
ABSTRACT
A complex landscape lies on an alpine plateau in the high Sierra Nevada, California, and comprises spatially organized physical and ecological features that interact to create a unique ecosystem. At the upslope end a tree-limited ribbon forest of whitebark pines grows on the crest of a short, steep slope, which is the only deformation on the otherwise flat plateau. A long-persistent snowbank forms on the slope in the lee of the ribbon forest; meltwater provides moisture to support a productive wet meadow just below. Below the meadow is a narrow, linear “rampart” comprising large sorted stones, and below this lies a large patterned-ground area of cryogenically sorted circles. The soil domains are densely covered with species-rich vegetation and are abruptly segregated from stone domains. The latter serve as distributed springheads in the otherwise dry patterned-ground terrain. American pikas have colonized the rocky nets of the stone domains and forage plants of the soil circles. The unusual context of this landscape enables pika territories to be more tightly packed than in traditional habitat. We propose a scenario for the Pleistocene origin of this ecosystem and a unique lithologic subsurface barrier that contributes to spring formation during warm intervals.
Introduction
Spatially organized landforms occur in many environments and climate zones. Patterned ground of periglacial origin is common in cold polar regions, where features occur in many forms, shapes, and sizes (Washburn Citation1956; Van Vliet-Lanoë Citation2014). Sorted circles are a type of patterned ground (PG; Krantz, Gleason, and Caine Citation1988). At the ground surface these appear as circular to polygonal in shape and are characterized by distinct segregation (sorting) of grain size. Commonly, fine-grained sediments dominate the center of the circles (soil domain) and larger clasts form a border around the circles (stone domain; Washburn Citation1956). Sorted circles typically develop on flat to shallow slopes with low relief. In their mature forms circles can attain diameters of 0.8 m to > 5 m; small features (<0.5 m in diameter) usually indicate nascent forms (Wilkerson Citation1995). Active features—that is those that are ongoing in formation—may lack vegetation in the soil circles and lack lichens on stones of the borders.
Washburn (Citation1956) evaluated nineteen hypotheses to explain the formation of PG, including sorted circles. Common features for development include the presence of a cold climate where repeat freezing and thawing occurs and permafrost is present. Aside from these, Washburn concluded that causality remained far from understood and multiple paths of origin were the rule, not the exception. Although significant progress has been made toward understanding how these forms arise, many specific aspects remain enigmatic (Hallet Citation2013). Unifying theories of self-organization within a freeze–thaw (F-T) environment were proposed for PG formation nearly half a century after Washburn’s pioneering work. Proposals for underlying processes that drive formation of PG, however, remain numerous and include particle sorting, F-T, deformation of frozen soil, soil creep, free convection, and combinations of these dynamics (Ray et al. Citation1983; Krantz Citation1990; Kessler and Werner Citation2003).
Whereas PG features are best known and studied from polar regions, they also occur in temperate environments, where permafrost is not considered essential for formation (Washburn Citation1956; Krantz, Gleason, and Caine Citation1988; Wilkerson and Schmid Citation2016). Climate conditions conducive to initiation and development of PG in these environments include cold winters, relatively warm summers that allow for many freeze‐thaw cycles, and enough precipitation to allow ground ice to develop but not so much that glaciers develop (Wilkerson and Schmid Citation2016). PG features found in mountains of temperate western North American mountains are mostly considered inactive and relicts of past cold climates (Krantz, Gleason, and Caine Citation1988). Active forms have been described for only a few very high, generally dry ranges, including the Wind River Range, Wyoming (Billings and Mooney Citation1959), Absaroka Range, Wyoming, Sangre de Cristo Mountains, Colorado (Krantz Citation1990), Rocky Mountain Front Range, Colorado (J. B. Benedict Citation1992), and the White Mountains, California (Mitchell, Lamarche, and Lloyd Citation1966; Wilkerson Citation1995).
The Sierra Nevada, California, is a relatively low-latitude, warm-temperate mountain range with a climate dominated by winter Pacific Ocean precipitation and Mediterranean-type summers. Remaining glaciers are small and few (Basagic and Fountain Citation2011), although active as well as relict rock glaciers are common (Millar and Westfall Citation2019). PG was described and mapped at high elevations as part of a survey of rock glaciers and related periglacial features in the central part of the range (Millar and Westfall Citation2008). In that survey, a small number of sorted circles were included in the PG class; those occurred on high, exposed, often windswept, snow-free plateaus and benches. Some PG features occurred along margins of alpine lakes in positions that were assumed to be submerged during summer and exposed in autumn. Aside from this limited survey, we know of no studies of sorted circles or related PG in the Sierra Nevada.
Cryogenic PG of cold regions is not the only context where spatially ordered, repeating landforms develop. Large-scale, recurring patterns of dominantly ecological origin occur at different levels of biological organization and in environments from tropical to polar (Rietkerk and Van De Koppel Citation2008; Tarnita et al. Citation2017). Examples range from dotted landscapes of soil mounds, such as the Mima Mounds of western Washington (Cramer and Barger Citation2014) to recurring patterns of termite “fairy circles” in Kenya, Zambia, and other hot desert regions (Bonachela et al. Citation2015). In high-temperate mountains, spatially patterned vegetation can be associated with periglacial, spatially patterned features (Mitchell, Lamarche, and Lloyd Citation1966; Millar et al. Citation2015; Apple, Ricketts, and Martin Citation2019) or with other aspects of mountain topography (Björk and Molau Citation2007).
Two types of patterned vegetation in high mountains not associated with periglacial PG include ribbon forests and snowbed plant communities (Holtmeier Citation2003; Björk and Molau Citation2007). Ribbon forests are linear strips of trees found in high-elevation mountain environments. These occur in several patterns, all strongly influenced by wind (Bekker and Malanson Citation2008). Some ribbon forests are oriented parallel to wind, and others develop perpendicular to the wind. The latter are little studied and their origins least understood (Bekker and Malanson Citation2008). Deep snowdrifts accumulate in the lee of ribbon forests, which can persist longer into summer than snow on adjacent slopes. This leads to the establishment of unique snowbed plant communities that are adapted to short growing seasons and saturated, poorly developed soils (Tomaselli Citation1991; Björk and Molau Citation2007). Other than a brief account of snow-dependent plants in the Sierra Nevada, included as part of a vegetation classification project for western North America (Peinado et al. Citation2005), the occurrence and ecological role of ribbon forests and snowbeds have not been studied in the Sierra Nevada.
A complex PG landscape occurs on Kavanaugh Plateau in the high Sierra Nevada, California. A portion of the plateau supports patterned features of physical and ecological nature that appear to arise as an interconnected ecosystem. Features of the ecosystem include a ribbon forest, snowbed slope, wet meadow forefield, rock-terrace rampart, and a large area of PG sorted circles that take on diverse shapes and sizes and support distinct vegetation communities. An unusual feature of this landscape is the occurrence of springs distributed across the PG network, characterized by water emerging from rock wells that border the soil circles and flowing a short distance onto adjacent soil circles. A further unexpected component is a population of American pikas (Ochotona princeps) that uses the flat PG ecosystem as habitat. Pikas are small herbivorous mammals related to rabbits that range across mountains of western North America (A. T. Smith and Weston Citation1990). Adapted to cold climates, pikas are poor thermoregulators and have been widely studied for their sensitivity to warming (A. S. Smith Citation2020). Pikas are restricted in habitat type and live only within talus and related broken-rock landforms.
Our initial objective for this study was to better understand the use of PG on the Kavanaugh Plateau by pikas. As we pursued questions related to pikas, however, it became clear that the geomorphology and broader ecology of the site were complex and novel, especially for a warm-temperate range such as the Sierra Nevada. Because the plateau environment has elements that appear to integrate cryogenic processes of PG and biogenic processes of spatial patterning, and exhibits features and processes unique to this site, we expanded our goals to include the larger system. Our guiding study questions were the following:
What are the primary components of the Kavanaugh Plateau PG ecosystem, including forms and processes that appear to be cryogenic (e.g., sorted circles, rock nets and wells), biogenic (e.g., ribbon forest, snowbed, forefield meadow, vegetation and plant cover), and novel situations to this location (rock-terrace rampart, dispersed springheads, overland flow, and pika use of PG)?
How do hydrologic and thermal conditions of the Kavanaugh Plateau ecosystem relate to previously described cryogenic processes for PG in colder environments?
As pika habitat, how do physical, thermal, and ecological conditions of the PG ecosystem compare to pika environments elsewhere?
Armed with insight from these individual elements, we propose an integrated scenario for the origin and functioning of the Kavanaugh PG ecosystem.
Methods
Study area
Kavanaugh Plateau is a broad shallow slope situated between Dunderberg Peak (3,772 m) and the rocky summits of Kavanaugh Ridge (high point, 3,426 m) in the eastern Sierra Nevada, California (). Taken together these uplands extend 6 km from Virginia Canyon in the south to Green Creek Canyon in the north. Kavanaugh Plateau (~0.9 km2) slopes to the northeast from an abrupt southwest edge (~3,350 m), where an escarpment drops to the East Fork Green Creek >600 m below. The south edge of the plateau extends to the low morainal ridge of a Pleistocene glacier that flowed downslope from one of several cirques under Dunderberg Peak (Wahrhaftig et al. Citation2019); rocky slopes of Kavanaugh Ridge define the north edge. Surface material of the Kavanaugh Plateau is composed of fragmented Paleozoic metasedimentary rocks, most likely derived as slopewash or glacial outwash till from adjacent Dunderberg Peak moraines (pers. comm., Greg Stock, National Park Service, 6 November 2020), and Kavanaugh Ridge to the north comprises Cretaceous-age granitic substrates (Huber, Bateman, and Wahrhaftig Citation1989). The plateau and ridge were unglaciated during the late Pleistocene (Wahrhaftig et al. Citation2019).
Figure 1. Map of the central Sierra Nevada, California, with location of the Kavanaugh Plateau and patterned-ground study site east of the range crest in Mono County.
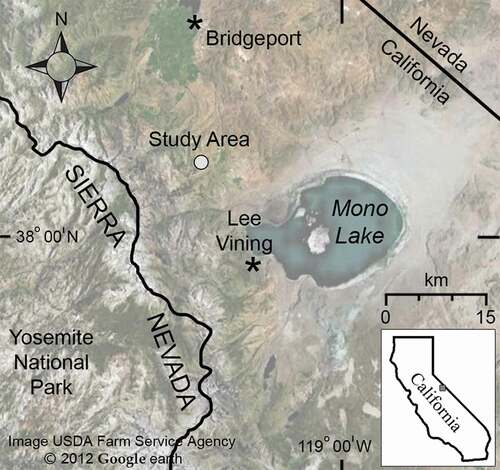
Figure 2. Overview of Kavanaugh Plateau and Dunderberg Peak. The patterned-ground study site (polygon) lies on the shallow slope in the middle of Kavanaugh Plateau and is surrounded by regions of higher relief. (a) View north toward Kavanaugh Ridge (3,389 m). (b) View south–southwest to Dunderberg Peak (3,772 m). Taluses on adjacent slopes that were monitored for pika activity are noted with asterisks.
