Abstract
The USDA-ARS breeding program has identified a recessively-controlled genetic variant in highbush blueberry (V. corymbosum L.) that results in parthenocarpic fruit development. Current studies are underway to refine our understanding of this variant, and to move this trait into desirable and useful genetic backgrounds. In rabbiteye blueberry (V. ashei Reade), a species with critical cross-pollination requirements, significant variation in ability to set fruit without insect pollination has been observed under greenhouse conditions. A current study is evaluating linkages between greenhouse and field behavior of rabbiteye cultivars with regard to fruit set and development, with the long-term goal of enhancing productivity. Breeders often select naturally for cultivars with parthenocarpic tendencies, but improved understanding of the sources of parthenocarpy, and their genetic control will facilitate further advances in this area.
INTRODUCTION
Parthenocarpy in any fruit-bearing crop is a very desirable trait because it holds the possibility of reduced concerns about many typical pollination worries. In a cross-pollinated crop like blueberry, parthenocarpy could address at least three pollination concerns: (1) sub-optimal pollination weather during the bloom period, (2) declines of pollinators due to parasitic infections, and (3) desirability of cross-pollination versus self-pollination for optimum yield.
Normal fruit development is initiated by gibberellins or auxins generated by the developing ovule. These compounds stimulate early fruit development and also promote fruit set (i.e., the prevention of abscission) (CitationCrane, 1964). In some cases, parthenocarpy, the development of fruit without seed, may occur. In parthenocarpic genotypes, pollination either is not needed, or if it does occur, it does not result in fertilization and seed development. Natural parthenocarpy has been documented in various crops, including banana (CitationGustafson, 1939), grape (CitationWong, 1941), mandarins (CitationTalon et al., 1992), opuntia (CitationWeiss et al., 1993), and tomato (CitationGroot et al. 1987; CitationWong, 1941). Furthermore, considerable work has been conducted on the induction of parthenocarpic fruit using growth regulators in a wide variety of horticultural crops either to study fruit development or to evaluate potential utilization in production. Among such crops are cucumber (CitationKim et al., 1994a, Citation1994b), grape (CitationFellman et al., 1991), kiwi fruit (CitationOhara et al., 1997), pears (CitationYamada et al., 1991), pepino (CitationErcan and Akilli, 1996), strawberry (CitationMudge et al., 1981), and watermelon (CitationPak, 1993).
In blueberry, limited work has been done, but several groups have demonstrated that normal fruit development exhibits high levels of gibberellic acid (GA) production during early stages of development in both highbush and lowbush blueberries (CitationKender and Desroches, 1970; CitationMainland and Eck, 1968, Citation1971). In addition, it has been shown that parthenocarpy may be induced by GA3 in cases of pollination failure (CitationCano-Medrano and Darnell, 1998; CitationMainland and Eck, 1969; CitationNesmith et al., 1995). Several authors have noted what appear to be tendencies toward parthenocarpy in blueberry (i.e., some parthenocarpic fruit, low seed numbers, high pulp to seed ratios) (CitationEhlenfeldt, 2001; CitationEhlenfeldt and Hall, 1996; CitationHarrison et al. 1994; CitationMacKenzie, 1997).
RESULTS AND DISCUSSION
Parthenocarpy Model in Highbush Blueberry
Previously, we developed a genetic model of parthenocarpy in progeny derived from the northern highbush (V. corymbosum L.) clone, G-176 (CitationEhlenfeldt and Vorsa, 2007). This model postulates that parthenocarpy is the result of a homozygous, single-gene, recessive allele at a locus with incomplete dominance. In the model, PP_ _ genotypes yield normally-seeded fruit, Pppp genotypes yield “diminished seed” fruit (i.e., fewer and smaller seed), and pppp genotypes are parthenocarpic (). The major obstacles to the further use of this germplasm were that the presumed fully recessive types, while parthenocarpic, also appeared to suffer from low overall productivity. The diminished-seed types (simplex heterozygotes), however, were often relatively large-fruited and considerably more productive. We have subsequently endeavored to do two things: (1) to further verify the inheritance of this character primarily by evaluating crosses of diminished-seed x diminished-seed phenotypes (Pppp x Pppp), which should yield high frequencies of both diminished-seed phenotypes (50%) and parthenocarpic phenotypes (25%), and (2) crosses of diminished-seed phenotypes x F1s (Pppp x PPpp), where the F1s have been selected primarily for productivity and vigor. These should also yield relatively high frequencies of diminished-seed (42%) and parthenocarpic phenotypes (8%), and, hopefully, some of them will benefit from the parental selection for vigor.
