
ABSTRACT
Predicted warming in Tibetan Plateau may have profound effects on plant community structure and function. We used open-top chambers to artificially warm two different plant communities in Tibet from 2012 to 2016. We recorded species richness, vegetation height, and graminoid, forb, legume, and litter cover each year of the experiment and leaf growth and chemical traits of the most dominant species after four years of warming. Our results showed that vegetation height increased under warming in both the alpine meadow and the swamp. Warming also marginally increased legume cover and C:N ratio of all species in the alpine meadow but not in the swamp, suggesting that plant growth rate and nitrogen cycling are higher in the meadow in a warmer future. An observed tradeoff between water use efficiency and nitrogen use efficiency in the alpine meadow, but not in the swamp, also indicated that resource use may be more efficient in plants in the drier meadow under future warming. Overall, our results support predictions that soil moisture may modify plant community responses to climate warming and that changes in carbon and nutrient cycling may be more pronounced in drier alpine meadows than in wetter swamps under climate warming.
Introduction
The Tibetan Plateau is regarded as the Earth’s third pole. It is the highest mountain plateau in the world, with the largest area of high-elevation permafrost. Stronger warming has been observed on the Tibetan Plateau than elsewhere, with an increase of 1.8°C over the past fifty years (Xu et al. Citation2017). Future warming in this region is predicted to be greater than the global average (Stocker et al. Citation2013). Climate warming can change the moisture regime in the shallow active layer of permafrost areas, which, when combined with higher temperatures, may primarily affect ecosystem processes and individual plant species performance and plant community structure (Christensen et al. Citation2003; Yang et al. Citation2018).
Meta-analyses of climate warming effects on alpine and arctic tundra have found that vegetation responses to higher temperatures may depend on soil moisture, because climate warming increased the abundance of some functional groups in wetter but not drier sites (Elmendorf, Henry, Hollister, Björk, Boulanger-Lapointe et al. Citation2012). This may suggest that plants growing under moist conditions are more resource acquisitive, whereas plants growing under drier conditions are resource conservative, resulting in less growth of the latter group (Bjorkman et al. Citation2018). On the Tibetan Plateau, a climate warming experiment increased above net primary production (ANPP) in an alpine mesic meadow but decreased ANPP in an alpine steppe with lower soil moisture (Ganjurjav et al. Citation2016). Furthermore, across the whole Tibetan Plateau, the soil moisture regime has been shown to mediate ANPP response to warming (Yang et al. Citation2018). A combined heating and watering experiment conducted in a moist alpine tundra habitat in North America increased ANPP (Winkler, Chapin, and Kueppers Citation2016). Overall, previous research suggests that water limitation can offset or even reverse plant growth under higher temperatures. Thus, further research is required to fully understand how alpine plant communities respond to climate warming and how these responses differ according to soil moisture regime.
Taller vegetation and increased biomass production due to warming may have cascading effects on ecosystem processes, such as competition for light and space, which again may affect plant species diversity (Klein, John, and Zhao Citation2004; Yang et al. Citation2015) and function (Hudson, Henry, and Cornwell Citation2011; Yang and Yang Citation2011). Plant functional traits related to C gain and water and nutrient use efficiency (Craine and Towne Citation2010) may determine the performance of individual species in a warmer future (Violle et al. Citation2007). Thus, the effects of climate warming may be species specific (Klein, John, and Zhao Citation2004; Yang et al. Citation2015).
Climate warming effects on individual plant species and their potential effect on ecosystem function can be studied by the use of plant traits such as leaf growth (specific leaf area (SLA) and leaf dry matter content (LDMC)) and chemical (C and N and C (δ13C) and N (δ15N) isotopes) traits that are associated with water and resource use efficiency. SLA, LDMC, C, and N are all associated with growth rate (Venn, Pickering, and Green Citation2013). Higher SLA suggests faster growth and more efficient or acquisitive resource use, resulting in larger and thinner leaves (Poorter and Bongers Citation2006) and thus more competitive plants. LDMC is a common indicator of plant species resource use strategy and usually correlates negatively with relative growth rate potential (Pérez-Harguindeguy et al. Citation2013). Leaf δ13C can be used as a measure of long-term water use efficiency (WUE) because they are highly correlated, indicating that plants with higher δ13C are more drought stressed. Leaf δ15N can be used to assess resource use (Hudson, Henry, and Cornwell Citation2011), which generally increases with increased net N input and decreased internal N cycling.
