ABSTRACT
‘Crimson Seedless’ is a table grape (Vitis vinifera) cultivar that often fails to develop adequate color, particularly in warm climate regions such as the San Joaquin Valley, California. Growers generally apply ethephon to ‘Crimson Seedless’ grapes to improve their color, but results are erratic. The plant hormone abscisic acid (ABA) stimulates anthocyanin production in some other varieties of grape, so applications of ABA might help improve the color of ‘Crimson Seedless’. Application of ABA at veraison maximized anthocyanin accumulation, but later applications also increased pigmentation and, at the same time, reduced ABA's negative effect on berry puncture resistance. Concentrations of 150 and 300 mg·L− 1 ABA increased anthocyanin levels more than did 75 mg·L− 1 ABA. The relationships between anthocyanin content of the berry skins and the color characteristics lightness (L*), and hue angle (h°), were well described by the functions L* = 22.56x− 0.0865, and h° = 3.356x− 0.513, where x = skin anthocyanin content. Thus, at skin anthocyanin contents ≥ 0.03 mg·cm− 2, relatively large increases in anthocyanin content have relatively small effects on L* and h° of ‘Crimson Seedless’ grapes. Similar results have been obtained with other cultivars of grape. For this reason, we suggest to include direct color measurements, rather than relying only on anthocyanin content when evaluating color on grapes.
INTRODUCTION
The ‘Crimson Seedless’ table grape was released in 1989 by the USDA and has since become popular among table grape growers worldwide. Unfortunately, in California and other warm-climate regions, ‘Crimson Seedless’ grapes often fail to develop adequate color because high temperatures impair anthocyanin accumulation in grape berry skins (CitationSpayd et al., 2002). ‘Crimson Seedless’ grapes have a long ripening period, so they mature late in the season and the berries are thus exposed to high temperatures at a critical stage for color accumulation. These high temperatures and the length of the maturation period could help explain why the berries of this cultivar are often more difficult to color than ‘Flame Seedless’ or ‘Redglobe’, which usually mature before ‘Crimson Seedless’.
We have demonstrated that application of abscisic acid to ‘Flame Seedless’ and ‘Redglobe’ grapes could improve their color, but efficacy varied according to the cultivar and the timing and concentration of the ABA solutions applied (CitationPeppi et al., 2006, 2007). Recently, CitationCantin et al. (2007) demonstrated that application of 150 or 300 mg·L−1 ABA to ‘Crimson Seedless’ at veraison improved berry color, but the efficacy of other application times or concentrations of ABA was not tested. Moreover, CitationCantin et al. (2007) did not determine the effects of ABA on skin anthocyanin content; a quality variable that is more widely reported than color when evaluating the quality of grapes. Therefore, the primary objective of this research was to determine the effects of different ABA concentrations, application times, and harvest dates, on the quality of ‘Crimson Seedless’ grapes, especially with respect to the puncture resistance, pigmentation, and the color of the berries. A secondary objective was to characterize the relationship between skin anthocyanin levels and the color of ‘Crimson Seedless’ grapes.
MATERIALS AND METHODS
Plant Material
The studies were conducted on own-rooted ‘Crimson Seedless’ grapevines (V. vinifera L.) planted in 1997 at the Kearney Agricultural Center, Parlier, CA. Vines were spaced 2.4 m within rows and 3.6 m between rows and supported by an open gable trellis. The vines used in 2004 were trained to quadrilateral cordons and spur-pruned, whereas those used in 2005 were bilateral cordon trained and cane-pruned. The vineyard was drip irrigated and standard cultural practices were followed every year, with the exception of ethephon applications that were not applied in 2004 but were included as a treatment in 2005. Crop load was regulated taking into account initial cluster number and cluster size. Vines of similar vigor were chosen for each of the experiments.
Experimental Design and Treatments
Abscisic acid concentrations tested were 0, 75, 150, and 300 mg·L− 1 ABA. In 2005, 250 mg L−1 ethephon was included as a PGR treatment. Within each vine, grape clusters were randomly assigned to receive treatments composed of two factors; timing of ABA application and PGR treatment. In 2004 four application times were tested (28 May [preveraison], 28 June [veraison], 26 July [postveraison I] or 23 August [postveraison II]); while in 2005 three application times were tested (11 July [veraison], 8 August [postveraison I], and 5 September [postveraison II]). The first harvest was after soluble solids reached 16.5° Brix and at least one of the treatments had well-colored fruits. The third harvest of 2004 was intended to occur at the end of October, but the fruits were harvested sooner due to impending rain. Grapevines were assigned to one of three harvest dates (1 September, 30 September, or 4 October in 2004 and 2 August, 6 September, or 6 October in 2005) in a randomized complete block design in 2004 (application timing and concentration factors were nested within harvest dates) and in a split-split plot design in 2005 (the main plot factor was harvest date, the subplot was application time, and the sub-subplot was PGR treatment). Both seasons there were six replicates. Treatments were applied with a hand-held sprayer until runoff and plastic shields prevented overspray or runoff from contacting other clusters.