FIGURE 1 Ranges of seed development expressed in families segregating for parthenocarpy: (a) normal, (b) diminished-seed type, and (c) parthenocarpic type (scale in cm). (Source: CitationEhlenfeldt and Vorsa, 2007).

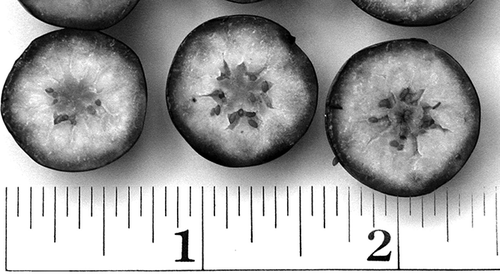
Since the parthenocarpic allele was initially found in the domesticated gene pool, we believe that this or similar alleles circulate in the highbush gene pool and that breeders often, directly or indirectly, select for these alleles. Several of the hybrids initially produced for the inheritance study had the parentage ‘Duke’ x G-176 (G-176 was the original parthenocarpic clone). Two of these plants have set large fruit under field conditions that (in retrospect) we would characterize as diminished-seed types (Pppp) (). This implies that ‘Duke’ may already carry some dosage of these recessive parthenocarpic alleles and could be of the genotype PPPp, PPpp, or Pppp, and, thus, may have contributed a gamete of the composition Pp in the cross generating these two hybrids. This concept is supported by the finding of CitationDraper and Scott (1969) that large fruit size is inherited recessively.
FIGURE 2 Fruit and seed development in an offspring of ‘Duke’ × G-176 (scale in inches).
Advancing the p Allele in the Hexaploid Gene Pool
A number of years ago, we crossed a synthetic 6x southern highbush (shb) clone, NJ 89–158–6, with G–176 (the original parthenocarpic clone). From this cross, we selected US 1232, a probable pentaploid, that was not notable except that it produced some fruit and was relatively vigorous. Under a conservative assumption, the allelic composition of US 1232 is PPPpp, and it might, if it produces balanced and randomly assorted 3x gametes, produce all of the following: PPP gametes (10% relative frequency), PPp gametes (60% relative frequency), and Ppp gametes (30% relative frequency). US 1232 has been used as a male with clones of rabbiteye (re), V. constablaei Gray (6x con), US 865 and US 872 (6x shb/con), and US 1056 (6x shb/con/re). US 1056 already seems to express parthenocarpic tendencies (high fruit set, high yield, low seediness), although its exact nature is unstudied. Conservatively, the best-case allelic composition of offspring from crosses utilizing US 1232 with any of these selections could be PPP/Ppp (30% frequency). Obviously, further backcrossing or sib-mating among these clones would probably be needed to recover a genotype expressing the parthenocarpic phenotype, but a single generation might be sufficient to recover the desired types, depending upon the relative effects of incomplete dominance within a hexaploid genome.
Parthenocarpy among Rabbiteye Blueberry Cultivars
Under greenhouse conditions, several rabbiteye blueberry cultivars regularly set fruit and develop it to maturity without pollination. Among the more notable cultivars exhibiting this behavior are ‘Baldwin’ and ‘Premier’. Since self-fruitfulness (or lack of) is a critical issue for rabbiteye, and a potential concern for northern-adapted rabbiteye types, we undertook a study of the relative propensity of 48 potted rabbiteye cultivars to set fruit under greenhouse conditions. We measured fruiting interval, fruit set, ripening interval, and fruit size, and also monitored temperature conditions during the study with the goal of not only documenting the performance of different cultivars, but also understanding as much as possible about the conditions leading to parthenocarpy. This study is in its third and (hopefully) final year.
Although the data have not yet been fully analyzed, a few cultivars stand out as having distinct ranges of parthenocarpy when not pollinated. Three older cultivars, ‘Early May’, ‘Ethel’, and ‘Owen’ have virtually no fruit-set without pollination. ‘Baldwin’, ‘Premier’, and ‘Maru’, three modern cultivars, have relatively large fruit at maturity (10–12 mm), but modest overall set (). In contrast, ‘Chaucer’, a modern Florida cultivar, has extremely high fruit-set without pollination, but its mature fruit are relatively small, typically 5–7 mm (). Is there an inverse relationship between parthenocarpic fruit size and degree of fruit set? Probably not. ‘Chaucer’ is unique in its relatively high fruit set, but it is easy to find cultivars with low set and small fruit (i.e., not an inverse relationship).
