage. Considering bone and otoliths weight was the common variable between both samples, we used it to estimate the fish density (grams per litre). Having this, we subsequently calculated absolute and relative frequencies in the total number of specimens (NSP), number of unidentified specimens (UID), number of anatomically identified specimens (NAISP), number of specimens identifiable to taxon (NISP) and minimum number of individuals (MNI) (Grayson Citation1984; Reitz and Wing Citation1999). Finally, we presented distributions of the most relevant species along the stratigraphic sequence of CV3. Fish systematic nomenclature follows Fricke, Eschmeyer, and Van der Laan (Citation2020).
We also included a qualitative perspective to assess the effect of sampling techniques and analytical decisions in data interpretation and discussed skeletal representation in both samples, considering categories previously established in Sample 1 (Carter Citation2016), and its relation to taxonomic identification. The latter also considers the presence of fish species linked to habitat and possible fishing strategies employed.
Results
As shown in , the data available for Samples 1 and 2 do not allow for global comparisons of both assemblages (as already mentioned, data from Sample 1 were not produced within a proper ichthyoarchaeological analysis). As fish weight is the quantitative variable present in both samples, we decided to use this variable to evaluate similarities and variations in fish density between samples.
Table 2. Quantitative ichthyoarchaeological data for Samples 1 (Carter Citation2016) and 2 (Rebolledo Citation2019).
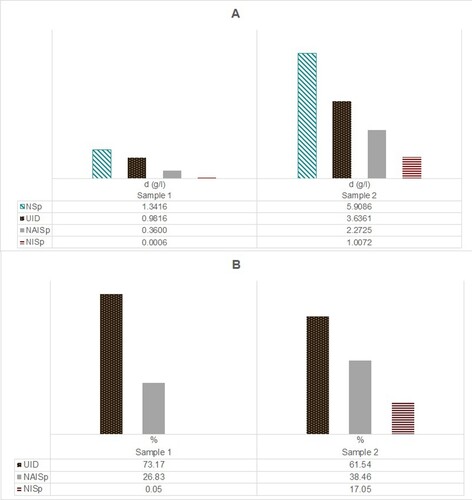
In CV3, Sample 1 is composed of 1207.41 g of fish remains, whereas Sample 2 shows a total weight of 420.25 g. In terms of fish density, Sample 2 shows a higher value (4.77 g/L) than Sample 1 (1.34 g/L) ((A)). When comparing fish density proportions of identified (NAISP) and unidentified fish (UID), Sample 1 shows a lesser percentage of NAISP (26.83%) than Sample 2 (61.54%) ((B)). Also, NISP density is higher in Sample 2 (17.05%) than in Sample 1 (0.05%).
Figure 3. Distribution of fish remains and zooarchaeological indices according to: (A) Density (g/L); and (B) Relative abundance (% of weight) of unidentified and identified specimens.
The differences between NAISP and NISP densities can be explained by pre- and post-depositional processes, as well as by the identification of diagnostic features in specimens. In this regard, there is no evidence of butchery marks, and the percentage of burned bones is particularly low in Sample 2 (1%) (Rebolledo Citation2019). The level of fragmentation is the most important taphonomic variable due to its high percentage in Samples 1 and 2, even though both assemblages present bones and otoliths in a very good condition (Carter Citation2016; Rebolledo Citation2019). The presence of mineral and/or calcareous concretions was also observed on some remains. In fact, the latter affected the taxonomic identification of some bones and otoliths in Sample 2 (Rebolledo Citation2019).
In contrast, divergences in %NISP density between assemblages can be explained by analytical decisions: in Sample 1, we considered mostly otoliths for taxonomic identification, whereas in Sample 2, we included a larger proportion of skeletal elements. Despite this, absolute frequencies in the second assemblage present an NISP of 899 specimens: only 5% of the Sample 2 total. This could be attributed to the recovery of minute fragments which cannot be identified.
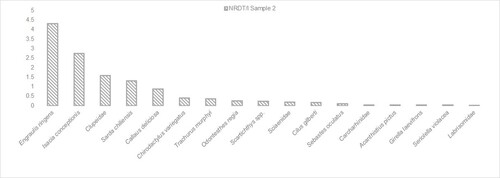
Concerning the taxonomic composition, Sample 1 presents four identified taxa, whereas Sample 2 exhibits 15 fish taxa (). Regarding the species represented in Sample 1, three are included in Sample 2: Callaus deliciosa (syn. Sciaena deliciosa) (lorna drum), Isacia conceptionis (cabinza grunt) and Cilus gilberti (corvina drum). In terms of MNI/l, C. deliciosa and I. conceptionis are the most represented species in Sample 1, whereas I. conceptionis and Engraulis ringens (anchoveta) are the most numerous in Sample 2. We observed a different situation when considering the NISP/l in Sample 2 (), where the small pelagic fish E. ringens was the most common.
Figure 4. Taxa distribution (MNI/l) in Samples 1 and 2.
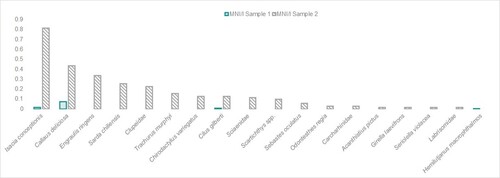
Figure 5. Taxa distribution (NISP/l) in Sample 2.