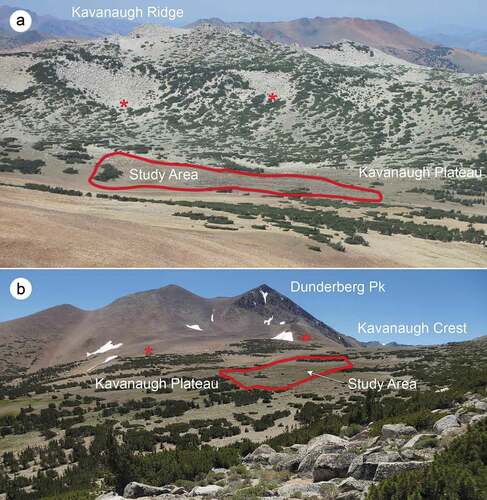
The PG ecosystem, which defines our study area, lies in the center of Kavanaugh Plateau (mean elevation 3,265 m; ; additional photos, Supplemental Material (SM) )). The 7 ha roughly rectangular area extends eastward from a linear strip (ribbon) of forest composed of densely packed, short whitebark pines (Pinus albicaulis) across gently sloping ground that comprises discrete environmental units, or zones. To characterize the features of the study area we delineated zones () as follows: ribbon forest (Zone 1, ); snowbed slope, which supports a long-lasting rectangular snowdrift (Zone 2, ); forefield meadow (Zone 3, ); rock-terrace rampart (Zone A; ); and three zones of PG (Zones B, C, and D, 2 ha total; ). The PG zones comprise sorted features of circular to ellipsoidal shapes, with soil domains in the center and densely packed stone-domain nets partially or fully around them. Many of the rock wells comprising the stone domains are transiently occupied by springs (). The upslope PG zone (B) has unsorted as well as sorted circles, with dispersed and mostly discontinuous rock wells (). The middle and lower slope PG zones (C and D) contain highly sorted circles with continuous nets of rock borders and wells ().
Figure 3. Overview of Kavanaugh Plateau patterned-ground study area. Ribbon forest, snowbed slope with snowdrift, soil circles, and rock wells are visible in this westward view.

Figure 4. Zones delineated for physical and ecological measurements, Kavanaugh Plateau. Ribbon forest, snowbed slope, forefield meadow, terrace rampart, and patterned-ground zones are indicated.
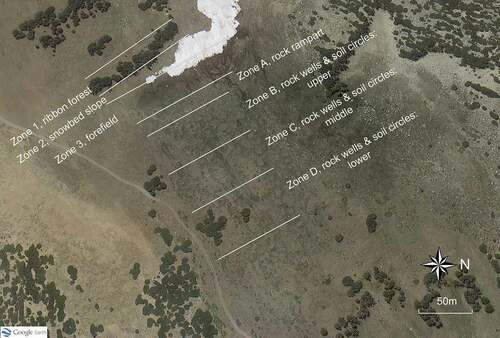
Figure 5. Features of the patterned-ground ecosystem within the Kavanaugh Plateau study area. (a) Edge of ribbon forest and snowbed slope. (b) Forefield meadow. (c) Rock terrace rampart. (d) Patterned ground with soil circles and rock wells. All views are southward.

Figure 6. Rock well springs in the patterned ground zones. Following spring snowmelt from most slopes of the Kavanaugh Plateau except the snowbed, multiple springs across the patterned-ground zones emerge from rock wells and flow a short distance onto soil circles below them. (a) View westward to the ribbon forest and snowpack remaining on the snowbed slope (14 July 2017). (b) View eastward of a spring in the lower portion of the patterned-ground zones showing the water flow onto downslope soil circle (3 July 2019).

Figure 7. Types of patterned ground in study area. (a) Unsorted circles in the upper portion of the study area. Rock wells are mostly discontinuous and do not always surround soil circles. View westward to the rock rampart, snowbed slope, and ribbon forest. (b) Sorted circles in the lower portion of the study area. Rock domains border the soils circles and are mostly continuous with other rock domains across the slope. View northward.

Figure 8. Temperatures from four positions in the Kavanaugh Plateau study site. (a) Air temperature recorded at 2.5 m in the ribbon forest. (b) Forefield meadow (buried). (c) Soil circle (buried). (d) Rock well springhead.
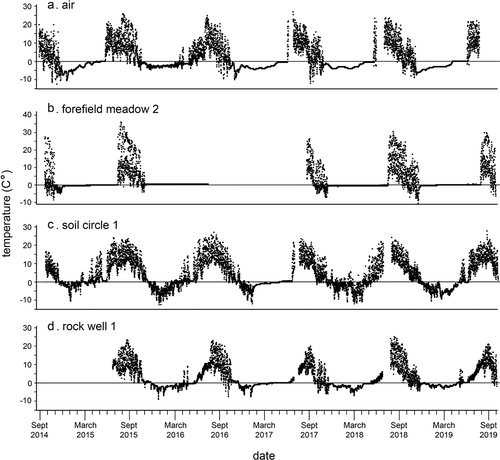
Figure 9. Temperatures from one Kavanaugh Plateau haypile site, September 2014 to September 2019. Missing data are either failures of datalogger or gaps in exchange of units. (a) Haypile surface. (b) Haypile matrix.
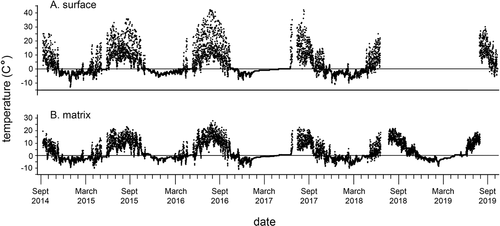
Figure 10. Fresh haypiles in rocky borders of sorted and unsorted circles, 21 September 2014. (a) Haypile in upper unsorted circle (Zone B). (b) Haypile in lower sorted circle (Zone D).

Figure 11. Location of fifty-three haypiles, including old and fresh, on the patterned-ground ecosystem. Cumulative across seven years of observations.

Figure 12. Number of haypiles observed on the patterned-ground zones from 2014 to 2020, freshness of vegetation in the haypiles, and relative size of fresh haypiles. Ranks: 0, old haypile; 0.5, one-year-old haypile; 1–5, fresh (green) haypiles by relative size, from 1 (smallest) to 5 (largest).
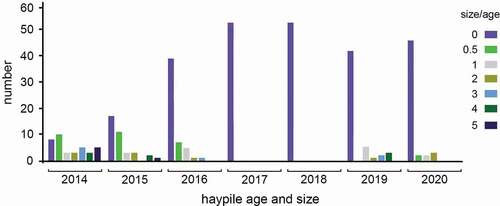
Figure 13. Cross-section views of the Kavanaugh Plateau showing stratification of substrates. A solid granitic formation dips under the unconsolidated metamorphic substrate of the patterned-ground slope in the central portion of the plateau. (a) Photo taken from a promontory on the edge of Kavanaugh Crest looking back (southeast) toward the plateau. (b) Google Earth image showing the shallow central slope with the substrate stratification (in circle; view northeast). The snowbed with snowdrift still present is in the middle of the slope and the patterned-ground ecosystem lies beyond (east) it at arrow. Google Earth image, 11 August 2017.

Other than the ribbon forest, only a few clusters of stunted trees (whitebark pine krummholz and low shrub juniper, Juniperus communis) occur in the study area. This situation exists although the plateau lies below regional whitebark–pine species line (sensu Körner Citation2012). Abundant krummholz stands are present on Kavanaugh Ridge to the north and at the south edge of the plateau beyond the study area (). Strong west and southwest winds regularly blast the flat area of the plateau and likely contribute to minimal tree presence.
Physical measurements
Structural components
Using Google Earth Pro (GEP), we estimated the slope inclination of each zone using the downslope width of the zone and the difference in elevation. We measured the inclination of the soil circles in the field with a carpenter’s level and clinometer and in the office with GEP. The field methods were more accurate and precise, given the low resolution of elevation change in GEP across short distances.
In each zone we measured rock (clast) size. For Zones 1 to 3 and A we opportunistically selected ten points across the north–south extent of the zone, and at each point we measured ten clasts on their long dimension. For each of the PG zones, we measured ten clasts from each of ten rock wells and from each of ten soil circles per zone. We selected the center points of soil circles and the deepest part (springhead) of rock wells to measure clast sizes.
In addition to clast size, in the PG zones we measured two dimensions of ten soil circles and ten rock wells. For soil circles we measured the longest dimension and the dimension orthogonal to it. We estimated the area of soil circles based on the formula for area of an ellipse; in many cases these converged on circular. Rock wells were difficult to standardize in their dimensions, given the variable range of shapes and their continuous and discontinuous nature. Where rock wells were discontinuous (i.e., did not surround the soil circles), they had a roughly triangular shape (). Thus, for both discontinuous and continuous () rock wells we measured the two longest dimensions and estimated their areas using the equation for the area of a triangle. Rock wells were regularly and conspicuously lower than the surface elevations of adjacent soil circles (; SM ). We estimated the depth of the well at its lowest point relative to the surface elevation of the adjacent soil circle.
Because of the unbalanced nature of the data for clast size samples, whereby Zones B through D had two features and the remaining zones only one, the full mixed model analysis of variance (ANOVA) could not be used. This model comprised the main effects of zones and features , the zone × feature interaction, plots nested in zones and features. Samples in plots was the error (residual) term. In this model, the main effects were not estimable. In this model, the main effects were not estimable. Instead, we used a fully hierarchical design where features were nested in zones and plots nested in zones and features. Least squares mean contrasts were used to test differences between certain groups of zones and features. For areas of soil circle and rock well features, we used a factorial model including the interaction, where zones and features were main effects and Zone × Features was the interaction; plots in zones and features were the error term. Differences in well depth were tested with a one-way ANOVA among zones. ANOVAs were conducted in JMP (v12.2; SAS Institute Citation2015).
