
ABSTRACT
Despite its importance for plant survival in cold biomes, little is known about the variation in the morphological traits of root systems of alpine plants. In this study, we investigated the difference in biomass allocation and morphological traits of belowground organs of Potentilla matsumurae between snowbeds and fellfields in an alpine tundra in northern Japan. The fellfields were characterized to have drier, colder, and less fertile conditions with more soil freeze–thaw cycles and longer growing seasons than snowbeds. The allocation to the aboveground part relative to the root system was significantly higher in the fellfields than in the snowbeds. The allocation to the root system relative to the aboveground part was significantly lower in the fellfields than in the snowbeds. For the morphological traits of fine roots, the specific root length and the root tip density were smaller in the fellfields than in the snowbeds. Together, distinct intraspecific differentiation of biomass allocation and morphological traits in the root system was found for P. matsumurae between the snowbed and fellfields populations in an alpine ecosystem. The distinct variation in root morphology could make P. matsumurae grow across the sharp gradient of snow conditions in alpine environments.
Introduction
The global climate has been warming over the past half century and is predicted to continue warming over this century (IPCC Citation2013). Alpine ecosystems are at the edges of climatic clines, and the vegetation mosaic and species diversity of plant communities are considered to be highly vulnerable to climate warming and coincidental drought (Kim and Donohue Citation2013; Amagai, Kudo, and Sato Citation2018). Plasticity and/or adaptation of functional traits are important to support the growth and survival of plants as well as to determine plant responses to climate change (Soudzilovskaia et al. Citation2013; Chiatante et al. Citation2015; Pérez-Harguindeguy et al. Citation2016). For instance, preferential biomass allocation to roots can maximize nutrient uptake with a limited amount of photosynthate obtained as a whole plant (Mokany, Raison, and Prokushkin Citation2006; Reich et al. Citation2014). Simultaneously, the larger biomass allocation to roots leads to the increased resistance of plants to predicted drought stress (Chiatante et al. Citation2015). In alpine ecosystems, because of the cold temperature, short growing season, and limited nutrient availability, the adaptive development of root systems is critical for their survival and sustainable growth (Körner Citation2003). Despite its importance, little is known about the regulatory mechanism of morphological traits of the root system in alpine plants.
In alpine ecosystems, a sharp gradient of snow accumulation is characteristic and creates distinct habitats: snowbeds and fellfields. Generally, snowbeds are characterized by more snow accumulation, a shorter growing season for alpine plants due to the later snowmelt, and higher nutrient and water availability than fellfields (Miller Citation1982). In fellfields, due to the small accumulation of snow cover that acts as an insulator, plants are exposed to frost, strong wind, and coincidental desiccation (McGraw and Antonovics Citation1983; Shimono et al. Citation2009).
Due to the harsh environments, plants inhabiting fellfields tend to have a compact stature, whereas plants inhabiting snowbeds with milder conditions tend to have larger aboveground parts (Shimono et al. Citation2009). Furthermore, it is known that the functional traits of aboveground parts can vary between fellfield and snowbed populations. For instance, leaf traits such as specific leaf area and nitrogen concentration are larger in snowbeds than in fellfields (Kudo, Nordenhall, and Molau Citation1999; Kudo, Molau, and Wada Citation2001). The large difference in flowering phenology between fellfield and snowbed populations results in the temporal isolation of mating opportunity and can even drive genetic divergence and coincidental trait variation between local populations (Shimono et al. Citation2009; Hirao et al. Citation2019). To the best of our knowledge, little is known about the relationship between the gradient of snow accumulation and plant root structure in alpine ecosystems, which makes it problematic to interpret the regulatory mechanisms to cope with a short growing season for the whole plant. Given the highest vulnerability of roots to cold temperature among all organs of a plant (Sakai and Larcher Citation1987) and cold temperature in the soil of alpine ecosystems, differentiation of root traits can occur along the sharp gradient of snow accumulation.