To examine climate warming effects within a permafrost region on the Tibetan Plateau and whether such effects differ according to soil moisture regime, we examined community structure and functional traits of the most common species in two communities, an alpine meadow and an alpine swamp. Open-top chambers (OTCs) were used to investigate whether (1) four years of experimental warming affected (a) air and soil temperature and soil moisture; (b) species richness, cover of different functional groups, litter, and vegetation height; and (c) leaf functional traits of dominant plant species and (2) the response to warming differed between the alpine meadow and alpine swamp plant communities. We hypothesized that warming by OTCs would increase plant cover and height in both the meadow and the swamp but more so in the swamp because moisture is not limited in that habitat. We predicted that plant functional trait responses to warming would reflect different resource use strategies between the drier meadow and mesic swamp.
Methods
Study site
This study was conducted in Fenghuoshan (92°50′–93°30′ E, 34°40′–34°48′ N) in the hinterland of the Tibetan Plateau, China. The mean annual temperature and precipitation are −5.3°C and 380.2 mm, respectively, with 80 percent of the precipitation falling during the growing season between May and September (Yang et al. Citation2015). The study sites are within a region of continuous permafrost with an active layer of 0.8 to 1.5 m (Zhou et al. Citation2000).
Alpine meadows and swamps are the two most common plant communities in the area. Alpine meadows have moderate water availability and consist mainly of cold-tolerant perennial herbs. Alpine swamps are waterlogged or moist, mainly occurring in patches or strips in the mountains, and are dominated by perennial hygrophilous or hygro-mesophilic herbs (Li et al. Citation2011). Alpine meadows consist primarily of graminoids, such as Kobresia pygmaea, Carex scabrirostris, C. moorcroftii, and Poa pratensis, and forbs, such as Oxytropis deflexa, Halerpestes tricuspis, Polygonum viviparum, Pedicularis longiflora var. tubiformis, Androsace tanggulashanensis, and Leontopodium leontopodioides (Yang and Yang Citation2011). Alpine swamps also consist primarily of graminoids with some of the same species as those in the meadows, such as K. tibetica, C. scabriostris, and P. pratensis, in addition to K. humilis, K. capillifolia, C. atrofusca, Elymus mutans, and a few forbs, such as Saussurea superba, P. viviparum, Oxytropis, Pedicularis oederi var. sinensis, and Gentiana squarrosa. The study sites were grazed by yak before the experiment was set up but fenced, and therefore not grazed, during the experimental period.
Experimental design
We used OTCs (6-mm transparent Plexiglas, hexagonal, 30° sloped sides, 40 cm height, ~1 m2 area) as a passive warming device to artificially warm plant communities (Marion et al. Citation1997). The OTCs were left in the field throughout each year of the experimental period.
We had six replicate blocks of one OTC and one control plot in each of the two sites (twelve plots per site in total). The distance between an OTC and control plot was approximately 3 to 4 m, and the distance between the blocks ranged from 6 to 8 m. All plots were randomly positioned within one homogenous area; that is, with approximately similar microhabitat characteristics, surface conditions, vegetation cover, and species composition within each site.
Data collection
We measured air temperature at 20 cm above ground level, soil temperature at 20 cm depth, and soil water content at 5 cm depth in OTC and control plots in both the alpine meadow and swamp. One sensor per treatment was installed at each site. Measurements were taken at 30-minute intervals by thermometers and thermistor sensors (EM50 Data Collection System, Decagon Devices, Inc.) during the whole experimental period. A meteorology station was installed 10 m from the site to measure precipitation.
The percentage cover of all vascular plant species in all plots was visually estimated during the peak growing season in each year from 2012 to 2016. A 50 × 50 cm quadrat divided into a grid of 100 5 × 5 cm subquadrats was fixed in the center of each plot. All species present in each subquadrat were recorded. Vegetation height was measured at ten fixed points in each plot. We selected the most common species in the meadow (sedges: K. pygmaea and C. scabriostris, grass: Elymus nutans) and in the swamp (sedges: K. tibetica and C. scabrirostris) as target species and harvested leaves to measure leaf traits in one year (August 2016). Ten fully expanded and sun-exposed leaves of each species were selected from each plot. The leaves were clipped off the plant and placed in ziplock bags for leaf trait measurements.
To calculate SLA and LDMC, the fresh leaves were weighed and scanned (CanoScan LiDE 110, Canon China Inc., Beijing, China), and the area was measured with ImageJ (National Institutes of Health). The leaves were then oven dried at 65°C for 48 hours and weighed again. To prepare the leaves for nutrient analyses, we ground them using a mill and passed them through a 0.15-mm sieve. The leaves were analyzed for C and N concentrations and δ13C and δ15N on a stable isotope ratio mass spectrometer (Vario PYRO Cube, Elementar Analysensysteme, Germany).