Temperature
To assess temperature differences and snow/hydrologic conditions relevant to the PG processes and for pika habitat, we installed temperature dataloggers (Maxim iButtons, model DS1921G or DS1922L) across seven features of the Kavanaugh PG ecosystem. These included buried 10 cm in soil above (west of) the ribbon forest (three units distributed south to north); 2.5 m height in the ribbon forest (one unit on a tree in the middle of the zone); buried 10 cm in the soil of the forefield meadow (nine units in a grid pattern across the zone; SM Figure 14); buried 10 cm in the soil at the center of the soil circles (four units); surface of haypiles in rock wells (four units); rocky matrices below haypiles 15 to 20 cm deep in rock wells (four units); and springheads in 15 to 25 cm of water (when spring was flowing) in rock wells (four units). All units were sealed with silicon, secured in a wire mesh, and inserted into a 17-cm white PVC funnel. The PVC packages were tethered with wire and tied to either branch, rock, or a tent stake, depending on position. iButtons were installed in September 2014 and programmed to record at four-hour intervals, starting at noon on the day of deployment. All units were exchanged at least once annually, attempting to maintain continuous measurement, although gaps resulted, and were uninstalled in October 2019.
Measurements derived from the dataloggers included mean annual daily temperature (MAT), seasonal mean daily temperature, number of days below 0°C, number of F-T events (defined as the number of days/year when the daily minimum was <0°C and daily maximum temperature was >0°C), water flow and desiccation, and onset and melt dates of snow. We defined snow events to have occurred when ground temperatures dropped to near freezing and the diurnal range in temperature was less than 1°C (Lundquist and Lott Citation2008). Using this criterion, dates of onset of autumn/winter snow cover and release from spring snow cover for features of interest could be estimated from the temperature records. Similarly, spring/summer onset of water flow in rock wells and summer/autumn desiccation were estimated for seasonal dates when daily variance in temperatures increased from near zero but remained low with the onset of water flow and increasing greatly on desiccation. Changes in daily variance coincided with increases in maximum temperature (onset of water flow) and large increases in maximum temperature (desiccation).
To assess differences in mean daily temperatures we first conducted ANOVA with a nested design that included zones and features in zones (JMP; SAS Citation2015). We used paired tests for contrasts that tested differences in means between specific features of interest that were nonsignificant. The objective of the paired test is to determine whether daily temperatures between positions were consistently different; for example, whether daily mean summer surface temperatures were consistently greater than those in the matrices. However, we found that daily temperatures were highly autocorrelated and fit an autoregressive moving average (ARMA) process whereby the former reflects daily thermal momentum and the latter reflects synoptic weather patterns. This autocorrelation results in pseudoreplication, so the effective degrees of freedom are much less than the actual. We fit surface temperature and sometimes matrix temperature with an ARMA model (usually ARMA(2,2), representing the number of lagged years in the model) and then used the residuals of the fitted model as weights in the paired tests (JMP; SAS Citation2015).
Ecological measurements
Ribbon forest
To estimate age of trees in the ribbon forest, we selected five locations along the east-to-west length of the ribbon forest and established two 2-m-diameter plots at each location, one on the west side and one on the east side of the ribbon forest (ten plots total). Using standard dendrochronological methods, we retrieved a single increment core at 0.5 m height from two live trees per plot (twenty trees total). We sanded the cores and counted the number of rings from the bark to pith to estimate tree age in autumn 2020; we tested differences in ages of the trees at the west versus east edge using a two-tailed t-test. In each plot we also counted the number and diameter of pine stems and pine seedlings (<0.5 m height) and estimated percentage cover of pine vegetation.
Shrub and ground vegetation
Vascular plant species lists were compiled for the forefield meadow (Zone 2) and soil circles (Zones B–D) of the PG ecosystem and for talus forefields of occupied pika habitat on slopes adjacent to the study area (Jepson Flora Project [Citation2020] for taxonomic authority). Plant cover was estimated by ocular observation using 2-m-diameter circular plots (3.1 m2) with ten plots in the PG forefield meadow, eighteen plots in the soil circles, and ten plots in the adjacent talus forefields. Plots were located so they did not include any portion of rock wells or talus. In addition to live vegetation, cover of litter, bare ground, and rocks was estimated. Species were assigned plant functional traits (adapted from Kӧrner Citation2003; Millar et al. Citation2015) for life form, woodiness, and leaf form.
American pika activity
We surveyed for the presence of new and old pika haypiles (winter food caches) within the PG study area. We defined haypiles as fresh (current year) when vegetation was green or when drying leaves were obviously from the current summer’s harvest. Haypiles were scored as old (score 0) when they were composed of woody, leafless stems and/or obviously long-dead foliage. Fresh haypiles were also rated by size, from a few sprigs (score 1) to large mounds of vegetation (score 5). Haypiles were surveyed throughout summer and early autumn annually from 2014 to 2020.
Results
Physical measurements
Structural components
Despite an overall flat or gently sloping nature of the Kavanaugh Plateau, there were discrete changes in slope inclination within and just adjacent to the study area (). Above (southwest of) the ribbon forest the slope was much steeper (22 percent) than either the ribbon forest (8 percent) or PG zones (8 percent). Similarly, the slope declined below (northeast of) the PG zones (14 percent). The steepest slope in the area was the short snowbed slope (36 percent) below the ribbon forest. Despite the general slope of the PG zones, the soil circles were nearly flat (mean, 2.7 percent) and in the field appear tilted relative to the background ().
Table 1. Physical measurements
The PG features were significantly larger in the upper portion of the ecosystem (Zone B) than in the lower two zones (C, D; ). Soil circles in Zone B were more than twice as large, with mean areas estimated as 31.5 m2 versus 12.7 m2 for soil circles in Zones C and D (p < .001). Similarly, rock wells were significantly larger in the upper part of the ecosystem (Zone B), with areas estimated as 8.2 m2 versus 5.9 m2 for rock wells in Zones C and D (p < .001). Mean clast sizes were significantly larger (p < .0001; ) in the rock-terrace rampart (Zone A; 41.6 cm longest dimension, ) and rock wells (Zones B, C, D; 30.2 cm) than in the ribbon forest (3.6 cm) or soil circles (3.5 cm). Clasts in the rampart were also significantly larger than those in rock wells (all zones; p < .0001), and clast sizes in rock wells of Zone B (34.1 cm) were larger than those in Zones C and D (mean, 31.9 cm; p < .0001). At their lowest points, rock wells averaged 54.1 cm below the surfaces of adjacent soil circles.
Table 2. Significance values for select statistical tests of differences between class means of physical features in the study area. See for location of zones
Temperature
Mean annual air temperature (MAT), measured across five years at 2.0 m height in the ribbon forest, was 1.5°C, with mean winter temperature of −3.8°C and mean summer temperature of 8.8°C (, ). Differences among years in MAT were significant (p = .0074; ), with 2015 the warmest year (2.2°C) and 2017 the coldest (0.9°C). Spring and summer temperatures were especially important in contributing to differences among years. The low diurnal variance during winter might suggest that our loggers were covered by snow, as does the short “zero curtain” in late spring. Temperatures during most of the winter season, however, were far below freezing, suggesting that if there was snow on the logger, it was thin enough to allow thermal conduction. The north aspect of the loggers and their situation in dense shade would further attenuate daily air temperature fluctuations in winter. Prior experience with similar situations suggests that these temperatures are representative of winter means.
Table 3. Temperatures at six features of the Kavanaugh Plateau study area, 2014–2019. Mean annual daily temperatures and seasonal daily temperatures, with standard deviations, minima, and maxima
Table 4. Annual and seasonal air temperatures (2.0 m in ribbon forest) for 2015–2018. Data for 2014 and 2019 are incomplete due to duration of study
Soil temperatures differed significantly among the features monitored (, ), with the forefield meadow soils (MAT, 2.2°C) significantly cooler (p < .001) than MAT of the soil circles (3.5°C) and of the soils on the slope above the ribbon forest (3.6°C). In winter, soil circles (−3.1°C) were on average colder than soil in the forefield meadow (−2.2°C). In summer, the opposite was the case, with the soil circles (11.9°C) significantly warmer than the forefield meadow soils (7.5°C, p < .0001).
Mean annual temperature across five years at pika haypile surfaces in the PG was equal to the mean for rocky matrices under haypiles (both 2.7°C; , ). In paired tests, summer temperatures in matrices (10.6°C) were significantly cooler than surfaces (11.9°C, p < .001; , ), although by a small difference. For most of the plots and across the years of the study, MAT of haypile matrices (2.7°C) was significantly warmer than that of rock wells (2.2°C; p < .001), although differences of some pairs were not significant. Summer differences followed the same pattern, with mean haypile matrix temperatures (10.6°C) warmer than those of rock wells (9.4°C) in most cases.
Using changes in daily variance and temperatures relative to freezing (0°C), we were able to estimate annual dates (2014–2019) when snow first covered features and when snow melted (; ). Of particular interest were comparisons of the slope above the ribbon forest (beyond the PG ecosystem), the forefield meadow, and the soil circles. Snow-onset dates within a year were relatively similar across these positions, although there was variability across years, with 2016 being the winter of earliest onset and 2014 and 2018 being latest onset. Onset dates were variable by location in that early autumn snowfall (usually in early October) often melted quickly from some areas and not others. Relative order of snowmelt across the features was as follows: soil circles (earliest annual melt), slope above the ribbon forest, and forefield meadow (latest melt). This corroborates the late-lying snowdrift that extends below the snowbed slope onto the forefield meadow, with melt occurring earliest at the downslope (eastward) position and proceeding upward toward the ribbon forest. Relative order of spring snowmelt across the years varied slightly by position. For the slope above the ribbon forest, 2016 was earliest snowmelt and 2017 was latest; for soil circles, 2016 and 2018 were earliest and 2017 was latest; and for the forefield meadow, 2015 was earliest and 2019 was latest.
Table 5. Seasonal dates of (A) onset of snow cover in autumn/winter and spring/summer melt date of snow from soils of the slope above the ribbon forest, forefield meadow below the ribbon forest, and soil circles. Forefield meadow: west transect, 1–3; center, 4–5; east, 6–9. (B) onset and dessication of waterflow in rock wells and rocky matrices below pika haypiles
Similarly, we estimated annual dates for onset of water flow and desiccation of water in the rock well springs (, ). One well was dry throughout the warm season of 2016; water was estimated to be present in all wells and years otherwise. Dates for onset of water flow in spring varied by year, with the earliest recorded 1 May (2018) and latest 17 June (2017), a difference of six weeks. The relative order of years for onset of water flow in rock wells, from earliest to latest, was 2018, 2016, 2019, 2015, 2017. Desiccation ranged from 16 June (2016) through 14 August (2017), a range of eight weeks.
