
ABSTRACT
Tree architecture describes plant form by defining the spatial organization of different structures. Shoot branching is an important step in understanding the tree architecture. Such studies are required for analyzing phenotypic diversity of plant architecture. Because such studies are rare on fig trees, the architecture diversity among nine Tunisian fig cultivars grown in Chott-Mariem (center-east Tunisia) was explored. Quantitative morphological descriptors were noted on six annual shoots and studied during four yearly growth cycles, from 2007–10. Coding strategy was adopted for fig branch description. Branch growth characteristics, location, and distribution of axillary shoots were determined for three axis orders. Results showed a great variability of fig branches, particularly in terms of branching density and position, branching angle, and shoot dimensions. Shoot length was closely related to its architectural position in the branch, i.e., all low-order shoots in all cultivars had more growth than high-order shoots. The meso-basitony tendency was frequent. Thus, an architectural typology characterizing all cultivars was established and three principal groups were distinguished. The first one (Type I) with orthotropic branches located at the basal and median parts of the bearer axis had dense and continuous ramification. The second group (Type II) with a diffuse branching differed in lateral shoot distribution, and finally, the third group (Type III) had open and diffuse branching located on the basal zone of the carrier axis. This typology could be used in future genetic improvement programs of local fig trees in Tunisia.
Introduction
The majority of fruit species are directly identifiable by their canopy or growth habit. These physiognomic concepts are certainly useful in the recognition of trees, but do not allow understanding their development. The purpose of the architectural analysis of trees is to interpret the total structure of the tree and include mechanisms that underlie its development. At each stage, a tree is analyzed in its totality, considering the whole of structures. Based on the observation of fundamental morphological characters (Caraglio and Barthélémy, Citation1997), this qualitative approach of tree development is a preliminary essential step to quantify and simulate plant organization. Several studies have quantified the various parts of a plant, i.e., growth unit, annual shoots, and axis (Aїachi-Mezghani et al., Citation2008; Ben Sadok et al., Citation2013; Costes and Guédon, Citation2002; Lauri and Térouanne, Citation1995; Lauri et al., Citation1997). Some horticultural and agronomic applications with regard to the distribution of vegetative and floral organs, developed for better management practices, were also applied at the tree and orchard scales (Costes, Citation2003; Costes et al., Citation2006; Lauri and Laurens, Citation2005; Lauri and Lespinasse, Citation2001). In this context, the study of architectural and morphological variability in fruit species is entirely applicable for clonal selection and control of tree cultural practices. Tree architecture is the result of a balance between its genetic potential and its environment. In orchards, the initial choice of cultivation system, plant spacing, and training system strongly affect tree growth and fruiting. The architectural analysis, a prerequisite to any reflection of tree management and also, in return, to better analyze the effect of training manipulations carried out on the growth and fruiting at the whole tree level, constitutes a basic, yet little explored, framework (Lauri and Laurens, Citation2005). In addition, the interactions between tree architecture and the agronomic practices then can constitute a second step in the investigations of the branching process of fruit trees, based on the knowledge of the tree developmental rules (Costes et al., Citation2006).
Thus, the integration of architectural traits in breeding programs is a promising way to improve the control of vegetative development and bearing regularity (Costes et al., Citation2004). Nevertheless, such research aspects on fig trees (Ficus carica L.) were limited in the literature and some of them have been studied by Lauri and Caraglio (Citation1993) and Caraglio and Barthélémy (Citation2005). Fig trees are grown in all of Tunisia with more than 2,500,000 trees occupying about 33,800 ha (MARH, Citation2012). There are several Tunisian fig cultivars with a good adaptation to local agro-ecological conditions. Figs are consumed both fresh and dried with a preference for fresh products (Mars et al., Citation2009). Indeed, fig tree constitutes an adequate model for architectural study considering the great varietal diversity of this species in Tunisia (Gaaliche et al., Citation2012; Mars et al., Citation2008). Moreover, understanding the organization of shoot growth and branching is an important step for the comprehension of the architecture of the whole tree. Such studies are crucial for better selection of morphological and architectural criteria for further use in genetic breeding programs of fig tree.
