
ABSTRACT
Across the circumpolar north, the degradation of permafrost has resulted in an increase in the extent of thermokarst landforms. Within thermally disturbed ecosystems, climatically driven changes to hydrology and temperature regimes have the potential to modify nutrient cycling processes. To assess the impacts of changing moisture and thermal conditions on the mineralization of nutrients in the Hudson Bay Lowlands, subarctic Canada, soil cores were extracted along gradients of moisture and topographic position (peat plateaus, sedge lawns, channel fens, and thermokarst collapse scars). Soil subsamples were then subjected to a factorial design of temperature (4°C, 12°C, and 20°C) and moisture treatments (saturated, field moist, and air dried) in oxic conditions for three weeks. Nitrogen transformation rates were highly variable across landscape units (ranging from −1 to 96 μg N-NO3− and from −53 to 73 μg N-NH4+ g−1 dry soil for the incubation period). Shoreline collapse scar material showed the greatest potential for nitrification, with rates two orders of magnitude higher than other landscape positions, under warm (20°C) and saturated conditions. This work shows the potential of increased plant-available nitrate for rapid vegetative colonization of thermokarst collapse scars, and provides novel insight into nutrient cycling processes in permafrost peatland landscapes.
Introduction
Many northern cold-region landscapes are favorable environments for the accumulation of peat (Kuhry and Turunen Citation2006), where the decomposition rate of organic matter is retarded by low mean annual soil temperatures and prolonged anoxia due to a perched water table overlying relatively impermeable permafrost (Woo Citation2012). The insulating properties of peat are favorable for the development and preservation of underlying ground frost (Vitt, Halsey, and Zoltai Citation1994; Woo and Young Citation2003). Climatic changes, and in particular warming global temperatures, are projected to be amplified at northern latitudes where permafrost peatland coverage is currently widespread (Callaghan et al. Citation2010; Cohen et al. Citation2014; Kaufman et al. Citation2009). Long-term observational evidence has shown that permafrost has thawed extensively in the past century (Jorgenson, Shur, and Pullman Citation2006; Payette et al. Citation2004) and is projected to continue to do so (Lawrence, Slater, and Swenson Citation2012; Schaefer et al. Citation2011; Schuur et al. Citation2013). Recent permafrost degradation has been shown to change both hydrologic pathways (Connon et al. Citation2014; Walvoord and Kurylyk Citation2016) as well as carbon (C) cycling regimes (Schuur et al. Citation2008). However, the potential for changes to nutrient cycling from climatic changes and permafrost thaw are less constrained (Vonk et al. Citation2015), and have important implications for further modifying the ecological and C storage functions of northern ecosystems.
In northern peatlands, the internal cycling of nutrients stored in organic matter often represents a much larger fraction of an annual nutrient budget than external inputs and total export (Malmer and Nihlgård Citation1980). The supply of nutrients plays a large role in C cycling and peat accumulation rates, because peat-forming terrestrial biomass and aquatic productivity are typically chiefly limited by nitrogen (N) and phosphorus (P) (Cargill and Jefferies Citation1984; Shaver and Chapin Citation1980; Symons, Arnott, and Sweetman 2012). However, the low annual average soil temperatures in northern peatlands limit the rates at which biogeochemical processes occur (Stark Citation2007). Generally, microbial decomposition remains the rate-limiting factor in nutrient cycling (Andersen et al. Citation2013). Thus, soil and air temperatures are often the limiting factors dictating the rate of microbial decomposition of soil organic matter in cold regions (Jones et al. Citation2009; Uhlířová, Šantrůčková, and Davidov Citation2007; Wallenstein, McMahon, and Schimel Citation2009). It is predicted that enhanced soil microbial activity from both increased temperatures (increased rate) as well as a longer snow-free season (increased active period) will result in the increased liberation of N and P in bioavailable forms from organic matter (Brown and Braaten Citation1998; Weintraub and Schimel Citation2005). Experimental warming of boreal and arctic soils has increased soil inorganic N and P pools (Natali et al. Citation2011; Schimel, Bilbrough, and Welker Citation2004) through increased ammonification, nitrification, and P mineralization (Rustad et al. Citation2001). Temperature projections for northern regions show increases in temperature between 1°C and 10°C, varying across space and seasons; however, changes to precipitation regimes are less certain and more variable across space (Bintanja and Andry Citation2017). As such, some regions may become drier and others may become wetter. Moreover, with permafrost thaw resulting in geomorphic disturbance, previously dry upland areas may become ephemerally or permanently saturated (Payette et al. Citation2004). Although the individual effects of changes in moisture or temperature on nutrient transformations have been examined, less is known about the combined effects of simultaneous moisture and temperature manipulations on net mineralization rates.
Typically, the unsaturated zone maintains oxic conditions dominated by efficient aerobic processes, resulting in high decomposition rates (Golovchenko, Tikhonova, and Zvyagintsev Citation2007). With a drawdown in the water table, an increase in unsaturated zone depth (Macrae et al. Citation2013; Roulet et al. Citation1992), and the associated additional oxygen supply in drier soils, there will be an increase in available plant nutrients resulting from products of oxidation of organic matter. Loss of soil structure through permafrost and ground-ice thaw may lead to the development of thermokarst collapse scars, which can facilitate the transport of nutrients and solutes from areas that would otherwise be hydrologically inaccessible (Jorgenson and Osterkamp Citation2005; Lafrenière and Lamoureux Citation2013). These geomorphic disturbances have the potential to expose deeper portions of the soil profile (typically inundated and suboxic/anoxic) to oxic conditions when exposed to the surface. Because peat is generally a poor thermal conductor, surface peat is more responsive to air temperatures while deeper peat is less sensitive. However, with thermokarst activity, deeper portions of the soil profile may warm and dry rapidly when exposed to the surface climate through structural disturbance. These hydrologic changes may alter nutrient transformations in a manner that depends on the consistency and duration of new moisture regimes. For example, repeated drying-wetting cycles enhance phosphatase activity and may therefore result in the release of phosphate (Song, Zoh, and Kang Citation2007). Alternatively, sustained drying followed by rewetting can decrease phosphorus supply as a result of increased metal complexing (Richardson Citation1985). In the case of nitrogen, sustained drying as well as re-wetting cycles can result in large increases to ammonification and denitrification rates, respectively (Venterink et al. Citation2002). Sustained wetting can also lead to elevated N and P mineralization by increasing heat transfer through conduction and elevated temperatures (Lipson et al. Citation2012). Thus, while the effects of changing moisture conditions on nutrient cycling are complex, it is necessary to consider the effects of temperature and moisture regime changes in tandem rather than in isolation from one another because of their natural linkages.
The rate and climatic controls of nutrient mineralization through organic-matter decomposition are strongly related to the quality of the litter parent material and recalcitrance, and consequently are not uniform through heterogeneous patterned landscapes (Eppinga et al. Citation2010; van Breemen and Finzi Citation1998). The partitioning of peatland ecosystems into landscape units is a common practice used to understand how biogeochemical dynamics vary with topography and hydrological responses (Allan, Roulet, and Hill Citation1993; Quinton, Hayashi, and Pietroniro Citation2003), and can be used to study how different elements on the landscape, and their differing litter parent material, contribute to whole-system nutrient cycling. Thus, it is important to determine how peat biogeochemical properties vary between depths and landscape unit position and, further, to determine which properties are the key drivers in nutrient mineralization responses to temperature and moisture conditions. To address these needs, the objectives of this study were to: (1) assess the variability in nutrient pools (total and extractable) between different landscape units and through depth in a thermokarst-impacted permafrost bog; (2) experimentally determine the individual and combined effects of temperature and moisture on the net mineralization rates of N and P; and (3) determine if and how these controls differ as a function of litter quality.
