
ABSTRACT
The morphogenesis of the mammalian secondary plate is a series of highly dynamic developmental process, including the palate shelves vertical outgrowth, elevation to the horizontal plane and complete fusion in the midline. Extracellular matrix (ECM) proteins not only form the basic infrastructure for palatal mesenchymal cells to adhere via integrins but also interact with cells to regulate their functions such as proliferation and differentiation. ECM remodeling is essential for palatal outgrowth, expansion, elevation, and fusion. Multiple signaling pathways important for palatogenesis such as FGF, TGF β, BMP, and SHH remodels ECM dynamics. Dysregulation of ECM such as HA synthesis or ECM breakdown enzymes MMPs or ADAMTS causes cleft palate in mouse models. A better understanding of ECM remodeling will contribute to revealing the pathogenesis of cleft palate.
Introduction
The morphogenesis of the mammalian secondary palate begins with the outgrow of two palatal shelves from the maxillary processes on both sides of the tongue on an embryonic day (E) 12.Citation1 The two vertically oriented palatal shelves soon elevate horizontally and opposite each other on E 14–15.Citation1 Then, the palatal shelves epithelia disintegrate in the midline and their mesenchymal compartment fuse completely to form an intact palatal roof.Citation1 Cells in the palatal shelves originate from three sources of embryonic tissue/structures: the superficial palatal epithelium is derived from the embryonic ectoderm, the underlying palatal mesenchyme mainly from the neural crest.Citation1,Citation2 Supporting these cells is the infrastructure composed by complex extracellular matrix network.
The extracellular matrix (ECM) is a three-dimensional, highly dynamic non-cellular architectural scaffold present in all tissues. In mammals, the ECM is composed of a complex protein network including collagens (Col), proteoglycans (PGs), glycoproteins, and Proteoglycans (PGs).Citation3 ECM not only support the tissue integrity and elasticity but also control tissue homeostasis.Citation4 ECM remodeling is an important process in the morphogenesis of many organs such as lungs, intestine, and mammary glands.Citation5 During development, ECM is undergoing dynamic deposition, degradation by growth factors-controlled synthesis and proteolysis by matrix-degrading enzymes.Citation4 Abnormal ECM remodeling can lead to embryonic lethality or abnormal morphogenesis or pathological conditions such as fibrosis and cancer.Citation5
In the palate, ECM not only forms the basic infrastructure where cells adhere via integrins but also play important roles in integrating and regulating growth factors network. They store and release growth factor, therefore controlling the bioavailability of active growth factors such as Tgf-βs, which in turn remodel ECM dynamics and palatal cell differentiation.Citation6-Citation8 They accumulate water, bind other ECM molecules, mediating palatal shelf growth, expansion, and elevation.Citation9-Citation14 Multiple signaling pathways important for palatogenesis such as FGF, TGF β, BMP, and SHH regulate ECM dynamics during palate development ().Citation6-Citation8,Citation14-Citation18 In this review, we will summarize the dynamic deposition and degradation of ECM during palate development.
Figure 1. ECM remodeling during palate development.
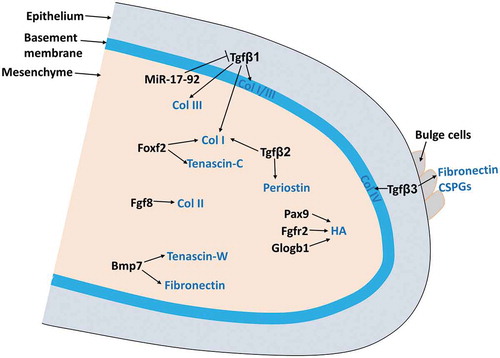
ECM molecules remodeling in the palate
Collagens (Col)
Collagens are the major components of ECM in connective tissues. There are 28 distinct collagens composed of α1, α2, α3 subunits combination and classified into fibrillar collagens (Collagen I–III, V, and XI) and non-fibrillar forms (Collagen VI, IX, IV, etc.).Citation5,Citation19 Fibrillar collagens form strong and stable fibrils and organize the fibrils into three-dimensional network, for example, Collagen I fibrils for bones and Collagen II fibrils for cartilages.Citation5 Non-fibrillar forms of collagens include Fibril-Associated Collagens and basement collagens. Fibril-Associated Collagens, such as Collagen IX, associate with collagen fibrils and bind them together to form thicker collagen fibers. Basement collagens are sheet-forming collagens such as Collagen IV, which form the two-dimensional network for all basal laminae.Citation5,Citation19 A variety of collagens are highly expressed in the palate and dynamically remodeled during palatogenesis ( and ).
Table 1. Cleft palate-related ECM gene mutation in human and mice.
Table 2. ECM molecules expression, function, and remodeling pattern during palatogenesis.
Col I and III are widely expressed in the palatal mesenchyme before and after palate shelf elevation.Citation7,Citation20-Citation25 During palatogenesis, their degradation is highly regulated. For example, Col I is downregulated in the palatal shelves of Foxf2−/- embryos which failed to elevate palate shelves.Citation15 Interestingly, in human palatal fibroblasts derived from orofacial cleft patients, COL I and III mRNA levels are strongly decreased, in contrast, the protein levels are increased compared to the control sample possibly contributing to decreased collagen degradation by MMPs and increased collagen cross-links.Citation24 COL I mRNA and protein levels in the palate are also downregulated in chondroitin sulfate proteoglycan defective mutants which have thinner palate due to abnormal bone and cartilage development.Citation14 TGF-β1, one of the most important growth factors during palate development, can induce palatal mesenchymal cells proliferation and Col I and III synthesis, which can be inhibited by MiR-17-92 clusters.Citation16
Col IV is expressed in the epithelial cell basement membrane.Citation10,Citation17,Citation20,Citation25 Col IV expression is reduced in the Tgf-β3 mutant in which palatal shelves failed to fuse.Citation17 Addition of Tgf-β3 in the palate culture increases Col IV deposition in the basement membrane of MEE cells.Citation17
In human, mutation of COL2A1, COL11A1, COL11A2, COL9A1, and COL9A2 cause a group of hereditary conditions known as Stickler syndrome I–V, respectively, characterized by high myopia, retinal detachment, hearing loss, midfacial underdevelopment, and cleft palate is only described in Stickler syndrome I–III ().Citation26-Citation32
Type II collagen is the major extracellular matrix component of cartilage and essential for endochondral bone formation.Citation5 In the palate mesenchyme, only a few osteoblast precursors express Col II in the palatal mesenchyme.Citation8 Inactivate Type II collagen in mouse develops short bones and cleft palate.Citation33-Citation38 In these mutants, both chondrocyte differentiation and intramembranous ossification are disrupted. Augmented fibroblast growth factor 8 (FGF8) signaling in the anterior hard palate by using Shox2Cre causes a subset of palatal mesenchymal cells differentiating into Col II+ chondrogenic cells at the expense of osteogenic cell fate.Citation8 These results indicate that appropriate level of Type II collagen is necessary for palate mesenchymal cell fate determination.