FIGURE 3 Parthenocarpic fruit development in (a) ‘Baldwin’ and (b) ‘Chaucer’ rabbiteye cultivars under insect-free greenhouse conditions.

The most critical issue with this experiment is how greenhouse data will relate to field results. To address this, we are working with Dr. Stephen Stringer of the USDA in Poplarville, Mississippi to compare parameters of field-grown fruit (size, weight, seed/pulp ratios) to see if cultivars with superior parthenocarpic performance in the greenhouse also exhibit less need for pollination to achieve superior yields under field conditions. In a future phase, we expect to also examine self-fertility in relation to parthenocarpy and field fruit development. Prospectively, we have made crosses of the high set/small fruit size cultivar ‘Chaucer’ with low set/large fruit size cultivars (‘Powderblue’, ‘Maru’) to further evaluate inheritance of these two factors.
The Value of Parthenocarpy Beyond the Obvious
Blueberry breeders of several generations have had interest in hybridizing highbush and rabbiteye, and transferring genes between them. This interest has resulted in many southern highbush cultivars with rabbiteye germplasm in their background. A few rabbiteye cultivars possess non-rabbiteye germplasm (‘Snowflake’, ‘Pink Lemonade’), but to date little outside germplasm has been utilized in rabbiteye breeding. Many breeders freely acknowledge that mainstream rabbiteye could benefit from the incorporation of some of the fruit characteristics of highbush and southern highbush (organic acid composition, sweetness). A few cultivars are pentaploid first-generation crosses between highbush and rabbiteye (‘Pearl River’, ‘Robeson’), but breeders will also acknowledge that in general, among such first generation crosses, there is often low productivity, generally attributed to low fertility. Certain rabbiteye-highbush combinations seem to work better than others, but in most cases, ploidy has to be re-established at either the 4x or 6x level for good fertility and fruitfulness. However, imagine a rabbiteye x highbush cross where both parents have pronounced parthenocarpic tendencies. Would this produce hybrids with adequate fruit production in which fertility would be much less of a concern? We don't fully understand the interaction of parthenocarpy with fertility, and a threshold level of fertility would probably be needed for optimum results, but the possibilities are tantalizing.
CONCLUSIONS
The nature of parthenocarpy, as we have deduced it, thus far, for both highbush and rabbiteye blueberries is an imperfect system. Parthenocarpy, however, doesn't have to be a perfect system to be useful. In terms of fruit set and fruit development, even a modest improvement in fruit set and development could have substantial value. Our functional goal is to help the bees, not replace them!
ACKNOWLEDGMENT
I wish to thank Robert B. Martin Jr. for technical support.
Notes
This article not subject to US copyright law.
LITERATURE CITED
- Cano-Medrano , R. and Darnell , R.L. 1998 . Effect of GA3 and pollination on fruit set and development in rabbiteye blueberry . HortScience , 33 : 632 – 635 .
- Crane , J.C. 1964 . Growth substances in fruit setting and development . Ann. Rev. Plant Physiol. , 15 : 203 – 226 .
- Draper , A.D. and Scott , D.H. 1969 . Fruit size inheritance in highbush blueberries, Vaccinium australe small . J. Amer. Soc. Hort. Sci. , 94 : 417 – 418 .
- Ehlenfeldt , M.K. 2001 . Self- and cross-fertility in recently released highbush blueberry cultivars . HortScience , 36 : 133 – 135 .
- Ehlenfeldt , M.K. and Hall , M.R. 1996 . Metrical analysis of a putative source for semi-seedlessness in rabbiteye blueberry, Vaccinium ashei Reade . HortScience , 31 : 272 – 274 .
- Ehlenfeldt , M.K. and Vorsa , N. 2007 . Inheritance patterns of parthenocarpic fruit development in highbush blueberry (Vaccinium corymbosum L.) . HortScience , 42 : 1127 – 1130 .
- Ercan , N. and Akilli , M. 1996 . Reasons for parthenocarpy and the effects of various hormone treatments on fruit set in pepino (Solanum muricatum Ait.) . Sci. Hort. , 66 : 141 – 147 .
- Fellman , C. , Hoover , E. , Ascher , P.D. and Luby , J. 1991 . Gibberellic acid-induced seedlessness in field-grown vines of Swenson red grape . HortScience , 26 : 873 – 875 .