The objectives of this work were to: (i) quantify the branch characteristics of nine fig cultivars according to the different relative branching orders and evaluate the importance of the different annual growth unit, (ii) describe the branching pattern relating to the lateral shoot position and distribution along the main axis, and finally (iii) classify the studied cultivars with respect to main morphological criteria developed.
Materials and methods
Plant material
The research was carried out in an experimental fig orchard (altitude, 18 m; latitude, 35° 55ʹ N; longitude, 10° 34ʹ E) located at a coastal semi-arid bioclimatic zone characterized by mild winter, without frost periods, and hot dry summer. Rainfall is irregular and stormy. The soil is sandy loam and porous with alkaline pH. The study of the architectural development was followed during four growing seasons (2007–10) on nine fig cultivars (). Two representative trees for each cultivar were chosen because they had good sanitary status, and were 15 years old, spaced at 8 m × 8 m, and cultivated under rain-fed conditions. The cultivars belong to two distinct groups, i.e., the Smyrna-type cultivars usually do not produce Breba crop (first crop). Syconia of the Main crop (second crop) form and ripen only if pollinated. San Pedro-type cultivars usually initiate a Breba crop that persist until maturity (parthenocarpic). The second crop requires pollination to mature (Gaaliche et al., Citation2011).
Table 1. Studied fig cultivars.
Branch description and measurements
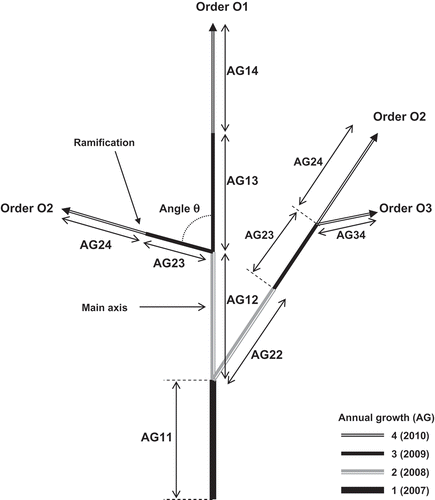
Three shoots of similar size, located at the periphery of each tree, were labeled in Autumn 2007 and their growth characteristics were assessed during the three following growing seasons. Observations and measurements were undertaken during winter, i.e., rest period after leaf fall (January and February), during 4 consecutive years. For fig branch description, we adopted the coding strategy for olive tree (Aїachi-Mezghani, Citation2009). The relative branching order was determined centrifugally, using the concept of relative order, beginning from the first main axis sampled at the tree periphery. The main axis was considered as the “first-order axis,” the second ramification on it as “second-order axis,” and the third ramification on it as “third-order axis” (). Length (cm), diameter (mm), internode number, internode length (mm), and branch angle (degree) relative to the parent shoot were recorded for four annual growths and three relative branching orders (). In fig tree, annual growth is defined as the shoot elongation developed during 1 year (Gaaliche et al., Citation2011). Branching density was expressed as the ratio of shoots number of order n (On) to the number of nodes on the branch of order n – 1 (On–1). Branching position was determined according to the different parts (basal, median, and top) of the main axis, irrespective of the annual shoots.
Figure 1. Architecture of a 4-year-old fig branch. Each annual growth (AG) is indexed by its branching order and year of growth, beginning in 2007 (1), respectively.
Topological representation of fig branches
Data collected for different fig branches were regrouped in the same file encrypted (MTG-Excel) according to a topological coding. This code preserves the topology information of the plant and represents the tree in the form of a multi-scale tree graph (MTG), which is associated with all measurements. This graph describes the topology of plants and associates with each measured botanical entity, qualitative, and quantitative attribute. The exploration of these multi-scale graphs is accessible by the AML language (AMAP; modeling language) or AML-Python (Godin et al., Citation1999). The following step was the modification of the file extension to ‘txt’ (MTG-txt) format and transfer in ‘Python’ software (Python Software Foundation, DE, USA). The first function ‘MTG’ permits to read the coding file and to check it. The second function ‘Plot’ permits to visualize the branch diagram. For each cultivar, topological representation of the branch was made using the average length of annual growths and those of branch angles for the six shoots. Branch diagrams were then treated and improved with Adobe Illustrator CS5 software (Adobe Systems Incorporated, CA, USA) used for graphic design.