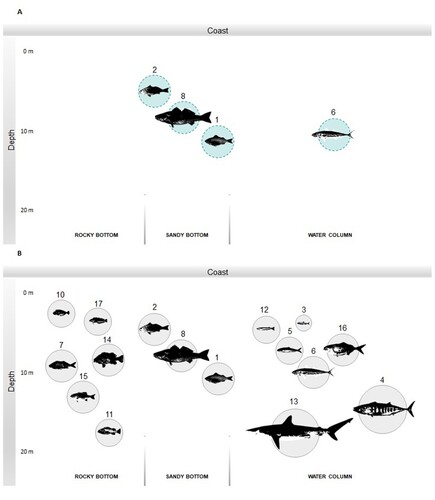
Considering habitat representation according to taxa in Sample 1, C. deliciosa, I. conceptionis, C. gilberti and Hemilutjanus macrophthalmos (grape-eye seabass) are all benthopelagic fishes which live over sandy and rocky bottoms ((A)). With the exception of H. macrophthalmos, Sample 2 presents the same coastal species, alongside Chirodactylus variegatus (Peruvian morwong), Acanthistius pictus (brick seabass), Girella laevifrons (nibbler), Scartichthys spp. (blennies), Sebastes oculatus (Patagonian redfish) and Labrisomidae (labrisomid blennies), suggesting the exploitation of the Caleta Vitor rocky surroundings ((B)). Additionally, small pelagic fish are well represented in Sample 2: E. ringens, Clupeidae (herrings) and Odontesthes regia (Chilean silverside), specifically. Medium-size fish such as Seriolella violacea (palm ruff), Trachurus murphyi (jack mackerel), and other larger fishes, such as Sarda chiliensis (bonito) and Carcharhinidae (requiem sharks) were also identified.
Figure 6. Fish species identified in CV3 according to habitat in (A) Sample 1; and (B) Sample 2. Shoreline is divided into rocky (left) and sandy (right) bottoms. 1: I. conceptionis, 2: C. deliciosa, 3: E. ringens, 4: S. chiliensis, 5: Clupeidae, 6: T. murphyi, 7: C. variegatus, 8: C. gilberti, 10: Scartichthys spp., 11: S. oculatus, 12: O. regia, 13: Carcharhinidae, 14: A. pictus, 15: G. laevifrons, 16: S. violacea, 17: Labrisomidae, 18: H. macrophthalmos. Fish species are illustrated according to size proportions.
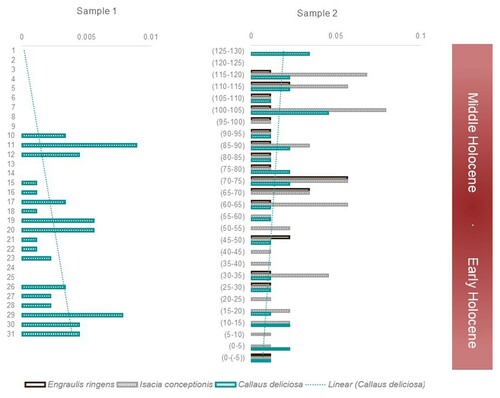
To examine how different habitats were represented, we compared the tendencies of C. deliciosa in Sample 1 with E. ringens and I. conceptionis in Sample 2 (). The former showed a decrease of C. deliciosa MNI/l whereas the opposite situation occurred in Sample 2. An increase of I. conceptionis and E. ringens was also observed. Indeed, the percentage of both species is highly represented when considering NISP/l, with E. ringens being the most abundant taxon since the Early–Middle-Holocene transition (70–75 cm). This coincides with a slight increase in taxa richness during the Middle Holocene, especially of pelagic species (Rebolledo Citation2019).
Figure 7. Taxa distribution through the sequence in Sample 1 (MNI/l C. deliciosa) and Sample 2 (MNI/l E. ringens, I. conceptionis, and C. deliciosa).
Discussion
This comparative analysis exemplifies how contrasting ichthyoarchaeological data arise when different recovery techniques are used. Our results take into account the importance of exhaustive recovery techniques to obtain accurate information on fish remains and increase available data, which in turn can provide further interpretations of fish captures and fishing dynamics. Comparative ichthyoarchaeological samples in CV3 have provided valuable information about fishing dynamics during Early to Middle-Holocene Chinchorro occupations but also have exposed scopes and inconsistencies of comparative analysis when dealing with sets recovered using different techniques. In this regard and before any archaeological interpretation, it is important to evaluate the advantages and limitations of excavation trenches versus column samples.
Excavation Trench or Column Sample?
As a fieldwork archaeological method, big areas but also excavation trenches provide a wider perspective on the archaeological reality under study, thus giving more possibilities to compare with other previously studied regional archaeological sites. Trench excavations provide evidence not only about the cultural materials but also of activity areas, considering the association between artefacts, ecofacts, structures and other spatial features. This is especially important in the case of fish remains, which are intrinsically related to other archaeological evidence such as fishing technology, food processing and other marine resources.
In contrast, the column (or section) sample enables a more detailed study due to its fixed, exhaustive and stratigraphically precise method. It allows for increased understanding of the structure of an archaeological site, providing a detailed understanding with increased chronological and stratigraphical control. Additionally, it is a useful tool for non-archaeological questions associated with other disciplines such as paleoecology, geology, among others. Concerning disadvantages, a column sample does not provide an archaeological spatial perspective, thus hindering interpretations at a local or regional scale.
For the study of fish remains, some authors have proposed to develop pre-excavation analysis using column samples (Nagaoka Citation1994; McKechnie Citation2005) and/or multi-mesh screens (Thomas Citation1969; Nagaoka Citation1994; Partlow Citation2006). In line with Wheeler and Jones (Citation1989), we agree on the necessity of flexibility in recovery techniques, but also that research and fieldwork objectives must consider previous studies to improve regional data and advance beyond just studying archaeological material from single sites.