Severe frost in snowless fellfield habitat causes enormous mechanical tension to roots during the freeze–thaw cycles in soil, which are better resisted by coarse roots (>2 mm as defined by Pregitzer et al. Citation2002) in alpine ecosystems (Körner Citation2003). On the other hand, plants invest large amount of biomass in roots and develop thin fine roots with higher specific root length (SRL) to uptake soil nutrients efficiently in infertile habitats (Freschet et al. Citation2017). By considering the infertile soil in fellfields, we hypothesize that the fellfield population allocates increased biomass to roots, especially to fine roots with efficient morphology (e.g., high SRL), to uptake the limited soil resources as compared to the snowbed population (nutrient hypothesis). Meanwhile, as an alternate hypothesis, it is also possible that thick roots (especially coarse roots with low SRL) rather than thin fine roots develop in fellfields rather than in snowbeds to ensure resistance to frost damage in fellfields (frost hypothesis). According to optimality theory, the root traits are supposed to vary with the most limiting resource or environmental stress (Chapin Citation1980; Bloom, Chapin, and Mooney Citation1985; Reich et al. Citation2014), which allows us to predict that one of the hypotheses would be relevant. To test the hypotheses, we investigated the biomass allocation and root morphological traits in snowbed and fellfield habitats of Potentilla matsumurae Th. Wolf (Rosaceae) in an alpine ecosystem of northern Japan.
Materials and methods
Study design
This study was conducted in the central part of the Taisetsu Mountains (43°13′ to 43°45′ N, 142°32′ to 143°19′ E) in Hokkaido, northern Japan. Near Lake Hisago, one fellfield plot (1,910 m a.s.l.) and two snowbed plots (1,840 and 1,790 m a.s.l.; two were different in their distance to fellfields and thus snowmelt timing) were established along a snow accumulation gradient. In the two snowbed plots, snow usually disappears by early July (hereafter shallow snowbed) and late July (hereafter deep snowbed). The bedrock of the research site is nonalkaline mafic volcanic rocks. The two snowbed plots were separated by 250 m and 400 m respectively from the fellfield plot (details of the plots were provided in Shimono et al. Citation2009).
Abiotic factors
Following the previous studies on plant phenology in our site, we estimated the length of growing season based on the temperature regime combined with snowmelt time (e.g., Kudo Citation1992; Kudo et al. Citation2010). In 2016, the temperatures at 10 cm of soil (in fellfields) and those at the soil surface (in snowbeds) were recorded throughout the year ). Soil in the fellfields is frozen during the winter and plants cannot to start to grow until soil thaw because plants cannot uptake water from frozen soil even when aboveground temperature is above zero. Therefore, soil temperature is an available measure to estimate the starting point of the growing season in the fellfields. In snowbeds, soil does not freeze under thick snow cover. Overwintering buds locate at the soil surface and bud break does not occur until snowmelt. Thus, surface temperature is an available measure to estimate the starting point of the growing season in snowbeds. In fellfields, the date when the temperature reached above 0°C and remained positive in the following days was identified as the start of the growing season, and the date when the temperature reached below 0°C and remained negative in the following days was identified as the end of the growing season. In snowbeds, the first date when the surface temperature started to fluctuate can be recognized as the date of the snow disappearance (due to the lack of insulation by snow) and thus the start of the growing season (Shimono et al. Citation2009). Although there are alpine plants that have snow roots and can start root growth under snow (e.g., Onipchenko et al. Citation2014), our target species does not have these special roots and the aboveground part does not start under snow. Similarly, the final date when the surface temperature fluctuated can be recognized as the time when snow started to accumulate and as the end of the growing season.
In all plots, soil nitrogen availability was measured in the middle of the growing season (from the end of July to the beginning of September 2016) by employing the resin bag method (Binkley and Matson Citation1983; ). In this method, soil solution nitrogen in the rhizosphere was measured using nylon bags filled with 10 g of ion exchange resin (Amberlite MB-1; Organo, Japan; hereafter resin bags) buried at five locations in each plot to a depth of 10 cm below the soil surface (n = 5 per plot). Once harvested, resin beads were analyzed according to Makoto et al. (Citation2012). Ammonium (NH4+) and nitrate (NO3−) are two of the important forms of nitrogen available for tundra plants (Michelsen et al. Citation1996). The concentrations of NH4+ and NO3− in 2 M potassium chloride resin extract were determined by indophenol blue absorptiometry and by naphthyl ethylenediamine dihydrochloride spectrophotometry, respectively, using an autoanalyzer (AACS-4, BL-TEC, Inc., Japan). For soil moisture, we collected the soils sampled from 0 to 10 cm from the soil surface at five locations in each plot. From each location, about 100 g of the soil was sampled. After sieving the soil (with a 2-mm sieve), a few grams of the soil was dried at 105°C over three days and the gravimetric soil moisture was determined. The top 10 cm of the soil profile is the zone most relevant for root activity and nutrient cycling in alpine tundra (Schimel, Bilbrough, and Welker Citation2004; Makoto et al. Citation2016).
