
Abstract
We investigated a panel of 14 compounds belonging to the monothiocarbamate (MTC) and dithiocarbamate (DTC) series against the β-carbonic anhydrase 3 (β-CA3) of Mycobacterium tuberculosis (Mtb). We also evaluated all compounds for toxicity using 1–5-day post fertilisation zebrafish embryos. 11 out of the 14 investigated derivatives showed effective nanomolar or submicromolar in vitro inhibition against the β-CA3 (KIs 2.4–812.0 nM), and among them four DTCs of the series (8–10 and 12) showed very significant inhibition potencies with KIs between 2.4 and 43 nM. Out of 14 compounds screened for toxicity and safety 9 compounds showed no adverse phenotypic effects on the developing zebrafish larvae at five days of exposure. The results of in vitro inhibition and the toxicological evaluation of our study suggest that 5 compounds are suitable for further in vivo preclinical characterisation in zebrafish model.
Introduction
Mtb is a highly infectious microbial species that causes tuberculosis (TB) in humans. The latest World Health Organisation report on infectious diseases estimated that annually, 10 million people develop TB and 1.6 million die from the diseaseCitation1. Quite worryingly, the same study reported >550 000 infections to be caused by rifampin-resistant strains that are unresponsive to this first-line drug against Mtb. Among these strains, 80% were multidrug-resistant (MDR-TB)Citation1. Therefore, there is an urgent need for antibiotics targeting novel physiological pathways of MtbCitation2–4.
Mycobacteria encode for at least three zinc-containing metalloenzyme carbonic anhydrases (CAs, EC 4.2.1.1) that belong to the β-CA gene familyCitation1,Citation2. Since the primary function of β-CAs in mycobacteria is to reversibly catalyse the hydration of CO2 to generate HCO3− and H+ ionsCitation5,Citation6, such enzymes are involved in a multitude of physiological processes closely related to pH homeostasis, biosynthetic processes as well as adaption to the environmentsCitation3,Citation7–10. In addition, mycobacterial β-CAs have been shown to be essential for the transport of extracellular DNA (eDNA)Citation11. It was also reported that the administration of the non-selective CA Inhibitor (CAI) ethoxzolamide (EZA) reduced transport of eDNA and affected the formation of biofilms in non-tuberculous mycobacteria (NTM)Citation11. Another study conducted on Mycobacterium tuberculosis (Mtb) strains, demonstrated that EZA inhibited the two-component PhoPR regulon as well as the Esx-1 protein secretion system, which are fundamental for the virulence of the bacteriumCitation12. EZA also showed efficacy in infected macrophages and mice suggesting that β-CAs perform very important roles in mycobacterial infectionsCitation12, and hence present themselves as potential drug targets. The three Mtb β-CAs (i.e. β-CA1, β-CA2 and β-CA3) are encoded by Rv1284, Rv3588c and Rv3273 genes, respectivelyCitation8,Citation13–16. Among the compounds tested in vitro for their inhibition properties against such enzymes are the classical CAIs of the sulphonamide typeCitation17,Citation18, and the recently reported dithio- and monothiocarbamatesCitation19,Citation20 (DTCs and MTCs) whose general structures are shown in .
Figure 1. General structures of dithiocarbamates (DTC) and monothiocarbamates (MTC).
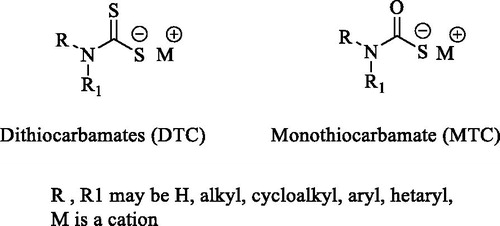
Both DTCs and MTCs have been explored, with good outcomes, against several CAs from pathogens expressing α-, β-, and δ-CA enzymesCitation19–23. A study conducted in our laboratory showed that a DTC-based compound (i.e. derivative 12 in this study) not only effectively inhibited the β-CAs of Mtb but also impaired the growth of Mycobacterium marinum, a close relative of Mtb, in zebrafish larvaeCitation2. The data obtained by means of in vitro and in vivo experiments involving DTCs and MTCs encouraged us to further investigate similar compounds against the Mtb β-CA3 with the aim to find new potential inhibitors targeting mycobacterial β-CAsCitation2,Citation21.