Statistical analyses
For all morphological traits, data were analyzed statistically by running ANOVA of Statistical Package for the Social Sciences (SPSS) version 13.0 (SPSS Inc., Chicago, IL, USA). The mean values were compared using Duncan’s multiple range test. For annual shoot growth, data were subjected to univariate analysis using a general linear model (GLM) procedure from SPSS including two fixed factors (cultivar and year) in order to check the effect of each factor and their interactions.
Results and discussion
Branch characteristics
Differences among cultivars were statistically significant (P < 0.01) for all branch characteristics and relative branching orders (). For all cultivars, axis dimensions and internode number decreased when the relative branching order increased. In fact, total axis length ranged from 33.6 cm to 50.8 cm for order 1, from 12.3 cm to 25.5 cm for order 2, and from 2.9 cm to 13.6 cm, for order 3. For the first order, the shortest shoot was found in cultivar Zidi, while the longest one was registered in cultivar Chetoui. At orders 2 and 3, the shortest shoots were noted in cultivars Bidhi (12.3 cm) and Hemri (2.9 cm), respectively; while the longest values were observed in cultivars Khedhri (25.5 cm) and Chetoui (13.6 cm), respectively (). The first order grows up to 2.0 to 3.3 more than the second order and 3.5 to 14 more than the third order. It is important to mention that cultivar Zidi did not develop third-order branches. Concerning shoot vigor, the lowest diameters were found in Bidhi for all relative branching orders, while the highest values were registered in Khedhri for the first and second order (). The internode number showed a similar tendency to total shoot length in all cultivars. It varied from 32.6 (Zidi) to 41.1 (Khedhri), from 13.7 (Bidhi) to 23.7 (Khedhri), and from 3.0 (Hemri) to 14.0 (Soltani), for orders 1, 2, and 3, respectively. The average internode length varied considerably between cultivars and according to relative branching orders. It is important to mention that the highest internodes lengths were recorded in Chetoui for all orders (). Regarding shoot direction, differences among fig cultivars were statistically significant at P < 0.01 (). We noted also that branch angle with the bearer axis decreased when the relative branching order increased. Branch angle values showed a large variation between cultivars and for both relative branching orders 2 and 3. Indeed, it varied between 50.0° (Soltani) and 75.9° (Hemri) for order 2, and ranged from 37.5° (Soltani) to 65.0° (Khedhri) for order 3 (). The branches of order 2 were horizontal, particularly in some cultivars, such as Hemri, Khedhri, and Zidi with average branch angles varying between 65.0° and 75.9° ().
Table 2. Average values of total shoot length, diameter, number, and internode length according to the branching orders (1, 2, and 3) of nine fig cultivars.z
Table 3. Variation of branch angle according to the branching orders for studied fig cultivarsz.