Biotic measurements
In each of the three plots, ten individuals of P. matsumurae were harvested at the end of the growing season (end of August 2017). For the excavation of roots, we carefully removed the surface litter and soil to avoid disturbing the root system as much as possible. The harvested samples were placed in a cooler box and transported carefully to the laboratory for further analysis.
The harvested individuals were gently washed and first separated into aboveground part and roots. The roots were further separated into coarse roots (diameter >2 mm) and fine roots (diameter <2 mm). Each part of the root system was scanned with an image scanner (GTS-630, Epson, Japan). Using ImageJ software, the length of the root systems of scanned images was measured (Abràmoff, Magalhães, and Ram Citation2004). Furthermore, we measured the number of root tips from the whole root system of each individual. After scanning, the organs were dried at 80°C for three days and the dry weight was measured. By using the dry weight of each part of the organs, we calculated the aboveground biomass, belowground biomass, shoot:root (S:R) ratio, coarse root biomass, fine root biomass, coarse root:fine root (C:F) ratio, SRL, and root tip density, which are important proxies for nutrient and water uptake by plants (Børja et al. Citation2008).
Statistical analysis
After verifying that the data frequency distributions did not deviate significantly from normality, we conducted a one-way analysis of variance (ANOVA), followed by Tukey’s honestly significant difference (HSD) multiple comparison tests. Data that did not follow a normal distribution were log-transformed and used for the analysis. The response variable was each of the plant traits (aboveground biomass, belowground biomass, S:R ratio, coarse root biomass, fine root biomass, C:F ratio, SRL, and root tip density) and the explanatory variable was the plot. In our study, we had three plots: fellfield, shallow snowbed, and deep snowbed. When the p value was less than .05, the difference in the trait among the plots was identified to be statistically significant. All statistical analyses were performed using R version 3.5.1 (R Development Core Team Citation2018).
Results
Environmental conditions
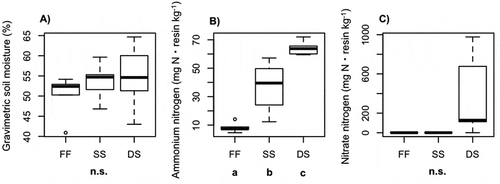
Based on the temperature measurement, the growing season was estimated to last from DOY (day of the year) 127 to DOY 295 in the fellfield, from DOY 189 to DOY 280 in the shallow snowbed, and from DOY 215 to DOY 280 in the deep snowbed (). The length of the growing season was 168 days, 91 days, and 65 days in the fellfield, shallow snowbed, and deep snowbed, respectively. During winter, while the soil was frozen in the fellfield, soil surface in the snowbed remained unfrozen (). For soil moisture, though there was an increasing trend of median with later snowmelt, there was no statistically significant difference among the three plots (ANOVA, p > .05; ). Soil NH4+ availability significantly varied among the three plots (ANOVA, F = 30.8, p < .001; ). Soil NH4+ availability significantly increased with later snowmelt (Tukey, p < .05; ). The ANOVA test revealed that there was a significant difference for soil NO3− availability among three plots (F = 10.2, p < .01; ). On the other hand, though there was an increasing trend of the median of NO3− availability with later snowmelt, the differences between any pair of plots were not statistically significant by multiple comparison test ().