The number of F-T days per year measured at the air temperature position (2.0 m in ribbon forest) ranged from 25.0 (2015) to 87.0 (2018) and mean of 59.0 days (). We did not have the full year of data for 2019, so could not estimate F-T for that year. The numbers of annual F-T days at matrices below haypiles and rock wells were similar to the air temperature position, with means of 57.1 and 42.2 days, respectively, although the ranges were smaller across years than for the ground positions. Soil circles had the lowest mean (38.6 days) and smallest range in F-T days (34.7–52.0); haypile surfaces had the highest mean F-T (73.6 days) and largest range (46.3–98.0 days).
Table 6. Freezing days and freeze–thaw events. Number of days when temperatures were <0°C and number of freeze–thaw days at four positions of the Kavanaugh Plateau study area
The mean number of days below freezing was 235 for the air temperature position with a small annual variance (9.0 days; ). Means were similar for the rock wells and haypile surface and matrix positions, although annual variances were two to three times larger. Soil circles had the lowest mean number of days below freezing across years (178.8).
Ecological measurements
Ribbon forest
Mean tree age of whitebark pines in the ribbon forest was 90.8 years in 2020 (); trees on the upslope/windward edge of the ribbon were significantly younger (56.2 years) than those on the downslope edge (mean = 125.4 years; p = .008). Diameters of cored trees (mean = 17.0 cm) and all trees measured in plots (17.8 cm) were small but included a few large stems (maximum = 72 cm diameter). Stems in the ribbon forest were dense, with a mean of 23.0 stems/plot (69,748/ha), yielding 100 percent cover by mature trees. No seedlings were observed in ribbon forest plots.
Table 7. Ribbon forest tree ages, diameters, density, basal area, and tree cover, Kavanaugh Plateau study area. Odd numbered plots were on the west side of the ribbon (windward); even numbered plots were on the east side
Shrub and ground vegetation
The zones and features of Kavanaugh Plateau supported different vegetation types and plant species conditions. Soil circles of the PG ecosystem had four times more plant species (forty-four) than the study area forefield meadow (ten) or forefields of pika talus habitat on slopes adjacent to the study area (twelve species; ). Plant functional traits also differed among the features, with soil circles supporting a high proportion of deciduous, herbaceous-perennial herbs and greater diversity in life forms (six total) and woodiness (four total) than the other communities. The forefield meadow had 100 percent deciduous and perennial species, with more graminoids (60 percent) than the other features. Species diversity was low in the forefields of adjacent talus slopes but mixed in life form, leaf form, and woodiness. Plant cover was high in the soil circles (80.4 percent) and forefield meadow (73.9 percent) and very low in the talus slope forefield (9.1 percent), where bare earth and rocks were common in plots. Species density per plot was highest in the soil circles (mean 9.2), moderate in the forefield meadow (5.2), and lowest in the talus slope forefields (2.5).
American pika activity
We mapped fifty-three distinct American pika haypiles, including fresh and old, distributed across the PG ecosystem (; ). Haypiles were located in the rock-terrace rampart and in stone domains of the rock wells. Fresh haypiles (green vegetation) were observed in 2014, 2015, 2016, 2019, and 2020, during which we also observed and heard pikas near haypiles in the PG ecosystem throughout the warm season. We counted the largest number of fresh haypiles in 2014 (19). All other years with fresh haypiles had half this number or less; 2016 and 2020 had fewest fresh haypiles (seven and five, respectively). No fresh haypiles and no direct or indirect signs of pikas were observed in 2017 or 2018 despite multiple surveys throughout summer and autumn. Size of fresh haypiles varied by year (, ), with the largest haypiles observed in 2014 and the smallest haypiles in 2020. Based on clustering of haypiles and average territory size found in typical pika habitat (~550 m2; A. T. Smith and Weston Citation1990), fifteen to seventeen pika territories would be indicated for the PG ecosystem, which yields a density of 8.0 to 9.0 pikas per hectare. Our observations of pika behavior, however, indicate that pikas were tending haypiles only at single or directly adjacent rock wells and that clustering of territories was much greater than in classic talus. We estimated territory sizes for pikas in the PG ecosystem to be ~100 m2, which yields a count of twenty-eight to thirty-two territories and density of fourteen to sixteen pikas per hectare. In addition to haypiles, we found fecal pellets—diagnostic signs of pikas—in nearly every well of the PG zones, indicating widespread use of the landform by pikas.
Table 8. Plant cover, species list and functional traits at Kavanaugh Plateau patterned-ground ecosystem ecosystem and on adjacent pika habitat
Table 9. Pika haypiles at the Kavanaugh Plateau patterned ground ecosystem, 2014–2020. Haypile numbers correspond to locations in
Eight active haypiles were found in 2014 in the talus slopes adjacent to the PG ecosystem (north and south) and monitored to 2020. These were separated from one another by more than 200 m and had evidence of occupation (fresh vegetation) in all years except 2017.
Discussion
We present the first study of cryogenic PG in the Sierra Nevada; evaluate spatially patterned ecological features that have been little investigated in North America outside the Rocky Mountains; and propose a scenario for complex functioning of the integrated ecosystem. The Kavanaugh Plateau landscape has aspects that resemble PG landscapes elsewhere and also elements that appear novel. We evaluate individual features first, starting with the upslope zones, and then interpret the system holistically.
Ribbon forest
The ribbon forest of whitebark pine is the only significant linear patch of conifers on the plateau and the only one that lies perpendicular to the dominant wind direction and fall line. Other pines on the plateau occur as individual, windswept stunted trees or as small patches of irregular shape (SM ).
Solitary linear strips of high-elevation conifers oriented perpendicular to the wind and fall line were classified as “tree-limit ribbon forests,” one of three subtypes described in a review by Bekker and Malanson (Citation2008). Unlike “extensive ribbon forests,” which occur as closely associated, repeating strips of forests (Buckner Citation1977), tree-limit ribbon forests occur singly and near upper treeline. Bekker and Malanson (Citation2008) considered tree-limit ribbon forests the least studied and poorest understood of the ribbon forest types, documented in the Northern Hemisphere only for the Rocky Mountains of the United States and Canada (similar ribbon forests of Eucalyptus were described from Australia, where ecological conditions are much different; Green and Venn Citation2012). These rarer types of ribbon forest occur on solifluction terraces or structurally defined bedrock ridges (Holtmeier Citation1985; Butler et al. Citation2003). The Kavanaugh tree-limit ribbon forest is likely associated with a structural deformation and not a solifluction terrace. Unlike conditions described in the Rocky Mountains, however, the ribbon forest sits along the crest of a one-sided slope rather than on a ridge. Above the Kavanaugh ribbon forest, the slope inclines gradually upward, and below the ribbon forest the slope falls in a short but very steep pitch. Further unlike the contexts in the Rocky Mountains, the slope of the Kavanaugh ribbon forest is limited to a short, marked deformation in the otherwise even-surfaced plateau. In some Rocky Mountain ribbon forests, melting of snowdrifts creates moist soils that favor conifer seedling establishment and expansion of the forest strip (Billings Citation1969). By contrast, the Kavanaugh ribbon forest appears to be stable and restricted to its current location atop the steep slope, with no evidence of spread in any direction.
Snowbed slope
Commonly associated with high-elevation linear forests of all types are deep snowdrifts that accumulate in the lee of the ribbon (Billings Citation1969; Buckner Citation1977; Earle Citation1993). Although snow accumulates on the downslope/lee side of individual pines and clusters of pines on the plateau, the snowbank of the Kavanaugh snowbed slope and forefield meadow is the largest and persists far longer than any other accumulations on the plateau (). The snowdrift below the Kavanaugh ribbon forest regularly extends from the top of the snowbed slope halfway across the forefield meadow (). Timing of spring snowmelt of the snowdrift proceeds from the east (downslope/distant from the ribbon forest), where the drift is thinnest and most exposed to sun, to the west (upslope/adjacent to the ribbon forest). Snow lingers in small patches on the snowbed slope and upper forefield meadow into mid-late summer, even in years of low winter snow ().
Snowbeds are typically described as small alpine landforms unrelated to ribbon forests that accumulate deeper snow than adjacent slopes and occur in topographic depressions (Björk and Molau Citation2007). Changes in wind velocity caused by the depression lead to a deep snowdrift forming within the depression, which melts to become a snowbed feature. In the Kavanaugh situation the snowbed develops along a steeply pitched, one-sided slope rather than in a depression and benefits from the added snow-fence effect of the ribbon forest. Common to snowbeds in general is the effect on plant growth and vegetation communities of snow dominance and, in turn, on soil temperature and soil moisture (Billings and Bliss Citation1959; Shimono and Kudo Citation2003). Although we did not measure temperatures on the Kavanaugh snowbed slope, the lack of soil development and absence of vegetation point to long annual snow cover that precludes plant establishment due to short growing seasons.
Forefield meadow
Wet meadows typically develop in flat areas where moisture accumulates in upper soil horizons and persists into the growing season. This situation favors establishment of graminoid plant species and other herbs and low shrubs adapted to saturated and wet, organic soils and excludes establishment of conifers (N. B. Benedict Citation1983). Cold soil temperatures typically are found for snowbeds and meadows (Ostler et al. Citation1982; Björk and Molau Citation2007). The Kavanaugh forefield meadow parallels these situations, where snowmelt from the persistent snowdrift below the ribbon forest saturates the soils on the flat bench below it, maintaining meadow conditions. The flat aspect of the meadow allows melting snow to inundate soils rather than draining downslope quickly, further promoting development of organic soils. Relatively colder soil temperatures and long duration under snow of the Kavanaugh forefield meadow parallel expectations elsewhere for wet meadows.
Species diversity in snowbeds studied in other regions generally is lower than that on adjacent uplands, although plant cover is relatively high and dominated by herbaceous species (and bryophytes where soils are especially saturated) with fewer shrubs (Kudo and Ito Citation1992; Björk and Molau Citation2007). Of the features we measured, vegetation on the Kavanaugh forefield meadow similarly had lowest species diversity (comparable to Björk and Molau Citation2007); highest proportions of herbaceous plants, including graminoids; lacked shrubs; and had relatively high plant cover. Four of the ten species identified on the meadow did not occur on the soil circles or talus slope forefields, but these are not rare species for the area, occurring in meadow and alpine habitats elsewhere in the region.