Results showed different patterns of vegetative growth related to the cultivar and relative branch order. All low-order shoots issued from all cultivars had more growth than high-order shoots. This result corroborated other reports in fruit species as for Olea europaea (Aїachi-Mezghani, Citation2009; Ben Sadok et al., Citation2013), Prunus persica (Lauri, Citation1991), Malus × domestica (Renton et al., Citation2006), and other forest species like Juglans nigra, Juglans nigra × regia, and Cedrus atlantica (Sabatier and Barthélémy, Citation1999; Sabatier et al., Citation1998). A greater growth of lower-order branches is related to apical dominance. Indeed, many fig cultivars display strong apical dominance, resulting in seasonal growth either from the terminal bud or the adjacent 1–2 buds. Lack of apical dominance, due to modification by pruning, can result in denser lateral growth, leading to less productive, bushy-type trees. When a cultivar ordinarily (without pruning) has weak apical dominance, it naturally displays bushy-type growth without pruning, yet still performs well in terms of productivity (Şahin and Balci, Citation1998). Branch length is closely related to its architectural position in the tree (Barthélémy et al., Citation1997; Suzuki, Citation2000). Some cultivars, such as Chetoui and Bither Abiadh 2, developed long vegetative shoots, whereas others like Zidi and Besbessi were characterized by short shoots. These characteristics depend on the culture conditions, pruning system, annual rainfall, and agro-ecological conditions, but they are mostly determined by the cultivar (Mars et al., Citation2009). Results are in accordance with those reported by Chatti et al. (Citation2004) in fig collection in center-east Tunisia and by Gaaliche et al. (Citation2012) in fig cultivars grown in Djebba (north-west Tunisia). For fig axis direction, low-order axes showed branch angles with the parent shoot larger than those of the superior order. Similar observations were noted in walnut (Solar and Stampar, Citation2003) and olive trees (Aїachi-Mezghani, Citation2009).
Branching patterns
Considering the entire shoot, branching density showed a great variation between fig cultivars (). The second-order shoot number developed by the first order varied between 0.33 shoots per node in Hemri and 0.80 shoots per node in Chetoui. Thus, it appeared that the branching mode of Chetoui was more or less continuous, while other cultivars, such as Hemri, Khedhri, Zidi, and Bither Abiadh 1, showed lower density inferior to 0.4. The third-order shoots developed by the second one decreased and the branching density becomes very low, even null (). This parameter was used for tree architecture classification (Hallé et al., Citation1978) for many fruit species like peach (Scorza, Citation1984), pear (Sansavini and Musacchi, Citation1993), and olive trees (Aïachi-Mezghani, Citation2009). Bassi et al. (Citation1994) reported that the dwarf form peach had denser branching than standard or columnar form. This last form produced noticeably fewer second-order shoots. Aïachi-Mezghani (Citation2009) showed on olive that bushy model has a compact branch distribution while mixed and upright models developed a diffuse branching.
Table 4. Branching density for studied fig cultivars.
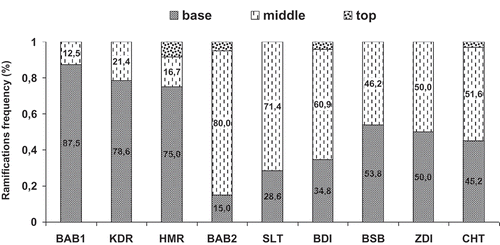
Positioning of the second-order shoots is illustrated in for each cultivar. For some cultivars, such as Bither Abiadh 1, Khedhri, and Hemri, lateral shoots were located on the basal zone of the parent axis with percentages of 87.5%, 78.6%, and 75%, respectively. While for others like ‘Bither Abiadh 2’, ‘Bidhi’, and ‘Soltani’, lateral axes were located in the median zone of the carrier axis and totaled 80.0%, 60.9%, and 71.4%, respectively (). Thus, these cultivars are characterized by a mesotonic branching along the parent axis. Moreover, for cultivars Besbessi and Chetoui, lateral shoots were developed in both parts (basal and median) of the bearer axis. Branch distribution in the distal zone was very weak even null in some cultivars, such as Bither Abiadh 1, Besbessi, Khedhri, Soltani, and Zidi (). It must be noticed that these gradients were analyzed along the parent axis, considered as a whole, and do not fully correspond to finer analyses performed at the annual shoot scale (Guédon et al., Citation2001). Other factors, such as branch vigor, competition between shoots, and gravimorphism are involved in this phenomenon (Barthélémy et al., Citation1997). The acrotonic tendency was rarely observed in the fig tree. This criterion was used for the description of tree architecture of many fruit species like peach, apricot, olive, and apple (Aїachi-Mezghani, Citation2009; Costes et al., Citation1996; Fournier, Citation1994; Lauri, Citation2007). Lauri and Caraglio (Citation1993) reported that fig tree exhibited a bushy architectural pattern by basal reiteration and natural layering. It is generally accepted that acrotony permitted the construction of tree form while basitony plays an important role in the construction of bushy form (Champagnat, Citation1947). The association between branch distribution patterns and tree form was not systematic (Bell, Citation1993; Lüttge et al., Citation1992). These concepts referred to the expression at the whole plant level rather than the shoot level. Plant architecture is the result of the balance between endogenous processes and exogenous constraints. Although environmental factors may modulate the sequence of differentiation, they almost never (except probably in extreme conditions) modify the inherent morphogenetic and ontogenetic constructional rules of plant organization (Barthélémy et al., Citation1997). For our study, it is important to mention that the studied trees were in the same orchard and were consequently submitted to the same environmental conditions.