Mutation of COL11A1 in human causes Marshall syndrome and Stickler syndrome type. 2Citation26 Col11a1 homozygote mice have craniofacial abnormalities including cleft palate, shortened head, and mandible, short limbs, protruding tongue et.al.Citation39,Citation40 The tongue protrusion is possible to obstruct palatal shelf elevation, contact, and fusion. However, the Col11a1 mutant palatal shelves can make contact and fuse when placed close to each other in organ culture.Citation39 It indicates that Col11a1 may play a role in palate growth. Mutation of COL11A2 in human cause Stickler syndrome type 3 and Nance-Insley syndrome.Citation26,Citation41 However, inactivation of Col11a2 in mice does not lead to cleft palate, although other clinical features such as hearing loss and abnormal skeleton development replicate the human phenotypes.Citation41,Citation42
Glycoproteins
Fibronectins (FN)
Fibronectin (FN) is a glycoprotein with a high molecular weight of 230–270 KD. As one of the most widely expressed ECM in the vertebrate, FN is composed of types I, II, and III repeating units.Citation4,Citation43 FN existing in multiple isoforms is encoded by a single FN gene, located in the human chromosome 2 and rodent chromosome 1.Citation44 Alternative splicing occurs at three regions, EIIIA/EDA and EIIIB/EDB and V region. FN has multiple sites for self-assembly and ligand binding for integrins, heparin, fibrin, collagen/gelatin, and growth factors, mediating biological processes such as cell adhesion, migration, differentiation.Citation4
FN is one of the most abundant ECM components in the palate (). Although FN null embryos are embryonic lethalCitation45,Citation46, conditional knockout FN in cranial neural crest cells and mesodermal cells leads to cleft palate.Citation47,Citation48 Before palate shelve elevation, FN locates in the palatal mesenchymeCitation20-Citation23 and around MEE, with strong expression around the bulging MEE cells.Citation17,Citation20 FN expression is totally absent in MEE cells and apical surface in Tgf-β3 mutant, and the addition of Tgf-β3 in the palate culture increase FN deposition on the MEE apical surface (Martinez-Sanz et al., 2008). FN production by the human fetal palatal mesenchymal cells also can be inhibited by retinoid acid, a known cleft palate inducer, in a dose-dependent manner.Citation49 Strong Fibronectin mRNA expression is also observed at the midline epithelial seam (MES) in E14.5 wildtype or BMP heterozygous embryos.Citation6 While the same stage of BMP homozygous embryos has delayed palatal shelf elevation, only little Fibronectin mRNA expression was found in their still vertical palatal shelves. Besides, anti-fibronectin antibody can block palate shelve adhesion.Citation17 Recently, fibronectin splice-isoform ED-A domain, essential for Tgfβ latency complex formation, is shown reduced in the palatal shelves of Foxf2−/- embryos which failed to elevate.Citation15 These results indicate fibronectin is important during palate shelve elevation and fusion, downstream of Foxf2 and TGF/BMP signaling.
Tenascins
Tenascins are a family of polymorphic glycoproteins with a molecular weight of 150–380 KD, including tenascin-C, -R, -W, -X and -Y.Citation43 Tenascins are composed of repeated domains including type III domains, EGF-like repeats, and a C-terminal globular domain.Citation43 Tenascin-R is mainly found in the central nervous system and Tenascin-X and -Y in skeletal muscles.Citation43 Tenascin-C and -W are widely expressed in developing tissues and play important roles in tissue morphogenesis and tumor growth.Citation43 The expression of tenascins is regulated by mechanical loading both during development and in adulthood.Citation50 Unlike most ECM proteins mediating cell adhesion and cytoskeletal organization, Tenascins modulate cell-matrix interactions and function as adaptors.Citation51,Citation52
Tenascin-C and Tenascin-W showed distinct spatial and temporal expression patterns during palatogenesis ().Citation1,Citation6,Citation53 At E13.5, before palatal shelves elevated, Tenascin-C expression was found in the mesenchyme close to the nasal and distal surface of the shelf; After elevation, Tenascin-C expression accumulated in the mesenchyme close to the MES. In contrast, before elevation, Tenascin-W weekly expressed in the proximal-nasal quadrant of the vertical shelves; After elevation, Tenascin-W was restricted to the dorsal mesenchyme around the MES, corresponding to the future osteogenic domains of hard palate. Tenascin-W, not Tenascin-C diminished in the palatal shelves of BMP7−/- embryos can be induced by Bmp7 in embryonic cranial fibroblasts in vitro.Citation6 While Tenascin-C expression is reduced in Foxf2−/- mutant palatal shelvesCitation15 and retinoid acid overexpressed human palatal mesenchymal cellsCitation49, it is possible that they are involved in different pathways regulating palate development.
Although Tenascin-X expression has not reported in the palate, Tenascin-X deficiency (CAH-X syndrome) exhibits a bifid uvula, a mildest form of cleft palate.Citation54,Citation55 Interestingly, three proteins important in palate development, TGF-β2, TGF-β3, MMP13, are all increased in Tenascin-X deficiency patient fibroblast and tissues.Citation56
Periostin
Periostin is a secreted 90KD glycoprotein identified from a mouse MC3T3-E1 osteoblastic cell line and originally named as osteoblast-specific factor 2.Citation57 Periostin promote cell motility via integrin-dependent cell adhesion.Citation58 Periostin also plays an essential role in bone and tooth development.Citation59 Periostin null mice are growth retarded, showing incisor enamel defects indicating important roles for tooth and bone development.Citation59-Citation61
Periostin protein and mRNA are spatiotemporally expressed in the palate ().Citation7,Citation53,Citation62,Citation63 In the anterior palate (hard palate), periostin is expressed by in the mesenchyme on the oral side and part of basal membrane.Citation7,Citation62 In contrast, periostin is highly expressed in the entire posterior palate (soft palate).Citation7,Citation62
Periostin intensified around MEE when they are undergoing EMT and transdifferentiating into MES.Citation62 In contrast, laminin and Type IVcollagen, two major ECM in MEE basement membrane, are degraded earlier than periostin.Citation62 This indicates that periostin is involved regulating MEE fate during the palate fusion process.Citation62
Both TGF-β2, Col I and periostin expression are detected in the palatine aponeurosis region of the soft palate.Citation7 Exogenous TGF-β2 can induce periostin and Col I expression in the palate tissue in organ culture which indicates that TGF signaling might regulate soft palate development by mediating periostin expression.Citation7
Laminins
Laminins are a group of heterotrimeric glycoproteins composed of α, β, γ polypeptide chains and contribute to the assembly of the basement membrane.Citation64 Laminins can bind to and interact with other ECMs such as Col IV and nidogen and activate cell receptors such as integrins, glycolipids, proteoglycans, and glycoproteinsCitation65, therefore mediate cell adhesion, migration, and differentiation.Citation64 Before palatal shelve elevation, laminin is present discontinuously in the basement membrane and intercellularly in MEE cells ().Citation17,Citation66 However, in the Tgf-β3 mutant which failed to fuse, laminins are upregulated and become continuous in the basement membrane under MEE cells.Citation17 It indicates that dynamic assembly of laminins in the basement membrane is regulated by the growth factors.
Fibrillins
Fibrillins are a group of large extracellular glycoproteins including three isoforms, Fibrillin-1, −2, −3.Citation43 They compose the core microfibrils in the ECM of elastic and non-elastic tissues, and interact with integrins directlyCitation43 or bind and activate Tgf-β.Citation67 Fibrillin-1 mRNA is weakly expressed in the palatal mesenchyme only before palatal shelf elevation ().Citation6 In contrast, Fibrillin-2 mRNA is enriched in the nasal side of palatal mesenchyme before palatal shelf elevation, then increased and expanded around Tgf-β3+MEE cells and in the oral side of palatal mesenchyme.Citation6 The close relationship of strong Fibrillin-2+mesenchyme cells and Tgf-β3+MEE cells indicates that Fibrillin-2 may be important for Tgf-β mediated palatal fusion. Retinoid acid, an important regulator during embryogenesis, dose-dependently inhibit fibrillin-2 production in human fetal palatal mesenchymal cells in vitro.Citation49
Proteoglycans (PGs)
Proteoglycans are a group of complex protein families characterized by anionic glycosaminoglycan (GAG) chains covalently bounding to core proteins.Citation68 GAG chains can be classified into the below 5 classes: chondroitin sulfate (CS), heparan sulfate (HS), keratan sulfate (KS), dermatan sulfate (DS), and hyaluronan (also called hyaluronic acid, HA). Hyaluronan is a non-sulfated glycosaminoglycan and not attached to a protein core. CS, HS, KS, and DS attaching to core proteins form CSPGs, HSPGs, KSPGs, and DSPGs, respectively. PGs are classified based on their cellular and subcellular location, overall gene/protein homology and protein modules within the respective protein cores.Citation68 Only pericellular and extracellular proteoglycans regarded as ECM proteins will be discussed in this review. Pericellular proteoglycans such as perlecan (HSPGs) located in the basement membrane interact with each other and participate in modulating growth factors.Citation68 Extracellular proteoglycans constitute the major structural complex, provide viscoelastic properties, retain water, and keep osmotic pressure and regulate cell migration, proliferation, apoptosis, and angiogenesis by interacting with several receptor tyrosine kinases.Citation68 During palate development, CSPGs, HSPGs, KSPGs, and DSPGs are all enriched in palatal shelves ().Citation11,Citation25,Citation69,Citation70 For a long time, GAGs accumulation and hydration were regarded as the main source of the intrinsic force for palatal shelf elevation, as cleft palate is induced after GAG biosynthesis suppression.Citation9,Citation71,Citation72 But evidence has emerged that proteoglycans are also essential for palatal adhesion and osteogenesis.