Statistical analysis
Repeated measures analysis of variance (ANOVA) was used to examine the effects of community (meadow vs. swamp), warming treatment (OTC vs. control), and their interaction with year (2012, 2013, 2014, 2015, 2016) as within-subject comparisons of species richness, vegetation height, and cover of graminoids, forbs, legumes, and litter in the alpine meadow and swamp. We used one-way ANOVA to examine the effects of warming on SLA, LDMC, leaf C and N concentrations, and δ13C and δ15N at each of the sites separately. Statistical significance was considered at p < .05. All analyses were performed in R 3.5.3 (R Core Team Citation2018) and citation(lme4) (Bates et al. Citation2015).
Results
Temperature and soil moisture
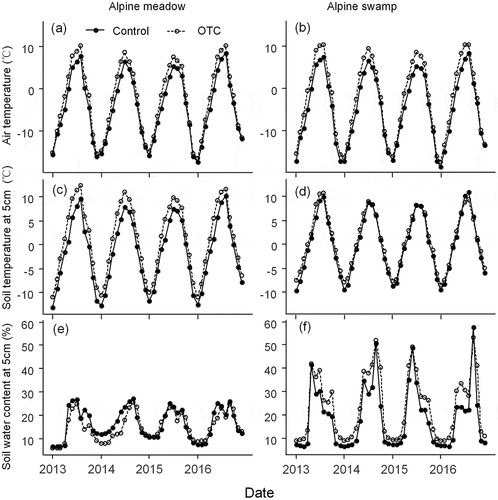
Over four years of warming, the air temperature in the OTCs was 2.52°C and 2.95°C higher than in the control plots in the alpine meadow and the swamp, respectively, during the growing season (May to September). Annual air temperature in the OTCs was 1.8°C and 2.2°C higher than those in control plots in the alpine meadow and the swamp, respectively (, ). Annual soil temperature at 5 cm depth increased in the OTCs by 2.6°C in the meadow and 0.8°C in the swamp (, ). Ambient soil moisture at 5 cm depth was on average 23.6 percent higher in the swamp than in the meadow during the growing season and decreased by 3.45 percent in the OTCs in the meadow and increased by 5.26 percent in the OTCs in the swamp when compared with the controls. Ambient mean annual soil moisture at 5 cm depth was 2.1 percent higher in the swamp than in the meadow and decreased by 1.3 percent in the OTCs in the meadow and increased by 5 percent in the OTCs in the swamp (). However, mean annual changes in soil moisture, soil temperature, and air temperature ranged from 1.1 to 1.6 percent, 0.2°C to 0.3°C, and 0.2°C to 0.4°C in both the meadow and swamp. Mean seasonal changes in soil moisture, soil temperature, and air temperature ranged from 0.6 to 2.8 percent, 0.3°C to 0.6°C, and 0.5°C to 0.7°C in both the meadow and swamp.
Figure 1. Mean monthly air temperature at 20 cm aboveground, mean monthly soil temperature at 5 cm depth, and mean monthly soil water content at 5 cm depth in control and warming plots measured in alpine meadow and alpine swamp in the Tibetan Plateau from 2013 to 2016. Control, solid circle; Open-Topped Chamber (OTC) warming, solid triangle
Warming effects on community structure
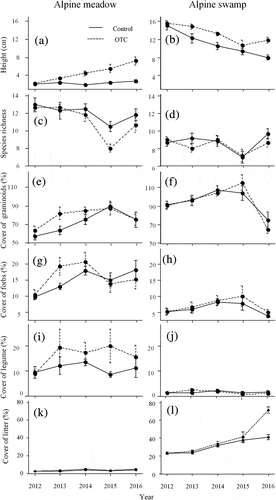
Vegetation height increased over time in the meadow, and the increase was significantly higher in the OTCs than in the controls (, ). In contrast, vegetation height decreased steadily over the study period in the alpine swamp, in both the controls and OTCs, but was generally higher in the OTCs (, ). Plant species richness decreased over four years in the alpine meadow, but there was no effect of the warming treatment in the meadow or swamp (, , ). There was no effect of warming on the cover of graminoids and forbs (, ). There was a marginal effect of warming and of the interaction between warming and community on the cover of legumes, with increasing cover in the OTCs in the meadow but not in the swamp (–2j, ). All community traits, including species richness, vegetation height, cover of functional groups, and litter, significantly varied between the alpine meadow and swamp. Litter cover in the alpine meadow slightly changed over time, ranging from 2.5 to 4 percent in the controls and from 2.5 to 5 percent in the OTCs. However, in the swamp, litter cover increased from 24 to 42 percent in controls and from 24 to 73 percent in the OTCs between 2012 and 2016 (–2l, ).