Data were subjected to the general linear models procedure (PROC GLM) of SAS statistical software (SAS Inst., Cary, NC). When interactions between factors were significant (P = 0.05), the effect of each factor was determined by separating the means by Duncan's New Multiple Range Test (DNMRT), or by Least Significant Difference (LSD), as appropriate (CitationGomez and Gomez, 1984).
Berry Sampling and Fruit Analyses
At harvest, 50 randomly selected berries per cluster were removed, weighed and then macerated for 5 min in a blender. The filtered juice was used for soluble solids and titratable acidity measurements as described in CitationPeppi et al. (2006). Additional 10 randomly selected berries were removed from each cluster by clipping their pedicels with shears. With a razor blade, a small area of equatorial flesh was exposed on each berry, and mesocarp puncture resistance measurements, indicators of firmness (CitationJensen et al., 1975), were made as described previously (CitationPeppi et al., 2006).
Anthocyanin Measurements
Ten additional berries from each cluster were collected, frozen and kept at − 20°C for anthocyanin measurements as described previously (CitationPeppi et al. 2006). Anthocyanins from skin disks from two opposite points at the berry equator were extracted in darkness at room temperature in methanol (10 mL·g− 1 skin fresh weight) for 48 hours (when no red pigment was observed in the skins), extracts were mixed, and a 2-ml aliquot was acidified with HCl to adjust the pH to 1.5 for maximum absorption determined at 520 nm using a spectrophotometer (Milton Roy 601, Spectronic, Rochester, NY). Data were expressed as mg of total anthocyanins as malvidin-3-glucoside per area of skin in cm2.
Color Measurements
There are many objective color measurement systems but the one with lightness (L*), chroma (C*), and hue (h°) is recommended by various researchers working on color of fruits and vegetables to characterize its appearance (CitationMcGuire, 1992; CitationWrolstad et al., 2005), and these variables have already been used in various species (CitationGazula et al., 2007; CitationNgo et al., 2007) and other grape cultivars (CitationPeppi et al., 2006, 2007) for color comparisons.
In 2005 measurements of L*, C*, and h° were made at three equidistant points at the equator of the berries collected for anthocyanin analyses, prior to freezing, with a colorimeter (CR-200, Minolta Inc., Ramsey, NJ). The colorimeter was calibrated following the manufacturer instructions, under the illuminant C and 2° the observer angle. Anthocyanin and color data were subjected to regression analyses by SigmaPlot (Systat Software Inc., Richmond, CA).
RESULTS
Experiments 2004
All the variables measured in 2004 differed between harvest dates as expected (data not shown). None of the ABA treatments affected berry weight. On the first harvest date titratable acidity was reduced from 5.35 to 5.00 g·L− 1 in fruit treated with ABA at or after veraison compared to preveraison applications, otherwise ABA did not affect fruit composition. Fruit puncture resistance was not affected by ABA treatments on any harvest date (data not shown).
At the first harvest date, berry skin anthocyanin levels were highly variable and did not differ between treatments if P = 0.05 (data not shown). However, using a slightly less conservative criteria, P = 0.07, fruits treated with ABA at veraison or one month after had twice the anthocyanin content as fruits treated preveraison or two months after veraison (data not shown). By the second harvest, 300 mg·L− 1 ABA applied at or after veraison more than doubled skin anthocyanins () and on the third harvest, 150 or 300 mg·L− 1 ABA applied at veraison, or eight weeks after, increased anthocyanins to levels that were two or three times greater than that of controls ().
TABLE 1. Effect of abscisic acid (ABA) application times and concentrations on ‘Crimson Seedless’ berry skin anthocyanins on 30 September 2004
TABLE 2. Effect of abscisic acid (ABA) or ethephon application times and concentrations on ‘Crimson Seedless’ berry skin anthocyanins on 4 October 2004
Experiments 2005
In 2005 berry weight and fruit composition changed with harvest date as expected (data not shown). Because ‘Crimson Seedless’ has a long maturity phase the application times and the harvest dates intervals were widely spaced (one month each), and the second and third applications were not made until after the first and second harvests. Thus, application time and harvest date usually interacted to affect most variables as expected. There was no consistent effect of ABA on berry weight (data not shown). In general soluble solids and pH were not affected by ABA treatments, but titratable acidity was slightly reduced by the veraison application (data not shown).