Rock-terrace rampart
The linear rock-terrace rampart that borders the forefield meadow is sorted to the extent that fine sediments are absent, clasts are regular in size (larger than those of the rock wells below), and the rocky lineament is elevated. Though we do not know of similar features elsewhere, the Kavanaugh rampart resembles miniature and more haphazard rocky borders that we have observed at the bases of long-persisting snowbanks in the region (Millar and Westfall Citation2008). These occur below snowdrifts that recur annually in the same locations, are not associated with PG, and lie at the base of moderate-to-steep northward aspect slopes. Their development appears to result as melting snow at the base of the broad snowbank percolates through rocky soils and filters fine sediments downslope, leaving larger clasts patchily along the base of the snowpack. The Kavanaugh rampart, by contrast, lies on a flat slope at the edge of a meadow. Further, although the snowbank above is long-persisting, its base is distant from the rampart and thus unlikely to be involved with maintenance of the rampart, at least under the present climate. We speculate that the rock-terrace formed, as described in more detail below, during former cold climate intervals.
Sorted circles
In many aspects the patterned ground of the Kavanaugh Plateau resembles sorted-circle landscapes described elsewhere (Hallet Citation2013; Rowley et al. Citation2015; Wilkerson and Schmid Citation2016). At Kavanaugh, stone domains are abruptly segregated from soil domains. The latter comprise surface soils of fine- to small-grained sediments and are densely colonized by vegetation. Dimensions, shapes, and environmental context of the soil circles are similar to those measured in other temperate North American ranges, including the Rocky Mountains, Colorado (J. B. Benedict Citation1992), and White Mountains, California (Mitchell, Lamarche, and Lloyd Citation1966), as well as in Arctic regions (Washburn Citation1956; Feuillet, Certini, and Ugolini Citation2014). The stony borders at Kavanaugh compare with conditions elsewhere as well, in stone size and regularity, lack of fine sediments, sunken character of the stone domains, and rocks with flat sides (J. B. Benedict Citation1992; Hallet Citation2013; Feuillet, Certini, and Ugolini Citation2014). Lichen development on some but not all stones of the borders (SM ) and dense vegetation on the soil circles are further indicators that these sorted circles are inactive at present.
Elevations of comparable PG systems in temperate North American mountains tend to be higher than at Kavanaugh, at more northerly latitudes, and/or in colder, more continental climates (J. B. Benedict Citation1992). At the one location south of the latitude of Kavanaugh that has been studied, the White Mountains of California, PG features occur 1,000 m higher than at Kavanaugh (Mitchell, Lamarche, and Lloyd Citation1966; Wilkerson Citation1995). Of fifteen PG features mapped in Millar and Westfall’s (Citation2008) Sierra Nevada survey, the four sorted circles described in that study occurred at mean elevation 200 m higher than Kavanaugh.
The mean annual air temperature (1.5°C) at Kavanaugh is considerably warmer than the 0°C threshold commonly assumed necessary for permafrost to develop (Péwé Citation1983). In some alpine environments, however, such as caves, rock glaciers, and taluses, discontinuous permafrost can develop at MAT >0°C (Harris Citation1986; Gude et al. Citation2003). This has been shown for the Sierra Nevada, where persistent ice has been interpreted to occur within rock glaciers and talus slopes, including elevations and MAT similar to Kavanaugh (Millar, Westfall, and Delany Citation2014). Nonetheless, the environmental context of the Kavanaugh PG ecosystem, with its broad, flat slope and lack of landforms associated with cold ventilation processes (Morard, Delaloye, and Lambiel Citation2010), makes it extremely unlikely that permafrost underlies the PG ecosystem now.
Other climatic features of the Kavanaugh environment appear conducive to PG development if MATs were cold enough. Slopes of the eastern Sierra Nevada escarpment lie in the snow shadow of the broad western Sierra Nevada, which experiences Pacific-dominated winter storms from the west. The exposed position of Kavanaugh Plateau lies far east of the Sierra crest, and its wind-blasted condition results in relatively shallow snow accumulation. Our temperature loggers in the PG soil circles corroborate shallow and transient snow throughout autumn and into early winter. During those seasons, temperatures decline from summer warmth in a jigsaw fashion, resulting in multiple F-T cycles. The mean number of F-T days at Kavanaugh (42), however, was far lower than temperate alpine situations observed elsewhere for PG. For instance, in the Rocky Mountains, active and relict PG was associated with microclimates having 63 to 140 F-T days (J. B. Benedict Citation1992), and in Idaho, where only relict PG features occurred, more than 200 F-T days were estimated (Malde Citation1964).
In their model for self-organization, Kessler and Werner (Citation2003) noted that PG develops where F-T cycles drive interactions between two feedback mechanisms. First, ice lenses that develop in freezing soils displace fine-grained sediments toward soil-rich domains and displace stones toward stone-rich domains, segregating clasts from an initially heterogenous substrate. Stones are moved perpendicular to the freezing front, which is commonly horizontal, in which case stones are heaved upwards to the ground surface. The freezing front can also be tilted vertically along the border of the soil and stone domains (see in Kessler and Werner 2008). In this situation, stones are forced laterally and directly into the stone domains. Freezing contributes to expansion in the soil domain, and as the soil domain mounds upward, stones that have been heaved to the ground surface avalanche by gravity toward the stone domains. Expansion in soil domains further causes stones to be squeezed and confined in the stone domain, intensifying segregation of domains.
Aside from current MAT being warm, conditions of the Kavanaugh PG corroborate development of sorted circles following key features of Kessler and Werner’s (Citation2003) model. Initial substrates are mixed in grain size, with an abundance of clasts in the range of 3 to 30 cm. The plateau is broad and relatively flat, the ground substrate is porous and uncompacted, and, even under the current climate, freezing and F-T cycles occur more in the stone domains than in the soil domains.
Vegetation development on soil circles
Given the complex soil development that occurs during PG formation, it is not surprising that soil circles support distinct vegetation structures, as has been documented from polar to alpine regions (Billings and Mark Citation1961; Mitchell, Lamarche, and Lloyd Citation1966; Cannone and Guglielmin Citation2010; D’Amico, Gorra, and Freppaz Citation2015). At Kavanaugh, vegetation in the soil circles relative to adjacent upland soils was species rich and had high plant cover dominated by perennial herbs in the soil centers. When present, shrubs were segregated to the edges of the soil domains. Similar vegetation patterning has been described for sorted circles in temperate PG terrain, as in the Rocky Mountains, Colorado (J. B. Benedict Citation1992), and several ranges of the South Island, New Zealand (Billings and Mark Citation1961). Plant species at Kavanaugh were not unique (restricted) to the soil circles but they were distinct in their concentration and density relative to adjacent upland slopes. Comparing the species present at Kavanaugh to sorted-circle vegetation elsewhere is not especially informative given the different climatic and vegetation conditions of the studies. Though functional trait analysis could be of greater comparative value than taxonomy, studies using these are limited and have addressed vegetation on PG forms other than sorted circles (Mitchell, Lamarche, and Lloyd Citation1966; Apple, Ricketts, and Martin Citation2019). Apparent stability of the vegetation, high plant cover on the soil circles, and lack of vegetation-free zones further corroborate the relict nature of the Kavanaugh PG (Mitchell, Lamarche, and Lloyd Citation1966; J. B. Benedict Citation1992; D’Amico, Gorra, and Freppaz Citation2015).
American pika behavior
Across their western North American species’ range, pikas are highly restricted to broken-rock landforms (A. T. Smith and Weston Citation1990) and, in the Sierra Nevada, dominantly to talus and rock-glacier slopes (Millar and Westfall Citation2010; Millar et al. Citation2018). These habitats provide escape from predation and, arguably most important, provide cool thermal regimes that are necessary for pika survival. The unique ventilation processes of taluses (Morard, Delaloye, and Lambiel Citation2010; Millar, Westfall, and Delany Citation2014) maintain cool temperatures within the talus subsurfaces where pikas spend much of their time, even when outside air temperatures rise over 35°C (Millar, Westfall, and Delany Citation2016). Matrices of pika talus habitat are regularly deep, extending one to many meters below the surface, with rock clasts within a range of 0.2 to 1 m diameter (A. T. Smith and Weston Citation1990). Taluses used by pikas in Great Basin mountain regions (aside from lava fields) are almost entirely on slopes and not flat ground or open plateau/benches (Millar et al. Citation2018). Pikas are cosmopolitan herbivores who feed on leaves of diverse plant species that occur in talus-forefield meadows. They commonly establish territories around the base of taluses and rarely venture more than 5 m from the talus edge before retreating to the cool and safety of talus matrices. Solitary in nature, typical pika territories and spacing are highly consistent, averaging ~550 m2 in size, separated from neighboring territories by ~25 m (A. T. Smith and Weston Citation1990). Estimated population densities are also consistent, with mean of approximately six animals per hectare (A. T. Smith and Weston Citation1990). Typical talus habitats are often extensive; smallest talus areas used as habitat range from 7,000 to 25,000 m2 (Stewart and Wright Citation2012; Stewart, Wright, and Heckman Citation2017).
Relevant to these conditions, use of the Kavanaugh PG ecosystem is novel in our experience of pikas in the Sierra Nevada/Great Basin; we know of no other pika population inhabiting sorted circles in the bioregion. Other characteristics that make the Kavanaugh PG unusual habitat for pikas include the broad, near-flat plateau; shallow depth of rock well matrices; small area of stone domains (i.e., comparable to a talus slope habitat; individual wells ~7 m2 in area and entire PG terrain of 13,000 m2); and clast size of stone domains at the small end of the range preferred by pikas.
Perhaps most striking about pika use of the Kavanaugh PG ecosystem is the tight packing of territories, twice the numbers given for typical talus (A. T. Smith and Weston Citation1990). The behavior that we have observed on the PG ecosystem is for a single pika territory to include two rock well networks and one to two soil circles. The dense, diverse vegetation in the soil circles appears to provide abundant long-lasting forage throughout the warm season and sufficient vegetation to harvest and cache adequate winter haypiles. Another distinction is the use of the entire rocky ground (PG) landform at Kavanaugh, in contrast to the typical behavior of pikas to use only talus margins. The capacity of the PG ecosystem to provide more pikas per square meter is corroborated locally by comparison to the adjacent slope taluses, where pika territories were separated by the typical distances, densities were lower, haypiles were situated at talus edges, and talus forefields had far less vegetation than on the sorted circles.
Despite the shallow depth of the stone domains and the flat, open slope of the PG ecosystem with its expansive solar exposure, the rock well temperatures were comparable to, or cooler than, pika habitat measured elsewhere in the Sierra Nevada. Millar, Westfall, and Delany (Citation2016) measured temperatures of thirty-seven widely separated pika taluses, analyzing them by elevation zones. For the zone above 3,060 m, the talus matrix MAT at 1 m depth below haypiles was warmer than at Kavanaugh, where temperatures were measured at only 20 cm depth or less (3.0°C vs. 2.7°C at Kavanaugh). Summer talus matrix temperatures in the broad study were also warmer compared to Kavanaugh (13.0°C vs. 10.6°C at Kavanaugh). This occurred although the MAT of the high-elevation sites in the broad study was identical to the MAT at Kavanaugh (both, 1.5°C), although summer air temperatures were warmer (10.7°C vs. 8.8°C at Kavanaugh). The cool temperatures of the rock well habitat, although ventilation typical of talus slopes is unlikely to operate, probably relates to the high elevation of the plateau and possibly to summer cooling afforded by the distributed underground and spring waters.