Figure 1. The dynamics of soil temperatures in 2016. Thin black line shows the soil temperature at 10-cm depth in the fellfield, thick black line shows surface temperature in the shallow snowbed, and thin gray line shows surface temperature in the deep snowbed, respectively
Figure 2. (A) Gravimetric soil moisture, (B) soil ammonium availability, and (C) soil nitrate availability in surface soil. Inorganic nitrogen availability was determined with the resin bag method (Binkley and Matson Citation1983). FF = fellfield, SS = shallow snowbed, DS = deep snowbed. Different letters shown under the habitat name indicate statistical difference by Tukey’s HSD multiple comparison test (p < .05). n.s. means not significant
Plant traits
Aboveground biomass was significantly different among the populations (ANOVA, F = 16.9, p < .001). Aboveground biomass was significantly higher in the shallow snowbed than in the fellfield and the deep snowbed (Tukey, p < .05; and 4). On the other hand, there was no significant difference in aboveground biomass between the deep snowbed and the fellfield ( and ). Similarly, belowground biomass was significantly different among three populations (ANOVA, F = 33.2, p < .001). Root biomass was highest in the shallow snowbed (mean = 1.00 ± 0.03 SE) among the three populations (Tukey, p < .05), whereas there was no significant difference between the deep snowbed (0.16 ± 0.02) and the fellfield (0.19 ± 0.03; and ). The S:R ratio significantly differed among the three populations (ANOVA, F = 14.5, p < .001). Interestingly, the S:R ratio was significantly higher in the fellfield population than in the snowbed population (Tukey, p < .05; ).
Figure 3. Differentiation of functional traits among fellfield (FF), shallow snowbed (SS), and deep snowbed (DS): (A) aboveground biomass, (B) belowground biomass, (C) aboveground:belowground (S:R) ratio, (D) coarse root biomass, (E) fine root biomass, (F) coarse root:fine root (C:F) ratio, (G) SRL, and (H) root tip density. Different letters shown under the habitat name indicate statistical difference by Tukey’s HSD multiple comparison test (p < .05). n.s. means not significant
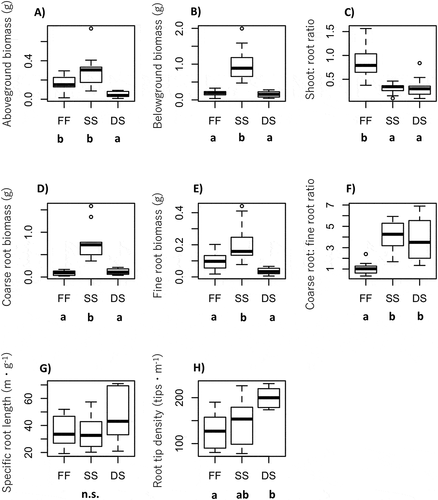
Figure 4. The representative individuals of Potentilla matsumurae in fellfield (FF), shallow snowbed (SS), and deep snowbed (DS)

For coarse root biomass, there was a significant difference among the three populations (F = 39.0, p < .001; and ). The shallow snowbed showed significantly higher values than the deep snowbed and the fellfield, whereas there was no significant difference in coarse root biomass between the deep snowbed and the fellfield (Tukey, p < .05; and ). Similar patterns with the highest root biomass were also found for fine roots in the shallow snowbed among the three populations (Tukey, p < .05; and ). The C:F ratio significantly differed among the three populations (ANOVA, F = 23.8, p < .001; ). Interestingly, the C:F ratio of the fellfield was significantly lower than that in the two snowbeds (Tukey, p < .05; ).
The morphological traits of fine roots differed partly among the three populations. Though there was no significant difference in SRL among the three populations, SRL tended to increase with later snowmelt (ANOVA, F = 1.9, p = .1; ). The root tip density differed significantly among the three populations (ANOVA, F = 10.6, p < .001; ). Root tip density was significantly higher in the deep snowbed than in the fellfield and the shallow snowbed (Tukey, p < .05;
Discussion
The allocation pattern and morphological traits of the root system differed significantly between the snowbed and fellfield populations of P. matsumurae. To the best of our knowledge, this is the first evidence showing the intraspecific variation in root structure of alpine plants with reference to the differences in snow accumulation and snowmelt time in alpine ecosystems.
Difference between fellfield and snowbed
Despite the infertile soil conditions, a small investment in root biomass and low root tip density were observed in the fellfield compared to the fertile snowbed (, , ). In addition, preferential allocation to fine roots relative to coarse roots was observed in the fellfield (). These results indicate that, partly consistent with the nutrient hypothesis, P. matsumurae increased the allocation to fine roots to deal with soil nutrient limitation in the fellfield habitat (consistent with our nutrient hypothesis) but changed other traits (root biomass and root tip density) in response to other environmental factors. An increase in fine root biomass relative to coarse root biomass can multiply the amount of root surface and thus their capacity for water and nutrient uptake per certain amount of biomass (Eissenstat Citation1992; McCormack et al. Citation2015). This pattern is especially beneficial in cold environments (Freschet et al. Citation2017) and in the soil of infertile tundra environments (Körner and Renhardt Citation1987), such as in alpine fellfield. Meanwhile, the low root tip density (and moderate SRL) in the fellfield (, 3H) did not support our nutrient hypothesis. Under low soil fertility conditions, plants would benefit more from having traits favoring resource conservation (e.g., low SRL) rather than a fast return on investment (Freschet et al. Citation2017). The increase in the production of fine root biomass might compensate for the low root tip density in the fellfield. Our results indicate that P. matsumurae develops its fine root morphology efficiently to uptake abundant nutrients by attaining a fast return from the investment. Although the instantaneous value of soil nutrient availability is generally small in fellfields (Miller Citation1982; and ), the integral amount of nutrient uptake over the growing season could increase due to the longer growing season in fellfields compared to snowbeds.