CSPGs
CSPGs are highly expressed in the palatal mesenchyme during the palate development ().Citation11,Citation14,Citation18 Interestingly, CS chains are transiently upregulated on the apical surface of palatal medial edge epithelial (MEE) cells when they become closer and make contact.Citation11,Citation18 Alteration of CS chain synthesis or its specific digestion disrupts palatal shelves adhesion in vitro, indicating CSPGs play a functional role at palatal adhesion.Citation18 The expression of CS chains is shown absent in the MEE cells of TGF-β3 null mutant mice, whose palatal shelves are unable to fuse in the midline.Citation18 However, the expression of CS chains, together with palatal shelf adhesion, can be re-induced by the addition of TGF-β3 in palate shelf organ culture.Citation18,Citation73 Besides CS chains, core proteins such as biglycan, decorin, versican are also significantly increased in MEE cells as the palatal shelf adhering.Citation12,Citation13 Inhibitor of TGFβ signaling with SB431542, a TGFβrI kinase inhibitorCitation74, caused the failure of palate shelf fusion together with the downregulation of biglycan and decorin protein from the MES. These studies indicate that biglycan and decorin are involved in palatal shelve adhesion downstream of TGFβ signaling. Although biglycan and decorin single or double knockout transgenic mice have no cleft palateCitation75, other factors might compensate their roles during palatal shelve adhesion.Citation13 All these results indicate that remodeling of proteoglycans by TGFβ signaling are important for the palatal adhesion process.
The protein and mRNA levels of biglycan and decorin are also found in the palatal mesenchymal cells at different palate development stages, although their expression is transiently downregulated when palatal shelves are elevating and closing.Citation13,Citation76 Interestingly, in retinoic acid included mice cleft palate, decorin positive area, not biglycan, is unable to downregulate in the mesenchyme when palatal shelves are elevating, indicating that decorin is more important in palatal shelf elevation than biglycan during palatogenesis.Citation76
Ectopic Hh signaling in the palatal mesenchyme leads to the defective palatine formation and fully penetrant cleft palate and defective osteogenesis.Citation77 In these mutants, significantly downregulation of the mRNA of decorin (Dcn) and lumican (Lum, a major KSPGs) in the palatal mesenchyme indicate that decorin and lumican also play roles in the palatal cell fate determination. Recently, another study showed that reducing half the abundance of CSPGs by knocking out a key CS biosynthesis glycosyltransferase caused malocclusion, skin hyperextension, severe intramembranous ossification, and cartilage formation defects in the craniofacial development.Citation14 These mutants exhibited significant thinner palate (5% mutant has a cleft palate), where Col I, Wnt3a, β-catenin are all downregulated.Citation14 Therefore, CSPGs in palatal mesenchyme probably mediate palatal mesenchyme osteogenesis by regulating the biosynthesis of collagen type 1 and deposition of CS-binding molecules Wnt3a during palate development.
HSPGs
Heparan sulfate (HS), a sulfated GAG, is dynamically expressed in the developing palate ().Citation11,Citation70,Citation78,Citation79 Before palate shelf elevation, HS expressed in the basement membrane of the whole palate and in the mesenchyme of the anterior palate.Citation11 The expression of HS in the mesenchyme become evident in the mesenchyme at the tip of whole palatal shelf when palatal shelf is elevating and gradually disappears when palatal shelf fused together.Citation11 The expression of HS in the basement membrane is stronger in the oral/lateral side than the nasal/medial side after palatal shelf elevation.Citation11 HS directly or indirectly regulates SHH and FGF signalingCitation80-Citation83, two key signaling pathways during palate development.Citation1,Citation84 HSPGs bind both FGFs and FGFRs directly, stable their ternary complex, remain FGFs concentration in the local area, and regulate signaling activation.Citation81,Citation82,Citation85 HSPGs can act as Shh co-receptors activating Shh signaling and promote cell proliferation.Citation80 Genetically abolish heparan sulfate in the lung epithelial cells leads to reduced SHH production in the epithelial and expanded Fgf10 expression in the underlying mesenchyme in the lung development.Citation83 Shh secreted by the palatal epithelium signals to the underlying palatal mesenchyme and regulate Fgfs expression.Citation1 Fgfs, in return, can either positively or negatively regulate SHH expression in the epithelium.Citation1,Citation84,Citation86,Citation87 But additional studies need to clarify if HSPGs coordinate SHH and FGF signaling transduction in the epithelial–mesenchymal interactions during palate development.
HA
Hyaluronic acid (HA) is a high molecular mass GAG, which helps to retain a large amount of water in the mesenchyme. As a major component of palatal mesenchyme, HA is shown accumulating in the nasal side and in the hinge region of the palatal mesenchyme with higher levers in the anterior/mid-part than anterior-most and posterior palate ().Citation9,Citation11,Citation78 Regionally specific accumulation of extracellular GAGs, predominantly HA, is proposed to be the intrinsic force to drive palatal shelf elevation.Citation10,Citation49 Mice homozygous for Fgfr2C342, Pax9 and Golgb1 mutation, which have a palatal shelf elevation defect, exhibit reduced HA accumulation in the palatal shelves.Citation9,Citation88,Citation89 HA synthase Has 1, 2, and 3, which synthesize HA at the plasma membrane, are disrupted in TGF-β3 mutant palatal shelves which failed to fuse in the midline, indicating that HA remodeling in palate is highly regulated by Tgf-β signaling pathways.Citation49
Integrin signaling
Integrins are a family of heterodimeric transmembrane receptors facilitating cell-ECM adhesion and signal transduction. By the combination of 18 α-subunits and 8 β-subunits, 24 distinct integrin heterodimers form and bind to different ECM proteins, such as α1β1, α2β1, α10β1, α11β1 for collagen; α3β4, α6β4, α7β4, α9β4 for laminin; α5β1, α8β1, αvβ1, αvβ6, etc., for RGD (a tripeptide sequence, present in ECM such as fibronectin and vitronectin).Citation90 Several integrin subunits are present in the palate. Integrins α5 is expressed by the palatal mesenchyme and apical side of MEE cells at E13.5, and its expression around MEE is absent in the Tgf-β3 mutant.Citation17 In contrast, Integrins β1 is absent from palatal mesenchymal cells but highly expressed by MEE cells at E13.5.Citation17,Citation62 Although the expression of Integrins β1 is no change in the Tgf-β3 mutant.Citation17 The addition of Tgf-β3 in the palate culture induces both Integrins α5 and Integrins β1 expression on the MEE apical surface.Citation17 Anti-Integrins α5 antibody blocks palate shelve adhesion in organ culture.Citation17 Inactivation of Integrins α5 from either palatal neural crest cells with TFAP2αIRESCre or from mesodermal cells with Mesp1Cre can cause cleft palate.Citation47,Citation48 But further studies are needed to clarify how Integrins α5 and Integrins β1 are involved in ECM remodeling and signaling transduction during palatal shelf elevation and adhesion. Integrin αV, β3, β5 are also highly expressed by MEE cells.Citation62 But their functions during palatal adhesion need to be further investigated. Loss of both Integrins α5 and αV from palatal neural crest cells with Wnt1Cre leads to cleft palate, where palatal shelves still remain small at E17.5Citation91, indicating that Integrins α5 and αV are essential for palatal shelve expansion. Only a small population of Integrin β8 heterozygous and homozygous embryos developed cleft palateCitation92, indicating other α subunits are required for cleft palate phenotype.