An important finding from our seven-year survey was the disappearance of pikas from the entire PG ecosystem (all territories) for two consecutive years (2017, 2018) and their subsequent recolonization to use levels similar to before. Although this pattern is not unusual for pikas, it is often overlooked in studies that conclude site extinction based on only a single year’s observation. In his classic work of the warm-marginal pika populations of Bodie, California, A. S. Smith (summarized in A. S. Smith Citation2020) documented persistent cycles of patch extirpation and recolonization. These were unrelated to annual temperature and depended on recolonization from neighboring patches in a classic metapopulation manner. At the PG ecosystem, recolonizations are likely to be sourced from the adjacent talus slope populations, which in themselves underwent annual patch extirpations and recolonizations during the years of our surveys.
The finding of many unused territories at the PG ecosystem each year (mean 19 percent occupied/year) is also not unusual. Many studies have shown that the average stable pika population is not equal to the carrying capacity of the habitat or equal to the number of existing (old + used) haypile sites (references in A. S. Smith Citation2020). One unique aspect of the PG habitat is that extirpation of territories likely is exacerbated by water in the rock wells that would submerge pika habitat and haypiles. In support of this, the 2017 water year had more than twice the snow level of years prior (Natural Resource Conservation Services Citation2020), and summer 2017 was the first year we observed site extirpation. Conversely, pikas were thriving at the PG ecosystem during record low snow years for the Sierra Nevada, including water year 2015, where snow depth was <17 percent that of water year 2017 (Natural Resource Conservation Services Citation2020), corroborating the results of A. T. Smith and Millar (Citation2018).
Scenario for the origin and integrated functioning of the patterned-ground ecosystem
We propose a scenario to explain the origin of the Kavanaugh PG ecosystem and the combined functioning of individual features as an integrated system. From analyses above, we take as given that the PG features are inactive under the present climate and are relics from past colder climates. Lacking further study, times of origin could have been as recent as the Little Ice Age (LIA; 1400–1920 CE) or as old as the late Pleistocene (>10,000 years ago). In the Sierra Nevada, decreases in summer minimum temperature of 0.2°C to 2.0°C and increases in winter precipitation of 3 to 26 cm compared to modern times have been interpreted for the LIA (Bowerman and Clark Citation2011). Even colder temperatures characterized the Sierra Nevada during the late Pleistocene (Millar and Westfall Citation2008). Temperatures interpreted for either of these times would be cold enough to promote and sustain permafrost on the Kavanaugh Plateau. Considering the large size and well-developed nature of the PG, which we interpret would require millennia for development, we speculate that the sorted circles formed in the late Pleistocene, became inactive in warm periods of the Holocene, and reactivated during the LIA. Deeper snowpacks and cooler summers of the late Pleistocene and LIA would provide meltwater that hydrated the plateau soils in general, and the plateau’s exposure and high winds would keep it snow-free in late summer and autumn, promoting F-T cycles, all factors favoring permafrost. The flat aspect of the plateau and the coarse-grained nature of the surface substrate would have provided sufficient conditions for development of a sorted-circle PG landscape as described by the Kessler and Werner (Citation2003) model.
The steep snowbed slope of the central plateau appears to be a structural deformation that contributed to focus development of the sorted circle terrain to one specific area. During the late Pleistocene, treeline was likely depressed below its current elevation and below the elevation of the current ribbon forest (Millar, Delany, and Westfall Citation2020). With greater snow depth and cooler summer temperatures, the structural deformation would promote snowbank accumulation in its lee even without the benefit of a ribbon forest. Localization of this snowbank would focus summer snowmelt downslope along the width of the drift, contributing to high soil moisture on the slope directly below.
During colder, wetter intervals of the late Pleistocene and possibly LIA, the persistent snowbank that would build annually below the snowbed slope would have been much larger than at present. We propose that during such intervals, the base of the summer snowdrift extended to the edge of the current rock-terrace rampart. The density of a larger snowbank may have been great enough to promote, in combination with an underlying permafrost layer, slumping of the ground below the snowbank via gelifluction (Hu et al. Citation2021), causing soil and rock debris to be “plowed up” at the snowdrift front (SM Figure 15). Abundant meltwater percolating through debris from a much larger snowbank would remove fine sediments, leaving the rocky rampart that we observe at present.
Although the Kavanaugh Plateau was likely barren of pines during the late Pleistocene and colder temperatures would have restricted vegetation to extreme cold-adapted alpine plants and cryptogams, we propose that pikas could have been present at that time. Pikas are currently abundant throughout the eastern Sierra Nevada across broad elevation ranges and extend into climates far colder than at Kavanaugh (Millar and Westfall Citation2010; Millar et al. Citation2018). Lack of glacial cover and development of sorted circles during the late Pleistocene may have enabled and conditioned the Kavanaugh Plateau PG for colonization by pikas. The talus slopes adjacent to the PG connect with others at lower elevation and could have provided dispersal corridors for pikas to enter the plateau during the Pleistocene. The talus slopes adjacent to the PG ecosystem, also unglaciated during the late Pleistocene (Wahrhaftig et al. Citation2019), would have been suitable habitat, providing a classic metapopulation environment on the plateau. Once pikas discovered the PG sorted circles they may well have inhabited the ecosystem to the present, in the manner of the interannual patch extirpation/recolonization pattern we observed.
As temperatures warmed in the early Holocene, permafrost would diminish and disappear from the Kavanaugh Plateau, the cryogenic PG-forming processes would decline and cease, and the sorted circles would become inactive. Concomitantly, appropriate conditions for migration upward and establishment of diverse alpine herb and shrub communities would emerge. Whitebark pines would also advance upslope. Unlike pikas, however, whitebark pines likely have been intermittent on the plateau during the Holocene, as warm and cold intervals caused shifting of treeline above and below the current plateau elevation (Millar, Delany, and Westfall Citation2020). Though the ages of trees in the current ribbon forest do not offer insight into the original (early Holocene?) establishment of a ribbon forest atop the PG ecosystem, they are informative about the most recent colonization. The young ages of the trees relative to species longevity (>500 years; Arno and Hoff, Citation1989) suggest that the current ribbon forest established in the early twentieth century as the LIA waned; older trees near the crest of the slope, along with a few considerably older trees, point to a thinner strip of forest during the cold LIA centuries. This scenario is corroborated by fringe forests of whitebark pine that have been documented along the crests of large convex hollows in the Sierra Nevada (Millar et al. Citation2004). These mid-elevation hollows are areas where formerly late-lying snowbanks precluded conifer establishment under colder (LIA) climates, and subsequent warming of the twentieth century allowed snowbanks to melt earlier and pine seedlings to establish (Millar et al. Citation2004). This could portend the future situation at Kavanaugh with continued warming.
Despite a decline in size and summer persistence of the Kavanaugh snowdrift that would be driven by warming temperatures of the Holocene, the effect would be counteracted somewhat by the appearance of the ribbon forest and its snow fence role. In climates as warm as present, when the snowbank retreats toward the snowbed slope, meltwater contributes to development and maintenance of the diverse and productive forefield meadow and its vegetation communities. This meadow unlikely existed during the late Pleistocene, when a snowbank would have covered the entire flat area to the rampart for too long during the growing season to allow plants to establish.
The distributed springs emanating from the rock wells are one of the most interesting and unique aspects of the Kavanaugh PG ecosystem. With warming climates of the Holocene and associated cessation of cryogenic processes, the sorted circles would be expected to become inactive and erode over time (Wilkerson Citation1995). We propose a process at Kavanaugh, based on lithology and the unique aspects of the PG ecosystem, to explain how the springs originated. We further hypothesize that the springs both contributed to development of PG under Pleistocene climates and serve to maintain at least some of the integrity of the sorted circles during warm Holocene climates.
The coarse and unconsolidated surface mantle of the Kavanaugh Plateau, derived from till of the Dunderberg Peak moraines, overlies an extensive granitic formation (Huber, Bateman, and Wahrhaftig Citation1989). At the latitude of Kavanaugh Plateau, the metamorphic mantle is shallow (<30 m at the western crest, estimated using GEP’s measuring tool), and the underlying granitic block appears solid, as can be seen from the edge of the plateau (). We propose that this granitic formation serves as a barrier beneath the plateau surface and that it retards surface water from percolating deep into the ground. In particular, melting water from the large snowbank below the ribbon forest of the PG ecosystem would percolate into the soil of the forefield meadow, hit the granitic boundary, and be deflected surface-ward downslope. The path of least resistance upward would be through the voids of the stony wells of the sorted circles, thus leading to the springs distributed throughout the PG ecosystem that we observe in early–mid-summer. Desiccation of the springs follows closely upon the annual melt dates of the snowbank, corroborating that the spring waters derive from a nearby source. The existence of a granitic boundary layer seems more likely to serve these functions than a seasonal subsurface ice layer, which is unlikely to develop in the soils and current climate of Kavanaugh.
We propose that water emerging from the springs contributes to maintaining the integrity of the soil circles even during warm Holocene conditions and contributes to the vegetation conditions we observed on the soil domains. In mid-summer when the springs are active, water emerges from the springs with considerable energy/pressure. This leads to the rock wells (springheads) being cleansed of fine sediments by the flowing water and also reduces lichen development from stones in the center of the wells, which initially confused us about age of the sorted circles. Thus, even during warm intervals when the cryogenic sorting processes are not active, segregation of rock from soil domains is maintained by the action of spring water.
Water flowing from the distributed springheads rapidly loses energy once it emerges from the rock wells and encounters relatively flat terrain. Downslope from each rock well, as a result of the regular organization of the PG, lies a soil circle. Water from the rock wells flows onto the downslope soil circle, depositing sediments as it loses velocity, and dissipating on the surface (, SM ). In this way, the springs also function as a distributed “irrigation system” to water the soil circles. High soil moisture in turn leads to development of the dense vegetation we documented for the circles, as well as flattening of the surface of the soil domain (PG circles elsewhere often have mounded or convex surfaces) beyond the inclination of the plateau slope.