Materials and methods
Chemistry
Compounds 1-14 considered in this manuscript were synthesised according to the procedures previously reported by some of us, and they were properly characterised by means of 1H-NMR, 13 C-NMR, and mass spectra analysisCitation24–27.
In vitro carbonic anhydrase inhibition assay
The CA-catalyzed CO2 hydration activity was assayed on an Applied Photophysics stopped-flow instrument using phenol red (at a concentration of 0.2 mM) as a pH indicator with 20 mM Hepes (pH 7.5) as the buffer, 20 mM Na2SO4, and following the initial rates of the CA-catalyzed CO2 hydration reaction for a period of 10 − 100 s and working at the maximum absorbance of 557 nmCitation28–30. The CO2 concentrations ranged from 1.7 to 17 mM. For each inhibitor six traces of the initial 5 − 10% of the reaction have been used in order to determine the initial velocity. The uncatalyzed reaction rates were determined in the same manner and subtracted from the total observed rates. Stock solutions of inhibitor (0.1 mM) were prepared in distilled water, and dilutions up to 0.01 nM were prepared. Solutions containing the inhibitor and enzyme were preincubated for 15 min at room temperature prior to assay in order to allow the formation of the E − I complex. The inhibition constants were obtained as nonlinear least-squares protocols using PRISM 3Citation28–30 and are the mean from at least three different measurements. All hCAs were recombinant ones and were obtained in houseCitation28–30.
Toxicity evaluation
Inhibitors
Compounds 1–14 were either dissolved in Embryonic medium [5.0 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, and 0.1% w/v Methylene Blue (Sigma-Aldrich, Germany)] or in dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO) to prepare 100 mM stock solutions. Before the start of each experiment, the series of dilutions were made from the above stock in the embryonic medium.
Maintenance of zebrafish
The wild type adult zebrafish (AB strains) were maintained at 28.5 °C. 3–5 pairs of male and female fish were moved to breeding tanks overnightCitation31. Next morning, 1–2-h post fertilisation (hpf), embryos were collected in a sieve and rinsed with embryonic medium, and the collected embryos were maintained in an incubator at 28.5 °C overnightCitation31. The toxicity evaluation studies of the inhibitors were initiated with fish embryos 24-hpf. All the fish experiments were performed at the zebrafish core facility of Tampere University according to the protocol used in our laboratoryCitation32.
Ethical statement
The research unit at Tampere University has an established zebrafish core facility authorised by the National Animal Experiment Board (ESAVI/7975/04.10.05/2016). The experiments using developing zebrafish embryos were performed according to the Provincial Government of Eastern Finland Province Social and Health Department Tampere Regional Service Unit protocol # LSLH-2007–7254/Ym-23. Care was taken to ameliorate suffering by euthanizing the 5 dpf larvae by prolonged immersion in a petri dish containing an overdose of Tricaine (Sigma-Aldrich, St. Louis, MO) before fixing in buffered formaldehyde for histochemical analysis.
Determination of median lethal concentration 50 (LC50)
The LC50 values for all the MTC and DTC compounds were determined using 24-hpf embryos with 10–12 different concentrations for each compound. For every concentration of the inhibitor, we used 30 24-hpf embryosCitation31,Citation32. For each compound, the fish were exposed to different concentrations of the inhibitors rangeing from 5 μM to 2.5 mM. Dose response curve (DRC) was calculated using DRM of the DRC R packageCitation33. The control group larvae constituted an equal number of larvae not treated with any inhibitor compound and the larvae that were treated with 1% of DMSO. Toxicological evaluation studies were performed in 24-well plates (Corning V R Co-star V R cell culture plates). One or two 24-hpf embryos in were placed per well in 1 mL of embryonic medium containing a diluted inhibitor. A minimum of three sets of experiments were carried out for each inhibitor. Mortality of the larvae was checked every 24 h until 5 days after exposure to the inhibitors.