- Groot , S.P.C. , Bruinsma , J. and Karssen , C.M. 1987 . The role of endogenous gibberellin in seed and fruit development of tomato: Studies with a gibberellin-deficient mutant . Physiol. Plant. , 71 : 184 – 190 .
- Gustafson , F.G. 1939 . The cause of natural parthenocarpy . Amer. J. Bot. , 26 : 135 – 138 .
- Harrison , R.E. , Luby , J.J. and Ascher , P.D. 1994 . Pollen source affects yield components and reproductive fertility of four half-high blueberry cultivars . J. Amer. Soc. Hort. Sci. , 119 : 84 – 89 .
- Kender , W.J. and Desroches , J.C. 1970 . Changes in endogenous auxin, gibberellin-like substances, and inhibitors in developing lowbush blueberry fruits . J. Amer. Soc. Hort. Sci. , 95 : 699 – 702 .
- Kim , I.S. , Okubo , H. and Fujieda , K. 1994a . Studies on parthenocarpy in Cucumis sativus L.: IV. Effects of exogenous growth regulators on induction of parthenocarpy and endogenous hormone levels in cucumber ovaries . J. Kor. Soc. Hort. Sci. , 35 : 187 – 195 .
- Kim , I.S. , Yoo , K.C. , Fujieda , K. and Okubo , H. 1994b . Studies on parthenocarpy in Cucumis sativus L.: V. Influence of exogenous plant growth regulators on growth and diffusible IAA level of cucumber ovaries . J. Kor. Soc. Hort. Sci. , 35 : 196 – 200 .
- MacKenzie , K.E. 1997 . Pollination requirements of three highbush blueberry (Vaccinium corymbosum L.) cultivars . J. Amer. Soc. Hort. Sci. , 122 : 891 – 896 .
- Mainland , C.M. and Eck , P. 1968 . Induced parthenocarpic fruit development in highbush blueberry . Proc. Amer. Soc. Hort. Sci. , 92 : 284 – 289 .
- Mainland , C.M. and Eck , P. 1969 . Fruiting response of the highbush blueberry to gibberellic acid under field conditions . J. Amer. Soc. Hort. Sci. , 94 : 21 – 23 .
- Mainland , C.M. and Eck , P. 1971 . Endogenous auxin and gibberellin-like activity in highbush blueberry flowers and fruit . J. Amer. Soc. Hort. Sci. , 96 : 141 – 145 .
- Mudge , K.W. , Narayanan , K.R. and Poovaiah , B.W. 1981 . Control of strawberry Fragaria-ananassa cultivar Ozark Beauty fruit set and development with auxins . J. Amer. Soc. Hort. Sci. , 106 : 80 – 84 .
- Nesmith , D.S. , Krewer , G. , Rieger , M. and Mullinix , B. 1995 . Gibberellic acid-induced fruit set of rabbiteye blueberry following freeze and physical injury . HortScience , 30 : 1241 – 1243 .
- Ohara , H. , Izawa , J. , Kimura , S. , Hiroi , N. , Matsui , H. , Hirata , N. and Takahashi , E. 1997 . Induction of fruit set and growth of parthenocarpic ‘Hayward’ kiwifruit with plant growth regulators . J. Jap. Soc. Hort. Sci. , 66 : 467 – 473 .
- Pak , H.Y. 1993 . Effects of plant growth regulators on parthenocarpic fruit development in watermelon (Citrullus vulgaris Schrad.) . J. Korean Soc. Hort. Sci. , 34 : 167 – 172 .
- Talon , M. , Zacarias , L. and Primo , M.E. 1992 . Gibberellins and parthenocarpic ability in developing ovaries of seedless mandarins . Plant Physiol. , 99 : 1575 – 1581 .
- Weiss , J. , Nerd , A. and Mizrahi , Y. 1993 . Vegetative parthenocarpy in the cactus pear Opuntia ficus-indica (L.) Mill . Ann. Bot. , 72 : 521 – 526 .
- Wong , C.Y. 1941 . Chemically induced parthenocarpy in certain horticultural plants, with special reference to the watermelon . Bot. Gazette , 103 : 64 – 86 .
- Yamada , H. , Nakajima , K. , Yamazawa , Y. and Kuroi , I. 1991 . Effect of pollination and gibberellin treatments on fruit set and development of the European pear Pyrus communis var. sativa, cultivar Le Lectier . J. Jap. Soc. Hort. Sci. , 60 : 267 – 274 .