The functioning of the granitic subsurface boundary layer to deflect meltwater may well have been active during colder intervals of the Holocene and late Pleistocene as well as warm times such as the present. In the former case, a lithologic boundary layer function, as we are proposing for the granitic formation, would complement the action of permafrost and serve to intensify development of PG.
Conclusions
The spatially organized terrain situated on Kavanaugh Plateau exhibits known features of cryogenic PG and ecologically patterned features, as well as novel elements. In combination, these features interact as a complex and apparently integrated ecosystem. Given indications for the current inactive condition of the sorted-circle area, we propose a late Pleistocene origin for the PG landscape, during which times temperatures were cold enough to support permafrost on the plateau. Other features, including the windswept condition, unglaciated cover, and location east of the range crest, suggest that the plateau would be snow-free by late summer and early autumn even in the late Pleistocene, exposing the plateau to multiple freeze–thaw cycles. Water melting from the large snowbank that develops under the ribbon forest on the otherwise unwatered plateau would further provide late-season soil moisture downslope. These factors, with the addition of coarse, rocky, and unconsolidated surface substrates, provide the necessary combination of conditions to support development of a sorted-circle PG terrain.
With warming of the Holocene, plant species, including whitebark pine, would shift upslope. The windswept nature of the plateau makes tree establishment difficult, except at the crest and in the lee of the single deformation, a steeply pitched short slope in mid-plateau. A tree-limited ribbon forest formed, which, through snow fence action, accentuated accumulation of a large snowbank, although conditions were warmer and likely drier than the Pleistocene. The smaller Holocene snowbank provided meltwater for development of the large forefield meadow and encouraged establishment of wet meadow–adapted herbaceous vegetation. Although the sorted circles would become inactive during warm Holocene times, we propose that the large granitic formation underlying the shallow surface mantle of the plateau serves as a boundary layer to deflect meltwater from percolating deep into the ground. Instead, water is forced back to the surface, emerging in the rock wells of the PG stone domains. The water from the springheads emerges with pressure and maintains the wells free of fine sediments. The flat slope, however, causes the flowing water to quickly lose energy, and it dissipates on adjacent downslope soil domains, adding fine sediments to their surfaces, irrigating them, and maintaining a species-rich, dense, and distinct vegetation community.
American pika are well established on the PG terrain, using the stone domains as habitat and foraging on vegetation from the soil domains. We propose that pikas colonized the PG ecosystem as it developed in the late Pleistocene and that the ecosystem has supported a population ever since. Despite the apparently shallow nature of the stone border networks and lack of conditions for typical talus ventilation, the matrices of the rock wells are cool and compare to pika habitat elsewhere. The geometry of the PG terrain enables pikas to pack more territories into smaller areas than in typical habitats. Interannual patch extirpation and recolonization occur in the PG as elsewhere in pika landscapes. Extirpation coincided with heavy snow years, a situation we have observed elsewhere, and likely exacerbated at the Kavanaugh ecosystem by meltwater drowning the rock wells.
The Kavanaugh Plateau complex PG ecosystem is especially notable for its occurrence in a warm-temperate, Mediterranean-dominated mountain range. Though other landscapes such as the Kavanaugh situation may exist in the range or elsewhere in the Great Basin, they have been overlooked and understudied. This unique alpine landscape provides a window into former periglacial environments and processes and exhibits contemporary integrated functioning of physical, hydrologic, climatic, floristic, and faunal elements. The Kavanaugh PG ecosystem is worthy of protection, especially because it is in an area of heavy off-road vehicle use, for further study and interpretation.
Supplemental Material
Download Zip (3 MB)Acknowledgments
We thank Diane Delany (USDA Forest Service) for developing the graphics, Greg Stock (National Park Service) for discussions about geology, and Chrissy Howell (USDA Forest Service) for review of the draft manuscript. Operational funds from the USDA Forest Service supported this study.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental material for this article can be accessed on the publisher’s website
Additional information
Funding
References
- Apple, M. E., M. K. Ricketts, and A. C. Martin. 2019. Plant functional traits and microbes vary with position on striped periglacial patterned ground at Glacier National Park, Montana. Journal of Geographical Sciences 29 (7):1127–41. doi:https://doi.org/10.1007/s11442-019-1649-3.
- Arno, S. F., and R. J. Hoff 1989. Silvics of whitebark pine (Pinus albicaulis). United States Department of Agriculture Forest Service Intermountain Research Station General Technical Report INT-253. 11 .
- Basagic, H. J., and A. G. Fountain. 2011. Quantifying 20th century glacier change in the Sierra Nevada, California. Arctic, Antarctic, and Alpine Research 43 (3):317–30. doi:https://doi.org/10.1657/1938-4246-43.3.317.
- Bekker, M. F., and G. P. Malanson. 2008. Linear forest patterns in subalpine environments. Progress in Physical Geography: Earth and Environment 32 (6):635–53. doi:https://doi.org/10.1177/0309133308101384.
- Benedict, J. B., 1992. Field and laboratory studies of patterned ground in a Colorado alpine region. University of Colorado, Institute of Arctic and Alpine Research, Occasional Paper 49, 38.
- Benedict, N. B. 1983. Plant associations of subalpine meadows, Sequoia National Park, California. Arctic and Alpine Research 15 (3):383–96. doi:https://doi.org/10.2307/1550833.
- Billings, W. D. 1969. Vegetational pattern near alpine timberline as affected by fire-snowdrift interactions. Vegetatio Acta Geobotanica 19 (1–6):192–207. doi:https://doi.org/10.1007/BF00259010.
- Billings, W. D., and A. F. Mark. 1961. Interactions between alpine tundra vegetation and patterned ground in the mountains of southern New Zealand. Ecology 42 (1):18–31. doi:https://doi.org/10.2307/1933264.
- Billings, W. D., and H. A. Mooney. 1959. An apparent frost hummock-sorted polygon cycle in the alpine tundra of Wyoming. Ecology 40 (1):16–20. doi:https://doi.org/10.2307/1929918.
- Billings, W. D., and L. C. Bliss. 1959. An alpine snowbank environment and its effects on vegetation, plant development, and productivity. Ecology 40 (3):388–97. doi:https://doi.org/10.2307/1929755.
- Björk, R. G., and U. Molau. 2007. Ecology of alpine snowbeds and the impact of global change. Arctic, Antarctic, and Alpine Research 39 (1):34–43. doi:https://doi.org/10.1657/1523-0430(2007)39[34:EOASAT]2.0.CO;2.
- Bonachela, J. A., R. M. Pringle, E. Sheffer, T. C. Coverdale, J. A. Guyton, K. K. Caylor, S. A. Levin, and C. E. Tarnita. 2015. Termite mounds can increase the robustness of dryland ecosystems to climatic change. Science 347 (6222):651–55. doi:https://doi.org/10.1126/science.1261487.
- Bowerman, N. D., and D. H. Clark. 2011. Holocene glaciation of the central Sierra Nevada, California. Quaternary Science Reviews 30 (9–10):1067–85. doi:https://doi.org/10.1016/j.quascirev.2010.10.014.
- Buckner, D. L. 1977. Ribbon forest development and maintenance in the Central Rocky Mountains of Colorado. PhD dissertation. Boulder. University of Colorado.
- Butler, D. R., G. P. Malanson, M. F. Bekker, and L. M. Resler. 2003. Lithologic, structural, and geomorphic controls on ribbon forest patterns in a glaciated mountain environment. Geomorphology 55 (1–4):203–17. doi:https://doi.org/10.1016/S0169-555X(03)00140-5.
- Cannone, N., and M. Guglielmin. 2010. Relationships between periglacial features and vegetation development in Victoria Land, continental Antarctica. Antarctic Science 22 (6):703–13. doi:https://doi.org/10.1017/S0954102010000751.
- Cramer, M. D., and N. N. Barger. 2014. Are mima-like mounds the consequence of long-term stability of vegetation spatial patterning? Palaeogeography, Palaeoclimatology, Palaeoecology 409:72–83. doi:https://doi.org/10.1016/j.palaeo.2014.04.026.
- D’Amico, M., R. Gorra, and M. Freppaz. 2015. Small-scale variability of soil properties and soil–vegetation relationships in patterned ground on different lithologies (NW Italian Alps). Catena 135:47–58. doi:https://doi.org/10.1016/j.catena.2015.07.005.
- Earle, C. J. 1993. Forest dynamics in a forest-tundra ecotone, Medicine Bow Mountains, Wyoming. PhD dissertation. Seattle. University of Washington.
- Feuillet, T., G. Certini, and F. C. Ugolini. 2014. Sorted patterned ground. Encyclopedia of Planetary Landforms 8. doi:https://doi.org/10.1007/978-1-4614-9213-9_536-1.
- Green, K., and S. Venn. 2012. Tree-limit ribbons in the Snowy Mountains, Australia: Characterization and recent seedling establishment. Arctic, Antarctic, and Alpine Research 44 (2):180–87. doi:https://doi.org/10.1657/1938-4246-44.2.180.
- Gude, M., S. Dietrich, R. Mausbacher, C. Hauck, R. Molenda, V. Ruzicka, and M. Zacharda. 2003. Probable occurrence of sporadic permafrost in non-alpine scree slopes in central Europe. In Permafrost, ed. M. Phillips, S. M. Springman, and L. U. Arenson, 331–36. Lisse: Swets and Zeitlinger.
- Hallet, B. 2013. Stone circles: Form and soil kinematics. Philosophical Transactions of the Royal Society A 371 (20120357):1–17. doi:https://doi.org/10.1098/rsta.2012.0357.
- Harris, S. A. 1986. Permafrost distribution, zonation and stability along the eastern ranges of the Cordillera of North America. Arctic 39 (1):29–38. doi:https://doi.org/10.14430/arctic2042.
- Holtmeier, F.-K. 1985. Climatic stress influencing the physiognomy of trees at the polar and mountain timberline. In Establishment and tending of subalpine forest: Research and management, Proceedings of the 3rd IUFRO Workshop, Eidgenössische Anstalt für das forstliche Versuchswesen, Berichte 270, ed. H. Turner and W. Tranquilini, 31–40.
- Holtmeier, F.-K. 2003. Mountain timberlines ecology, patchiness, and dynamics, 369. Dordrecht: Kluwer.
- Hu, Y., L. Liu, X. Wang, L. Zhao, T. Wu, J. Cai, X. Zhu, and J. Hao. 2021. Quantification of permafrost creep provides kinematic evidence for classifying a puzzling periglacial landform. Earth Surface Processes and Landforms 46:465–77. doi:https://doi.org/10.1002/esp.5039.