Table 1. Results (F and p values) of repeated measures ANOVA on the effects of the warming treatment (OTC vs. control), site, and their interactions on species richness, vegetation height, and cover of graminoids, forbs, legumes, and litter
Figure 2. Vegetation height; species richness; and cover of graminoids, forbs, legumes, and litter in control and warming plots in alpine meadow and alpine swamp. Control, solid line; OTC warming, dashed line. Data are shown as mean ± SE (n = 6)
Warming effects on leaf functional traits
Warming increased the SLA of E. nutans in the alpine meadow and of C. scabrirostris in the swamp (, , ). Warming also increased the LDMC of E. nutans in the meadow and of C. scabrirostris in both communities (, ).
Table 2. Results (F and p values) of ANOVA on the effects of the warming treatment (OTC vs. control) on SLA, LDMC, carbon isotope values (δ13C), nitrogen isotope values (δ15N), carbon concentration, nitrogen concentration, and C:N
Figure 3. Specific leaf area and leaf dry matter content for the most common species in control and OTC plots in alpine meadow and swamp in the Tibetan Plateau. Data are shown as mean ± SE (n = 6). Asterisks indicate significant (p < .05) differences between control and OTCs
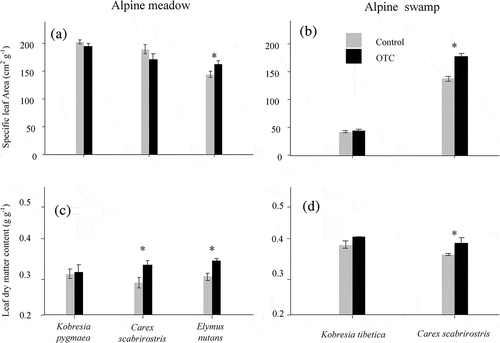
Figure 4. Leaf C and N isotopes, C and N concentrations, and C:N for the most common species in control and warming plots in alpine meadow and alpine swamp of the Tibetan Plateau. Data are shown as mean ± SE (n = 6). Asterisks indicate significant (p < .05) differences between control and OTCs
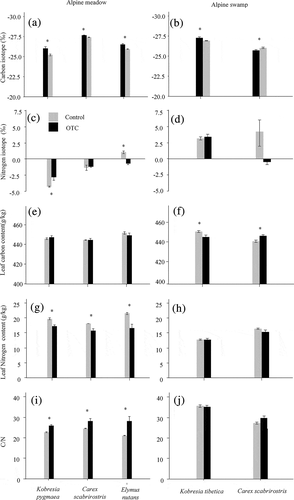
Warming decreased δ13C in all species in both communities, except for K. tibetica in the swamp, where δ13C increased with warming (, ). Furthermore, warming increased δ15N in K. pygmaea but decreased δ15N in E. nutans in the meadow (, ). Warming did not affect δ15N in any species in the swamp (, ). Warming had no effect on leaf C concentration in any of the species in the alpine meadow (, ) but affected both species in the swamp, with a decrease in leaf C in K. tibetica and an increase in C. scabrirostris (, ). Warming decreased leaf N concentration and increased leaf C:N ratio in all species in the alpine meadow (, 4i, ) but had no effect on any of the species in the swamp (–4j, ).
Discussion
Our four-year warming experiment in the permafrost region of the Tibetan Plateau showed that plant community structure and functional traits responded slightly different according to moisture regime, in line with our hypothesis. Vegetation height increased as a response to warming in both the meadow and the swamp, which agrees with other studies showing increased canopy height under climate warming in alpine and arctic tundra (Elmendorf, Henry, Hollister, Björk, Bjorkman et al. Citation2012; Elmendorf, Henry, Hollister, Björk, Boulanger-Lapointe et al. Citation2012; Bjorkman et al. Citation2018). The decrease in vegetation height observed over time in the swamp in both the OTCs and control plots, however, was not expected. This may have been caused by our exclusion of herbivores reducing grazing, thus increasing litter (Luo et al. Citation2010). A thick cover of litter reduces light and creates a mechanical barrier for new seedling and shoot emergence, thus inhibiting plant growth and establishment (Klein, John, and Zhao Citation2004), and may decrease vegetation height over time.