In the case of CV3, comparative inconsistencies associated with sample size, research objectives, recovery methods and analytical procedures were partially solved thanks to the fish density index. Although none of the aforementioned projects in CV3 were exclusively focused on fish remains, it has still been possible to discuss methodological procedures and to analyse the ichthyoarchaeological materials from this site.
Fish Data Standardisation
The importance of data standardisation to perform comparative analyses from a local to a macro-regional scale should also be stressed. Several archaeofaunal studies have used density to assess the spatial and temporal distribution of animal remains (e.g. Jerardino Citation2016), and the use of weight in density estimation has already been highlighted for its advantages in standardising archaeological information (Orton Citation2000). In our case, and considering the information available, we used weight to compare fish density in Samples 1 and 2. The results showed higher values in Sample 2, although the volume excavated was much smaller than for Sample 1. The increase of fish density in this sample suggests an increase in abundance per volume excavated, possibly explained by the recovery techniques employed. But how advantageous are these procedures for anatomical and taxonomical identification?
Regarding anatomical representation, there was a large proportion of unidentified remains (UID) possibly due to the level of fragmentation of some specimens and to a lesser extent to other post-depositional processes (e.g. presence of concretions, weathering). In contrast, the NAISP includes cranial and postcranial parts in both samples, although the use of skeletal elements in taxonomic identification differed. In Sample 1, otoliths were the main element used in fish identifications. Otoliths have been widely used in ichthyoarchaeological analysis due to their density and diagnostic features (e.g. Llagostera Citation1979; Falabella, Vargas, and Meléndez Citation1994; Llagostera, Kong, and Irachet Citation1997; Béarez Citation2000; Disspain, Ulm, and Gillanders Citation2016a, Citation2016b). However, their preservation potential in an archaeological context is highly dependent on depositional processes, soil pH and, most importantly, on the species. In the case of CV3, the largest proportion corresponds to Haemulidae and Sciaenidae, both medium-sized fishes with well-developed otoliths compared to other species represented in Sample 2. In this sample, postcranial elements and other cranial bones contributed to the taxonomic identification of different fish species.
Even though we observe an increase in the richness of species, this is not proportional to the percentage of taxonomic identification considering the total number of specimens in Sample 2. This problem has already been discussed by several researchers (e.g. Watson Citation1972; Casteel Citation1976a; Nagaoka Citation1994, Citation2005; Orton Citation2000), who pointed out the importance of constant revision regarding an archaeological site features, recovery techniques and laboratory procedures when working with ichthyoarchaeological material.
The variety of pelagic fish in Sample 2 is an important feature that stands out, especially because the identification of so many fish species would not have been possible using the recovery techniques and analysis methods used for Sample 1 (). This actually has important consequences regarding how ichthyoarchaeological evidence is used for archaeological interpretations, and hence archaeological reconstructions regarding fishing dynamics for the Chinchorro culture.
Big Fish or Small Fish?
Although fish remains have been recorded in northern Chile since the beginning of excavations (Bird Citation1943), these materials have not been part of a systematic anatomo-taxonomic analysis. Contemporary archaeological sites close to Caleta Vitor 3 show differences based on a variety of recovery techniques and analyses that often do not allow for comparisons from published data. According to published data from Acha-2 site, only fish vertebrae were considered (Muñoz, Arriaza, and Aufderheide Citation1993), whereas at Tiliviche 1B site total fish remains were counted (Hesse Citation1984). There is little information about the types of fish present in both archaeological sites: at Acha-2, authors proposed the capture of different sizes of fish, as well as the presence of pelagic species captured after stranding during the summer months (Muñoz, Arriaza, and Aufderheide Citation1993). At Tiliviche 1B site, the presence of small freshwater fish such as Trichomycteridae, Characidae and Atherinidae is mentioned (Núñez and Hall Citation1982). Meanwhile, nine taxa from coastal waters were identified at Camarones 14 site: S. chiliensis, T. murphyi, Paralichthys sp., Semicossyphus sp., Cilus gilberti, Sciaena sp., Scomber japonicus, Mugil cephalus and Myliobatis peruvianus (Schiappacasse and Niemeyer Citation1984).
At CV3, we observed that coastal species were well represented in both assemblages. The difference lies in pelagic fish, which are most represented in Sample 2 (). Among the species of the different size found in this sample, the presence of small fish species stands out. Furthermore, fluctuations in the abundance of Clupeiformes through the occupational sequence suggest temporal variations in fishing activity. This could indicate access to other habitats farther from the shoreline, and/or environmental fluctuations that affected some species in the coastal area. In any case, this information will add to discussions on fishing strategies of Early and Mid-Holocene occupations in the area.
Previous studies have proposed the use of nets from the Caleta Vitor shoreline since the beginning of human occupation (Carter Citation2016; Disspain et al. Citation2016b; Santoro et al. Citation2017, Citation2020; Martens and Cameron Citation2019). Moreover, Martens and Cameron (Citation2019) have proposed a long fibre technology tradition in Chinchorro and later coastal communities, among other productive activities. The earliest archaeological evidence of fibre technology is present in the Middle Holocene occupations in Caleta Vitor 1, where use of vegetal and animal raw materials for producing fibres have been identified. In CV3, indirect evidence from a shell bead from an Early Holocene layer and large amounts of small fish support the idea of cordage and fishing nets, respectively (Carter Citation2016; Martens and Cameron Citation2019). Additionally, Disspain et al. (Citation2016b) have suggested the use of nets since the Early Holocene due to the presence of Sciaenidae, a fish family that can be easily caught using nets due to their proximity to the shoreline.