Materials and methods
Experimental design overview
Four replicate peat cores were collected and sectioned into three depths (0–10 cm, 10–20 cm, > 20 cm) at each of four landscape units (collapse scar, channel fen, sedge lawn, peat plateau) in a subarctic peatland catchment. Using a factorial design, peat subsamples were subjected to three temperature (4°C, 12°C, and 20°C) and three moisture (air dry, field moist, saturated) treatments and were incubated for three weeks in a controlled laboratory setting to determine the net mineralization rates of NO3−, NH4+, and PO43- under the different conditions.
Study site and soil sampling
The study site is a peat plateau bog near Churchill, Manitoba (58°48ʹ N, 94°09ʹ W) in the catchment of a lake in a recent thermokarst shoreline collapse expansion state. The climate of the region is strongly influenced by its proximity to Hudson Bay (Rouse Citation1991) and is underlain by near-continuous permafrost with an active-layer depth ranging from 40 cm to 1 m (Dyke and Sladen Citation2010). Four landscape units were selected in the catchment that were representative of the variability in topographic, vegetative, and moisture conditions in the region, following the classification scheme and approach of Quinton, Hayashi, and Pietroniro (Citation2003; ). Fen channels (FC) are characterized by near-surface saturation and may develop in frost wedge polygon cracks or as diffuse pathways over land (Woo and Young Citation2006). Fen channels are dominated by Carex aquatilis and Plagiomnium spp. Sedge lawns (SL) are intermediary zones between upland frost-driven landforms and low-lying FCs. Given the transitional nature of SL, a wide variety of vascular plants (e.g., Vaccinium, Rhubus), sedges (Carex mitis), mosses (primarily Dichranum with some Sphagnum), and lichens (Cladina, Cetraria spp.) may be present, in addition to the presence of sparse tree islands (Picea mariana, Picea glauca, Larix laricina; Mamet and Kershaw Citation2013). Peat plateaus (PP) are dry upland areas developed via frost action between peat and underlying substrates (Sannel and Kuhry Citation2009). In contrast with the dominance of Carex spp. in FC and SL landforms, Carex are absent in PP and vegetation communities are instead lichen-heath communities (e.g., Empetrum, Rhubus, Dichranum, Cladina, Cetraria, Vaccinium, Ledum, with some Salix). The collapse scar (CS) is a thermokarst feature examined in this work, and is a collapsed upland area encompassing the shoreline of the small lake resulting from inundation and enhanced heat conduction leading to the failure of soil structure through permafrost thaw. In this region, thermokarst features often develop adjacent to existing pond edges and are either bare of any living vegetation or colonized by Carex spp., and are characterized by the presence of dying trees (Payette and Delwaide Citation2000). In this landscape, catchment pore water is generally higher in dissolved nutrient concentrations than surface water, with most dissolved inorganic N occurring as NH4+ (< 0.01–0.57 mg NO3–N L−1, < 0.01–4.17 mg NH4+-N L−1, < 0.01–1.12 mg soluble reactive P L−1 in groundwater; < 0.01–0.50 mg NO3–N L−1, < 0.01–1.12 mg NH4+-N L−1, < 0.01–0.02 mg soluble reactive P L−1 in surface water; Morison et al. Citation2017).
Figure 1. Pictures showing four major landscape units in this study: (a) peat plateau, (b) sedge lawn, (c) channel fen, and (d) thermokarst collapse scar

Soil samples were collected September 19–21, 2015, to capture a period when the seasonal ground frost had fully thawed and to facilitate the collection of the entire active layer. A 10.16 cm interior diameter PVC core was inserted from the surface until frozen ground was encountered at four sampling sites within each landscape unit (sixteen cores in total, four sites for each of four landscape units). Cores were packed on ice and stored at −4°C to simulate winter freezing conditions until being gradually thawed over seven days at 4°C, following Kelley et al. (Citation2012). In the laboratory, each core was then sectioned into three depths: 0–10 cm, 10–20 cm, and > 20 cm, with the total depth varying between cores—the shallowest core being 45 cm and the deepest being 75 cm. The four spatial samples for each landscape unit at each depth were composited (three depths at four landscape units) following Chambers, Marshall, and Danehy (Citation2004) and DeBusk and Reddy (Citation1998) to maintain representatives from multiple sites within each landscape unit, while providing an estimation of central tendency of the population (Boone et al. Citation1999; Osborne and DeLaune Citation2013). All visible roots were manually removed from samples, and all samples were manually blended in sealed plastic bags until visibly homogenous. This compositing technique, although a form of pseudoreplication (Davies and Gray Citation2015), has been employed previously in laboratory incubation studies measuring nutrient mineralization (Austin and Vitousek Citation1998; Long et al. Citation2017; Whalen, Chang, and Olson Citation2001) to explore differences in treatments, rather than within-site differences. While within-landscape unit heterogeneity in nutrient mineralization rates requires further investigation to fully understand ecosystem nutrient cycling, this was not the primary aim of this study.
Extraction and incubation procedures
In the laboratory, 5 g (field moist mass) of each peat sample was extracted in 50 ml of either 2 M KCl (for NO3− and NH4+) or deionized water (for water-extractable phosphorus) by shaking at 350 rpm for one hour on an orbital shaker table and then gravity filtering through 2.5 μm filter paper (Whatman No.4). Analytical triplicates of each of the twelve soil groups were immediately extracted to determine nutrient pools prior to incubation.
Four analytical replicates of 5 g from each soil group were then incubated aerobically in darkness in a sterile plastic cup in one of nine different treatment groups (three temperatures × three moisture conditions) for a 21 d period (n = 4 analytical replicates × 12 soil groups × 3 temperatures × 3 moisture treatments = 432 samples incubated). Temperature levels were set at 4°C, 12°C, and 20°C in separate growth chambers with HOBO temperature loggers deployed to monitor deviations from the target temperature. Three moisture conditions were attained in each soil group by (1) air-drying the soil for 24 h in a ventilated area with paired sealed control samples exposed to the same room temperature for preincubation extraction comparisons, (2) maintaining field moisture conditions, and (3) wetting to saturation with deionized water. Temperature and moisture contents remained within the targeted values throughout the incubation period () and fell within the range of values recorded through one season (May–September 2014) of in situ weekly monitoring of surface (0–5 cm) temperature and moisture values at the study site where the cores were collected. The gravimetric moisture content of each soil group at each of the three moisture conditions was determined by oven-drying at 95°C for 24 h. During the 21 d incubation period, samples were weighed every 48 h and were re-wetted with deionized water to their initial weight to replace the moisture lost to evaporation, with total wet sample mass change never exceeding 10 percent during a 48 h period. Following incubation, samples were extracted in the same manner as the paired samples prior to incubation.
Table 1. Targeted and measured values of moisture and temperature conditions, reported as mean (standard deviation)
At the University of Waterloo Biogeochemistry Laboratory, extracted samples were analyzed for nitrate, ammonium, and orthophosphate concentrations using colorimetric methods (Bran-Luebbe AutoAnalyzer III, Seal Analytical: Methods G-102–93 [NH4+-N]; G-103–93 [SRP-P], G-109–94 [NO3–N]). Results for NH4+, NO3−, and soluble reactive P have a detection limit of 0.001 mg N L−1, or 0.001 mg P L−1. No samples were below the detection limit during the analysis of ammonium and orthophosphate. During nitrate analysis, when a sample was determined to have a concentration below the detection limit (0.001 mg N-NO3− L−1, which occurred in 14 percent of samples) the concentration was assumed to be half of the detection limit (0.0005 mg N-NO3− L−1) for the purposes of determining net nitrification rates that did not represent a substantial error in the calculation. The mineralization rate was calculated as the difference between the concentration of incubated sample extract, normalized per gram of dry soil, and the concentration of initial extract, normalized per gram of dry soil. Extractable pools of N and P in soil are expressed as mass of N or P per mass of dry soil, while net mineralization rates are expressed as mass of N or P per mass of dry soil per incubation day.