Figure 2. Frequency of the ramifications of order 2 according to the insertion level on the branch in the different studied fig cultivars.
Annual shoot growth
Annual shoot length measured on fig branches exhibited fluctuations between years and differed according to relative branching order (). For order 1, some cultivars, such as Bither Abiadh 2, Besbessi, Bidhi, Khedhri, and Soltani, noted the most growth in shoot length on the last annual growth (2010), while others, such as Bither Abiadh 1 and Chetoui, showed more growth on the first annual growth (2007). For orders 2 and 3, no clear tendency was observed between annual growths for the different cultivars. A general trend of shoot vigor loss was noticed for the different annual growths with values ranging from 6.0 cm (Zidi) to 14.0 cm (Chetoui) for order 1, from 4.5 cm (Zidi, Soltani, and Bidhi) to 12.0 cm (Khedhri) for order 2, and from 2.3 cm (Bidhi) to 7.8 cm (Bither Abiadh 2) for order 3 (). It seems that annual shoot length varied between years and cultivars. However, statistical analysis confirmed this result where annual shoot length recorded for all relative branching orders differed depending significantly on the cultivar and year, even though the two factors had a significant interaction (). These results corroborate those of Aїachi-Mezghani (Citation2009) on olive tree. However, considering the low length of the annual shoots, the fluctuations observed with years do not correspond to ontogenetic gradients usually observed in trees and previously reported in forest (Caraglio et al., Citation1990; Jay-Allemand et al., Citation1995) and fruit trees (Costes et al., Citation2006; Ben Sadok et al., Citation2013; Renton et al., Citation2006). In our observations, the variations of annual shoot length can be attributed to different climatic conditions between the seasons (Ateyyeh and Sadder, Citation2006) and to fruiting conditions in each year (Gaaliche et al., Citation2011). Flaishman et al. (Citation2008) also mentioned that the decrease of temperature in autumn, the cold winter conditions, the growth temperature, and rain affect vegetative growth. Results were also in accordance with those of Ersoy et al. (Citation2008), stating that fig shoot length of Turkish cultivars ranged between 6.15 cm in Sarilop and 7.6 cm in Beyaz Orak. Whereas, Şahin and Balci (Citation1998) found that the average shoot length varied from 11.5 cm to 22.3 cm for Susak and Akça, respectively. These length values also reflect the growth level of mature trees that have overtaken most of their ontogenetic gradients and have reached an almost stable phase (Barthélémy and Caraglio, Citation2007).
Table 5. Average values of annual shoots length (cm) according to the branching orders (1, 2, and 3) for studied fig cultivarsz.