Although many studies have reported that graminoids in general increase in abundance under warmer climates (Walker et al. Citation2006; Elmendorf, Henry, Hollister, Björk, Boulanger-Lapointe et al. Citation2012; Klanderud, Vandvik, and Goldberg Citation2015), the response of grasses and sedges to warming may depend on soil moisture (Elmendorf, Henry, Hollister, Björk, Boulanger-Lapointe et al. Citation2012). Interestingly, we observed no warming effect on graminoids in our study and even a decrease in graminoid cover over time in the alpine swamp. Conversely, warming marginally increased the cover of legumes in the meadow. This result agrees with those reported in other studies from the alpine steppe of the Tibetan Plateau (Ganjurjav et al. Citation2016), prairies in North America (Whittington, Tilman, and Powers Citation2013; Cowles et al. Citation2016) and grasslands in France (Cantarel, Bloor, and Soussana Citation2012), where legumes also increased in abundance under experimental warming. These studies proposed that legume abundance increased because soil moisture decreased (Ganjurjav et al. Citation2016) in the warming plots, which is also in line with the decreased soil moisture in our warmed meadow plots. Drier soils and cryoturbation induced by warming may lead to nitrogen deficiency in the upper soil layer (Chang et al. Citation2017). In habitats with nitrogen as a limiting factor for plant growth, such as alpine tundra, plants with alternative strategies for acquiring resources, such as nitrogen-fixing legumes, may have a competitive advantage (Halvorson, Smith, and Franz Citation1991; Nadelhoffer et al. Citation1996; Vitousek et al. Citation2002). Thus, our results support predictions and previous findings from the northern Mongolian steppe (Cowles et al. Citation2018) that plant community response to warmer climates may depend on soil moisture conditions and that legumes may increase in abundance in sites where soil moisture and nitrogen are limited.
We also hypothesized that plant functional trait responses to warming would reflect resource use strategies related to soil moisture. The C:N ratio has been used to evaluate long-term N use efficiency (NUE; Livingston, Guy, and Ethier Citation1999), with higher NUE indicating plant growth using less N resources. Thus, the higher leaf C:N ratio in the meadow and unchanged C:N ratio in the swamp under experimental warming indicate that plants in the meadow are more efficient in utilizing N for growth than in the swamp. Warming increased the SLA of two of the dominant species (E. nutans and C. scabrirostris) in both the alpine meadow and the swamp, suggesting increased resource acquisition in warmer climates, in line with many other studies (e.g., Guittar et al. Citation2016). That the higher SLA was observed in only two of the dominant species in our warmed plots suggests that the ability to utilize improved growing conditions—that is, changing from resource conservative to acquisitive in warmer climates (Grady and Whitham Citation2013)—may be species specific. This may also result in changes in competitive interactions between species in these communities. The increase in LDMC in the warming plots in our study indicates tougher leaves related to more drought stress and less productivity. LDMC increased in both C. scabrirostris and E. nutans in the meadow, which may support the prediction that the drier communities are becoming even more limited by soil moisture in a warmer future, which is also in line with previous studies from seminatural grasslands (Cantarel, Bloor, and Soussana Citation2012).
δ15N can be used to qualitatively assess resource use and partitioning and is affected by many factors, such as N source, root depth, mycorrhizal associations, and the form of N taken up (Evans Citation2001). In our study, warming increased the δ15N of K. pygmaea but decreased δ15N of E. nutans in the alpine meadow. No effect was observed in any species in the alpine swamp. The interspecific difference in the response of δ15N to warming can be attributed to differences in N uptake (S. P. Wang et al. Citation2012; W. Y. Wang et al. Citation2014), suggesting that there may be a shift in species toward those that are able to utilize available N under future warmer climates.
To adapt to changes in environmental conditions, plants could change the C:N ratio, δ13C, or δ15N to reach a new equilibrium. In our study, we observed opposite responses of δ13C and C:N to warming in the alpine meadow, indicating a trade-off between WUE and NUE. This is consistent with previously reported negative correlations between WUE and NUE in Abies fabri in eastern Tibet (Yang et al. Citation2012) and in boreal spruce species (Patterson, Guy, and Dang Citation1997) and may suggest that plants increase their NUE by sacrificing WUE to increase growth in the alpine meadows.
Our results from the permafrost region of the Tibetan Plateau highlight that vegetation responses to warming may differ between communities according to soil moisture conditions (Elmendorf, Henry, Hollister, Björk, Boulanger-Lapointe et al. Citation2012; Cowles et al. Citation2018). Indeed, a meta-analysis based on data from the Tibetan Plateau found that aboveground biomass was positively related to precipitation and soil moisture and negatively related to temperature in alpine grasslands (Sun, Cheng, and Li Citation2013). This also agrees with results of another previous warming experiment, highlighting the importance of soil moisture modifying plant productivity and community species composition responses to warming in alpine meadows and steppe on the Tibetan Plateau (Ganjurjav et al. Citation2016). The different responses in terms of resource use strategies between alpine meadow and swamp communities to climate warming highlight that different management strategies should be implemented for different vegetation types on the Tibetan Plateau.