Phenotypic analysis of control and inhibitor treated larvae
After exposure to the inhibitors, we evaluated the effects of these inhibitors on the zebrafish larvae and analysed eight phenotypic parameters: (1) mortality, (2) hatching, (3) oedema, (4) swimming pattern, (5) yolk sack utilisation, (6) heartbeat, (7) body shape, and (8) swim bladder development. The images of the developing larvae were taken using a Lumar V1.12 microscope attached to a camera (Carl Zeiss MicroImaging GmbH, Göttingen, Germany). The images were analysed with AxioVision software versions 4.7 and 4.8 as described in our standard protocol for assessment of toxicity and safety of the chemical compoundsCitation32.
Swim pattern analysis
The swim pattern of the zebrafish larvae was studied after 5 days of exposure to these inhibitors. For the analyses of swim pattern, about 10–15 zebrafish larvae were placed in a 35 mm X 15 mm petri dish containing embryonic medium and the larvae were allowed to settle in the petri dish for 1 min. The movement of the zebrafish larvae was observed under the microscope for 1 min. The swim patterns were compared with the control group zebrafish larvae that were not treated with any inhibitor.
Histological studies
The histochemical analyses were done to assess the effect of inhibitors on the morphology of tissues of the larvae that were treated with different concentrations of inhibitors. The control group larvae were treated with embryonic medium alone or with 1% DMSO in embryonic medium. After 5 days of treatment the larvae were washed with phosphate buffered saline (PBS) and immersed in excess amounts of Tricaine to anaesthetize them. The larvae were transferred to a 1.5 mL microcentrifuge tube and fixed in buffered formaldehyde (4% formaldehyde solution, pH 6.9) in PBS for 3 h at room temperature or overnight at 4 °C. After the fixation, the larvae were transferred to 70% ethanol and stored at 4 °C before embedding in paraffin. The samples were then sectioned into 5 μm thin slices for the histochemical staining. The sections were deparaffinized in xylene, rehydrated in an alcohol series, and stained with Mayer's Haematoxylin and Eosin Y (both from Sigma-Aldrich). After dehydration, the slides were mounted with Entellan® Neu (Merck; Darmstadt, Germany). The slides containing the tissues were examined for morphological changes and photographed using a Nikon Microphot microscope (Nikon Microphot- FXA, Japan). All the procedures were carried out at room temperature unless stated otherwise.
Results and discussion
Chemistry
The main structural differences among the compounds tested in this study are related to the metal binding moieties of CAs. The compounds included both monothiocarbamates (MTCs) 1–7 and dithiocarbamates (DTCs) 8–14 ().
Figure 2. Chemical structures of MTCs 1-7 and DTCs 8-14.
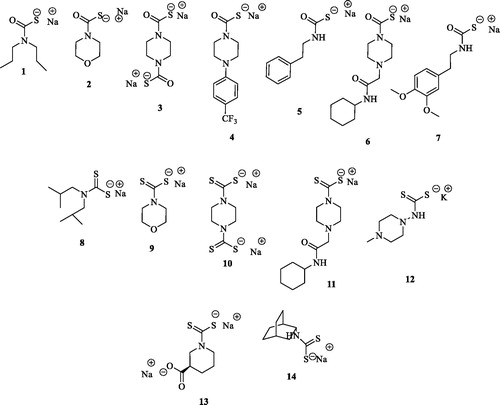
In order to efficiently explore the inhibition potencies of the MTC and DTC binding moieties on Mtb β-CA3, we decided to consider small size and lined-shaped scaffolds lacking any bulky substitution. Such an approach was also justified by the narrower size of the β CA’s cavities when compared to the α onesCitation34. As reported in almost all compounds responded to such a rule and the only exceptions were the 3-carboxy substituted piperazine 13 and the bicyclo[2.2.2]octan-2-amine 14.
CA inhibition
Compounds 1–14 were tested in vitro for their inhibitory properties against the Mtb β-CA3 and the data are reported in .
Table 1. Inhibition data of Mtb β-CA3 and human CA isoforms hCA I and II for compounds 1–14, determined by the stopped-flow CO2 hydrase assayCitation35, using acetazolamide (AAZ) as a standard drug.