- Huber, N. K., P. C. Bateman, and C. Wahrhaftig. 1989. Geologic map of Yosemite National Park and vicinity, California. Denver, Colorado: Misc. Investigations Series Map I-1874. USDI Geological Survey.
- Jepson Flora Project (eds.). 2020. Jepson eFlora. Accessed July 2020. https://ucjeps.berkeley.edu/eflora.
- Kessler, M. A., and B. T. Werner. 2003. Self-organization of sorted patterned ground. Science 299 (5605):380–83. doi:https://doi.org/10.1126/science.1077309.
- Kӧrner, C. 2003. Alpine plant life; Functional plant ecology of high mountain ecosystems. 2nd ed. Berlin: Springer.
- Körner, C. 2012. Alpine treelines. Functional ecology of the global high elevation tree limits, 220. Basel: Springer Science and Business Media.
- Krantz, W. B. 1990. Self-organization manifest as patterned ground in recurrently frozen soils. Earth-Science Reviews 29 (1–4):117–30. doi:https://doi.org/10.1016/0012-8252(90)90031-P.
- Krantz, W. B., K. J. Gleason, and N. Caine. 1988. Patterned ground. Scientific American 259 (6):68–76. doi:https://doi.org/10.1038/scientificamerican1288-68.
- Kudo, G., and K. Ito. 1992. Plant distribution in relation to length of growing season in a snow-bed in Taisetsu Mountains, northern Japan. Vegetatio 98 (2):165–74. doi:https://doi.org/10.1007/BF00045554.
- Lundquist, J. D., and F. Lott. 2008. Using inexpensive temperature sensors to monitor the duration and heterogeneity of snow covered areas. Water Resources Research 44 (4). doi: https://doi.org/10.1029/2008WR007035.
- Malde, H. E. 1964. Patterned ground in the western Snake River Plain, Idaho, and its possible cold-climate origin. Geological Society of America Bulletin 75 (3):191–208. doi:https://doi.org/10.1130/0016-7606(1964)75[191:PGITWS]2.0.CO;2.
- Millar, C. I., D. L. Delany, K. A. Hersey, M. R. Jeffress, A. T. Smith, K. J. Van Gunst, and R. D. Westfall. 2018. Distribution, climatic relationships, and status of American pikas (Ochotona princeps) in the Great Basin, USA. Arctic, Antarctic, and Alpine Research 50 (1):e1436296, 19. doi:https://doi.org/10.1080/15230430.2018.1436296.
- Millar, C. I., D. L. Delany, and R. D. Westfall. 2020. From treeline to species line: Thermal patterns and growth relationships across the krummholz zone of whitebark pine, Sierra Nevada, California, USA. Arctic, Antarctic, and Alpine Research 52 (1):390–407. doi:https://doi.org/10.1080/15230430.2020.1794098.
- Millar, C. I., and R. D. Westfall. 2008. Rock glaciers and related periglacial landforms in the Sierra Nevada, CA, USA; inventory, distribution and climatic relationships. Quaternary International 188 (1):90–104. doi:https://doi.org/10.1016/j.quaint.2007.06.004.
- Millar, C. I., and R. D. Westfall. 2010. Distribution and climatic relationships of the American pika (Ochotona princeps) in the Sierra Nevada and western Great Basin, USA; periglacial landforms as refugia in warming climates. Arctic, Antarctic, and Alpine Research 42 (1):76–88. doi:https://doi.org/10.1657/1938-4246-42.1.76.
- Millar, C. I., and R. D. Westfall. 2019. Geographic, hydrological, and climatic significance of rock glaciers in the Great Basin, USA. Arctic, Antarctic, and Alpine Research 51 (1):232–49. doi:https://doi.org/10.1080/15230430.2019.1618666.
- Millar, C. I., R. D. Westfall, A. Evenden, J. G. Holmquist, J. Schmidt-Gengenbach, R. S. Franklin, J. Nachlinger, and D. L. Delany. 2015. Potential climatic refugia in semi-arid, temperate mountains: Plant and arthropod assemblages associated with rock glaciers, talus slopes, and their forefield wetlands, Sierra Nevada, California, USA. Quaternary International 387:106–21. doi:https://doi.org/10.1016/j.quaint.2013.11.003.
- Millar, C. I., R. D. Westfall, and D. L. Delany. 2014. Thermal regimes and snowpack relations of periglacial talus slopes, Sierra Nevada, California, USA. Arctic, Antarctic, and Alpine Research 46 (2):483–504. doi:https://doi.org/10.1657/1938-4246-46.2.483.
- Millar, C. I., R. D. Westfall, and D. L. Delany. 2016. Thermal components of American pika habitat—How does a small lagomorph encounter climate? Arctic, Antarctic, and Alpine Research 48 (2):327–43. doi:https://doi.org/10.1657/AAAR0015-046.
- Millar, C. I., R. D. Westfall, D. L. Delany, J. C. King, and L. J. Graumlich. 2004. Response of subalpine conifers in the Sierra Nevada, California, USA, to 20th-century warming and decadal climate variability. Arctic, Antarctic, and Alpine Research 36 (2):181–200. doi:https://doi.org/10.1657/1523-0430(2004)036[0181:ROSCIT]2.0.CO;2.
- Mitchell, R. S., V. C. Lamarche Jr, and R. M. Lloyd. 1966. Alpine vegetation and active frost features of Pellisier Flats, White Mountains, California. American Midland Naturalist 75 (2):516–25. doi:https://doi.org/10.2307/2423408.
- Morard, S., R. Delaloye, and C. Lambiel. 2010. Pluriannual thermal behaviour of low elevation cold talus slopes in western Switzerland. Geographica Helvetica 65 (2):124–34. doi:https://doi.org/10.5194/gh-65-124-2010.
- Natural Resource Conservation Services. 2020. Virginia Lakes Ridge SNOTEL Site 846. Accessed September 5, 2020. https://wcc.sc.egov.usda.gov/nwcc/site?sitenum=846.
- Ostler, W. K., K. T. Harper, K. B. McKnight, and D. C. Anderson. 1982. The effects of increasing snowpack on a subalpine meadow in the Uinta Mountains, Utah, USA. Arctic and Alpine Research 14 (3):203–14. doi:https://doi.org/10.2307/1551153.
- Peinado, M., J. L. Aguirre, J. Delgadillo, and J. M. Martínez-Parras. 2005. A phytosociological survey of the chionophilous communities of western North America. Part I: Temperate and Mediterranean associations. Plant Ecology 180 (2):187–241. doi:https://doi.org/10.1007/s11258-005-3027-8.
- Péwé, T. L. 1983. Alpine permafrost in the contiguous United States: A review. Arctic and Alpine Research 15 (2):145–56. doi:https://doi.org/10.2307/1550917.
- Ray, R. J., W. B. Krantz, T. N. Caine, and R. D. Gunn. 1983. A model for sorted patterned-ground regularity. Journal of Glaciology 29 (102):317–37. doi:https://doi.org/10.1017/S0022143000008376.
- Rietkerk, M., and J. Van De Koppel. 2008. Regular pattern formation in real ecosystems. Trends in Ecology & Evolution 23 (3):169–75. doi:https://doi.org/10.1016/j.tree.2007.10.013.
- Rowley, T., J. R. Giardino, R. Granados-Aguilar, and J. D. Vitek. 2015. Periglacial processes and landforms in the critical zone. In Developments in earth surface processes, Vol. 19, ed. J. R. Giardino and C. Houser, 397–447. Oxford: Elsevier.
- SAS Institute. 2015. SAS online, Version 12. Cary, NC: JMP®Statistics and Graphics Guide.
- Shimono, Y., and G. Kudo. 2003. Intraspecific variations in seedling emergence and survival of Potentilla matsumurae (Rosaceae) between alpine fellfield and snowbed habitats. Annals of Botany 91 (1):21–29. doi:https://doi.org/10.1093/aob/mcg002.
- Smith, A. S. 2020. Conservation status of American pikas (Ochotona princeps). Journal of Mammalogy 1–23. doi:https://doi.org/10.1093/jmammal/gyaa110.
- Smith, A. T., and C. I. Millar. 2018. American pika (Ochotona princeps) population survival in winters with low or no snowpack. Western North American Naturalist 78 (2):126–32. doi:https://doi.org/10.3398/064.078.0203.
- Smith, A. T., and M. L. Weston. 1990. Ochotona princeps. Mammalian Species 352 (352):1–8. doi:https://doi.org/10.2307/3504319.
- Stewart, J. A., and D. H. Wright. 2012. Assessing persistence of the American pika at historic localities in California’s northern Sierra Nevada. Wildlife Society Bulletin 36 (4):759–64. doi:https://doi.org/10.1002/wsb.220.
- S*tewart, J. A., D. H. Wright, and K. A. Heckman. 2017. Apparent climate-mediated loss and fragmentation of core habitat of the American pika in the Northern Sierra Nevada, California, USA. PLoS ONE 12 (8):e0181834. doi:https://doi.org/10.1371/journal.pone.0181834.
- Tarnita, C. E., J. A. Bonachela, E. Sheffer, J. A. Guyton, T. C. Coverdale, R. A. Long, and R. M. Pringle. 2017. A theoretical foundation for multi-scale regular vegetation patterns. Nature 541 (7637):398–401. doi:https://doi.org/10.1038/nature20801.
- Tomaselli, M. 1991. The snow-bed vegetation in the Northern Apennines. Vegetatio 94 (2):177–89. doi:https://doi.org/10.1007/BF00032630.
- Van Vliet-Lanoë, B. 2014. Patterned ground and climate change. In Permafrost: Distribution, composition and impacts on infrastructure and ecosystems, Chapter 2. ed. O. Podrovsky, 67–106. Happaugue: Nova Science Publishers.
- Wahrhaftig, C., G. M. Stock, R. G. McCracken, P. Sasnett, and A. J. Cyr. 2019. Extent of the last glacial maximum (Tioga) glaciation in Yosemite National Park and Vicinity, California (No. 3414). Menlo Park: US Geological Survey.
- Washburn, A. L. 1956. Classification of patterned ground and review of suggested origins. Geological Society of America Bulletin 67 (7):823–66. doi:https://doi.org/10.1130/0016-7606(1956)67[823:COPGAR]2.0.CO;2.
- Wilkerson, F. D. 1995. Rates of heave and surface rotation of periglacial frost boils in the White Mountains, California. Physical Geography 16 (6):487–502. doi:https://doi.org/10.1080/02723646.1995.10642567.
- Wilkerson, F. D., and G. L. Schmid. 2016. Periglacial processes and landforms. In International Encyclopedia of Geography: People, the Earth, Environment and Technology: People, the Earth, Environment and Technology, ed. D. Richardson. Hoboken: Wiley.