Acknowledgments
We thank three anonymous reviewers and the editor for constructive comments on previous versions of the article. Our gratitude extends to the Fenghuoshan Field Station of the Northwest Research Institute Co., Ltd., of China Railway Engineering Corporation for their assistance in the field and the Alpine Ecosystem Observation and Experiment Station of Mt. Gongga, CAS, for their assistance in chemical analysis. We thank Xiaopeng Chen, Chunlin Song, and Xiaoxiang Zhao for help in the field and lab and Xiaoyan Song for advice on data analysis.
Additional information
Funding
References
- Bates, D., M. Maechler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67 (1):1–48. doi:https://doi.org/10.18637/jss.v067.i01.
- Bjorkman, A. D., I. H. Myers-Smith, S. C. Elmendorf, S. Normand, N. Rüger, P. S. A. Beck, A. Blach-Overgaard, D. Blok, J. H. C. Cornelissen, B. C. Forbes, et al. 2018. Plant functional trait change across a warming tundra biome. Nature 562(7725):57–62. doi:https://doi.org/10.1038/s41586-018-0563-7.
- Cantarel, A. A. M., J. M. G. Bloor, and J. F. Soussana. 2012. Four years of simulated climate change reduces above‐ground productivity and alters functional diversity in a grassland ecosystem. Journal of Vegetation Science 24 (1):113–26. doi:https://doi.org/10.1111/j.1654-1103.2012.01452.x.
- Chang, R. Y., G. X. Wang, Y. H. Yang, and X. P. Chen. 2017. Experimental warming increased soil nitrogen sink in the Tibetan permafrost. Journal of Geophysical Research Biogeoscience 122:1870–79. doi:https://doi.org/10.1002/2017JG003827.
- Christensen, T. R., A. Ekberg, L. Ström, M. Mastepanov, N. Panikov, M. Öquist, B. H. Svensson, H. Nykänen, P. J. Martikainen, and H. Oskarsson. 2003. Factors controlling large scale variations in methane emissions from wetlands. Geophysical Research Letters 30 (7):1414–19. doi:https://doi.org/10.1029/2002GL016848.
- Cowles, J. M., B. Boldgiv, P. Liancourt, P. S. Petraitis, and B. B. Casper. 2018. Effects of increased temperature on plant communities depend on landscape location and precipitation. Ecology and Evolution 8:5267–78. doi:https://doi.org/10.1002/ece3.3995.
- Cowles, J. M., P. D. Wragg, A. J. Wright, J. S. Powers, and D. Tilman. 2016. Shifting grassland plant community structure drives positive interactive effects of warming and diversity on aboveground net primary productivity. Global Change Biology 22:741–49. doi:https://doi.org/10.1111/gcb.13111.
- Craine, J. M., and E. G. Towne. 2010. High leaf tissue density grassland species consistently more abundant across topographic and disturbance contrasts in a North American tallgrass prairie. Plant & Soil 337 (1–2):193–203. doi:https://doi.org/10.1007/s11104-010-0515-y.
- Elmendorf, S. C., G. H. Henry, R. D. Hollister, R. G. Björk, A. D. Bjorkman, T. V. Callaghan, L. S. Collier, E. J. Cooper, J. H. Cornelissen, and T. A. Day. 2012. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecology Letters 15 (2):164–75. doi:https://doi.org/10.1111/j.1461-0248.2011.01716.x.
- Elmendorf, S. C., G. H. R. Henry, R. D. Hollister, R. G. Björk, N. Boulanger-Lapointe, E. J. Cooper, J. H. C. Cornelissen, T. A. Day, E. Dorrepaal, and T. G. Elumeeva. 2012. Plot-scale evidence of tundra vegetation change and links to recent summer warming. Nature Climate Change 2 (6):453–57. doi:https://doi.org/10.1038/nclimate1465.
- Evans, R. D. 2001. Physiological mechanisms influencing plant nitrogen isotope composition. Trends in Plant Science 6 (3):121–26. doi:https://doi.org/10.1016/S1360-1385(01)01889-1.
- Ganjurjav, H., Q. Gao, E. S. Gornish, M. W. Schwartz, Y. Liang, X. Cao, W. Zhang, Y. Zhang, W. Li, and Y. Wan. 2016. Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai–Tibetan Plateau. Agricultural & Forest Meteorology 223:233–40. doi:https://doi.org/10.1016/j.agrformet.2016.03.017.