Peat core properties
Subsamples from each soil group were analyzed for organic content using loss on ignition (LOI), where homogenized samples were heated to 550°C for 4 h following a drying stage of heating to 95°C for 24 h, following Heiri, Lotter, and Lemcke (Citation2001). Additional subsamples were freeze-dried, ball-milled, and analyzed for bulk composition of total C and N at the University of Waterloo Environmental Isotope Laboratory through a 4010 Elemental Analyzer (Costech Instruments) and for total P through the digestion method of Parkinson and Allen (Citation1975) and subsequent analysis of digested P as phosphate through standard colorimetric methods at the University of Waterloo Biogeochemistry Laboratory (Bran-Luebbe AutoAnalyzer III, Seal Analytical: Method G-103–93). Using 5 percent duplicate analytical replicates throughout all steps of the method, data for total C and N both show analytical precision of < 1 percent (average of 0.2 and 0.3 percent, respectively), while total P data showed an average analytical precision of 1.8 percent.
Statistical techniques
To address the first objective of this study, we ran a one-way analysis of variance (ANOVA; F statistic) of all phosphate, nitrate, and ammonium mineralization-rate data using landscape unit as the fixed factor. To address the second objective, we ran a two-way ANOVA of all net mineralization-rate data using temperature (4°C, 12°C, and 20°C) and moisture (air dry, field moist, saturated) treatments as fixed factors. In each case, data were tested for normality (with a Shapiro-Wilk test) and homoscedasticity. If these assumptions were not met, a nonparametric two-way ANOVA on ranks (Scheirer-Ray-Hare test; H statistic) was used instead. Following each ANOVA, provided there was a significant effect of an individual factor without a significant interaction term between factors, post hoc comparisons were assessed with a t test (if data were normally distributed) or Mann-Whitney U test (if data were not normal), with a Bonferroni adjustment for the number of comparisons. To address the third objective of this study and determine the control of initial extractable nutrient pools and total nutrient stoichiometry (totals and ratios of N, P, C) on mineralization rates, Pearson product moment correlations were performed between mineralization rates and these soil properties.
Due to a contamination error in one subsample in the experimental design (peat plateau, 0–10 cm depth subsample for analysis of water-extractable phosphorus assigned to air dry and 20°C treatments), this sample was removed from all analyses. All statistical analyses were conducted with R software (version 3.3.2, R Development Core Team Citation2016).
Results
Peat core properties
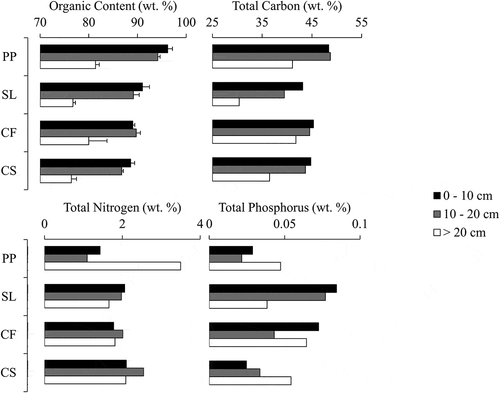
Organic content (as LOI550°C) generally decreased with depth (from 0 cm to > 20 cm) in all landscape units from approximately 90 percent in surface peat to 78 percent in material deeper than 20 cm (). Organic content was greatest in PP, least in CS, and intermediate in SL and CF at all depths. Total C generally decreased with depth (from 0 cm to > 20 cm) in all landscape units, and was least in SL and greatest in PP. Total N varied most with depth in PP (from 1.1 percent to 3.5 percent) but was more consistent through depth in SL (mean of 1.9 ± standard deviation of 0.2 percent), CF (1.9 ± 0.1), and CS (2.3 ± 0.3). In shallower material (0–10 cm and 10–20 cm), SL and CF had greater total P than PP and CS but all landscape units had similar total P in deep material. C:N ratios fluctuated most with depth in PP (ranging from 12 to 44), and were less variable across other landscape units (all other values ranging from 17 to 26). C:N ratios were greatest in the surface peat (0–10 cm) in all other landscape units except PP (). N:P ratios tended to be greatest in CS in surface samples (0–20 cm), driven by low P content, while neither N:P or C:P ratios appear to have a consistent pattern through depth across landscape units.
Table 2. Stoichiometry of major nutrients in different landscape units and depths
Figure 2. Peat properties through depth in each of the four landscape units: loss-on-ignition as a proxy for organic content, percent carbon by mass, percent nitrogen by mass, and percent phosphorus by mass in sample. Error bars for loss-on-ignition data are the standard deviation of three replicate samples
Extractable nutrient pools
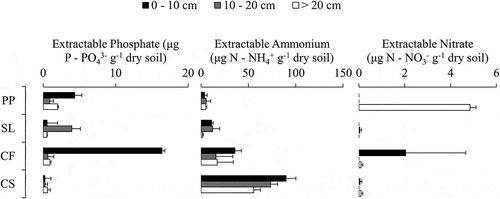
In the preincubated extracted samples, the inorganic N pool was dominated by NH4+ (). Across all soil groups an average of 95 percent of total inorganic N (TIN; NH4+ + NO3−) was NH4+ with two major exceptions, surface peat (0–10 cm) in CF and deep material (> 20 cm) in PP, which had appreciable nitrate concentrations. Other samples had little detectable NO3− (< 0.1 μg N − NO3− g−1 dry soil). Extractable NH4+ pools tended to decrease with depth and increase along a moisture gradient in landscape units (lowest in PP, greatest in CS). Extractable P pools were greatest in surface peat (0–10 cm) driven by high concentrations in CF (16.4 ± 8.2 μg P − PO43- g−1 dry soil) and PP (4.4 ± 1.1 μg P − PO43- g−1 dry soil).
Figure 3. Extractable nutrient pools of soluble reactive phosphorus, ammonium, and nitrate for each landscape unit through depth. Error bars represent the standard deviation of triplicate extractions
Net mineralization rates
Ammonium
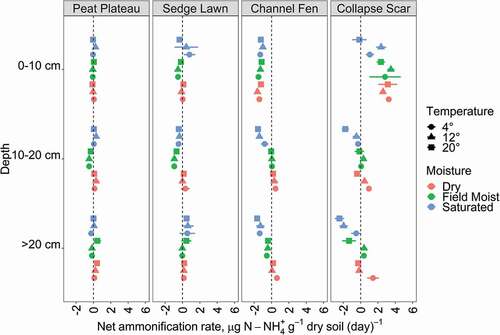
Net ammonification varied by landscape position and depth where CF and CS had much more variable responses to treatments than the drier areas (PP and SL; ). With the exception of the surface layer in the CS, the median net ammonification rates across all treatments were zero or slightly below zero. In most but not all cases across the treatments, saturation favored negative rates, whereas dry conditions favored positive rates; however, this was most apparent in the wetter areas (CS, CF) and very subtle in the dry (PP, SL) areas. A notable exception to this was the surface layer in the CF, which showed the reverse pattern (i.e., net ammonification under saturated conditions). Temperature did not change these overall relationships, but instead appeared to amplify them in these two landscape units, although the responses to temperature were variable within the moisture treatments. For example, in the wet areas (CF, CS), the negative ammonification observed under saturated conditions was greatest in warm peat and dampened in cold peat. In contrast, the positive net ammonification observed under the dry treatment was greatest for cold peat and dampened in warm peat. These differences with temperature were not observed for the drier (PP, SL) areas. In general, CS material displayed the widest response (both negative and positive net ammonification) through depth and across treatments compared to nondisturbed peat. The CS displayed the greatest range of response to temperature and moisture treatments within each depth category. There was a significant effect of moisture (H2,96 = 7.664, p < 0.001) with air-drying treatments, resulting in significantly greater net ammonification than saturated (p < 0.01) or field-moist (p < 0.001) samples, with no significant effect of temperature or interaction between temperature and moisture effects. There was also a significant effect of landscape unit (H3,96 = 6.445, p < 0.001) with a significantly lesser (negative rate) median in CF than all other landscape units (p < 0.05 in each case), and no significant differences among PP, SL, and CS ammonification rates.