Nets can be very diverse both in their construction and in the way they are used. They can be passive (gillnet) or active (beach seine), and their mesh size can be adapted depending on the species targeted. Indeed, variations in habitat exploitation or in the size or behaviour of fish may have implied the use of one or the other type of net/mesh to capture a wide variety of species. The handling of nets often further requires the investment of several people, and their use could have implied another type of cooperative work related to the acquisition of raw materials, construction, fish procurement and gear maintenance, thus modelling the social organisation of the communities (Hilger Citation1951; Smith Citation1957; Antropova Citation1964; Newton Citation1985; Parmentier Citation2011; Beresford-Jones et al. Citation2018).
Direct and indirect evidence of the use of fishing nets is present throughout the arid Pacific coast, from southern Peru (Sandweiss et al. Citation1989, Citation1998; deFrance and Umire Citation2004, Citation2009; Béarez Citation2012; Reitz et al. Citation2017), to the southernmost part of the Atacama Desert (Llagostera Citation1989; Casteletti Citation2007; Rebolledo et al. Citation2016; Salazar et al. Citation2018). These authors also highlight important fluctuations in fish composition between the Early and Mid-Holocene human occupations, strongly related to the change of fishing strategies. In most cases, this is explained by the appearance of large pelagic species (see Olguín, Salazar, and Jackson Citation2014; Béarez et al. Citation2016), and even though small fish species are present at these sites, these remains are often hardly discussed.
Along with changes in fishing strategies, these fluctuations could also be explained by a seasonal use of archaeological sites. At CV3, the species most represented in Samples 1 and 2 such as drums and rock fish allow year-round access, and although anchoveta approach shorelines in summer, it is not possible to infer seasonality patterns in fishing practices from available data.
Understanding variations and continuities of fishing dynamics require that we consider all fish species and the whole spectrum of devices used to capture them. Specific to ichthyoarchaeological studies, interpretations tend to focus on a particular kind of fish, which obscures fishing strategies associated with other species and habitats (Matsui Citation1996; Partlow Citation2006). In the Atacama Desert Coast, the discussion has centred on medium- and large-sized fishes that inhabit waters farther from the shoreline, and the study of fishing technologies such as fishhooks, harpoons, and watercraft. As a consequence, the archaeological narrative tends to emphasise the importance of large-size resources, reproducing the terrestrial paradigm of big-game hunting. Establishing new research questions through the study of different fish species and fishing strategies could shed new light on fishing and its implications for coastal communities.
Conclusion
The 9000-year-old historical sequence present at Caleta Vitor constitutes a promising example to evaluate fishing practices in northern Chile. Comparative analysis of samples from CV3 Early to Middle Holocene occupations reveals how recovery techniques and methods of analysis influence ichthyoarchaeological data production. In this sense, small mesh screen size allows for a more comprehensive retrieval of fish remains, including a broader skeletal and taxonomical representation. Furthermore, Sample 2 contributes new fish species which advocates for a reinterpretation of fishing dynamics by the earlier occupations in Caleta Vitor.
Despite their small spatial representativity, both samples established preliminary approaches to fishing strategies through time. Further research must consider a spatial and synchronic perspective to provide a more complete interpretation of cultural processes associated with fishing during the earlier occupations of Caleta Vitor. Access to open data sources will be necessary to develop new research dynamics and to perform comparative analysis at a macro-regional scale using statistical software. Such studies have the potential to reveal a new level of aspects linked to fishing by the prehistoric coastal communities and the inhabitants’ relationship with the Pacific Ocean.
Acknowledgments
The authors are grateful to V. Hurtado, M. Yilales, S. Monsalve and C. Pozo for help in the field and curating Sample 2 from CV3, within the FONDECYT 1150763 project. The authors thank Diego Salazar from Departamento de Antropología, Universidad de Chile for providing the reference collection used in ichthyoarchaeological analysis of Sample 2. The authors would also like to thank Jill Cucchi for copy editing. The authors gratefully acknowledge the suggestions of the two anonymous reviewers. Finally, we acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Disclosure Statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Sandra Rebolledo
Sandra Rebolledo is an archaeozoologist focused on ichthyoarchaeology and social archaeology. Particularly, she is interested in subsistence strategies in hunter-gatherer-fisher communities along the South American western coast. She has worked in the Atacama Desert Coast (northern Chile) and the Chiloean Archipelago.
Philippe Béarez
Philippe Béarez is an archaeozoologist specialised in the study of interactions between humans and aquatic environments over time. He is also interested in historical ecology and human impacts on aquatic ecosystems. He carries out archaeozoological work mostly in South America (eastern Pacific coast, Amazonia) but also in the Indian ocean (Arabian sea coast), Oceania (Melanesia, Polynesia) and France.
Débora Zurro
Débora Zurro is particularly interested in the methodological development of archaeology and her research focus is making much more visible secondary resources, such as plant consumption. She has worked for several years in (ethno)archaeological projects about coastal hunter-gatherer economies in Southamerica and also on the analysis of androcentrism in archaeological discourses.
Calogero M. Santoro
Calogero M. Santoro seeks to understand the human life systems that have developed and transformed the ecosystems of the Atacama Desert, from the most remote past to the present and to contribute to the understanding of human history in general and to the debate on the sustainability of the planet.