The kinetic data reported in clearly showed that among the MTCs tested the derivative 4 was the most potent compound in inhibiting the Mtb β-CA3 with a KI value of 83.3 nM, followed by the phenylethyl 5 (KI 93.0 nM) and its 3,4-dimethoxyphenylethyl derivative 7 (KI 97.9 nM), and thus slightly more potent than the reference CAI AAZ (KI 104 nM). All the other remaining MTCs (i.e. 1–3 and 6) were high nanomolar range inhibitors of the β-CA3 with KI values spanning between 558.6 and 812.0 nM. Interestingly MTCs 4, 5 and 7 were ineffective inhibitors of the hCA I (KIs >2000 for compounds 4 and 5, 891 nM for 7 respectively). Conversely, compounds 4, 5 and 7 showed good inhibition data on hCA II being up to 3.7 fold more potent (KIs of 43.6, 43.7 and 26.7 nM, respectively). An analogous kinetic trend was observed for the remaining MTCs 1, 3 and 6 which were quite effective inhibitors of the hCA II over the hCA I isoform. The only exception is represented by the MTC based morpholine 2 which showed high nanomolar KI value on the hCA I (KI 569 nM) and no effects on the hCA II isoform (KI >2000).
As for the isosteric DTC derivatives 8–14 a rather different kinetic profile was observed. For instance, the morpholine DTC 9 was the most potent inhibitor within the series against Mtb β-CA3 (KI of 2.4 nM) and quite interestingly it was 311.5 fold more potent when compared to its MTC analogous compound 2 (see ). A similar kinetic trend was reported also for the bis-piperazine derivative DTC 10 in comparison to its MTC counterpart 3 with the former being 101.5 fold more potent on the β-CA3 (KIs of 8.0 and 812.0 nM, respectively). An opposite kinetic trend was reported for the DTC 11 which was ineffective on the β-CA3, whereas its MTC derivative 6 was a high nanomolar inhibitor (KIs of >2000 and 780.7 nM, respectively). Among the DTCs reported in this study, the N-methylpiperazine 12 was the second most potent inhibitor against the β-CA3 just after the morpholine derivative 9 (KI of 2.6 and 2.4 nM, respectively). The structural differences between 9 and 12 (i.e. an oxygen and an N-methyl moiety at 4-position of the scaffold, respectively) did not affect the inhibition potencies against the β-CA3, whereas they proved crucial for the inhibition of the hCAs I and II. As reported in the morpholine derivative 9 was a sub-nanomolar inhibitor of the hCAs reported (KIs of 0.88 and 0.95 nM for the hCA I and II, respectively), whereas the N-methyl piperazine 12 was a medium potency inhibitor (KIs of 33.5 and 33.5 nM, respectively), that makes the latter more selective against the β-CA3 isoform. Interestingly, the substitution of the alkyl ring in 9 or 12 with a bis-alkyl chain, as in compound 8, or its modification to produce 13 and 14, resulted in detrimental reductions for the inhibition potency against the β-CA3 (see ).
Evaluation of safety and toxicity
Determination of inhibitor LC50 concentrations
The lethal concentrations of the MTC and DTC 1–14 inhibitors were tested on developing zebrafish embryos. The lowest concentration that caused death of half of the embryos was reported for the DTC 8 (i.e. LD50 2 μM), which was therefore considered highly toxic and not suitable for further testing in our studies. The LC50 values of all the compounds are shown in .
Table 2. LC50 of the CAIs 1–14.
Phenotypic analyses of zebrafish larvae treated with inhibitor compounds
To assess the toxic effects of the inhibitors on 5-day-old zebrafish during development after 4-day exposure to the compounds, we analysed seven observable phenotypic parameters () using a stereo microscope and recorded the observations for each group. The zebrafish treated with CAIs were compared with the control groups not treated with any compound or with 1% DMSO. The maximum concentration of each compound that does not induce any phenotypic changes in the larvae at 5 days of exposure was considered as safe () and can be used for in vivo inhibition of M. marinum in zebrafish larvaeCitation2. The images in show representative larvae exposed to MTCs and DTCs in which no apparent phenotypic defects were observed except the compound 8 that showed defects in the development of swim bladder (, arrow). DTC 12 that inhibited Mtb β-CAs in vitro efficiently in our earlier studiesCitation36, showed similar results and was used for in vivo inhibition of M. marinum in zebrafish studiesCitation1.