Figure 4. Net ammonification rates at each landscape unit (panels) and depth (y-axis), showing the effects of both temperature (shape) and moisture (color) treatments, with each point and bars showing the mean and standard deviation of four experimental replicates
Nitrate
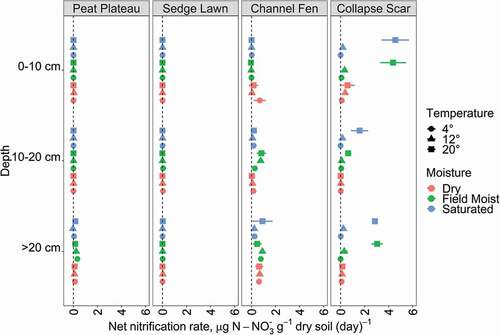
Across all treatments, landscape units, and depths, net nitrification was generally low (typically close to zero; ). No appreciable negative nitrification was observed (study-wide minimum value of −0.005 mg N g dry soil−1 over the entire 21 d incubation period). There was very low net nitrification (< 10 μg N g−1) in PP and SL, and minimal (primarily < 2 μg N g−1 d−1) responses to treatments in CF material. Net nitrification in the CS material generally increased at all three depths, although muted at the 10–20 cm depth, and only for the 20°C treatment in the field-moist and saturated samples. There were no significant individual effects or interactions of temperature and moisture treatments. Marginal changes were observed for dry samples or cooler temperatures across most landscape units and depth. There was a significant effect of landscape unit (H3,96 = 17.051, p < 0.001), with significantly higher nitrification occurring in both CF and CS than PP and SL (p < 0.01 in each case).
Figure 5. Net nitrification rates at each landscape unit (panels) and depth (y-axis), showing the effects of both temperature (shape) and moisture (color) treatments, with each point and bars showing the mean and standard deviation of four experimental replicates
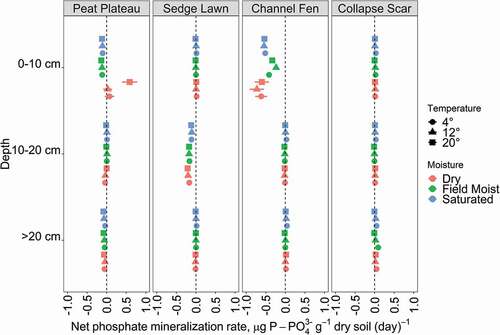
Phosphate
Overall, net P mineralization was very low, typically close to zero, with the exceptions of surficial material (0–10 cm) in PP and CF landscape units and in SL material at 10–20 cm (). Similar to net ammonification, there was a mixed response to treatments with both negative and positive net P mineralization rates, with a significant effect of landscape unit (H3,96 = 11.950, p < 0.001). There was significantly more negative P mineralization in PP, SL, and CF than CS (p < 0.001 in each case), but no significant differences among PP, SL, and CF net P mineralization rates. Both PP and CF had different phosphate transformations under dry conditions than were observed in field-moist or saturated treatments; however, the changes observed differed in direction between the two units. In the surface (0–10 cm) PP, drying marginally increased net phosphate mineralization, whereas in surface CF, drying led to marginally more negative rates (), although moisture and temperature did not have significant main effects or interaction.
Figure 6. Net phosphate mineralization rates at each landscape unit (panels) and depth (y-axis), showing the effects of both temperature (shape) and moisture (color) treatments, with each point and bars showing the mean and standard deviation of four experimental replicates
Relationships between peat properties and mineralization rates
There was a significant positive relationship between the initial ammonium pool and the net nitrification rate (r = 0.77, p < 0.010), indicating the potential for the nitrification of ammonium under oxic conditions. There were no other significant relationships between either net nitrification or ammonification with either initial extractable ammonium or nitrate pool size or total N. The presence of an appreciable initial water-extractable phosphorus pool was significantly negatively related to P mineralization rates (r = −0.96, p < 0.010), but did not significantly correlate with total P content.
Discussion
Extractable nutrient pools varied between landscape units and depth in this study, with the majority of TIN as NH4+, which tended to decrease with depth and increase downslope across landscape units. Extractable P pools were greatest in surface peat and diminished with depth. Temperature did not have a significant effect on any net mineralization rates in this study, while moisture had a significant effect on net ammonification. In each of net ammonification, nitrification, and P mineralization, there was a significant effect of landscape unit on cycling rates. There were two strong relationships between extractable pools and mineralization rates: a strong positive relationship between the initial ammonium pool and the net nitrification rate, and a strong negative relationship between the initial extractable phosphate pool and net P mineralization rate.
The significant correlation between the initial extractable pool of NH4+ and the net nitrification rate suggests that available ammonium may be readily nitrified under oxic conditions. This result, primarily as changes to extractable ammonium as opposed to nitrate, is consistent with results reported in field-based studies conducted by Rosswall and Granhall (Citation1980), Williams and Wheatley (Citation1988), and Bayley, Thormann, and Szumigalski (Citation2005). Similarly, in a lab-based study under oxic conditions, Venterink et al. (Citation2002) found that an increase in extractable nitrate in dried soils was attributed to increased ammonification and nitrification, not decreased denitrification, in which soil drying stimulated net total N mineralization (three times higher) and reduced denitrification (five times lower) compared to continuously wet soils, and that re-wetting may significantly increase P availability. In contrast, Macrae et al. (Citation2013) found that drying conditions decreased net nitrification rates across the landscape, but did not affect net PO43- mineralization rates. In this study, the greatest negative rates of P mineralization corresponded to the biggest initial pools of PO43- (), and changes to net P mineralization with moisture or temperature were not observed in landscape units with low initial PO43- pools.
Both net ammonification and net nitrification rates in our study are comparable with results from a wide range of both field-based and laboratory-based investigations of N and P mineralization in northern peatland and subarctic tundra soils, when normalized to a gravimetric and per-day basis (). In our study, CF and PP show both negative and positive net mineralization of phosphate in surface peat, which is within the upper range/orders of magnitude of other peatlands, comparable to a set of natural and harvested fen and bog sites in the Netherlands (Verhoeven et al. Citation1990), Canadian western boreal plains (Wood et al. Citation2016), and tundra/heath systems in Alaska (Nadelhoffer et al. Citation1991). As well, in collapse scar material in this study, which is absent in previous mineralization studies, the potential for extreme nitrification rates under different temperature and moisture treatments indicate distinct N dynamics in thermally disturbed material from undisturbed landscape units in this study and studies from across other boreal and northern environments. The difference in magnitude of rates between our study and others may also be related to length of incubation period and initial decomposition status. Decomposition and net mineralization rates have been shown to be dynamic over time and thus average rates (normailzed to a per-day basis) will differ depending on the incubation length (Arunachalam et al. Citation1996). Thormann and Bayley (Citation1997) show decreasing rates of decomposition throughout the course of a long-term incubation experiment (456 d) in a boreal peatland. This could potentially indicate that this relatively short-term study (and other studies of similar length) would capture higher rates than long-term studies when rates are normalized to a per-day basis, but this comparison is complicated by the fact that long-term in situ incubations would be carried out across multiple seasons with changing temperature and moisture regimes. Our incubation period of 21 d is comparable with the shorter end of the range of incubation lengths of several other in situ and laboratory studies (), which spans from a minimum of three weeks to multiyear incubations.