We believed that more coarse roots were present in the fellfields due to their high resistance to freeze–thaw cycles (). However, our results were inconsistent with our frost hypothesis. Whereas coarse roots are more resistant to the frost damage than fine roots (Körner Citation2003), coarse roots can also act as important stores of carbohydrates and nitrogen, which allow the rapid growth of plants in early spring to be competitive in the plant community (Chapin, McKendrick, and Johnson Citation1986; Kleijn, Treier, and Müller-Schärer Citation2005). In fact, at the same site as our study, Shimono et al. (Citation2009) reported faster aboveground growth of P. matsumurae in the snowbed than in the fellfield after snowmelt. The fast growth of the aboveground part of P. matsumurae results in high competitiveness for light interception in the snowbed, where vegetation in snowbeds commonly has larger biomass, higher primary productivity, and greater plant cover in the aboveground parts than that in the fellfields (Miller Citation1982). However, the underlying mechanism of high competitiveness in the snowbed populations of P. matsumurae is not fully understood. Based on our findings, it can be suggested that the high competitiveness of snowbed populations is attained partly due to the high biomass allocation to coarse roots as a storage function. Furthermore, the longevity of coarse roots is longer than that of fine roots (Wells and Eissenstat Citation2001). Given the limited amount of carbon fixation during the short growing season in snowbeds, snowbed plants might develop coarse roots not only for carbohydrate storage but also for increasing the longevity of roots to reduce the loss of carbon assimilated during the short growing season.
Variation within the snowbed
Even within the snowbeds, some root traits were substantially different between the shallow and deep populations. Although the allocation patterns between shoots and roots and between coarse roots and fine roots were similar between the two snowbeds, the aboveground biomass, coarse root biomass, fine root biomass, and belowground biomass (the sum of coarse root biomass and fine root biomass) were all smaller in the deep snowbed than in the shallow snowbed (). The overall small biomass in the deep snowbed could be due to the limited amount of carbon fixation and light availability during the short growing season. The difference in the length of growing season was twenty-six days between the shallow snowbed and the deep snowbed (). A twenty-six-day difference in the middle of summer could be a substantial length to cause carbon limitation for the plants. On the other hand, we observed a strategic change in the morphology of fine roots between the snowbed populations. The increasing root tip density in the deep snowbed corresponded to the increasing soil nutrient availability in the middle of the growing season (). A high root tip density and SRL may enable the efficient of temporally limited nutrients to support plant life in the deep snowbeds.
Implication for climate change
Populations that are locally adapted to alpine conditions are expected to be at especially high risk of extinction due to environmental changes (Holt Citation1990; Kim and Donohue Citation2013). Such risk can be decreased if specific traits of local populations exhibit some advantages under predicted climate change or phenotypic plasticity to altered climatic factors (Kim and Donohue Citation2013). Increased allocation to the root system relative to the aboveground system is assumed to be adaptive under warm and dry climate conditions (Chiatante et al. Citation2015), which is predicted worldwide under global change (e.g., Liu et al. Citation2018). In northern Japan, snow depth has been decreasing in the past ninety years (Makoto et al. Citation2014). Given that the small S:R ratio in the fellfield habitat is not a phenotypic but a genotypic difference, fellfield populations with small root systems are predicted to be highly sensitive to climate change. One study reported that the variation in S:R ratio of a herbaceous species in Japan (East Asian wild radish) is caused by a genetic divergence along with temperature gradient (Ishizuka et al. Citation2020), though this is not the case of alpine plants along with a snowmelt gradient. To predict the consequence of climate change, it is necessary to distinguish the driver of phenotypic variations in P. matsumurae by conducting a transplant experiment together with the combined analysis of genetic structure and climate manipulation (e.g., Kudo et al. Citation2010). Importantly, Blume-Werry, Jansson, and Milbau (Citation2017) found that though the growth of aboveground plant parts responded to the manipulation of snowmelt timing in arctic tundra, that of fine roots did not. This could imply that the importance of phenotypic plasticity for trait variations, which was observed between habitats with different snowmelt timing within the same species, differs between the aboveground and belowground organs of tundra plants.