- Grady, K. C., and T. G. Whitham. 2013. Conservative leaf economic traits correlate with fast growth of genotypes of a foundation riparian species near the thermal maximum extent of its geographic range. Functional Ecology 27 (2):428–38. doi:https://doi.org/10.1111/1365-2435.12060.
- Guittar, J., D. Goldberg, K. Klanderud, R. Telford, and V. Vandvik. 2016. Can trait patterns along gradients predict plant community responses to climate change? Ecology 97 (10):2791. doi:https://doi.org/10.1002/ecy.1500.
- Halvorson, J. J., J. L. Smith, and E. H. Franz. 1991. Lupine influence on soil carbon, nitrogen, and microbial activity in developing ecosystems at Mount St. Helens. Oecologia 87:162–70. doi:https://doi.org/10.1007/BF00325253.
- Hudson, J., G. Henry, and W. Cornwell. 2011. Taller and larger: Shifts in Arctic tundra leaf traits after 16 years of experimental warming. Global Change Biology 17 (2):1013–21. doi:https://doi.org/10.1111/gcb.2010.17.issue-2.
- Klanderud, K., V. Vandvik, and D. Goldberg. 2015. The importance of biotic vs. abiotic drivers of local plant community composition along regional bioclimatic gradients. PLoS One 10 (6):e0130205. doi:https://doi.org/10.1371/journal.pone.0130205.
- Klein, J. A., H. John, and X. Q. Zhao. 2004. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecology Letters 7 (12):1170–79. doi:https://doi.org/10.1111/j.1461-0248.2004.00677.x.
- Li, N., G. Wang, Y. Yang, Y. Gao, and G. Liu. 2011. Plant production, and carbon and nitrogen source pools, are strongly intensified by experimental warming in alpine ecosystems in the Qinghai-Tibet Plateau. Soil Biology & Biochemistry 43 (5):942–53. doi:https://doi.org/10.1016/j.soilbio.2011.01.009.
- Livingston, N. J., R. D. Guy, and G. J. Ethier. 1999. The effects of nitrogen stress on the stable carbon isotope composition, productivity and water use efficiency of white spruce (Picea glauca (Moench) Voss) seedlings. Plant Cell and Environment 22:281–89. doi:https://doi.org/10.1046/j.1365-3040.1999.00400.x.
- Luo, C. Y., G. P. Xu, Z. G. Chao, S. P. Wang, X. W. Lin, Y. G. Hu, Z. H. Zhang, J. C. Duan, X. F. Chang, A. L. Su, et al. 2010. Effect of warming and grazing on litter mass loss and temperature sensitivity of litter and dung mass loss on the Tibetan Plateau. Global Change Biology 16:1606–17. doi:https://doi.org/10.1111/gcb.2010.16.issue-5.
- Marion, G. M., G. H. R. Henry, D. W. Freckman, J. Johnstone, G. Jones, M. H. Jones, E. Lévesque, U. Molau, P. Mølgaard, and A. N. Parsons. 1997. Open-top designs for manipulating field temperature in high-latitude ecosystems. Global Change Biology 3 (s1):20–32. doi:https://doi.org/10.1111/j.1365-2486.1997.gcb136.x.
- Nadelhoffer, K., G. Shaver, B. Fry, A. Giblin, L. Johnson, and R. McKane. 1996. 15N natural abundances and N use by tundra plants. Oecologia 107 (3):386–94. doi:https://doi.org/10.1007/BF00328456.
- Patterson, T. B., R. D. Guy, and Q. L. Dang. 1997. Whole-plant nitrogen- and water-relations traits, and their associated trade-offs, in adjacent muskeg and upland boreal spruce species. Oecologia 110 (2):160–68. doi:https://doi.org/10.1007/s004420050145.
- Pérez-Harguindeguy, N., S. Díaz, E. Garnier, S. Lavorel, H. Poorter, P. Jaureguiberry, M. S. Bret-Harte, W. K. Cornwell, J. M. Craine, D. E. Gurvich, et al. 2013. New handbook for standardized measurement of plant functional traits worldwide. Australian Journal of Botany 61:167–234. doi:https://doi.org/10.1071/BT12225.
- Poorter, L., and F. Bongers. 2006. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 87 (7):1733–43. doi:https://doi.org/10.1890/0012-9658(2006)87[1733:LTAGPO]2.0.CO;2.
- R Core Team. 2018. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. http://www.R-project.org/.