Table 3. Ranges of values for extractible nutrient pools and net mineralization rates presented in a nonexhaustive set of comparable study regions (both peatland and subarctic sites, either permafrost or seasonally frozen). We exclude studies/sites that include experimental treatments or samples from non-pristine environments. Total inorganic nitrogen (TIN) is the sum of ammonium and nitrate. All laboratory incubations were completed under oxic conditions
Given the strong relationships between initial extractable ammonium pools and nitrification rates as well as between phosphate pools and net P mineralization, it is also important to contextualize extractable pool sizes within other comparable study regions. Values of extractable TIN pools that are reported in this study for our non-thermally distributed landscape units (PP, SL, and CF) are comparable with ranges from several peatland and tundra sites (). The partitioning of TIN into NH4+ and NO3− in our study is similar to other boreal peatland regions as well, with the majority of TIN occurring as NH4+, as Wood et al. (Citation2016), Andersen et al. (Citation2013), and Macrae et al. (Citation2013) all report NH4+ as > 70 percent of TIN. The stratification of NH4+ with depth in our results in consistent with the findings of Williams and Wheatley (Citation1988) in a Scottish bog, attributed to similar stratification of decreasing concentrations of ammonifying bacteria with depth. In comparison with tundra communities, our extractable TIN pools are generally greater than results reported by Haag (Citation1974) for a tundra birch-willow-heath system in Alaska of 0.5–1.0 μg N-TIN g−1, although following fertilization experiments of up to 200 kg N ha−1, their extractable nitrogen pools are comparable with our results. The CS material in our study contained extractable NH4+, which were much greater than both the non-thermally distributed landscape units in this study (PP, SL, and CF) as well as reported values from many comparable studies (). The water-extractable P pool in this study is also similar to comparable environments, with the exception of CF and PP surface peat (corresponding as well to anomalously high P mineralization rates). All other extractable phosphate pools in our study are in the range of other permafrost environments, similar to our results in tundra heath (1–11 P − PO43- g−1) and sedge meadow (3–6 P − PO43- g−1) sites as reported by Haag (Citation1974), as well as tussock/sedge tundra and heath systems (0–5 μg P − PO43- g−1) in Alaska as reported by Nadelhoffer et al. (Citation1991).
The significant effect of landscape unit on the net mineralization of all nutrient species in this study indicates the varying response to similar environmental controls between different portions of ecosystems, which has been observed at the field scale (Bridgham, Updegraff, and Pastor Citation1998; Mettrop et al. Citation2014). Further, there appeared to be an effect of depth within landscape units on nutrient cycling, such as the increased net nitrification experienced at surface (0–10 cm) and deep (> 20 cm) material in CS material under warm and saturated conditions, but this effect was muted in the 10–20 cm material. There were differences in litter quality (total C:N:P ratios) in different landscape units and depths, but there were no significant relationships between total C, N, and P and any of ammonification, nitrification, and P mineralization rates. This suggests that there may be more complex factors, such as microbial community composition (Waring, Averill, and Hawkes Citation2013) and the availability of C:N:P in more labile forms (as opposed to bulk totals) may be the key factors for determining mineralization response to hydroclimatic drivers across landscape units and depth. Nutrient stoichiometric ratios of C:N:P in this study generally fell within the expected range for peatlands and tundra sites (). Wang et al. (Citation2015) also report C:N values from 50:1 to 20:1 generally decreasing with depth for bog peat across a gradient of 400 peat profiles and peatland types in Ontario, where our study found an average C:N of 23:1. Our C:N values are generally slightly higher than most subarctic tundra environments (Kelley et al. Citation2012; Nadelhoffer et al. Citation1991) driven by larger organic carbon content due to peat accumulation in the Hudson Bay Lowlands. Our N:P and C:P ratios are also slightly greater than those reported in literature at peatland sites because of low total P concentrations in our samples (, ). The relationship of C:P is close to the critical quotient for net P loss of 800–1,200 demonstrated by Moore et al. (Citation2008) and the C:N ratios in this study are primarily below the critical quotient of 33–68 for net N loss.
This work did not address the relative magnitude of all individual components in net ammonification, nitrification, and P mineralization, including microbial immobilization (Schmidt, Jonasson, and Michelsen Citation1999), denitrification (Aerts and Ludwig Citation1997), and ammonia volatilization (Bouwman et al. Citation1997), although not all of these processes are expected to contribute fluxes of any considerable magnitude depending on the pH and oxic status (Rosswall and Granhall Citation1980; Skrzypek et al. Citation2015). Through root removal and the absence of living vegetation, this study constrains some potential pathways for nutrient transformation, such as plant uptake (Larsen et al. Citation2012) and biological atmospheric fixation (Rousk et al. Citation2017), which are inherently present in field studies, and of non-negligible magnitude in northern environments (Stark Citation2007). The presence of roots (absent in this study) has a strong control on nutrient availability through controlling seasonal patterns of microbial biomass associated with temperature shifts above 0°C (Edwards et al. Citation2006), during which plants take up pulses of available nutrients (Edwards and Jefferies Citation2010). Further, other environmental processes, which are naturally incorporated in field-based studies, are absent here on multiple time scales. On an annual scale, atmospheric deposition of nutrients (both wet and dry) and hydrologic transport pathways both play a central role in nutrient supply and redistribution in the environment (Eppinga et al. Citation2010; Rosswall and Granhall Citation1980). On shorter time scales, diurnal and seasonal temperature fluctuations, both in summer and sub-snowpack in the winter (Buckeridge et al. Citation2010), influence nutrient transport and cycling processes, in contrast to the constant conditions maintained throughout the study period in this work.
Conclusions
This work has implications for future projections of nutrient dynamics in thawing permafrost ecosystems under a changing climate. Climatic changes will affect different components of the landscape distinctly, and thermokarst CS material particularly has the potential for unprecedented nitrogen turnover under warm conditions; specifically, net ammonification in surface material and net nitrification under moist conditions. As existing and newly thawed collapse scars become exposed to oxic conditions, pulses of plant-available nitrogen in the form of ammonium may be mineralized at a rate higher than undisturbed sites in warmer and either dryer or wetter conditions. There appears to be a temperature- and moisture-driven nitrification threshold in CS material, which was activated only in the 20°C treatment and in the two moist treatments. When collapse scars become exposed to oxic conditions through soil-structure disturbance or through a drawdown of the water table, increased net nitrification under warm and moist conditions may result in up to a 100-times increase in extractable nitrate concentrations from initial extractable pools over a matter of weeks. These results show potential for substantial plant-available nitrogen release to a nutrient-limited environment, where biological uptake of nitrogen has shown to be rapid in subarctic terrestrial (Edwards and Jefferies Citation2010) and aquatic (Eichel et al. Citation2014) systems. However, if the timing of nutrient release is outside of the primary periods for nutrient uptake (Larsen et al. Citation2012), nutrients may be subject to hydrologic redistribution to downstream fates (Frey and McClelland Citation2009). Phosphate concentrations were low across landscape units and depths, and generally showed a negative mineralization response under all temperature and moisture conditions and in all parts of the landscape, indicating the potential for microbial demand for the loosely bound (water-extractable) P pool. This suggests that phosphates liberated from decaying organic matter in permafrost peatlands may be rapidly immobilized into less labile pools, although process-based microbial ecology studies are required to understand the magnitude of this pathway. The significant effect of landscape unit controlling all mineralization rates in this study has important implications for understanding the complex controls on N and P mineralization potential, which likely are highly dependent on peat origin (landscape unit, depth) and chemical properties.
Acknowledgments
We thank D. Gibson for field assistance and J. Williamson, H. Rogers, K. Lutes, V. Lam, W. Mark, J. Harbin, J. Plach, and J. Cober for lab assistance.
Disclosure statement
The authors declare that they have no conflict of interest.
Additional information
Funding
Related Research Data
References
- Aerts, R., and F. Ludwig. 1997. Water-table changes and nutritional status affect trace gas emissions from laboratory columns of peatland soils. Soil Biology and Biochemistry 29 (11):1–15. doi:https://doi.org/10.1016/S0038-0717(97)00074-6.
- Allan, C. J., N. T. Roulet, and A. R. Hill. 1993. The biogeochemistry of pristine, headwater Precambrian shield watersheds: An analysis of material transport within a heterogeneous landscape. Biogeochemistry 22 (1):37–79. doi:https://doi.org/10.1007/BF00002756.