Figure 3. Images of zebrafish larvae treated with different inhibitors. The identification number of each compound is in parenthesis. The images of the zebrafish larvae after 5 days of exposure to inhibitors that are considered safe for in vivo use to inhibit M. marinum growth in larvaeCitation2. The concentrations of the MTCs and DTCs shown here are the highest ones generally not inducing phenotypic defects at the end of 5 days of exposure. For compound 8, even the lowest concentration used caused the absence of swim bladder (arrow), suggesting that this compound is not suitable for further characterizationCitation2.
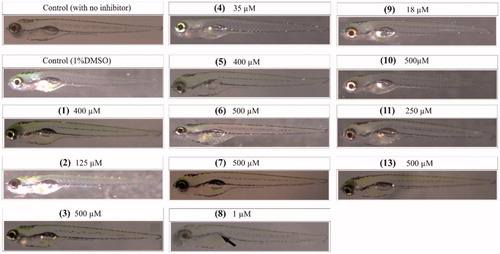
Table 3. Effect of inhibitors on phenotypic parameters of the larvae 5 days of after exposureTable Footnotea,Table Footnoteb.
Further analyses of the embryos to assess the toxicity of compounds at the concentrations that are considered as safe () on the observable phenotypic parameters showed that some of the compounds had no significant effects on any of the parameters assessed (). The compounds that were considered toxic, showed different level of toxicity by inducing high mortality, problem with hatching, oedema, heartbeat, yolk sac utilisation, body shape, and swim bladder development () as shown in .
Histochemical analysis
We analysed the sections of the zebrafish that were exposed to the inhibitors which did not have any significant effect on the phenotypic parameters () of the larvae at the end of 5 days of exposure to the inhibitors. The histologically stained sections of the inhibitor-treated larvae were compared with the stained sections of the control group larvae. None of the inhibitors showed any morphological changes of tissues of the zebrafish larvae exposed to the MTCs or DTCs (data not shown). The results of the histological examination suggest that these inhibitors cause no damage to the internal tissues at the concentrations tested and can be used for further characterisation.
Swim pattern analysis of larvae exposed to the inhibitors
Zebrafish embryos are easily affected by chemical compounds compared to adult zebrafish or other animal models and are hence suitable model organisms for assessing the subtle toxic effects of chemicalsCitation32,Citation37. In this study, we further assessed the subtle toxic effects of the inhibitors by analysing the swim patterns of the larvae during exposure to the inhibitors at concentration that are considered as safe shown in . The swim pattern analysis showed that 11 out of 14 compounds showed no abnormal or ataxic movement of the larvae that were exposed to the inhibitors (data now shown). Therefore, most of the compounds that efficiently inhibit Mtb β-CA3 show no or minimal toxicity at relevant concentrations and are thus safe for further characterisation in vivoCitation2,Citation32 ().
Table 4. The compounds that inhibit Mtb β-CA3 efficiently and show minimal toxicity.
Conclusions
In this study, we investigated a series of 14 CAIs belonging to the MTC and DTC zinc binding moieties possessing a variety of scaffolds and intended as inhibitors of the β-CA3 from M. tuberculosis. All compounds were investigated in vitro for their inhibition potencies against Mtb β-CA3 and compared to the human hCA I and II. The Mtb β-CA3 was efficiently inhibited by 11 of these derivatives with KIs in the range of 2.4–812 nM. We evaluated these compounds for their toxic effects on 1–5 dpf zebrafish larvae. The toxicological studies showed that the compounds 1, 3, 5, 7, and 10 exhibited minimal toxicity and were considered as safe for further characterisation in vivo. The results of the in vitro inhibition and toxicological evaluation studies showed that 5 compounds can be used for in vivo inhibition of the M. marinum growth in zebrafish larvae for further preclinical characterisation.