Berries were as resistant to puncture at the first harvest as they were on subsequent harvests (data not shown). High concentrations of ABA (150 or 300 mg·L− 1) applied at veraison reduced puncture resistance more than ethephon did (). However, when applied four weeks after veraison ABA and ethephon decreased berry puncture resistance to the same extent compared to untreated fruits. None of the PGRs applied eight weeks after veraison affected berry puncture resistance. The PGRs tested reduced puncture resistance equally whether they were applied at veraison or four weeks after, except for 300 mg·L−1 ABA, which caused more softening when applied at veraison than when applied four weeks after.
TABLE 3. Effect of abscisic acid (ABA) and ethephon application times and concentrations on ‘Crimson Seedless’ mesocarp puncture resistance in 2005
Anthocyanins increased with each harvest (data not shown). At the first harvest 150 and 300 mg·L−1 ABA applied at veraison were the only treatments that increased the berry skin anthocyanin content compared to the control (). At the second harvest, the only treatments that increased anthocyanins were 150 and 300 mg·L− 1 ABA applied at veraison or four weeks after. On the third harvest the only veraison treatment that still increased anthocyanins was 300 mg·L− 1 ABA, but 150 and 300 mg·L− 1 ABA applied four weeks after veraison also increased anthocyanins. Among the treatments applied eight weeks after veraison, only 300 mg·L−1 ABA had an effect on total anthocyanins compared to the control. Ethephon applied at any time, and evaluated at any harvest, was never superior to any ABA treatment or to the control fruit in the anthocyanin content.
TABLE 4. Effect of abscisic acid (ABA) and ethephon application times and concentrations on ‘Crimson Seedless’ berry skin anthocyanins at three harvest dates in 2005
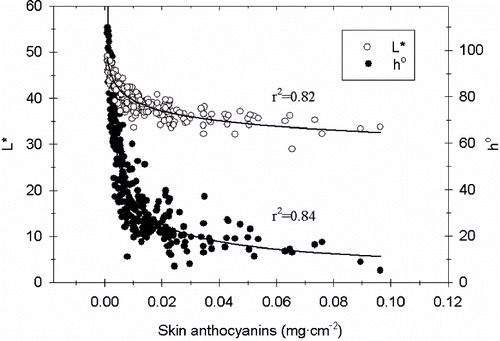
Regardless of harvest date, fruits treated at veraison with any PGR had lower L* values, and were thus more dark-colored, than nontreated fruits (). When treated one month after veraison, only fruits subjected to >150 mg·L− 1 ABA or ethephon had a darker color than control fruits, and, two months after veraison, only fruits treated with > 150 mg·L−1 ABA were darker than control fruits. The most effective treatment in darkening the fruit was 300 mg·L−1 ABA applied at veraison. The same concentration of ABA applied later also reduced fruit L*, but the effect was diminished compared to the earlier treatment. Chroma values ranged from 9.72 to 11.11 and only decreased in ethephon-treated fruit (data not shown). Chroma values tend to increase with pigment concentration; however, as skin berry color shifts to a darker coloration, chroma decreases (CitationWrolstad et al., 2005). Applications of ABA or ethephon at veraison, or four weeks after, decreased the h° (increased the red color) of the berries (), but any application eight weeks after veraison did not affect the h°. At veraison the best treatments were 150 and 300 mg·L− 1 ABA, followed by ethephon, but the same ABA concentrations applied four weeks later reduced the h° to the same extent as ethephon. There was an inverse, curvilinear relationship between the content of anthocyanins in the berries' skins and the fruits' L* and h° (), but there was no obvious relationship between anthocyanin content and C* (data not shown). As skin anthocyanin levels accumulated beyond about 0.03 mg·cm− 2 the relatively large changes in skin anthocyanin content had relatively small effects on L* and h°.
TABLE 5. Effect of different abscisic acid (ABA) and ethephon application times and concentrations on the lightness (L*) and hue angle (h°) of ‘Crimson Seedless’ berries in 2005
FIGURE 1. Relationship between anthocyanins per area and L* (lightness) and h° (hue angle) on ‘Crimson Seedless’ berries. L* and h° curves were described by y = axb. For L* curve a = 22.56 and b = − 0.08647, for h° curve a = 3.356 and b = − 0.5135.