- Andersen, R., C. Wells, M. Macrae, and J. Price. 2013. Nutrient mineralisation and microbial functional diversity in a restored bog approach natural conditions 10 years post restoration. Soil Biology and Biochemistry 64:37–47. doi:https://doi.org/10.1016/j.soilbio.2013.04.004.
- Arunachalam, A., H. N. Pandey, R. S. Tripathi, and K. Maithani. 1996. Fine root decomposition and nutrient mineralization patterns in a subtropical humid forest following tree cutting. Forest Ecology and Management 86 (1–3):141–50. doi:https://doi.org/10.1016/S0378-1127(96)03784-X.
- Austin, A. T., and P. M. Vitousek. 1998. Nutrient dynamics on a precipitation gradient in Hawai’i. Oecologia 113 (4):519–29. doi:https://doi.org/10.1007/s004420050405.
- Bayley, S. E., M. N. Thormann, and A. R. Szumigalski. 2005. Nitrogen mineralization and decomposition in western boreal bog and fen peat. Écoscience 12 (4):455–65. doi:https://doi.org/10.2980/i1195-6860-12-4-455.1.
- Bintanja, R., and O. Andry. 2017. Towards a rain-dominated Arctic. Nature Climate Change 7 (4):263–67. doi:https://doi.org/10.1038/nclimate3240.
- Boone, R. D., D. F. Grigal, P. Sollins, R. J. Ahrens, and D. E. Armstrong. 1999. Soil sampling, preparation, archiving, and quality control. In Standard soil methods for long-term ecological research, eds. G. P. Robertson, D. C. Coleman, C. S. Bledsoe, and P. Sollins, 3–28. New York: Oxford University Press.
- Bouwman, A. F., D. S. Lee, W. A. H. Asman, F. J. Dentener, K. W. van der Hoek, and J. G. J. Olivier. 1997. A global high‐resolution emission inventory for ammonia. Global Biogeochemical Cycles 11 (4):561–87. doi:https://doi.org/10.1029/97GB02266.
- Bridgham, S. D., K. Updegraff, and J. Pastor. 1998. Carbon, nitrogen, and phosphorus mineralization in northern wetlands. Ecology 79 (5):1545–61. doi:https://doi.org/10.1890/0012-9658(1998)079[1545:CNAPMI]2.0.CO;2.
- Brown, R. D., and R. O. Braaten. 1998. Spatial and temporal variability of Canadian monthly snow depths, 1946–1995. Atmosphere-Ocean 36 (1):37–54. doi:https://doi.org/10.1080/07055900.1998.9649605.
- Buckeridge, K. M., Y. P. Cen, D. B. Layzell, and P. Grogan. 2010. Soil biogeochemistry during the early spring in low arctic mesic tundra and the impacts of deepened snow and enhanced nitrogen availability. Biogeochemistry 99 (1–3):127–41. doi:https://doi.org/10.1007/s10533-009-9396-7.
- Callaghan, T. V., F. Bergholm, T. R. Christensen, C. Jonasson, U. Kokfelt, and M. Johansson. 2010. A new climate era in the sub‐Arctic: Accelerating climate changes and multiple impacts. Geophysical Research Letters 37 (14). doi: https://doi.org/10.1029/2009GL042064.
- Cargill, S. M., and R. L. Jefferies. 1984. Nutrient limitation of primary production in a sub-arctic salt marsh. Journal of Applied Ecology 21 (2):657–68. doi:https://doi.org/10.2307/2403436.
- Chambers, C., J. D. Marshall, and R. J. Danehy. 2004. Nitrogen uptake and turnover in riparian woody vegetation. Oecologia 140 (1):125–34. doi:https://doi.org/10.1007/s00442-004-1565-8.
- Cohen, J., J. A. Screen, J. C. Furtado, M. Barlow, D. Whittleston, D. Coumou, J. Francis, K. Dethloff, D. Entekhabi, J. Overland, et al. 2014. Recent Arctic amplification and extreme mid-latitude weather. Nature Geoscience 7 (9):627–37. doi:https://doi.org/10.1038/ngeo2234.
- Connon, R. F., W. L. Quinton, J. R. Craig, and M. Hayashi. 2014. Changing hydrologic connectivity due to permafrost thaw in the lower Liard River valley, NWT, Canada. Hydrological Processes 28 (14):4163–78. doi:https://doi.org/10.1002/hyp.v28.14.
- Davies, G. M., and A. Gray. 2015. Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecology and Evolution 5 (22):5295–304. doi:https://doi.org/10.1002/ece3.1782.
- DeBusk, W. F., and K. R. Reddy. 1998. Turnover of detrital organic carbon in a nutrient-impacted Everglades marsh. Soil Science Society of America Journal 62 (5):1460–68. doi:https://doi.org/10.2136/sssaj1998.03615995006200050045x.
- Devito, K. J., C. J. Westbrook, and S. L. Schiff. 1999. Nitrogen mineralization and nitrification in upland and peatland forest soils in two Canadian Shield catchments. Canadian Journal of Forest Research 29 (11):1793–804. doi:https://doi.org/10.1139/x99-148.
- Dyke, L. D., and W. E. Sladen. 2010. Permafrost and peatland evolution in the northern Hudson Bay Lowland, Manitoba. Arctic 63 (4):429–41. doi:https://doi.org/10.14430/arctic3332.
- Edwards, K. A., and R. L. Jefferies. 2010. Nitrogen uptake by Carex aquatilis during the winter-spring transition in a low Arctic wet meadow. Journal of Ecology 98 (4):737–44. doi:https://doi.org/10.1111/j.1365-2745.2010.01675.x.
- Edwards, K. A., J. McCulloch, G. P. Kershaw, and R. L. Jefferies. 2006. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biology and Biochemistry 38 (9):2843–51. doi:https://doi.org/10.1016/j.soilbio.2006.04.042.
- Eichel, K. A., M. L. Macrae, R. I. Hall, L. Fishback, and B. B. Wolfe. 2014. Nutrient uptake and short-term responses of phytoplankton and benthic algal communities from a subarctic pond to experimental nutrient enrichment in microcosms. Arctic, Antarctic, and Alpine Research 46 (1):191–205. doi:https://doi.org/10.1657/1938-4246-46.1.191.
- Eppinga, M. B., M. Rietkerk, L. R. Belyea, M. B. Nilsson, P. C. D. Ruiter, and M. J. Wassen. 2010. Resource contrast in patterned peatlands increases along a climatic gradient. Ecology 91 (8):2344–55.
- Fellman, J. B., and D. V. D’Amore. 2007. Nitrogen and phosphorus mineralization in three wetland types in southeast Alaska, USA. Wetlands 27 (1):44–53. doi:https://doi.org/10.1672/0277-5212(2007)27[44:NAPMIT]2.0.CO;2.
- Frey, K. E., and J. W. McClelland. 2009. Impacts of permafrost degradation on arctic river biogeochemistry. Hydrological Processes 23 (1):169–82. doi:https://doi.org/10.1002/hyp.v23:1.
- Golovchenko, A. V., E. Y. Tikhonova, and D. G. Zvyagintsev. 2007. Abundance, biomass, structure, and activity of the microbial complexes of minerotrophic and ombrotrophic peatlands. Microbiology 76 (5):630–37. doi:https://doi.org/10.1134/S0026261707050177.
- Haag, R. W. 1974. Nutrient limitations to plant production in two tundra communities. Canadian Journal of Botany 52 (1):103–16. doi:https://doi.org/10.1139/b74-014.
- Hartsock, J. A., M. House, and D. H. Vitt. 2016. Net nitrogen mineralization in boreal fens: a potential performance indicator for peatland reclamation. Botany 94 (11):1027–1040.
- Heiri, O., A. F. Lotter, and G. Lemcke. 2001. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. Journal of Paleolimnology 25 (1):101–10. doi:https://doi.org/10.1023/A:1008119611481.