Acknowledgements
We thank Aulikki Lehmus and Marianne Kuuslahti for the histochemical analyses of the zebrafish samples. We also thank Leena Mäkinen and Hannaleena Piippo for the assistance with zebrafish embryos for the experiments.
Disclosure statement
The authors declare no conflict of interests.
Additional information
Funding
References
- WHO. Global Tuberculosis Report. 2018; ISBN 978-92-4-156564-6:1–277.
- Aspatwar A, Hammaren M, Koskinen S, et al. beta-CA-specific inhibitor dithiocarbamate Fc14-584B: a novel antimycobacterial agent with potential to treat drug-resistant tuberculosis. J Enzyme Inhib Med Chem 2017;32:832–40.
- Aspatwar A, Winum JY, Carta F, et al. Carbonic anhydrase inhibitors as novel drugs against Mycobacterial beta-Carbonic anhydrases: an update on in vitro and in vivo studies. Molecules 2018;23:291.
- Supuran CT. Carbonic anhydrases. Bioorg Med Chem 2013;21:1377–8.
- Supuran CT. Structure and function of carbonic anhydrases. Biochem J 2016;473:2023–32.
- Supuran CT. Carbonic anhydrases: novel therapeutic applications for inhibitors and activators. Nat Rev Drug Discov 2008;7:168–81.
- Supuran CT, Capasso C. An overview of the bacterial carbonic anhydrases. Metabolites 2017;7:56.
- Carta F, Maresca A, Covarrubias AS, et al. Carbonic anhydrase inhibitors. Characterization and inhibition studies of the most active beta-carbonic anhydrase from Mycobacterium tuberculosis Rv3588c. Bioorg Med Chem Lett 2009;19:6649–54.
- Sassetti CM, Boyd DH, Rubin EJ. Genes required for mycobacterial growth defined by high density mutagenesis. Mol Microbiol 2003;48:77–84.
- Sassetti CM, Rubin EJ. Genetic requirements for mycobacterial survival during infection. Proc Natl Acad Sci U S A 2003;100:12989–94.
- Rose SJ, Bermudez LE. Identification of bicarbonate as a trigger and genes involved with extracellular DNA export in mycobacterial biofilms. mBio 2016;7:e01597–e01616.
- Johnson BK, Colvin CJ, Needle DB, et al. The carbonic anhydrase inhibitor ethoxzolamide inhibits the Mycobacterium tuberculosis PhoPR Regulon and Esx-1 secretion and attenuates virulence. Antimicrobial Agents Chemother 2015;59:4436–45.
- Suarez Covarrubias A, Larsson AM, Hogbom M, et al. Structure and function of carbonic anhydrases from Mycobacterium tuberculosis. J Biol Chem 2005;280:18782–9.
- Covarrubias AS, Bergfors T, Jones TA, Högbom M. Structural mechanics of the pH-dependent activity of beta-carbonic anhydrase from Mycobacterium tuberculosis. J Biol Chem 2006;281:4993–9.
- Minakuchi T, Nishimori I, Vullo D, et al. Molecular cloning characterization and inhibition studies of the Rv1284 beta-carbonic anhydrase from Mycobacterium tuberculosis with sulfonamides and a sulfamate. J Med Chem 2009; 52:2226–32.
- Nishimori I, Minakuchi T, Vullo D, et al. Carbonic anhydrase inhibitors. Cloning characterization and inhibition studies of a new beta-carbonic anhydrase from Mycobacterium tuberculosis. J Med Chem 2009;52:3116–20.
- Pacchiano F, Carta F, Vullo D, et al. Inhibition of beta-carbonic anhydrases with ureido-substituted benzenesulfonamides. Bioorg Med Chem Lett 2011;21:102–5. 201
- Maresca A, Carta F, Vullo D, et al. Carbonic anhydrase inhibitors. Inhibition of the Rv1284 and Rv3273 beta-carbonic anhydrases from Mycobacterium tuberculosis with diazenylbenzenesulfonamides. Bioorg Med Chem Lett 2009;19:4929–32.