Conclusion
Distinct intraspecific differentiation of biomass allocation and morphological traits in the root system was found between the snowbed and fellfield populations of P. matsumurae in an alpine ecosystem. P. matsumurae is one of the few species that is distributed from fellfield to deep snowbeds in our system (Kudo et al. Citation2010). The distinct morphological adaptation of the root system could make it possible for this species to grow across the sharp gradient of snow conditions in alpine ecosystems. To elucidate the adaptive significance of root traits in an alpine ecosystem, we should consider the relationship between the amount of available nutrients and fine root traits across the whole growing season in future studies.
Acknowledgments
We thank Dr. Tetsuo Kohyama and Kota Onizawa for their support during the fieldwork and Dr. Pamela H. Templer for information on the resin bag method. We also thank Juthi Rani Mitra and Hiroko Oiwa for their support for the root image analysis.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Abràmoff, M. D., P. J. Magalhães, and S. J. Ram. 2004. Image processing with imageJ. Biophotonics International 11:36–41. doi:https://doi.org/10.1201/9781420005615.ax4.
- Amagai, Y., G. Kudo, and K. Sato. 2018. Changes in alpine plant communities under climate change: Dynamics of snow-meadow vegetation in northern Japan over the last 40 years. Applied Vegetation Science 2018:1–11. doi:https://doi.org/10.1111/avsc.12387.
- Binkley, D., and P. Matson. 1983. Ion exchange resin bag method for assessing forest soil nitrogen availability. Soil Science Society of America Journal 47 (5):1050–52. doi:https://doi.org/10.2136/sssaj1983.03615995004700050045x.
- Bloom, A. J., F. S. Chapin III, and H. A. Mooney. 1985. Resource limitation in plants-an economic analogy. Annual Review of Ecology and Systematics 16 (1):363–92. doi:https://doi.org/10.1146/annurev.es.16.110185.002051.
- Blume-Werry, G., R. Jansson, and A. Milbau. 2017. Root phenology unresponsive to earlier snowmelt despite advanced above-ground phenology in two subarctic plant communities. Functional Ecology 31 (7):1493–1502. doi:https://doi.org/10.1111/1365-2435.12853.
- Børja, I., H. A. De Wit, A. Steffenrem, and H. Majdi. 2008. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiology 28 (5):773–84. doi:https://doi.org/10.1093/treephys/28.5.773.
- Chapin, F. S. 1980. The mineral nutrition of wild plants. Annual Review of Ecology and Systematics 11 (1):233–60. doi:https://doi.org/10.1146/annurev.es.11.110180.001313.
- Chapin, F. S., III, J. D. McKendrick, and D. A. Johnson. 1986. Seasonal changes in carbon fractions in Alaskan tundra plants of differing growth form: Implications for herbivory. Journal of Ecology 74 (3):707–31. doi:https://doi.org/10.2307/2260393.
- Chiatante, D., R. Tognetti, G. S. Scippa, T. Congiu, B. Baesso, M. Terzaghi, and A. Montagnoli. 2015. Interspecific variation in functional traits of oak seedlings (Quercus ilex, Quercus trojana, Quercus virgiliana) grown under artificial drought and fire conditions. Journal of Plant Research 128 (4):595–611. doi:https://doi.org/10.1007/s10265-015-0729-4.
- Eissenstat, D. M. 1992. Costs and benefits of constructing roots of small diameter. Journal of Plant Nutrition 15 (6–7):763–82. doi:https://doi.org/10.1080/01904169209364361.
- Freschet, G. T., O. J. Valverde-Barrantes, C. M. Tucker, J. M. Craine, M. L. McCormack, C. Violle, F. Fort, C. B. Blackwood, K. R. Urban-Mead, C. M. Iversen, et al. 2017. Climate, soil and plant functional types as drivers of global fine-root trait variation. Journal of Ecology 105 (5):1182–96. doi:https://doi.org/10.1111/1365-2745.12769.