- Jones, D. L., K. Kielland, F. L. Sinclair, R. A. Dahlgren, K. K. Newsham, J. F. Farrar, and D. V. Murphy. 2009. Soil organic nitrogen mineralization across a global latitudinal gradient. Global Biogeochemical Cycles 23 (1). doi: https://doi.org/10.1029/2008GB003250.
- Jorgenson, M. T., and T. E. Osterkamp. 2005. Response of boreal ecosystems to varying modes of permafrost degradation. Canadian Journal of Forest Research 35 (9):2100–11. doi:https://doi.org/10.1139/x05-153.
- Jorgenson, M. T., Y. L. Shur, and E. R. Pullman. 2006. Abrupt increase in permafrost degradation in Arctic Alaska. Geophysical Research Letters 33 (2). doi: https://doi.org/10.1029/2005GL024960.
- Kaufman, D. S., D. P. Schneider, N. P. McKay, C. M. Ammann, R. S. Bradley, K. R. Briffa, G. H. Miller, B. L. Otto-Bliesner, J. T. Overpeck, B. M. Vinther, et al. 2009. Recent warming reverses long-term Arctic cooling. Science 325 (5945):1236–39. doi:https://doi.org/10.1126/science.1173983.
- Kelley, A. M., H. E. Epstein, C. L. Ping, and D. A. Walker. 2012. Soil nitrogen transformations associated with small patterned‐ground features along a North American Arctic Transect. Permafrost and Periglacial Processes 23 (3):196–206. doi:https://doi.org/10.1002/ppp.1748.
- Kuhry, P., and J. Turunen. 2006. The postglacial development of boreal and subarctic peatlands. In Boreal peatland ecosystems, eds. R. K. Wieder, and D. H. Vitt, 25–46. Berlin Heidelberg: Springer.
- Lafrenière, M. J., and S. F. Lamoureux. 2013. Thermal perturbation and rainfall runoff have greater impact on seasonal solute loads than physical disturbance of the active layer. Permafrost and Periglacial Processes 24 (3):241–51. doi:https://doi.org/10.1002/ppp.1784.
- Larsen, K. S., A. Michelsen, S. Jonasson, C. Beier, and P. Grogan. 2012. Nitrogen uptake during fall, winter and spring differs among plant functional groups in a subarctic heath ecosystem. Ecosystems 15 (6):927–39. doi:https://doi.org/10.1007/s10021-012-9555-x.
- Lawrence, D. M., A. G. Slater, and S. C. Swenson. 2012. Simulation of present-day and future permafrost and seasonally frozen ground conditions in CCSM4. Journal of Climate 25 (7):2207–25. doi:https://doi.org/10.1175/JCLI-D-11-00334.1.
- Lipson, D. A., D. Zona, T. K. Raab, F. Bozzolo, M. Mauritz, and W. C. Oechel. 2012. Water-table height and microtopography control biogeochemical cycling in an Arctic coastal tundra ecosystem. Biogeosciences 9 (1):577–91. doi:https://doi.org/10.5194/bg-9-577-2012.
- Long, M. S., C. M. Litton, C. P. Giardina, J. Deenik, R. J. Cole, and J. P. Sparks. 2017. Impact of nonnative feral pig removal on soil structure and nutrient availability in Hawaiian tropical montane wet forests. Biological Invasions 19 (3):749–63. doi:https://doi.org/10.1007/s10530-017-1368-6.
- Macrae, M. L., K. J. Devito, M. Strack, and J. M. Waddington. 2013. Effect of water table drawdown on peatland nutrient dynamics: Implications for climate change. Biogeochemistry 112 (1–3):661–76. doi:https://doi.org/10.1007/s10533-012-9730-3.
- Malmer, N., and B. Nihlgård. 1980. Supply and transport of mineral nutrients in a subarctic mire. Ecological Bulletins 30:63–95.
- Mamet, S. D., and G. P. Kershaw. 2013. Environmental influences on winter desiccation of Picea glauca foliage at treeline, and implications for treeline dynamics in northern Manitoba. Arctic, Antarctic, and Alpine Research 45 (2):219–28. doi:https://doi.org/10.1657/1938-4246-45.2.219.
- Mettrop, I. S., C. Cusell, A. M. Kooijman, and L. P. Lamers. 2014. Nutrient and carbon dynamics in peat from rich fens and Sphagnum-fens during different gradations of drought. Soil Biology and Biochemistry 68:317–28. doi:https://doi.org/10.1016/j.soilbio.2013.10.023.
- Moore, T. R., J. A. Trofymow, M. Siltanen, and L. M. Kozak. 2008. Litter decomposition and nitrogen and phosphorus dynamics in peatlands and uplands over 12 years in central Canada. Oecologia 157 (2):317–25. doi:https://doi.org/10.1007/s00442-008-1076-0.
- Morison, M. Q., M. L. Macrae, R. M. Petrone, and L. Fishback. 2017. Seasonal dynamics in shallow freshwater pond‐peatland hydrochemical interactions in a subarctic permafrost environment. Hydrological Processes 31 (2):462–75. doi:https://doi.org/10.1002/hyp.11043.
- Nadelhoffer, K. J., A. E. Giblin, G. R. Shaver, and J. A. Laundre. 1991. Effects of temperature and substrate quality on element mineralization in six arctic soils. Ecology 72 (1):242–53. doi:https://doi.org/10.2307/1938918.
- Natali, S. M., E. A. Schuur, C. Trucco, C. E. Hicks Pries, K. G. Crummer, and A. F. Baron Lopez. 2011. Effects of experimental warming of air, soil and permafrost on carbon balance in Alaskan tundra. Global Change Biology 17 (3):1394–407. doi:https://doi.org/10.1111/j.1365-2486.2010.02303.x.
- Osborne, T. Z., and R. D. DeLaune. 2013. Soil and sediment sampling of inundated environments. In Methods in biogeochemistry of Wetlands, eds. R. D. DeLaune, K. R. Reddy, C. J. Richardson, and J. P. Megonigal, 21–40. Madison: Soil Science Society of America.
- Parkinson, J. A., and S. E. Allen. 1975. A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Communications in Soil Science and Plant Analysis 6 (1):1–11. doi:https://doi.org/10.1080/00103627509366539.
- Payette, S., and A. Delwaide. 2000. Recent permafrost dynamics in a subarctic floodplain associated with changing water levels, Québec, Canada. Arctic, Antarctic, and Alpine Research 32 (3):316–23.
- Payette, S., A. Delwaide, M. Caccianiga, and M. Beauchemin. 2004. Accelerated thawing of subarctic peatland permafrost over the last 50 years. Geophysical Research Letters 31 (18). doi: https://doi.org/10.1029/2004GL020358.
- Quinton, W. L., M. Hayashi, and A. Pietroniro. 2003. Connectivity and storage functions of channel fens and flat bogs in northern basins. Hydrological Processes 17 (18):3665–84. doi:https://doi.org/10.1002/(ISSN)1099-1085.
- R Development Core Team. 2016. R: A language and environment for statistical computing, R foundation for statistical computing. Vienna, Austria. Accessed January 10, 2017. http://www.R-project.org.
- Richardson, C. J. 1985. Mechanisms controlling phosphorus retention capacity in freshwater wetlands. Science 228 (4706):1424–26. doi:https://doi.org/10.1126/science.228.4706.1424.
- Rosswall, T., and U. Granhall. 1980. Nitrogen cycling in a subarctic ombrotrophic mire. Ecological Bulletins 30:209–34.
- Roulet, N., T. I. M. Moore, J. Bubier, and P. Lafleur. 1992. Northern fens: Methane flux and climatic change. Tellus B 44 (2):100–05. doi:https://doi.org/10.3402/tellusb.v44i2.15429.
- Rouse, W. R. 1991. Impacts of Hudson Bay on the terrestrial climate of the Hudson Bay Lowlands. Arctic and Alpine Research 23 (1):24–30. doi:https://doi.org/10.2307/1551433.