- Monti SM, Maresca A, Viparelli F, et al. Dithiocarbamates are strong inhibitors of the beta-class fungal carbonic anhydrases from Cryptococcus neoformans Candida albicans and Candida glabrata. Bioorg Med Chem Lett 2012;22:859–62.
- Nocentini A, Vullo D, Del Prete S, et al. Inhibition of the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa with monothiocarbamates. J Enzyme Inhib Med Chem 2017;32:1064–70.
- Maresca A, Carta F, Vullo D, Supuran CT. Dithiocarbamates strongly inhibit the beta-class carbonic anhydrases from Mycobacterium tuberculosis. J Enzyme Inhib Med Chem 2013;28:407–11.
- Vullo D, Del Prete S, Nocentini A, et al. Dithiocarbamates effectively inhibit the beta-carbonic anhydrase from the dandruff-producing fungus Malassezia globosa. Bioorg Med Chem 2017;25:1260–5.
- Bua S, Bozdag M, Del Prete S, et al. Mono- and di-thiocarbamate inhibition studies of the delta-carbonic anhydrase TweCAdelta from the marine diatom Thalassiosira weissflogii. J Enzyme Inhib Med Chem 2018;33:707–13.
- Carta F, Aggarwal M, Maresca A, et al. Dithiocarbamates: a new class of carbonic anhydrase inhibitors. Crystallographic and kinetic investigations. Chem Commun 2012;48:1868–70.
- Carta F, Aggarwal M, Maresca A, et al. Dithiocarbamates strongly inhibit carbonic anhydrases and show antiglaucoma action in vivo. J Med Chem 2012;55:1721–30.
- Vullo D, Durante M, Di Leva FS, et al. Monothiocarbamates strongly inhibit carbonic anhydrases in vitro and possess intraocular pressure lowering activity in an animal model of glaucoma. J Med Chem 2016;59:5857–67.
- Bozdag M, Carta F, Vullo D, et al. Synthesis of a new series of dithiocarbamates with effective human carbonic anhydrase inhibitory activity and antiglaucoma action. Bioorg Med Chem 2015;23:2368–76.
- Carta F, Vullo D, Maresca A, et al. New chemotypes acting as isozyme-selective carbonic anhydrase inhibitors with low affinity for the offtarget cytosolic isoform II. Bioorg Med Chem Lett 2012;22:2182–5.
- Carta F, Vullo D, Maresca A, et al. Mono-/dihydroxybenzoic acid esters and phenol pyridinium derivatives as inhibitors of the mammalian carbonic anhydrase isoforms I II VII IX XII and XIV. Bioorg Med Chem 2013;21:564–9.
- Angeli A, Ferraroni M, Nocentini A, et al. Polypharmacology of epacadostat: a potent and selective inhibitor of the tumor associated carbonic anhydrases IX and XII. Chem Commun 2019;55:5720–3.
- Gourmelon A, Delrue N. Validation in support of internationally harmonised OECD test guidelines for assessing the safety of chemicals. Adv Exp Med Biol 2016;856:9–32.
- Aspatwar A, Hammaren MM, Parikka M, Parkkila S. Rapid evaluation of toxicity of chemical compounds using zebrafish embryos. J Vis Exp 2019;150:e59315.
- Ritz C, Baty F, Streibig JC, Gerhard D. Dose-Response analysis using R. PloS One 2015;10:e0146021.
- De Simone G, Alterio V, Supuran CT. Exploiting the hydrophobic and hydrophilic binding sites for designing carbonic anhydrase inhibitors. Expert Opin Drug Dis 2013;8:793–810.
- Khalifah RG. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J Biol Chem 1971;246:2561–73.
- Maresca A, Vullo D, Scozzafava A, et al. Inhibition of the beta-class carbonic anhydrases from Mycobacterium tuberculosis with carboxylic acids. J Enzyme Inhib Med Chem 2013;28:392–6.
- Aspatwar A, Becker HM, Parvathaneni NK, et al. Nitroimidazole-based inhibitors DTP338 and DTP348 are safe for zebrafish embryos and efficiently inhibit the activity of human CA IX in Xenopus oocytes. J Enzyme Inhib Med Chem 2018;33:1064–73.