- Hirao A. S., Y. Shimono, K. Narita, N. Wada, and G. Kudo. 2019. Ecotypic divergences of the alpine herb Potentilla matsumurae adapted to fellfield-snowbed habitats across a series of mountain sky islands. American Journal of Botany 106: 772–787. doi:https://doi.org/10.1002/ajb2.2019.106.issue-6. 6
- Holt, R. D. 1990. The microevolutionary consequences of climate change. Trends in Ecology and Evolution 5 (9):311–15. doi:https://doi.org/10.1016/0169-5347(90)90088-U.
- IPCC. 2013. Climate Change 2013: The Physical Science Basis; Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. New York: Cambridge University Press. 1535 pp.
- Ishizuka, W., K. Hikosaka, M. Ito, and S.-I. Morinaga. 2020. Temperature-related cline in the root mass fraction in East Asian wild radish along the Japanese archipelago. Breeding Science 70 (3):321–30. doi:https://doi.org/10.1270/jsbbs.18201.
- Kim, E., and K. Donohue. 2013. Local adaptation and plasticity of Erysimum capitatum to altitude: Its implications for responses to climate change. Journal of Ecology 101 (3):796–805. doi:https://doi.org/10.1111/1365-2745.12077.
- Kleijn, D., U. A. Treier, and H. Müller-Schärer. 2005. The importance of nitrogen and carbohydrate storage for plant growth of the alpine herb Veratrum album. New Phytologist 166 (2):565–75. doi:https://doi.org/10.1111/j.1469-8137.2005.01321.x.
- Körner, C. 2003. Alpine plant life: Functional plant ecology of high mountain ecosystems. 2nd ed. Berlin Heidelberg: Springer.
- Körner, C., and U. Renhardt. 1987. Dry matter partitioning and root length/leaf area ratios in herbaceous perennial plants with diverse altitudinal distribution. Oecologia 74 (3):411–18. doi:https://doi.org/10.1007/BF00378938.
- Kudo, G. 1992. Effect of snow-free duration on leaf life-span of four alpine plant species. Canadian Journal of Botany 70 (8):1684–88. doi:https://doi.org/10.1139/b92-207.
- Kudo, G., U. Molau, and N. Wada. 2001. Leaf-trait variation of tundra plants along a climatic gradient: An integration of responses in evergreen and deciduous species. Arctic, Antarctic and Alpine Research 33 (2):181–90. doi:https://doi.org/10.1080/15230430.2001.12003421.
- Kudo, G., U. Nordenhall, and U. Molau. 1999. Effects of snow-melt timing on leaf traits, leaf production and shoot growth of alpine plants: Comparisons along a snow-melt gradient in northern Sweden. Ecoscience 6 (3):439–50. doi:https://doi.org/10.1080/11956860.1999.11682543.
- Kudo, G., M. Kimura, T. Kasagi, Y. Kawai, and A. S. Hirao. 2010. Habitat-specific responses of alpine plants to climatic amelioration: Comparison of fellfield to snowbed communities. Arctic, Antarctic, and Alpine Research 42 (4):438–48. doi:https://doi.org/10.1657/1938-4246-42.4.438.
- Liu, H., Z. Mi, L. Lin, Y. Wang, Z. Zhang, F. Zhang, H. Wang, L. Liu, B. Zhu, G. Cao, et al. 2018. Shifting plant species composition in response to climate change stabilizes grassland primary production. Proceedings of the National Academy of Sciences of the United States of America 115 (16):4051–56. doi:https://doi.org/10.1073/pnas.1700299114.
- Makoto, K., H. Shibata, Y. S. Kim, T. Satomura, K. Takagi, M. Nomura, F. Satoh, and T. Koike. 2012. Contribution of charcoal to short-term nutrient dynamics after surface fire in the humus layer of a dwarf bamboo-dominated forest. Biology and Fertility of Soils 48 (5):569–77. doi:https://doi.org/10.1007/s00374-011-0657-y.
- Makoto, K., S. V. Bryanin, V. V. Lisovsky, K. Kushida, and N. Wada. 2016. Dwarf pine invasion in an alpine tundra of discontinuous permafrost area: Effects on fine root and soil carbon dynamics. Trees - Structure and Function 30: 431–439. doi:https://doi.org/10.1007/s00468-015-1192-5.