Claudio Latorre
Claudio Latorre is a Quaternary paleoecologist interested in understanding the complex relationship between climate, ecology, and human history in arid and semiarid environments including the Atacama Desert. Together with his colleagues and students, his research spans from historical times to the last glacial period.
References
- Albarella, U.. 2017. “Zooarchaeology in the Twenty-First Century: Where We Come from, Where We Are Now, and Where We Are Going.” In The Oxford Handbook of Zooarchaeology edited by Umberto Albarella, Mauro Rizzetto, Hannah Russ, Kim Vickers, and Sarah Viner-Daniels, 3–21. Oxford: Oxford University Press . ISBN 9780199686476.
- Antropova, V. V. 1964. “The Koryaks.” In The Peoples of Siberia, edited by M. G. Levin, and L. P. Potapov, 851–875. Chicago, IL: University of Chicago Press.
- Arriaza, B. T. 1995. “Chinchorro Bioarchaeology: Chronology and Mummy Seriation.” Latin American Antiquity 6 (1): 35–55.
- Arriaza, B. T., M. Doubrava, V. G. Standen, and H. Haas. 2005. “Differential Mortuary Treatment among the Andean Chinchorro Fishers: Social Inequalities or in Situ Regional Cultural Evolution?” Current Anthropology 46: 662–671. doi:10.1086/432747.
- Arriaza, B. T., V. G. Standen, V. Cassman, and C. M. Santoro. 2008. “Chinchorro Culture: Pioneers of the Coast of the Atacama Desert.” In The Handbook of South American Archaeology, edited by Helaine Silverman and William H. Isbell, 45–58. New York: Springer. doi:10.1007/978-0-387-74907-5.
- Barrett, J. H. 1997. “Fish Trade in Norse Orkney and Caithness: A Zooarchaeological Approach.” Antiquity 71 (273): 616–638. doi:10.1017/S0003598X00085367.
- Béarez, P. 2000. “Archaic Fishing at Quebrada de los Burros, Southern Coast of Peru. Reconstruction of Fish Size by Using Otoliths.” Archaeofauna 9: 29–34.
- Béarez, P. 2012. “Los Peces y la Pesca.” In Prehistoria de la Costa Extremo-sur del Perú, edited by Danièle Lavallée, and Michèle Julien, 99–123. Lima: Instituto Francés de Estudios Andinos - Pontificia Universidad Católica del Perú.
- Béarez, P., F. Fuentes-Mucherl, S. Rebolledo, D. Salazar, and L. Olguín. 2016. “Billfish Foraging Along the Northern Coast of Chile During the Middle Holocene (7400-5900 cal BP).” Journal of Anthropological Archaeology 41: 185–195. doi:10.1016/j.jaa.2016.01.002.
- Beresford-Jones, D., A. Pullen, G. Chauca, L. Cadwallader, M. García, I. Salvatierra, O. Whaley, et al. 2018. “Refining the Maritime Foundations of Andean Civilization: How Plant Fiber Technology Drove Social Complexity During the Preceramic Period.” Journal of Archaeological Method and Theory 25 (2): 393–425. doi:10.1007/s10816-017-9341-3.
- Bird, J. 1943. “Excavations in Northern Chile.” Anthropological Papers of the American Museum of Natural History, New York 38 (4): 173–318.
- Buckley, H. R., and J. E. Buikstra. 2019. “Stone Agers in the Fast Lane? How Bioarchaeologists Can Address the Paleo Diet Myth.” In Bioarchaeologists Speak Out: Deep Time Perspectives on Contemporary Issues, edited by Jane E. Buikstra, 161–180. Cham: Springer.
- Butler, V. L. 1993. “Natural Versus Cultural Salmonid Remains: Origin of The Dalles Roadcut Bones, Columbia River, Oregon, U.S.A.” Journal of Archaeological Science 20: 1–24.
- Carter, C. P. 2016. “The Economy of Prehistoric Coastal Northern Chile: Case Study: Caleta Vitor.” Thesis submitted for the degree of Doctor of Philosophy of The Australian National University.
- Casteel, R. W. 1972. “Some Biases in the Recovery of Archaeological Faunal Remains.” Proceedings of the Prehistoric Society 38: 382–388. doi:10.1017/S0079497X00012172.
- Casteel, R. W. 1976a. “Comparison of Column and Whole Unit Samples for Recovering Fish Remains.” World Archaeology 8 (2): 192–196. doi:10.1080/00438243.1976.9979663.
- Casteel, R. W. 1976b. Fish Remains in Archaeology and Paleo-Environmental Studies. New York: Academic Press.
- Casteletti, J. 2007. “Patrón de asentamiento y uso de recursos a través de la secuencia ocupacional prehispana de la costa de Taltal.” Memoria para optar al grado de Magister en Antropología con mención en Arqueología, Universidad Católica del Norte y Universidad de Tarapacá.
- Colley, S. M. 1990. “The Analysis and Interpretation of Archaeological Fish Remains.” Archeological Method and Theory 2: 207–253.
- deFrance, S. D., and A. Umire. 2004. “Quebrada Tacahuay: Un Sitio Marítimo del Pleistoceno Tardío en la Costa sur del Perú.” Chungara, Revista de Antropología Chilena 36 (2): 257–278.
- deFrance, S. D., and A. Umire. 2009. “Quebrada Tacahuay and Early Maritime Foundations on Peru’s Far Southern Coast.” In Andean Civilization. A Tribute to Michael E. Moseley, edited by Joyce Marcus, and Patrick R. Williams, 55–74. Los Angeles: Cotsen Institute of Archaeology, University of California. doi:10.2307/j.ctvdmwx3 h.6.