Branch architecture typology
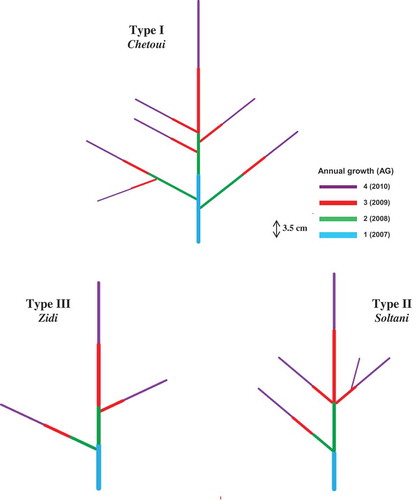
Typological classification of the different fig cultivars was established with respect to some morphological criteria, i.e., branch direction (orthotropic or plagiotropic), position (basitonic, mesotonic, and acrotonic), and ramification distribution (continuous or diffuse) (). This classification permitted us to regroup all cultivars in three principal groups, which were represented in . For all cultivars, branching mode was monopodial but differed among cultivars by other traits (). The first group (Type I) consisted of cultivars Bither Abiadh 2, Bidhi, and Chetoui with orthotropic branches located in the basal and median parts of the bearer axis and had dense and continuous ramification (, ). The cultivars Bither Abiadh 1, Besbessi, and Soltani formed the second group (Type II) with a diffuse branching and differ in lateral shoot position (, ), and finally the third group (Type III) constituted by Hemri, Khedhri, and Zidi, with open and diffuse branching (plagiotropic) located in the basal zone of the carrier axis (, ). In our study, we mentioned that studied fig trees were pruned and the natural forms were slightly modified. Lauri and Caraglio (Citation1993) studied three fig cultivars grown in France (Montpellier), and concluded that fig tree conformed to Rauh’s model proposed by Hallé et al. (Citation1978), i.e., all branches are orthotropic and monopodial, ramification is rhythmic, and localized at the top of each annual unit of extension (acrotony). Caraglio and Barthélémy (Citation2005) reported that fig tree develops a bushy architectural pattern by basal reiteration and natural layering. This result approved the basitony tendency showed for our studied fig branch architecture. Many classifications were also established based upon similar criteria and have been proposed for other fruit species, such as apple (Lespinasse and Delort, Citation1986), peach (Bassi et al., Citation1994), pear (Sansavini and Musacchi, Citation1993), and walnut trees (Germain, Citation1992).
Table 6. Typological classification of fig branches.
Figure 3. Topological representation of fig branch architecture for cultivars Chetoui (Type I), Soltani (Type II), and Zidi (Type III).
Conclusion
The present study allowed us to quantify how fig trees develop their branches and the pattern of lateral branch distribution. Results showed a different pattern of vegetative growth depending on the cultivar and relative branch order. All low-order shoots issued from all cultivars were longer than high-order shoots. A general trend of shoot vigor loss was observed for the different annual growths. Meso-basitonic tendency was frequently observed for the studied cultivars. Further investigations are required for predicting the fruiting location on the branch system and to analyze the relationship between vegetative and reproductive behavior within the tree. Gaaliche et al. (Citation2011) studied the interaction between annual vegetative growth and fruiting, thus concluding that fruit number in each fig generation was closely related to shoot length and more precisely to portion of shoots corresponding to growth rhythmicity within each year. Among cultivars, morphological variations were quantitatively based on the growth habit characteristics and ramification. They are mainly due to growth direction and branching position (basitonic and/or mesotonic). Most of these traits, although they appear to result from a genetic control can be modulated by exogenous factors. Moreover, these morphological characters were used to discriminate the studied fig cultivars. Therefore, some cultivars, such as Khedhri, Hemri, and Zidi, with open branching are considered as spreading and can thus be recommended for the extensive or semi-intensive cultivation system. By contrast, Soltani and Bidhi with more upright branching are classified as erected and can be advised for intensive plantations. These results give a basis for the selection of morphological and architectural criteria to take into account in future genetic improvement programs of fig tree.
Funding
This study is part of a research program of the Research Unit on Agro-biodiversity (UR03AGR04) financed by the Ministry of Higher Education and Scientific Research (Tunisia).
Acknowledgment
Authors would like to thank Mr. Foued Laabidi and Mrs. Ines Ben Sadok for their efficient collaboration.
Additional information
Funding
Literature cited
- Aїachi-Mezghani, M. 2009. Déterminisme de la diversité caulinaire et radiculaire chez l’olivier (Olea europaea L.) et modélisation de la croissance. Approche morphogénétique. Institut National Agronomique de Tunisie, Thèse de Doctorat en Sciences Agronomiques, 353 p.