DISCUSSION
Effects of ABA on berry weight were small and inconsistent, as observed by others (CitationLee et al., 1997). Recent experiments with ‘Flame Seedless’ and ‘Redglobe’ (CitationPeppi et al., 2006, 2007) showed that ABA treatments affected berry weight in some years but not all. Fruit composition was usually not affected by ABA, which agrees with other studies (CitationJeong et al., 2004; CitationLee et al., 1997; CitationPeppi et al., 2006, 2007) in which ABA had few or no effects on the soluble solids or titratable acidity of the berries.
Firm berries are a very important characteristic of quality table grapes. Ethephon is known to cause berry softening, which is undesirable (CitationJensen et al., 1975). Previously we found that ABA-induced softening of ‘Flame Seedless’ and ‘Redglobe’ grapes (CitationPeppi et al., 2006; 2007), and here we show that applications of ABA at veraison to ‘Crimson Seedless’ can cause as much or more softening as ethephon. This finding is in contrast with those of CitationCantin et al. (2007) who reported that ABA treatment did not affect the firmness of ‘Crimson Seedless’ berries. However, CitationCantin et al. (2007) treated all the clusters on a vine with ABA rather than certain individual clusters, so their population of berries within each plot probably contained a wider range of physiological ages than we had in the present study. This is important because our data indicate that when applied postveraison, fruits treated with ABA or ethephon were of similar firmness. Also, CitationCantin et al. (2007) collected berry samples from clusters of grapes after berries with quality defects were removed. This process likely eliminated berries that were excessively soft. If so, it suggests that the data here might overestimate the degree of softening caused by ABA; as soft berries, if not too numerous, could be clipped off of the cluster before reaching consumers.
Though ethephon and ABA both decreased fruit puncture resistance, ABA was more effective than ethephon at increasing anthocyanins in the skins of ‘Crimson Seedless’ grapes, confirming the findings of CitationCantin et al. (2007). Particularly interesting is the striking difference in the anthocyanin content between ABA-treated and ethephon-treated ‘Crimson Seedless’ fruits compared to ‘Flame Seedless’ (CitationPeppi et al., 2006) and ‘Redglobe’ (CitationPeppi et al., 2007). Furthermore, the ABA-timing effect differs considerably between ‘Crimson Seedless’ and these other cultivars. A review of the literature suggests that the efficacy of ABA varies between different grape cultivars, as we observed in our studies. For example, CitationKim et al. (1998) observed that ABA increased anthocyanins in several grape cultivars, but the same treatments decreased anthocyanins in others, and CitationJeong et al. (2004) found that ABA treatment caused a transient increase in anthocyanins of Cabernet Sauvignon that did not last until harvest. The effects of ABA on anthocyanin accumulation might also depend on environmental conditions, which could explain some of the observed annual variation in efficacy. In contrast, ethephon is often not effective enough to obtain good packable yields of ‘Crimson Seedless’ as suggested by our results 2005 (), and by the data of others (CitationDokoozlian et al., 1994).
Though ABA applications at veraison were the most effective at enhancing pigmentation and color, later applications between 150 and 300 mg·L−1 may be best for ‘Crimson Seedless’ because they still improve color but have less effect on berry puncture resistance. Besides the optimization in terms of anthocyanins, puncture resistance and general fruit quality, the effectiveness of late applications holds another great benefit in practical terms; growers could wait until relatively late in the season to assess whether or not their fruit are coloring well before spending the money on a PGR application.
The relationship between anthocyanins and color is not linear as observed previously with ‘Flame Seedless’ and ‘Redglobe’ (CitationPeppi et al., 2006, 2007). As observed for other grape cultivars L* and h° decreased sharply as anthocyanins increased up to about 0.01 mg·cm− 2 and then continued decreasing more gradually up to 0.03 mg cm− 2. As anthocyanins accumulated beyond that level, they had relatively less effect on L*, and especially on h°. The shapes of these curves validate color measurements as an important tool when evaluating the effect of cultural practices on table grape color, as relatively large differences in skin pigment content may have small effects on berry color.