- Disspain, M. C. F., S. Ulm, and B. M. Gillanders. 2016a. “Otoliths in Archaeology: Methods, Applications and Future Prospects.” Journal of Archaeological Science: Reports 6: 623–632. doi:10.1016/j.jasrep.2015.05.012.
- Disspain, M. C. F., S. Ulm, C. M. Santoro, C. Carter, and B. M. Gillanders. 2016b. “Pre-Columbian Fishing on the Coast of the Atacama Desert, Northern Chile: An Investigation of Fish Size and Species Distribution Using Otoliths from Camarones Punta Norte and Caleta Vitor.” Journal of Island and Coastal Archaeology 00: 1–23. doi:10.1080/15564894.2016.1204385.
- Falabella, F., L. Vargas, and R. Meléndez. 1994. “Differential Preservation and Recovery of Fish Remains in Central Chile.” In Fish Exploitation in the Past. Proceedings of the 7th meeting of the ICAZ Fish Remains Working Group, edited by W. van Neer, 25–35. Tervuren: Annales du Musèe Royal de L’Afrique Central, Science Zoologiques No. 274.
- Fariña, J. M., and A. Camaño. 2012. Humedales costeros de Chile. Aportes científicos a su gestión sustentable. Santiago de Chile: Ediciones Universidad Católica de Chile.
- Fricke, R., W. N. Eschmeyer, and R. Van der Laan, (eds). 2020. “Eschmeyer’s Catalog of Fishes: Genera, Species, References.” Accessed 5 May 2020. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Electronic version.
- Garson, A. G. 1980. “Comment upon the Economic Potential of Fish Utilization in Riverine Environments and Potential Archaeological Biases.” American Antiquity 45 (3): 562–567. doi:10.2307/279874.
- González, J., and J. Molina. 2017. Flora Nativa de la Región de Arica y Parinacota. Arica: Ediciones Universidad de Tarapacá.
- Grayson, D. K. 1984. Quantitative Zooarchaeology. Topics in the Analysis of Archaeological Faunas. Orlando: Academic Press.
- Gurven, M., and K. Hill. 2009. “Why do men Hunt? A Reevaluation of ‘Man the Hunter’ and the Sexual Division of Labor.” Current Anthropology 50 (1): 51–74. doi:10.1086/595620.
- Hernández, J., C. Estades, L. Faúndez, and J. Herreros. 2014. In Biodiversidad Terrestre de la Región de Arica y Parinacota, edited by Jaime Hernández, Cristián Estades, Luis Faúndez, and Jorge Herreros. Santiago: Ediciones Universidad de Chile.
- Herreros, J. 2011. Biodiversidad del Borde Costero de Arica. Arica: Ministerio del Medio Ambiente.
- Hesse, B. 1984. “Archaic Exploitation of Small Mammals and Birds in Northern Chile.” Estudios Atacameños. Arqueología y Antropología Surandinas 51 (7): 37–51. doi:10.22199/s07181043.1984.0007.00005.
- Hildebrandt, W. R., and K. R. McGuire. 2003. “Large-game Hunting, Gender-Differentiated Work Organization, and the Role of Evolutionary Ecology in California and Great Basin Prehistory: A Reply to Broughton and Bayham.” American Antiquity 68 (4): 790–792.
- Hilger, M. I. 1951. “Chippewa Child Life and its Cultural Background.” Bureau of American Ethnology Bulletin 146: 1–204.
- Hill, E. 2013. “Archaeology and Animal Persons: Toward a Prehistory of Human-Animal Relations.” Environment and Society 4: 117–136. doi:10.3167/ares.2013.040108.
- Jerardino, A. 2016. “On the Origins and Significance of Pleistocene Coastal Resource Use in Southern Africa with Particular Reference to Shellfish Gathering.” Journal of Anthropological Archaeology 41: 213–230. doi:10.1016/j.jaa.2016.01.001.
- Konner, M., and S. B. Eaton. 2010. “Paleolithic Nutrition: Twenty-Five Years Later.” Nutrition in Clinical Practice 25 (6): 594–602. doi:10.1177/0884533610385702.
- Landon, D. 2005. “Zooarchaeology and Historical Archaeology: Progress and Prospects.” Journal of Archaeological Method and Theory 12 (1): 1–36.
- Latorre, C., R. de Pol-Holz, C. Carter, and C. M. Santoro. 2017. “Using Archaeological Shell Middens as a Proxy for Past Local Coastal Upwelling in Northern Chile.” Quaternary International 427: 128–136. doi:10.1016/j.quaint.2015.11.079.
- Llagostera, A. 1979. “9,700 Years of Maritime Subsistence on the Pacific: An Analysis by Means of Bioindicators in the North of Chile.” American Antiquity 44 (2): 309–324.
- Llagostera, A. 1989. “Caza y Pesca Marítima (9.000 a 1.000 a.C.).” In Prehistoria: desde sus orígenes hasta los albores de la Conquista, edited by Jorge Hidalgo, Virgilio Schiappacasse, Hans Niemeyer, Carlos Aldunate, and Iván Solimano, 57–79. Santiago de Chile: Editorial Andrés Bello.
- Llagostera, A., I. Kong, and P. Irachet. 1997. “Análisis ictioarqueológico del sitio La Chimba 13 (II Región, Chile).” Chungara, Revista de Antropología Chilena 29 (2): 163–179.
- Martens, T., and J. Cameron. 2019. “Early Coastal Fiber Technology from the Caleta Vitor Archaeological Complex in Northern Chile.” Latin American Antiquity 30 (2): 287–299. doi:10.1017/laq.2018.78.