- Aїachi-Mezghani, M., A. Sahli, F. Labidi, K. Meddeb, A. Jebari Ben El, and S. Hadj. 2008. Analysis of primary and secondary growth and modelling growth dynamics of olive shoots (Olea europaea L.). J. Hort. Sci. Biotechnol. 83:411–419.
- Ateyyeh, A.F. and M.T. Sadder. 2006. Preliminary study on the vegetative and reproductive growth of six common fig (Ficus carica L.) cultivars in Jordan. Jordan J. Agr. Sci. 2:1–7.
- Barthélémy, D. and Y. Caraglio. 2007. Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Annu. Bot. 99:375–407.
- Barthélémy, D., Y. Caraglio, and E. Costes. 1997. Architecture, gradients morphogénétiques et âge physiologique chez les végétaux, p. 89–136. In: J. Bouchon, P. de Reffye, and D. Barthélémy (eds.). Modélisation et simulation de l’architecture des végétaux. Science Update, INRA Editions, Paris, France.
- Bassi, D., A. Dima, and R. Scorza. 1994. Tree structure and pruning response of six peach growth forms. J. Amer. Soc. Hort. Sci. 119:378–382.
- Bell, A.D. 1993. Les plantes à fleurs. Guide morphologique illustré. Edition, 342 p. Masson, Paris, Milan, Barcelone Bonn.
- Ben Sadok, I., N. Moutier, G. Garcia, F. Dosba, N. Grati- Kamoun, A. Rebai, B. Khadari, and E. Costes. 2013. Genetic determinism of the vegetative and reproductive traits in a F1 olive tree progeny: Evidence of tree ontogeny effect. Tree Genet. Genomes 9:205–221.
- Caraglio, Y. and D. Barthélémy. 1997. Revue critique des termes relatifs à la croissance et à la ramification des tiges des végétaux vasculaires, p. 11–88. In: J. Bouchon, P. de Reffye, and D. Barthélémy (eds.). Modélisation et simulation de l’architecture des végétaux. Sciences Update, Editions INRA, Paris, France.
- Caraglio, Y. and D. Barthélémy. 2005. Revue critique des termes relatifs à la croissance et à la ramification des tiges des végétaux vasculaires. Arbres. Sci. 13:1–61.
- Caraglio, Y., E. Elguero, I. Mialet, and H. Rey. 1990. Le peuplier: Modélisation et stimulation de son architecture (Populus nigra L. cultivars, exemple des clones I 45/54 et I 214). Rapport de convention I.D.F/unité de modélisation des plantes du C.I.R.A.D/G.E.R.D.A.T.
- Champagnat, P. 1947. Les principes généraux de la ramification des végétaux ligneux. Rev. Hort. 30:335–341.
- Chatti, K., A. Salhi-Hannachi, M. Mars, M. Marrakchi, and M. Trifi. 2004. Analyse de la diversité génétique de cultivars tunisiens de figuier (Ficus carica L.) à l’aide de caractères morphologiques. Fruits 59:49–61.
- Costes, E. 2003. Winter bud content according to position in 3-year-old branching systems of ‘Granny Smith’ apple. Ann. Bot. 92:581–588.
- Costes, E. and Y. Guédon. 2002. Modelling branching patterns on 1-year-old trunks of six apple cultivars. Annu. Bot. 89:513–524.
- Costes, E., Y. Guédon, M. Jay, A. Audubert, and J. Lichou. 1996. Modeling of flower and fruit distribution in relation to shoot organisation and apricot tree architecture. Acta Hort. 384:291–295.
- Costes, E., P.E. Lauri, F. Laurens, N. Moutier, A. Belouin, F. Delort, J.M. Legave, and J.L. Regnard. 2004. Morphological and architectural traits on fruit trees which could be relevant for genetic studies: A review. Acta Hort. 663:349–356.