- Stocker, T. F., D. H. Qin, G. K. Plattner, M. M. B. Tignor, S. K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex, and P. M. Midgley. 2013. Climate change 2013. The physical science basis: Working Group I contribution to the Fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK.
- Sun, J., G. W. Cheng, and W. P. Li. 2013. Meta-analysis of relationships between environmental factors and aboveground biomass in the alpine grassland on the Tibetan Plateau. Biogeosciences 10 (3):1707–15. doi:https://doi.org/10.5194/bg-10-1707-2013.
- Venn, S., C. Pickering, and K. Green. 2013. Spatial and temporal functional changes in alpine summit vegetation are driven by increases in shrubs and graminoids. AoB Plants 6 (6):490–552.
- Violle, C., M. L. Navas, D. Vile, E. Kazakou, C. Fortunel, I. Hummel, and E. Garnier. 2007. Let the concept of trait be functional! Oikos 116:882–92. doi:https://doi.org/10.1111/j.0030-1299.2007.15559.x.
- Vitousek, P. M., S. Hättenschwiler, L. Olander, and S. Allison. 2002. Nitrogen and nature. Ambio 31 (2):97–101. doi:https://doi.org/10.1579/0044-7447-31.2.97.
- Walker, M. D., C. H. Wahren, R. D. Hollister, G. H. Henry, L. E. Ahlquist, J. M. Alatalo, M. S. Bret-Harte, M. P. Calef, T. V. Callaghan, and A. B. Carroll. 2006. Plant community responses to experimental warming across the tundra biome. Proceedings of the National Academy of Sciences USA 103 (5):1342–46. doi:https://doi.org/10.1073/pnas.0503198103.
- Wang, S. P., J. C. Duan, G. P. Xu, Y. F. Wang, Z. H. Zhang, Y. C. Rui, C. Y. Luo, B. Xu, X. X. Zhu, X. F. Chang, et al. 2012. Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 93 (11):23652376. doi:https://doi.org/10.1890/11-1408.1.
- Wang, W. Y., H. K. Zhou, L. Yang, J. P. Li, and X. C. Wang. 2014. The uptake strategy of soil nitrogen nutrients by different plant species in alpine Kobresia tibetica meadow on the Qinghai-Tibet Plateau. Journal of Natural Resources 29 (2):249–55.
- Whittington, H. R., D. Tilman, and J. S. Powers. 2013. Consequences of elevated temperatures on legume biomass and nitrogen cycling in a field warming and biodiversity experiment in a North American prairie. Functional Plant Biology 40:1147–58. doi:https://doi.org/10.1071/FP12345.
- Winkler, D. E., K. J. Chapin, and L. M. Kueppers. 2016. Soil moisture mediates alpine life form and community productivity responses to warming. Ecology 97 (6):1553–63. doi:https://doi.org/10.1890/15-1197.1.
- Xu, Y. M., A. Knudby, H. C. Ho, Y. Chen, and Y. H. Liu. 2017. Warming over the Tibetan Plateau in the last 55 years based on area-weighted average temperature. Regional Environmental Change 17:2339–47. doi:https://doi.org/10.1007/s10113-017-1163-z.
- Yang, Y., J. Guo, G. Wang, L. Yang, and Y. Yang. 2012. Effects of drought and nitrogen addition on photosynthetic characteristics and resource allocation of Abies fabri seedlings in eastern Tibetan Plateau. New Forests 43 (4):505–18. doi:https://doi.org/10.1007/s11056-011-9295-3.
- Yang, Y., K. A. Hopping, G. X. Wang, J. Chen, A. H. Peng, and J. A. Klein. 2018. Permafrost and drought regulate vulnerability of Tibetan Plateau grasslands to warming. Ecosphere 9 (5):e02233. doi:https://doi.org/10.1002/ecs2.2233.
- Yang, Y., G. Wang, K. Klanderud, J. Wang, and G. Liu. 2015. Plant community responses to five years of simulated climate warming in an alpine fen of the Qinghai–Tibetan Plateau. Plant Ecology & Diversity 8 (2):211–18. doi:https://doi.org/10.1080/17550874.2013.871654.
- Yang, Y., and L. Yang. 2011. Responses in leaf functional traits and resource allocation of a dominant alpine sedge (Kobresia pygmaea) to climate warming in the Qinghai-Tibetan Plateau permafrost region. Plant and Soil 349 (1):377–87. doi:https://doi.org/10.1007/s11104-011-0891-y.
- Zhou, Y., D. Guo, G. Qiu, G. Cheng, and S. Li. 2000. Geocryology in China. Beijing: Chinese Academy of Sciences. [in Chinese].