CONCLUSIONS
The use of ABA to improve the color of ‘Crimson Seedless’ grapes may have commercial application because it is more effective than ethephon. However, care must be taken in the use of ABA because it can also cause more softening than ethephon. One solution may be to apply 300 mg·L− 1 ABA about one month after veraison, when it can still improve color without causing excessive softening. The curvilinear relationship between pigments, L*, and h° suggests again that color measurements should be collected more often by researchers when evaluating the effects of cultural practices on the color of grapes. These data are quick and easy to collect, are nondestructive and may be more repeatable than anthocyanin data.
Acknowledgments
This research was supported in part by grants from the California Table Grape Commission and Valent BioSciences. We thank Kimberley Cathline, Nona Ebisuda, Don Katayama, Karin Kawagoe, and Jorge Osorio for technical assistance and Andrew Walker and Doug Adams for their helpful comments on the manuscript.
Notes
Dokoozlian, N.K., D.A. Luvisi, P.L. Schrader, and M.M. Moriyama.1994. Influence of trunk girdle timing and ethephon on the quality of ‘Crimson Seedless’ table grapes, pp. 237–240. In: International Symposium on Table Grape Production Proceedings Anaheim. Amer. Soc. Enol. Viticult.
LITERATURE CITED
- Cantin , C.M. , Fidelibus , M.W. and Crisosto , C.H. 2007 . Application of abscisic acid (ABA) at veraison advanced red color development and maintained postharvest quality of ‘Crimson Seedless’ grapes . Postharvest Biol. Technol. , 46 : 237 – 241 .
- Dokoozlian, N.K., D.A. Luvisi, P.L. Schrader, and M.M. Moriyama.1994. Influence of trunk girdle timing and ethephon on the quality of ‘Crimson Seedless’ table grapes, pp. 237–240. In: International Symposium on Table Grape Production Proceedings Anaheim. Amer. Soc. Enol. Viticult.
- Gazula , A. , Kleinhenz , M.D. , Scheerens , J.C. and Ling , P.P. 2007 . Anthocyanin levels in nine lettuce (Lactuca sativa) cultivars: Influence of planning date and relations among analytic, instrumented, and visual assessments of color . HortScience , 42 : 232 – 238 .
- Gomez , K.A. and Gomez , A.A. 1984 . Statistical Procedures for Agricultural Research. , 2nd , New York : Wiley .
- Jensen , F.L. , Kissler , J. , Peacock , W. and Leavitt , G. 1975 . Effect of ethephon on color and fruit characteristics of Tokay and Emperor table grapes . Amer. J. Enol. Viticult. , 26 : 79 – 81 .
- Jeong , S.T. , Goto-Yamamoto , N. , Kobayashi , S. and Esaka , M. 2004 . Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins . Plant Sci. , 167 : 247 – 252 .
- Kim , S.K. , Kim , J.T. , Jeon , S.H. , Nam , Y.S. and Kim , S.H. 1998 . Effects of ethephon and ABA application on coloration, content and composition of anthocyanin in grapes (Vitis spp.). . J. Kor. Soc. Hort. Sci. , 39 : 547 – 554 .
- Lee , K.S. , Lee , J.C. , Hwang , Y.S. and Hur , I.B. 1997 . Effects of natural type (S)-(+)-abscisic acid on anthocyanin accumulation and maturity in ‘Kyoho’ grapes . J. Kor. Soc. Hort. Sci. , 38 : 717 – 721 .
- McGuire , R.G. 1992 . Reporting of objective color measurements . HortScience , 27 : 1254 – 255 .
- Ngo , T. , Wrolstad , R.E. and Zhao , Y. 2007 . Color quality of Oregon strawberries- Impact of genotype, composition, and processing . J. Food Sci. , 72 : 25 – 32 .
- Peppi , M.C. , Fidelibus , M.W. and Dokoozlian , N. 2006 . Abscisic acid application timing and concentration affect firmness, pigmentation and color of ‘Flame Seedless’ grapes . HortScience , 41 : 1440 – 1445 .
- Peppi , M.C. , Fidelibus , M.W. and Dokoozlian , N. 2007 . Application timing and concentration of abscisic acid affect the quality of ‘Redglobe’ grapes . J. Hort. Sci. Biotech. , 82 : 304 – 310 .
- Spayd , S.E. , Tarara , J.M. , Mee , D.L. and Ferguson , J.C. 2002 . Separation of sunlight and temperature effects on the composition of Vitis vinifera cv . Merlot berries. Amer. J. Enol. Viticult. , 53 : 171 – 182 .
- Wrolstad , R.E. , Durst , R.W. and Lee , J. 2005 . Tracking color and pigment changes in anthocyanin products . Trends Food Sci. Technol. , 16 : 423 – 428 .