- Matsui, A. 1996. “Archaeological Investigations of Anadromous Salmonid Fishing in Japan.” World Archaeology 27 (3): 444–460. doi:10.1080/00438243.1996.9980319.
- McKechnie, I. 2005. “Column Sampling and the Archaeology of Small Fish at Ts’ishaa.” In Ts’ishaa: Archaeology and Ethnography of a Nuu-chah-nulth Origin Site in Barkley Sound, edited by Alan D. McMillan and Denis E. st. Claire, 206–223. Burnaby: Archaeology Press. Simon Fraser University.
- Meadow, R. H. 1980. “Animal Bones: Problems for the Archaeologist Together with Some Possible Solutions.” Paléorient 6 (1): 65–77. doi:10.3406/paleo.1980.4260.
- Mueller, J. W. 1974. “I. The Theoretical Framework.” Memoirs of the Society for American Archaeology 28: 1–6.
- Muñoz, I., B. Arriaza, and A. Aufderheide. 1993. In Acha-2 y los orígenes del poblamiento humano en Arica, edited by Ivan Muñoz, Bernardo Arriaza, and Arthur Aufderheide. Arica: Ediciones Universidad de Tarapacá.
- Nagaoka, L. 1994. “Differential Recovery of Pacific Island Fish Remains: Evidence from the Moturakau Rockshelter, Aitutaki, Cook Islands.” Asian Perspectives 33 (1): 1–17.
- Nagaoka, L. 2005. “Differential Recovery of Pacific Island Fish Remains.” Journal of Archaeological Science 32 (6): 941–955. doi:10.1016/j.jas.2004.12.011.
- Newton, J. 1985. “Technology and Cooperative Labour among the Orokaiva.” MANKIND 15 (3): 214–222. doi:10.1111/j.1835-9310.1985.tb01406.
- Núñez, L., and H. J. Hall. 1982. “Análisis de dieta y movilidad en un campamento arcaico del norte de Chile.” Bulletin de l’Institut Français d’Études Andines 3–4: 91–113.
- Olguín, L., D. Salazar, and D. Jackson. 2014. “Tempranas evidencias de navegación y caza de especies oceánicas en la costa pacífica de Sudamérica (Taltal, ∼7,000 cal A.P.).” Chungara, Revista de Antropología Chilena 46 (2): 177–192.
- Orton, C. 2000. Sampling in Archaeology. Cambridge Manuals in Archaeology. Cambridge: Cambridge University Press.
- Parmentier, R. J. 2011. Culture Summary: Belauans. New Haven, CT: Human Relations Area Files.
- Partlow, M. A. 2006. “Sampling Fish Bones: A Consideration of the Importance of Screen Size and Disposal Context in the North Pacific.” Arctic Anthropology 43 (1): 67–79.
- Payne, S. 1972. “Partial Recovery and Sample Bias: The Results of Some Sieving Experiments.” In Papers in Economic Prehistory, edited by E. S. Higgs, 49–64. Cambridge: British Academy.
- Peregrine, P. N. 2002. “Sampling in Archaeology.” Geoarchaeology 17 (5): 483–484. doi:10.1002/gea.10024.
- Rebolledo, S. 2019. Informe ictioarqueológico Caleta Vitor 3. Report Fondecyt 1150763.
- Rebolledo, S., P. Béarez, D. Salazar, and F. Fuentes. 2016. “Maritime Fishing During the Middle Holocene in the Hyperarid Coast of the Atacama Desert.” Quaternary International 391: 3–11. doi:10.1016/j.quaint.2015.09.051.
- Reitz, E. J. 2009. “International Council for Archaeozoology (ICAZ) Professional protocols for archaeozoology.” http://alexandriaarchive.org/icaz/pdf/protocols2009.pdf. Electronic version accessed 10 nov 2020.
- Reitz, E. J., H. E. McInnis, D. H. Sandweiss, and S. D. DeFrance. 2017. “Variations in Human Adaptations During the Terminal Pleistocene and Early Holocene at Quebrada Jaguay (QJ-280) and the Ring Site, Southern Perú.” The Journal of Island and Coastal Archaeology 12 (2): 224–254. doi:10.1080/15564894.2016.1172381.
- Reitz, E. J., and E. S. Wing. 1999. Zooarchaeology. Cambridge Manuals in Archaeology. Cambridge: Cambridge University Press.
- Roberts, A., F. Donald Pate, B. Petruzzelli, C. Carter, M. C. Westaway, C. M. Santoro, J. Swift, et al. 2013. “Retention of Hunter-Gatherer Economies among Maritime Foragers from Caleta Vitor, Northern Chile, During the Late Holocene: Evidence from Stable Carbon and Nitrogen Isotopic Analysis of Skeletal Remains.” Journal of Archaeological Science 40 (5): 2360–2372. doi:10.1016/j.jas.2013.01.009.h.
- Ross, A., and R. Duffy. 2000. “Fine Mesh Screening of Midden Material and the Recovery of Fish Bone: The Development of Flotation and Deflocculation Techniques for an Efficient and Effective Procedure.” Geoarchaeology 15 (1): 21–41. doi:10.1002/(SICI)1520-6548(200001)15:1< 21::AID-GEA2 > 3.0.CO;2-L.
- Russell, N. 2011. Social Zooarchaeology: Humans and Animals in Prehistory. New York: Cambridge University Press. doi:10.1017/cbo9781139019712.