Talin (Tln) is one of the important intracellular proteins which activates integrins by binding to its β subunit.Citation93,Citation94 Two Tln isoforms are present in most vertebratesCitation95 and three in zebrafish.Citation96 In zebrafish, tln1 is required for the cranial neural crest cell proliferation during palate morphogenesis.Citation97 In mice, global loss of Tln1 leads to embryonic lethality during gastrulationCitation98, while Tln2 null mice are viable and fertile.Citation99 Conditional mouse models would provide more evidence on how Talin engages in Integrin signal transduction during palate development.
ECM remodeling by Extracellular metalloproteinases
ECM is dynamically remodeled by extracellular metalloproteinases, including Matrix metalloproteinases (MMPs) and their endogenous tissue inhibitors (TIMPs), a disintegrin and metalloproteinases (ADAMs), and ADAMs with thrombospondin motifs (ADAMTS). During palate development, MMPs and TIMPs are spatiotemporally expressed in the mouse embryos, correlating to their ECM substrates ().Citation100
Table 3. MMP associated with palate development.
MMPs 2, 3, 9, 13, 14, and 25 and TIMPs 1, 2, and 3 are spatiotemporally expressed in the mouse embryonic palate and MMP-9 and TIMP 4 are detected in the newborn human palate tissue suggesting ECM remodeling by MMPs and TIMPs are essential for palate development ().Citation100-Citation104 More importantly, MMP-13 and TIMP-2 are transiently highly upregulated in the MEE while palatal shelves are elevating and fusing.Citation100-Citation103 Their expression patterns preceded the decreases of their substrates, fibronectin, collagen I, and III.Citation100,Citation101,Citation103 In Tgf-β mutant mouse, which failed to fuse palate shelves in the midline, MMP-13 and TIMP-2 are significantly reduced or totally absent in the MEE.Citation105 And inhibition of MMP-13 synthesis and excessive TIMP-2 in palatal organ culture phenocopies Tgf-β mutant phenotypeCitation105,Citation106, indicating that ECM remodeling in the midline is essential for palatal fusion.
Almost 80% of double null of MMP-14 and −16 have a cleft palate.Citation107 Neither MMP-14 nor MMP-16 single mutant exhibits a cleft palate, indicating that MMP-14 and MMP-16 have overlapping roles on ECM remodeling during palate development.Citation107
MMP-25 protein and mRNA are increased in the tips of palatal shelve while palatal growing, where it significantly decreased when neutralize TGF-β3Citation108 Knockdown of MMP-25 in palatal organ culture impairs palate shelf fusion and persistent MES, indicating MMP-25 is a direct transcriptional target for Tgf-β3 in the palate development.Citation108
Adamts 20 was found universally in the palatal shelf mesenchyme, while Adamts 9 mainly in the palate microvascular endothelium.Citation12 Although Adamts 9 and 20 single mutant did not show cleft palate, haploinsufficiency of Adamts9 in Adamts 20 mutant mice (Adamts 9±;Adamts20bt/bt) showed a secondary cleft palate with 100% penetrance.Citation12 These mice had defects in palatal mesenchymal cell proliferation and versican cleaving.Citation12 Haploinsufficiency of versican (VCAN) in Adamts 20 mutant mice (Vcanhdf/+;Adamts20bt/bt) phenocopied the phenotype of Adamts 9±;Adamts20bt/bt mice.Citation12 The collectively versican proteolysis by ADAMTS 9 and 20 in the palate are important for regulating palatal mesenchyme cell proliferation.
Taken together, these studies indicate an important role of ECM remodeling by extracellular metalloproteinases for palatal shelve expansion and fusion. But the corresponding ECM substrates of most extracellular metalloproteinases during palatogenesis are still unknown. It will be interesting to further explore how breakdown of other ECM such as collagen, proteoglycans, fibronectin, etc., by extracellular metalloproteinases facilitate palatogenesis.
Conclusion and future direction
In summary, many ECM and related genes are found to be involved in the palate development. ECM not only form the basic infrastructure of palatal shelves, but also play pivotal roles regulating cell proliferation, adhesion, cell fate determination in the morphogenesis of the secondary palate. However, the complex ECM functions and remodeling for palatal shelf expansion, elevation, and fusion has not yet been identified. We still know only little about the interaction of ECM themselves, and ECM and growth factors at different stages of palate development. Although a variety of ECM proteins expressed temporospatial during palatogenesis, it is still unknown how their dynamical expression patter might contribute to the distinct anterior-posterior palatal shelf elevation behavior. Besides, multiple signaling pathway regulates palate ECM elasticity and stiffness. But the contribution of mechanical transduction of ECM stiffness to palate elevation remains largely unknown. Future genetic studies will help us further understand the function of ECM remodeling during palatogenesis.
Disclosure of potential conflicts of interest
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article. The authors report no conflict interest.
Additional information
Funding
References
- Li C, Lan Y, Jiang R. Molecular and cellular mechanisms of palate development. J Dent Res. 2017;96:1184–91. doi:10.1177/0022034517703580.
- Lan Y, Xu J, Jiang R. Cellular and molecular mechanisms of palatogenesis. Curr Top Dev Biol. 2015;115:59–84.
- Hynes RO, Naba A. Overview of the matrisome–an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol. 2012;4:a004903. doi:10.1101/cshperspect.a004903.
- Hynes RO. The extracellular matrix: not just pretty fibrils. Science (New York, NY). 2009;326:1216–19. doi:10.1126/science.1176009.
- Bonnans C, Chou J, Werb Z. Remodelling the extracellular matrix in development and disease. Nat Rev Mol Cell Biol. 2014;15:786–801. doi:10.1038/nrm3904.
- d’Amaro R, Scheidegger R, Blumer S, Pazera P, Katsaros C, Graf D, Chiquet M. Putative functions of extracellular matrix glycoproteins in secondary palate morphogenesis. Front Physiol. 2012;3:377.
- Oka K, Honda MJ, Tsuruga E, Hatakeyama Y, Isokawa K, Sawa Y. Roles of collagen and periostin expression by cranial neural crest cells during soft palate development. J Histochem Cytochem. 2012;60:57–68. doi:10.1369/0022155411427059.
- Xu J, Huang Z, Wang W, Tan X, Li H, Zhang Y, Tian W, Hu T, Chen YP. FGF8 signaling alters the osteogenic cell fate in the hard palate. J Dent Res. 2018;97:589–96. doi:10.1177/0022034517750141.
- Lan Y, Zhang N, Liu H, Xu J, Jiang R. Golgb1 regulates protein glycosylation and is crucial for mammalian palate development. Development. 2016;143:2344–55. doi:10.1242/dev.134577.
- Ferguson MW. Palate development. Development. 1988;103:Suppl:41–60.
- Vaziri Sani F, Kaartinen V, El Shahawy M, Linde A, Gritli-Linde A. Developmental changes in cellular and extracellular structural macromolecules in the secondary palate and in the nasal cavity of the mouse. Eur J Oral Sci. 2010;118:221–36. doi:10.1111/j.1600-0722.2010.00732.x.