- Makoto, K., T. Kajimoto, L. Koyama, G. Kudo, H. Shibata, Y. Yanai, and J. H. C. Cornelissen. 2014. Winter climate change in plant – soil systems: Summary of recent findings and future perspectives. Ecological Research 29 (4):593–606. doi:https://doi.org/10.1007/s11284-013-1115-0.
- McCormack, M. L., I. A. Dickie, D. M. Eissenstat, T. J. Fahey, C. W. Fernandez, D. Guo, H.-S. Helmisaari, E. A. Hobbie, C. M. Iversen, R. B. Jackson, et al. 2015. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytologist 207 (3):505–18. doi:https://doi.org/10.1111/nph.13363.
- McGraw, J., and J. Antonovics. 1983. Experimental ecology of Dryas octopetala ecotypes: I. Ecotypic differentiation and life-cycle stages of selection. Journal of Ecology 71 (3):879–97. doi:https://doi.org/10.2307/2259599.
- Michelsen, A., I. K. Schmidt, S. Jonasson, C. Quarmby, and D. Sleep. 1996. Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia 105 (1):53–63. doi:https://doi.org/10.1007/BF00328791.
- Miller, P. C. 1982. Environmental and vegetational variation across a snow accumulation area in montane tundra in central Alaska. Ecography 5 (2):85–98. doi:https://doi.org/10.1111/j.1600-0587.1982.tb01022.x.
- Mokany, K., R. J. Raison, and A. S. Prokushkin. 2006. Critical analysis of root : shoot ratios in terrestrial biomes. Global Change Biology 12 (1):84–96. doi:https://doi.org/10.1111/j.1365-2486.2005.001043.x.
- Onipchenko, V. G., A. M. Kipkeev, M. I. Makarov, A. D. Kozhevnikova, V. B. Ivanov, N. A. Soudzilovskaia, D. K. Tekeev, F. S. Salpagarova, M. J. A. Werger, J. H. C. Cornelissen, et al. 2014. Digging deep to open the white black box of snow root phenology. Ecological Research 29 (4):529–534. doi:https://doi.org/10.1007/s11284-013-1112-3.
- Pérez-Harguindeguy, N., S. Díaz, E. Garnier, S. Lavorel, H. Poorter, P. Jaureguiberry, M. S. Bret-Harte, W. K. Cornwell, J. M. Craine, D. E. Gurvich, et al. 2016. Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany 64 (8):715. doi:https://doi.org/10.1071/bt12225_co.
- Pregitzer, K. S., J. L. DeForest, A. J. Burton, M. F. Allen, R. W. Ruess, and R. L. Hendrick. 2002. Fine root architecture of nine North American trees. Ecological Monographs 72 (2):293. doi:https://doi.org/10.2307/3100029.
- R Development Core Team. 2018. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
- Reich, P. B., Y. Luo, J. B. Bradford, H. Poorter, C. H. Perry, and J. Oleksyn. 2014. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. Proceedings of the National Academy of Sciences of the United States of America 111 (38):13721–26. doi:https://doi.org/10.1073/pnas.1216053111.
- Sakai, A., and W. Larcher. 1987. Frost survival of plants. Berlin, Heidelberg: Springer.
- Schimel, J. P., C. Bilbrough, and J. M. Welker. 2004. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biology and Biochemistry 36 (2):217–27. doi:https://doi.org/10.1016/j.soilbio.2003.09.008.
- Shimono, Y., M. Watanabe, A. S. Hirao, N. Wada, and G. Kudo. 2009. Morphological and genetic variations of Potentilla matsumurae (Rosaceae) between fellfield and snowbed populations. American Journal of Botany 96 (4):728–37. doi:https://doi.org/10.3732/ajb.0800242.
- Soudzilovskaia, N. A., T. G. Elumeeva, V. G. Onipchenko, I. I. Shidakov, F. S. Salpagarova, A. B. Khubiev, D. K. Tekeev, and J. H. C. Cornelisse. 2013. Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proceedings of the National Academy of Sciences of the United States of America 110:18180–84. doi:https://doi.org/10.1073/pnas.1310700110.
- Wells, C. E., and D. M. Eissenstat. 2001. Marked differences in survivorship among apple roots of different diameters. Ecology 82: 882–892.