- Salazar, D., C. Arenas, P. Andrade, L. Olguín, J. Torres, C. Flores, G. Vargas, et al. 2018. “From the use of Space to Territorialisation During the Early Holocene in Taltal, Coastal Atacama Desert, Chile.” Quaternary International 473: 225–241. doi:10.1016/j.quaint.2017.09.035.
- Salazar, D., C. Flores, C. Borie, L. Olguín, S. Rebolledo, M. Escobar, and A. Cifuentes. 2020. “Marine Communities in the Atacama Desert. “Economic Organization and Social Dynamics of Middle-Holocene Hunter-Gatherer-Fisher Communities on the Coast of the Atacama Desert (Taltal, Northern Chile).” In New Perspectives on the Social Dynamics and Economic Interactions of Andean Maritime Communities, edited by Gabriel Prieto and DanielSandweiss, 74–100. Florida: University Press of Florida.
- Sandweiss, D. H., H. Mcinnis, R. L. Burger, A. Cano, B. Ojeda, R. Paredes, C. Sandweiss, et al. 1998. “Quebrada Jaguay : Early South American Maritime Adaptations.” Science 281 (5384): 1830–1832.
- Sandweiss, D., J. B. Richardson III, E. Reitz, and J. T. Hsu. 1989. “Early Maritime Adaptations in the Andes: Preliminary Studies at the Ring Site, Peru.” In Ecology, Settlement, and History in the Osmore Drainage, Peru, edited by D. S. Rice, C. Stanish, and P. R. Scar, 35–84. Oxford: BAR International Series 545 (i).
- Santoro, C. M., V. Castro, C. Carter, and D. Valenzuela. 2020. “Marine Communities in the Atacama Desert. Masters of the sub-Tropical Pacific Coast of South America.” In New Perspectives on the Social Dynamics and Economic Interactions of Andean Maritime Communities, edited by Gabriel Prieto and Daniel Sandweiss, 39–73. Florida: University Pres of Florida.
- Santoro, C. M., E. M. Gayo, C. Carter, V. G. Standen, V. Castro, D. Valenzuela, R. de Pol-Holz, P. A. Marquet, and C. Latorre. 2017. “Loco or no Loco? Holocene Climatic Fluctuations, Human Demography, and Community Based Management of Coastal Resources in Northern Chile.” Frontiers in Earth Science 5: 1–16. doi:10.3389/feart.2017.00077.
- Santoro, C. M., M. M. Rivadeneira, C. Latorre, F. Rothhammer, and V. G. Standen. 2012. “Rise and Decline of Chinchorro Sacred Landscapes Along the Hyperarid Coast of the Atacama Desert.” Chungara, Revista de Antropología Chilena 44 (4): 637–653. doi:10.4067/S0717-73562012000400007.
- Schiappacasse, V., and H. Niemeyer. 1984. “Descripción y análisis interpretativo de un sitio Arcaico Temprano en la Quebrada de Camarones.” Publicación Ocasional Museo Nacional de Historia Natural y Universidad de Tarapacá 41: 1–187.
- Schiffer, M. B., A. P. Sullivan, and T. C. Klinger. 1978. “The Design of Archaeological Surveys.” World Archaeology 10 (1): 1–28. doi:10.1080/00438243.1978.9979712.
- Shaffer, B. S. 1992. “Quarter-Inch Screening: Understanding Biases in Recovery of Vertebrate Faunal Remains.” American Antiquity 57 (1): 129–136.
- Shaffer, B. S., and J. L. J. Sanchez. 1994. “Comparison of 1/8″- and 1/4″-Mesh Recovery of Controlled Samples of Small-to-Medium-Sized Mammals.” American Antiquity 59 (3): 525–530.
- Smith, M. G. 1957. “Cooperation in Hausa Society.” Information 11: 1–20. Paris: International Social Science Council.
- Stahl, P. W. 1996. “The Recovery and Interpretation of Microvertebrate Bone Assemblages from Archaeological Contexts.” Journal of Archaeological Method and Theory 3 (1): 31–75. doi:10.1007/BF02228930.
- Standen, V. G., and C. M. Santoro. 2004. “Patrón funerario arcaico temprano del sitio Acha-3 y su relación con Chinchorro: Cazadores, pescadores y recolectores de la costa norte de Chile.” Latin American Antiquity 15 (1): 89–109. doi:10.2307/4141565.
- Standen, V. G., C. M. Santoro, B. T. Arriaza, and D. Coleman. 2018. “Hunting, Gathering, and Fishing on the Coast of the Atacama Desert: Chinchorro Population Mobility Patterns Inferred from Strontium Isotopes.” Geoarchaeology 33 (2): 162–176. doi:10.1002/gea.21594.
- Struever, S. 1968. “Flotation Techniques for the Recovery of Small-Scale Archaeological Remains.” American Antiquity 33 (3): 353–362.
- Thomas, D. H. 1969. “Great Basin Hunting Patterns: A Quantitative Method for Treating Faunal Remains.” American Antiquity 34 (4): 392–401. doi:10.2307/277736.
- Watson, J. P. N. 1972. “Fragmentation Analysis of Animal Bone Samples from Archaeological Sites.” Archaeometry 14 (2): 221–228. doi:10.1111/j.1475-4754.1972.tb00064.
- Wheeler, A., and A. Jones. 1989. Fishes. Cambridge: Cambridge University Press.
- Zohar, I., and M. Belmaker. 2005. “Size Does Matter: Methodological Comments on Sieve Size and Species Richness in Fishbone Assemblages.” Journal of Archaeological Science 32 (4): 635–641. doi:10.1016/S0305-4403(03)00037-2.