- Enomoto H, Nelson CM, Somerville RP, Mielke K, Dixon LJ, Powell K, Apte SS. Cooperation of two ADAMTS metalloproteases in closure of the mouse palate identifies a requirement for versican proteolysis in regulating palatal mesenchyme proliferation. Development. 2010;137:4029–38. doi:10.1242/dev.050591.
- Ibrahim I, Serrano MJ, Ruest LB, Svoboda KKH. Biglycan and decorin expression and distribution in palatal adhesion. J Dent Res. 2017;96:1445–50. doi:10.1177/0022034517722783.
- Ida-Yonemochi H, Morita W, Sugiura N, Kawakami R, Morioka Y, Takeuchi Y, Sato T, Shibata S, Watanabe H, Imamura T, et al. Craniofacial abnormality with skeletal dysplasia in mice lacking chondroitin sulfate N-acetylgalactosaminyltransferase-1. Sci Rep. 2018;8:17134. doi:10.1038/s41598-018-35412-5.
- Nik AM, Johansson JA, Ghiami M, Reyahi A, Carlsson P. Foxf2 is required for secondary palate development and Tgfbeta signaling in palatal shelf mesenchyme. Dev Biol. 2016;415:14–23. doi:10.1016/j.ydbio.2016.05.013.
- Li L, Shi JY, Zhu GQ, Shi B. MiR-17-92 cluster regulates cell proliferation and collagen synthesis by targeting TGFB pathway in mouse palatal mesenchymal cells. J Cell Biochem. 2012;113:1235–44. doi:10.1002/jcb.23457.
- Martinez-Sanz E, Del Rio A, Barrio C, Murillo J, Maldonado E, Garcillan B, Amorós M, Fuerte T, Fernández Á, Trinidad E, et al. Alteration of medial-edge epithelium cell adhesion in two Tgf-beta3 null mouse strains. Differentiation. 2008;76:417–30. doi:10.1111/j.1432-0436.2007.00226.x.
- Gato A, Martinez ML, Tudela C, Alonso I, Moro JA, Formoso MA, Ferguson MWJ, Martı́nez-Álvarez C. TGF-beta(3)-induced chondroitin sulphate proteoglycan mediates palatal shelf adhesion. Dev Biol. 2002;250:393–405. doi:10.1006/dbio.2002.0792.
- Mienaltowski MJ, Birk DE. Structure, physiology, and biochemistry of collagens. Adv Exp Med Biol. 2014;802:5–29.
- Silver MH, Foidart JM, Pratt RM. Distribution of fibronectin and collagen during mouse limb and palate development. Differentiation. 1981;18:141–49. doi:10.1111/j.1432-0436.1981.tb01115.x.
- Kurisu K, Ohsaki Y, Nagata K, Kukita T, Yoshikawa H, Inai T. Immunocytochemical demonstration of simultaneous synthesis of types I, III and V collagen and fibronectin in mouse embryonic palatal mesenchymal cells in vitro. Coll Relat Res. 1987;7:333–40. doi:10.1016/S0174-173X(87)80026-2.
- Kurisu K, Ohsaki Y, Nagata K, Inai T, Kukita T. Heterogeneous distribution of the precursor of type I and type III collagen and fibronectin in the rough endoplasmic reticulum of palatal mesenchymal cells of the mouse embryo cultured in ascorbate-depleted medium. Cell Tissue Res. 1992;267:429–35. doi:10.1007/BF00319365.
- Ohsaki Y, Nagata K, Kurisu K. Localization of types I and III collagen and fibronectin in the developing mouse palatal shelves. Acta Anat. 1995;153:161–67. doi:10.1159/000147696.
- Gagliano N, Carinci F, Moscheni C, Torri C, Pezzetti F, Scapoli L, Martinelli M, Gioia M, Stabellini G. New insights in collagen turnover in orofacial cleft patients. Cleft Palate-Craniofacial J. 2010;47:393–99. doi:10.1597/07-196.1.
- Fantauzzo KA, Soriano P. PDGFRbeta regulates craniofacial development through homodimers and functional heterodimers with PDGFRalpha. Genes Dev. 2016;30:2443–58. doi:10.1101/gad.288746.116.
- Higuchi Y, Hasegawa K, Yamashita M, Tanaka H, Tsukahara H. A novel mutation in the COL2A1 gene in a patient with Stickler syndrome type 1: a case report and review of the literature. J Med Case Rep. 2017;11:237. doi:10.1186/s13256-017-1396-y.
- Robin NH, Moran RT, Ala-Kokko L. Stickler syndrome. In: Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJH, Stephens K, editors. GeneReviews((R)). Seattle: University of Washington, Seattle University of Washington, Seattle. GeneReviews is a registered trademark of the University of Washington, Seattle; 1993. (All rights reserved).
- Melkoniemi M, Koillinen H, Mannikko M, Warman ML, Pihlajamaa T, Kaariainen H, Rautio J, Hukki J, Stofko JA, Cisneros GJ, et al. Collagen XI sequence variations in nonsyndromic cleft palate, Robin sequence and micrognathia. Eur J Hum Genet. 2003;11:265–70. doi:10.1038/sj.ejhg.5200950.
- Nikopensius T, Jagomagi T, Krjutskov K, Tammekivi V, Saag M, Prane I, Piekuse L, Akota I, Barkane B, Krumina A, et al. Genetic variants in COL2A1, COL11A2, and IRF6 contribute risk to nonsyndromic cleft palate. Birth Defects Res Part A Clin Mol Teratol. 2010;88:748–56. doi:10.1002/bdra.v88:9.
- Hoornaert KP, Vereecke I, Dewinter C, Rosenberg T, Beemer FA, Leroy JG, Bendix L, Björck E, Bonduelle M, Boute O, et al. Stickler syndrome caused by COL2A1 mutations: genotype-phenotype correlation in a series of 100 patients. Eur J Hum Genet. 2010;18:872–80. doi:10.1038/ejhg.2010.23.
- Richards AJ, Laidlaw M, Meredith SP, Shankar P, Poulson AV, Scott JD, Snead MP. Missense and silent mutations in COL2A1 result in Stickler syndrome but via different molecular mechanisms. Hum Mutat. 2007;28:639. doi:10.1002/()1098-1004.
- Richards AJ, Laidlaw M, Whittaker J, Treacy B, Rai H, Bearcroft P, Baguley DM, Poulson A, Ang A, Scott JD, et al. High efficiency of mutation detection in type 1 stickler syndrome using a two-stage approach: vitreoretinal assessment coupled with exon sequencing for screening COL2A1. Hum Mutat. 2006;27:696–704. doi:10.1002/humu.v27:7.
- Barbieri O, Astigiano S, Morini M, Tavella S, Schito A, Corsi A, Martino DD, Bianco P, Cancedda R, Garofalo S, et al. Depletion of cartilage collagen fibrils in mice carrying a dominant negative Col2a1 transgene affects chondrocyte differentiation. Am J Physiol Cell Physiol. 2003;285(6):C1504–12. doi:10.1152/ajpcell.00579.2002.
- Li SW, Prockop DJ, Helminen H, Fassler R, Lapvetelainen T, Kiraly K, Peltarri A, Arokoski J, Lui H, Arita M, et al. Transgenic mice with targeted inactivation of the Col2 alpha 1 gene for collagen II develop a skeleton with membranous and periosteal bone but no endochondral bone. Genes Dev. 1995;9(22):2821–30. doi:10.1101/gad.9.22.2821.
- Vandenberg P, Khillan JS, Prockop DJ, Helminen H, Kontusaari S, Ala-Kokko L. Expression of a partially deleted gene of human type II procollagen (COL2A1) in transgenic mice produces a chondrodysplasia. Proc Natl Acad Sci U S A. 1991;88(17):7640–44. doi:10.1073/pnas.88.17.7640.