- Rousk, K., P. A. Pedersen, K. Dyrnum, and A. Michelsen. 2017. The interactive effects of temperature and moisture on nitrogen fixation in two temperate-arctic mosses. Theoretical and Experimental Plant Physiology 29 (1):25–36. doi:https://doi.org/10.1007/s40626-016-0079-1.
- Rustad, L. E., J. Campbell, G. Marion, R. Norby, M. Mitchell, A. Hartley, J. Cornelissen, and J. Gurevitch. 2001. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126 (4):543–62. doi:https://doi.org/10.1007/s004420000544.
- Sannel, A. B. K., and P. Kuhry. 2009. Holocene peat growth and decay dynamics in sub‐arctic peat plateaus, west‐central Canada. Boreas 38 (1):13–24. doi:https://doi.org/10.1111/j.1502-3885.2008.00048.x.
- Schaefer, K., T. Zhang, L. Bruhwiler, and A. P. Barrett. 2011. Amount and timing of permafrost carbon release in response to climate warming. Tellus B 63 (2):165–80. doi:https://doi.org/10.1111/teb.2011.63.issue-2.
- Schimel, J. P., C. Bilbrough, and J. M. Welker. 2004. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biology and Biochemistry 36 (2):217–27. doi:https://doi.org/10.1016/j.soilbio.2003.09.008.
- Schmidt, I. K., S. Jonasson, and A. Michelsen. 1999. Mineralization and microbial immobilization of N and P in arctic soils in relation to season, temperature and nutrient amendment. Applied Soil Ecology 11 (2):147–60. doi:https://doi.org/10.1016/S0929-1393(98)00147-4.
- Schuur, E. A., J. Bockheim, J. G. Canadell, E. Euskirchen, C. B. Field, S. V. Goryachkin, S. Hagemann, P. Kuhry, P. M. Lafleur, H. Lee, et al. 2008. Vulnerability of permafrost carbon to climate change: Implications for the global carbon cycle. BioScience 58 (8):701–14. doi:https://doi.org/10.1641/B580807.
- Schuur, E. A. G., B. W. Abbott, W. B. Bowden, V. Brovkin, P. Camill, J. G. Canadell, J. P. Chanton, F. S. Chapin, T. R. Christensen, P. Ciais, et al. 2013. Expert assessment of vulnerability of permafrost carbon to climate change. Climatic Change 119 (2):359–74. doi:https://doi.org/10.1007/s10584-013-0730-7.
- Shaver, G. R., and F. S. Chapin. 1980. Response to fertilization by various plant growth forms in an Alaskan tundra: Nutrient accumulation and growth. Ecology 61 (3):662–75. doi:https://doi.org/10.2307/1937432.
- Skrzypek, G., B. Wojtuń, D. Richter, D. Jakubas, K. Wojczulanis-Jakubas, and A. Samecka-Cymerman. 2015. Diversification of nitrogen sources in various tundra vegetation types in the High Arctic. PloS One 10 (9). doi: https://doi.org/10.1371/journal.pone.0136536.
- Song, K. Y., K. D. Zoh, and H. Kang. 2007. Release of phosphate in a wetland by changes in hydrological regime. Science of the Total Environment 380 (1):13–18. doi:https://doi.org/10.1016/j.scitotenv.2006.11.035.
- Stark, S. 2007. Nutrient cycling in the tundra. In Nutrient cycling in terrestrial ecosystems, eds. P. Marschner, and Z. Rengel, 309–31. Berlin Heidelberg: Springer.
- Thormann, M. N., and S. E. Bayley. 1997. Decomposition along a moderate-rich fen-marsh peatland gradient in boreal Alberta, Canada. Wetlands 17 (1):123–37. doi:https://doi.org/10.1007/BF03160724.
- Uhlířová, E., H. Šantrůčková, and S. P. Davidov. 2007. Quality and potential biodegradability of soil organic matter preserved in permafrost of Siberian tussock tundra. Soil Biology and Biochemistry 39 (8):1978–89. doi:https://doi.org/10.1016/j.soilbio.2007.02.018.
- van Breemen, N., and A. C. Finzi. 1998. Plant-soil interactions: Ecological aspects and evolutionary implications. Biogeochemistry 42 (1–2):1–19. doi:https://doi.org/10.1023/A:1005962124317.
- Venterink, H. O., T. E. Davidsson, K. Kiehl, and L. Leonardson. 2002. Impact of drying and re-wetting on N, P and K dynamics in a wetland soil. Plant and Soil 243 (1):119–30. doi:https://doi.org/10.1023/A:1019993510737.
- Verhoeven, J. T., E. Maltby, and M. B. Schmitz. 1990. Nitrogen and phosphorus mineralization in fens and bogs. Journal of Ecology 78 (3):713–726.
- Vitt, D. H., L. A. Halsey, and S. C. Zoltai. 1994. The bog landforms of continental western Canada in relation to climate and permafrost patterns. Arctic and Alpine Research 26 (1):1–13. doi:https://doi.org/10.2307/1551870.
- Vonk, J. E., S. E. Tank, W. B. Bowden, I. Laurion, W. F. Vincent, P. Alekseychik, M. Amyot, M. F. Billet, J. Canario, R. M. Cory, et al. 2015. Reviews and syntheses: Effects of permafrost thaw on Arctic aquatic ecosystems. Biogeosciences 12 (23):7129–67. doi:https://doi.org/10.5194/bg-12-7129-2015.
- Wallenstein, M. D., S. K. McMahon, and J. P. Schimel. 2009. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils. Global Change Biology 15 (7):1631–39. doi:https://doi.org/10.1111/gcb.2009.15.issue-7.
- Walvoord, M. A., and B. L. Kurylyk. 2016. Hydrologic impacts of thawing permafrost—A review. Vadose Zone Journal 15 (6). doi: https://doi.org/10.2136/vzj2016.01.0010.
- Wang, M., T. R. Moore, J. Talbot, and J. L. Riley. 2015. The stoichiometry of carbon and nutrients in peat formation. Global Biogeochemical Cycles 29 (2):113–21. doi:https://doi.org/10.1002/gbc.v29.2.
- Waring, B. G., C. Averill, and C. V. Hawkes. 2013. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta‐analysis and theoretical models. Ecology Letters 16 (7):887–94. doi:https://doi.org/10.1111/ele.12125.
- Weintraub, M. N., and J. P. Schimel. 2005. Nitrogen cycling and the spread of shrubs control changes in the carbon balance of Arctic tundra ecosystems. BioScience 55 (5):408–15. doi:https://doi.org/10.1641/0006-3568(2005)055[0408:NCATSO]2.0.CO;2.
- Whalen, J. K., C. Chang, and B. M. Olson. 2001. Nitrogen and phosphorus mineralization potentials of soils receiving repeated annual cattle manure applications. Biology and Fertility of Soils 34 (5):334–41. doi:https://doi.org/10.1007/s003740100416.
- Williams, B. L., and R. E. Wheatley. 1988. Nitrogen mineralization and water-table height in oligotrophic deep peat. Biology and Fertility of Soils 6 (2):141–47. doi:https://doi.org/10.1007/BF00257664.
- Woo, M. K. 2012. Permafrost hydrology, 562. Berlin Heidelberg: Springer.
- Woo, M. K., and K. L. Young. 2003. Hydrogeomorphology of patchy wetlands in the High Arctic, polar desert environment. Wetlands 23 (2):291–309. doi:https://doi.org/10.1672/8-20.
- Woo, M. K., and K. L. Young. 2006. High Arctic wetlands: Their occurrence, hydrological characteristics and sustainability. Journal of Hydrology 320 (3):432–50. doi:https://doi.org/10.1016/j.jhydrol.2005.07.025.
- Wood, M. E., M. L. Macrae, M. Strack, J. S. Price, T. J. Osko, and R. M. Petrone. 2016. Spatial variation in nutrient dynamics among five different peatland types in the Alberta oil sands region. Ecohydrology 9 (4):688–99. doi:https://doi.org/10.1002/eco.v9.4.