- Metsaranta M, Garofalo S, Decker G, Rintala M, de Crombrugghe B, Vuorio E. Chondrodysplasia in transgenic mice harboring a 15-amino acid deletion in the triple helical domain of pro alpha 1(II) collagen chain. J Cell Biol. 1992;118:203–12. doi:10.1083/jcb.118.1.203.
- Garofalo S, Vuorio E, Metsaranta M, Rosati R, Toman D, Vaughan J, Lozano G, Mayne R, Ellard J, Horton W, et al. Reduced amounts of cartilage collagen fibrils and growth plate anomalies in transgenic mice harboring a glycine-to-cysteine mutation in the mouse type II procollagen alpha 1-chain gene. Proc Natl Acad Sci U S A. 1991;88:9648–52. doi:10.1073/pnas.88.21.9648.
- Savontaus M, Rintala-Jamsa M, Morko J, Ronning O, Metsaranta M, Vuorio E. Abnormal craniofacial development and expression patterns of extracellular matrix components in transgenic Del1 mice harboring a deletion mutation in the type II collagen gene. Orthod Craniofac Res. 2004;7:216–26. doi:10.1111/j.1601-6343.2004.00304.x.
- Lavrin IO, McLean W, Seegmiller RE, Olsen BR, Hay ED. The mechanism of palatal clefting in the Col11a1 mutant mouse. Arch Oral Biol. 2001;46:865–69. doi:10.1016/S0003-9969(01)00044-9.
- Li Y, Lacerda DA, Warman ML, Beier DR, Yoshioka H, Ninomiya Y, Oxford JT, Morris NP, Andrikopoulos K, Ramirez F, et al. A fibrillar collagen gene, Col11a1, is essential for skeletal morphogenesis. Cell. 1995;80(3):423–30. doi:10.1016/0092-8674(95)90492-1.
- McGuirt WT, Prasad SD, Griffith AJ, Kunst HP, Green GE, Shpargel KB, Runge C, Huybrechts C, Mueller RF, Lynch E, et al. Mutations in COL11A2 cause non-syndromic hearing loss (DFNA13). Nat Genet. 1999;23(4):413–19. doi:10.1038/70516.
- Li SW, Takanosu M, Arita M, Bao Y, Ren ZX, Maier A, Prockop DJ, Mayne R. Targeted disruption of Col11a2 produces a mild cartilage phenotype in transgenic mice: comparison with the human disorder otospondylomegaepiphyseal dysplasia (OSMED). Dev Dyn. 2001;222:141–52. doi:10.1002/()1097-0177.
- Halper J, Kjaer M. Basic components of connective tissues and extracellular matrix: elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins. Adv Exp Med Biol. 2014;802:31–47.
- Zollinger AJ, Smith ML. Fibronectin, the extracellular glue. Matrix Biol. 2017;60–61:27–37. doi:10.1016/j.matbio.2016.07.011.
- Astrof S, Crowley D, Hynes RO. Multiple cardiovascular defects caused by the absence of alternatively spliced segments of fibronectin. Dev Biol. 2007;311:11–24. doi:10.1016/j.ydbio.2007.07.005.
- Astrof S, Hynes RO. Fibronectins in vascular morphogenesis. Angiogenesis. 2009;12:165–75. doi:10.1007/s10456-009-9136-6.
- Liang D, Wang X, Mittal A, Dhiman S, Hou SY, Degenhardt K, Astrof S. Mesodermal expression of integrin alpha5beta1 regulates neural crest development and cardiovascular morphogenesis. Dev Biol. 2014;395:232–44. doi:10.1016/j.ydbio.2014.09.014.
- Wang X, Astrof S. Neural crest cell-autonomous roles of fibronectin in cardiovascular development. Development. 2016;143:88–100. doi:10.1242/dev.125286.
- Galloway JL, Jones SJ, Mossey PA, Ellis IR. The control and importance of hyaluronan synthase expression in palatogenesis. Front Physiol. 2013;4:10. doi:10.3389/fphys.2013.00010.
- Jarvinen TA, Jozsa L, Kannus P, Jarvinen TL, Hurme T, Kvist M, Pelto-Huikko M, Kalimo H, Järvinen M. Mechanical loading regulates the expression of tenascin-C in the myotendinous junction and tendon but does not induce de novo synthesis in the skeletal muscle. J Cell Sci. 2003;116:857–66. doi:10.1242/jcs.00303.
- Murphy-Ullrich JE. The de-adhesive activity of matricellular proteins: is intermediate cell adhesion an adaptive state? J Clin Invest. 2001;107:785–90. doi:10.1172/JCI12609.
- Sage EH. Regulation of interactions between cells and extracellular matrix: a command performance on several stages. J Clin Invest. 2001;107:781–83. doi:10.1172/JCI12683.
- Chiquet M, Blumer S, Angelini M, Mitsiadis TA, Katsaros C. Mesenchymal remodeling during palatal shelf elevation revealed by extracellular matrix and F-actin expression patterns. Front Physiol. 2016;7:392. doi:10.3389/fphys.2016.00392.
- Chen W, Kim MS, Shanbhag S, Arai A, VanRyzin C, McDonnell NB, Merke DP. The phenotypic spectrum of contiguous deletion of CYP21A2 and tenascin XB: quadricuspid aortic valve and other midline defects. Am J Med Genet Part A. 2009;149a:2803–08. doi:10.1002/ajmg.a.v149a:12.
- Merke DP, Chen W, Morissette R, Xu Z, Van Ryzin C, Sachdev V, Hannoush H, Shanbhag SM, Acevedo AT, Nishitani M, et al. Tenascin-X haploinsufficiency associated with Ehlers-Danlos syndrome in patients with congenital adrenal hyperplasia. J Clin Endocrinol Metab. 2013;98(2):E379–87. doi:10.1210/jc.2012-3148.
- Morissette R, Merke DP, McDonnell NB. Transforming growth factor-beta (TGF-beta) pathway abnormalities in tenascin-X deficiency associated with CAH-X syndrome. Eur J Med Genet. 2014;57:95–102.
- Takeshita S, Kikuno R, Tezuka K, Amann E. Osteoblast-specific factor 2: cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem J. 1993;294(Pt 1):271–78. doi:10.1042/bj2940271.
- Gillan L, Matei D, Fishman DA, Gerbin CS, Karlan BY, Chang DD. Periostin secreted by epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins and promotes cell motility. Cancer Res. 2002;62:5358–64.
- Kii I, Amizuka N, Minqi L, Kitajima S, Saga Y, Kudo A. Periostin is an extracellular matrix protein required for eruption of incisors in mice. Biochem Biophys Res Commun. 2006;342:766–72. doi:10.1016/j.bbrc.2006.02.016.
- Rios HF, Ma D, Xie Y, Giannobile WV, Bonewald LF, Conway SJ, Feng JQ. Periostin is essential for the integrity and function of the periodontal ligament during occlusal loading in mice. J Periodontol. 2008;79(8):1480–90. doi:10.1902/jop.2008.070624.
- Rios H, Koushik SV, Wang H, Wang J, Zhou HM, Lindsley A, Rogers R, Chen Z, Maeda M, Kruzynska-Frejtag A, et al. periostin null mice exhibit dwarfism, incisor enamel defects, and an early-onset periodontal disease-like phenotype. Mol Cell Biol. 2005;25:11131–44. doi:10.1128/MCB.25.24.11131-11144.2005.
- Kitase Y, Yamashiro K, Fu K, Richman JM, Shuler CF. Spatiotemporal localization of periostin and its potential role in epithelial-mesenchymal transition during palatal fusion. Cells Tissues Organs. 2011;193:53–63. doi:10.1159/000320178.
- Kruzynska-Frejtag A, Wang J, Maeda M, Rogers R, Krug E, Hoffman S, Markwald RR, Conway SJ. Periostin is expressed within the developing teeth at the sites of epithelial-mesenchymal interaction. Dev Dyn. 2004;229(4):857–68. doi:10.1002/()1097-0177.
- Yao Y. Laminin: loss-of-function studies. Cell Mol Life Sci. 2017;74:1095–115. doi:10.1007/s00018-016-2381-0.
- Miner JH, Yurchenco PD. Laminin functions in tissue morphogenesis. Annu Rev Cell Dev Biol. 2004;20:255–84. doi:10.1146/annurev.cellbio.20.010403.094555.
- Yamada T, Hirata A, Sasabe E, Yoshimura T, Ohno S, Kitamura N, Yamamoto T. TCDD disrupts posterior palatogenesis and causes cleft palate. J Cranio-maxillo-facial Surg. 2014;42:1–6. doi:10.1016/j.jcms.2013.01.024.
- Neptune ER, Frischmeyer PA, Arking DE, Myers L, Bunton TE, Gayraud B, Ramirez F, Sakai LY, Dietz HC. Dysregulation of TGF-beta activation contributes to pathogenesis in Marfan syndrome. Nat Genet. 2003;33:407–11. doi:10.1038/ng1116.
- Iozzo RV, Schaefer L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015;42:11–55. doi:10.1016/j.matbio.2015.02.003.
- Jakobsen LP, Borup R, Vestergaard J, Larsen LA, Lage K, Maroun LL, Kjaer I, Niemann CU, Andersen M, Knudsen MA, et al. Expression analyses of human cleft palate tissue suggest a role for osteopontin and immune related factors in palatal development. Exp Mol Med. 2009;41(2):77–85. doi:10.3858/emm.2009.41.2.010.
- Singh GD, Moxham BJ, Langley MS, Waddington RJ, Embery G. Changes in the composition of glycosaminoglycans during normal palatogenesis in the rat. Arch Oral Biol. 1994;39:401–07. doi:10.1016/0003-9969(94)90170-8.
- Brinkley LL, Morris-Wiman J. The role of extracellular matrices in palatal shelf closure. Curr Top Dev Biol. 1984;19:17–36.
- Singh GD, Moxham BJ, Langley MS, Embery G. Glycosaminoglycan biosynthesis during 5-fluoro-2-deoxyuridine-induced palatal clefts in the rat. Arch Oral Biol. 1997;42:355–63. doi:10.1016/S0003-9969(97)00031-9.
- Martinez-Alvarez C, Tudela C, Perez-Miguelsanz J, O’Kane S, Puerta J, Ferguson MW. Medial edge epithelial cell fate during palatal fusion. Dev Biol. 2000;220:343–57. doi:10.1006/dbio.2000.9644.
- Inman GJ, Nicolas FJ, Callahan JF, Harling JD, Gaster LM, Reith AD, Laping NJ, Hill CS. SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol Pharmacol. 2002;62:65–74. doi:10.1124/mol.62.1.65.
- Corsi A, Xu T, Chen XD, Boyde A, Liang J, Mankani M, Sommer B, Iozzo RV, Eichstetter I, Robey PG, et al. Phenotypic effects of biglycan deficiency are linked to collagen fibril abnormalities, are synergized by decorin deficiency, and mimic Ehlers-Danlos-like changes in bone and other connective tissues. J Bone Miner Res. 2002;17:1180–89. doi:10.1359/jbmr.2002.17.7.1180.
- Zhang YX, Mori T, Iseki K, Hagino S, Takaki H, Takeuchi M, Hikake T, Tase C, Murakawa M, Yokoya S, et al. Differential expression of decorin and biglycan genes during palatogenesis in normal and retinoic acid-treated mice. Dev Dyn. 2003;226(4):618–26. doi:10.1002/dvdy.10267.
- Hammond NL, Brookes KJ, Dixon MJ. Ectopic hedgehog signaling causes cleft palate and defective osteogenesis. J Dent Res. 2018;97:1485–93.
- Knudsen TB, Bulleit RF, Zimmerman EF. Histochemical localization of glycosaminoglycans during morphogenesis of the secondary palate in mice. Anat Embryol (Berl). 1985;173:137–42. doi:10.1007/BF00707312.
- Larsson KS. Studies on the closure of the secondary palate. IV. Autoradiographic and histochemical studies of mouse embryos from cortisone-treated mothers. Acta Morphol Neerl Scand. 1962;4:369–86.
- Witt RM, Hecht ML, Pazyra-Murphy MF, Cohen SM, Noti C, van Kuppevelt TH, Fuller M, Chan JA, Hopwood JJ, Seeberger PH, et al. Heparan sulfate proteoglycans containing a glypican 5 core and 2-O-sulfo-iduronic acid function as sonic hedgehog co-receptors to promote proliferation. J Biol Chem. 2013;288:26275–88. doi:10.1074/jbc.M112.438937.
- Rapraeger AC, Krufka A, Olwin BB. Requirement of heparan sulfate for bFGF-mediated fibroblast growth and myoblast differentiation. Science (New York, NY). 1991;252:1705–08. doi:10.1126/science.1646484.
- Matsuo I, Kimura-Yoshida C. Extracellular modulation of fibroblast growth factor signaling through heparan sulfate proteoglycans in mammalian development. Curr Opin Genet Dev. 2013;23:399–407. doi:10.1016/j.gde.2013.02.004.
- He H, Huang M, Sun S, Wu Y, Lin X. Epithelial heparan sulfate regulates sonic hedgehog signaling in lung development. PLoS Genet. 2017;13:e1006992. doi:10.1371/journal.pgen.1006992.
- Xu J, Liu H, Lan Y, Aronow BJ, Kalinichenko VV, Jiang R. A Shh-Foxf-Fgf18-Shh molecular circuit regulating palate development. PLoS Genet. 2016;12:e1005769. doi:10.1371/journal.pgen.1005769.
- Yayon A, Klagsbrun M, Esko JD, Leder P, Ornitz DM. Cell surface, heparin-like molecules are required for binding of basic fibroblast growth factor to its high affinity receptor. Cell. 1991;64:841–48. doi:10.1016/0092-8674(91)90512-W.
- Rice R, Spencer-Dene B, Connor EC, Gritli-Linde A, McMahon AP, Dickson C, Thesleff I, Rice DPC. Disruption of Fgf10/Fgfr2b-coordinated epithelial-mesenchymal interactions causes cleft palate. J Clin Invest. 2004;113:1692–700. doi:10.1172/JCI20384.
- Han J, Mayo J, Xu X, Li J, Bringas P Jr., Maas RL, Rubenstein JLR, Chai Y. Indirect modulation of Shh signaling by Dlx5 affects the oral-nasal patterning of palate and rescues cleft palate in Msx1-null mice. Development. 2009;136(24):4225–33. doi:10.1242/dev.036723.
- Snyder-Warwick AK, Perlyn CA, Pan J, Yu K, Zhang L, Ornitz DM. Analysis of a gain-of-function FGFR2 crouzon mutation provides evidence of loss of function activity in the etiology of cleft palate. Proc Natl Acad Sci U S A. 2010;107:2515–20. doi:10.1073/pnas.0913985107.
- Li C, Lan Y, Krumlauf R, Jiang R. Modulating Wnt signaling rescues palate morphogenesis in pax9 mutant mice. J Dent Res. 2017;96:1273–81. doi:10.1177/0022034517719865.
- Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002;110:673–87. doi:10.1016/S0092-8674(02)00971-6.
- Turner CJ, Badu-Nkansah K, Crowley D, van der Flier A, Hynes RO. alpha5 and alphav integrins cooperate to regulate vascular smooth muscle and neural crest functions in vivo. Development. 2015;142:797–808. doi:10.1242/dev.117572.
- Zhu J, Motejlek K, Wang D, Zang K, Schmidt A, Reichardt LF. beta8 integrins are required for vascular morphogenesis in mouse embryos. Development. 2002;129:2891–903.
- Tadokoro S, Shattil SJ, Eto K, Tai V, Liddington RC, de Pereda JM, Ginsberg MH, Calderwood DA. Talin binding to integrin beta tails: a final common step in integrin activation. Science (New York, NY). 2003;302:103–06. doi:10.1126/science.1086652.
- Calderwood DA, Fujioka Y, de Pereda JM, Garcia-Alvarez B, Nakamoto T, Margolis B, McGlade CJ, Liddington RC, Ginsberg MH. Integrin beta cytoplasmic domain interactions with phosphotyrosine-binding domains: a structural prototype for diversity in integrin signaling. Proc Natl Acad Sci U S A. 2003;100:2272–77. doi:10.1073/pnas.262791999.
- Senetar MA, McCann RO. Gene duplication and functional divergence during evolution of the cytoskeletal linker protein talin. Gene. 2005;362:141–52. doi:10.1016/j.gene.2005.08.012.
- Wu Q, Zhang J, Koh W, Yu Q, Zhu X, Amsterdam A, Davis GE, Arnaout MA, Xiong JW. Talin1 is required for cardiac Z-disk stabilization and endothelial integrity in zebrafish. FASEB J. 2015;29:4989–5005. doi:10.1096/fj.15-273409.
- Ishii K, Mukherjee K, Okada T, Liao EC. Genetic requirement of talin1 for proliferation of cranial neural crest cells during palate development. Prs-Glob Open. 2018;6:e1633.
- Monkley SJ, Zhou XH, Kinston SJ, Giblett SM, Hemmings L, Priddle H, Brown JE, Pritchard CA, Critchley DR, Fässler R. Disruption of the talin gene arrests mouse development at the gastrulation stage. Dev Dyn. 2000;219:560–74. doi:10.1002/1097-0177(2000)9999:9999<::AID-DVDY1079>3.0.CO;2-Y.
- Debrand E, Conti FJ, Bate N, Spence L, Mazzeo D, Pritchard CA, Monkley SJ, Critchley DR. Mice carrying a complete deletion of the talin2 coding sequence are viable and fertile. Biochem Biophys Res Commun. 2012;426(2):190–95. doi:10.1016/j.bbrc.2012.08.061.
- Morris-Wiman J, Burch H, Basco E. Temporospatial distribution of matrix metalloproteinase and tissue inhibitors of matrix metalloproteinases during murine secondary palate morphogenesis. Anat Embryol (Berl). 2000;202:129–41. doi:10.1007/s004290000098.
- Morris-Wiman J, Du Y, Brinkley L. Occurrence and temporal variation in matrix metalloproteinases and their inhibitors during murine secondary palatal morphogenesis. J Craniofac Genet Dev Biol. 1999;19:201–12.
- de Oliveira Demarchi AC, Zambuzzi WF, Paiva KB, da Silva-valenzuela M, Nunes FD, de Cassia Savio Figueira R, Sasahara RM, Demasi MAA, Winnischofer SMB, Sogayar MC, et al. Development of secondary palate requires strict regulation of ECM remodeling: sequential distribution of RECK, MMP-2, MMP-3, and MMP-9. Cell Tissue Res. 2010;340:61–69. doi:10.1007/s00441-010-0931-6.
- Mansell JP, Kerrigan J, McGill J, Bailey J, TeKoppele J, Sandy JR. Temporal changes in collagen composition and metabolism during rodent palatogenesis. Mech Ageing Dev. 2000;119:49–62. doi:10.1016/S0047-6374(00)00168-8.
- Smane-Filipova L, Pilmane M, Akota I. MMPs and TIMPs expression in facial tissue of children with cleft lip and palate. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2016;160:538–42. doi:10.5507/bp.2016.055.
- Blavier L, Lazaryev A, Groffen J, Heisterkamp N, DeClerck YA, Kaartinen V. TGF-beta3-induced palatogenesis requires matrix metalloproteinases. Mol Biol Cell. 2001;12:1457–66. doi:10.1091/mbc.12.5.1457.
- Brown NL, Yarram SJ, Mansell JP, Sandy JR. Matrix metalloproteinases have a role in palatogenesis. J Dent Res. 2002;81:826–30. doi:10.1177/154405910208101206.
- Shi J, Son MY, Yamada S, Szabova L, Kahan S, Chrysovergis K, Wolf L, Surmak A, Holmbeck K. Membrane-type MMPs enable extracellular matrix permissiveness and mesenchymal cell proliferation during embryogenesis. Dev Biol. 2008;313:196–209. doi:10.1016/j.ydbio.2007.10.017.
- Brown GD, Nazarali AJ. Matrix metalloproteinase-25 has a functional role in mouse secondary palate development and is a downstream target of TGF-beta3. BMC Dev Biol. 2010;10:93. doi:10.1186/1471-213X-10-93.
- Kondo H, Matsushita I, Nagata T, Hayashi T, Kakinoki M, Uchio E, Kondo M, Ohji M, Kusaka S. Novel mutations in the COL2A1 gene in Japanese patients with Stickler syndrome. Hum Genome Var. 2016;3(1):16018. doi:10.1038/hgv.2016.18.
- Dixon MJ, Marazita ML, Beaty TH, Murray JC. Cleft lip and palate: understanding genetic and environmental influences. Nat Rev Genet. 2011;12:167–78. doi:10.1038/nrg2933.
- Kannu P, Bateman J, Savarirayan R. Clinical phenotypes associated with type II collagen mutations. J Paediatr Child Health. 2012;48:E38–43. doi:10.1111/j.1440-1754.2010.01979.x.
- Fairfield H, Gilbert GJ, Barter M, Corrigan RR, Curtain M, Ding Y, D’Ascenzo M, Gerhardt DJ, He C, Huang W, et al. Mutation discovery in mice by whole exome sequencing. Genome Biol. 2011;12:R86. doi:10.1186/gb-2011-12-9-r86.
- Miller WL, Merke DP. Tenascin-X, congenital adrenal hyperplasia, and the CAH-X syndrome. Horm Res Paediatr. 2018;89:352–61. doi:10.1159/000481911.
- O’Connell M, Burrows NP, van Vlijmen-willems MJ, Clark SM, Schalkwijk J. Tenascin-X deficiency and Ehlers-Danlos syndrome: a case report and review of the literature. Br J Dermatol. 2010;163:1340–45. doi:10.1111/j.1365-2133.2010.09949.x.
- Kimata K, Barrach HJ, Brown KS, Pennypacker JP. Absence of proteoglycan core protein in cartilage from the cmd/cmd (Cartilage matrix deficiency) mouse. J Biol Chem. 1981;256:6961–68.
- Krueger RC Jr., Kurima K, Schwartz NB. Completion of the mouse aggrecan gene structure and identification of the defect in the cmd-Bc mouse as a near complete deletion of the murine aggrecan gene. Mamm Genome. 1999;10:1119–25. doi:10.1007/s003359901176.
- Sasaki S, Kurisu K. Effect of triamcinolone acetonide on proliferation and collagen and glycosaminoglycan syntheses in palatal mesenchymal cells from the mouse fetus. J Craniofac Genet Dev Biol. 1983;3:351–69.
- Foreman DM, Sharpe PM, Ferguson MW. Comparative biochemistry of mouse and chick secondary-palate development in vivo and in vitro with particular emphasis on extracellular matrix molecules and the effects of growth factors on their synthesis. Arch Oral Biol. 1991;36:457–71. doi:10.1016/j.ejmg.2013.12.004.
- Jin JZ, Tan M, Warner DR, Darling DS, Higashi Y, Gridley T, Ding J. Mesenchymal cell remodeling during mouse secondary palate reorientation. Dev Dyn. 2010;239:2110–17. doi:10.1002/dvdy.22339.