- Costes, E., P.E. Lauri, and J.L. Regnard. 2006. Analysing fruit tree architecture. Implication for tree management and fruit production. Hort. Rev. 32:1–61.
- Ersoy, N., S. Gözlekçi, L. Kaynak, B. Dal, and K. Yazıcı. 2008. Relationships among the shoot, leaf and fruit growth of some fig (Ficus carica) cultivars grown in Antalya conditions. Acta Hort. 798:143–148.
- Flaishman, M.A., V. Rover, and E. Stover. 2008. The fig: Botany, horticulture, and breeding. Hort. Rev. 34:113–197.
- Fournier, D. 1994. Analyse et modélisation des processus de croissance et développement qui contribuent aux performances agronomiques du Pêcher Prunus persica (L.) Batsch. de l’Université de Montpellier, France, Thèse de Doctorat en Sciences Agronomiques, 142 p.
- Gaaliche, B., P.E. Lauri, M. Trad, E. Costes, and M. Mars. 2011. Interactions between vegetative and generative growth and between crop generations in fig tree (Ficus carica L.). Sci. Hort. 131:22–28.
- Gaaliche, B., O. Saddoud, and M. Mars. 2012. Morphological and pomological diversity of fig (Ficus carica L.) cultivars in northwest of Tunisia. ISRN Agron. 2012:1–9.
- Germain, E. 1992. Le noyer, p. 620–632. In: A. Gallais and H. Bannerot (eds.). Dans: Amélioration des espèces végétales. Objectifs et Critères de Sélection. INRA, Paris, France.
- Godin, C., Y. Guédon, and E. Costes. 1999. Exploration of a plant architecture database with the AMAPmod software illustrated on an apple tree hybrid family. Agronomie 19:163–184.
- Guédon, Y., D. Bathélémy, Y. Caraglio, and E. Costes. 2001. Pattern analysis in branching and axillary flowering sequences. J. Theor. Biol. 212:481–520.
- Hallé, F., R.A.A. Oldeman, and P.B. Tomlinson. 1978. Tropical trees and forests. An architectural analysis, 441 p. Springer Verlag, Berlin.
- Jay-Allemand, C., P. Doumas, B. Sotta, H. Tranvan, and E. Miginiac. 1995. Juvenility and physiology of rhizogenesis in two woody species (Sequoia semper.irens and Juglans nigra × Juglans regia), p. 79–99. In: H. Saudermann Jr. and M. Bonnet-Masimbert (eds.). EUROSILVA contribution to forest tree physiology. INRA Editions, Paris.
- Lauri, P.E. 1991. Données sur l’évolution de la ramification et de la floraison du pêcher (Prunus percica L. Batsth) au cours de la croissance. Annu. Sci. Nat. Bot. Biol. Végét. 11:95–103.
- Lauri, P.E. 2007. Differentiation and growth traits associated with acrotony in the apple tree (Malus × domestica, Rosaceae). Annu. Bot. 94:1273–1281.
- Lauri, P.E. and Y. Caraglio. 1993. Architectural analysis of fig trees as a tool to study genetic diversity in cultivars. Acta Hort. 349:265–267.
- Lauri, P.E. and J.M. Lespinasse. 2001. Genotype of apple trees affects growth and fruiting responses to shoot bending at various times of year. J. Am. Soc. Hort. Sci. 126:169–174.
- Lauri, P.E. and F. Laurens. 2005. Architectural types in apple (Malus × domestica Borkh.), p. 1300–1314. In: R. Dris (ed.). Crops: Growth, quality and biotechnology. World Food Limited, Helsinki, Finland.
- Lauri, P.E. and E. Térouanne. 1995. Analyse de la croissance primaire de rameaux de pommier (Malus domestica Borkh.) au cours d’une saison de végétation. Can. J. Bot. 73:1471–1489.
- Lauri, P.E., E. Térouanne, and J.M. Lespinasse. 1997. Relationship between the early development of apple fruiting branches and the regularity of bearing. An approach to the strategies of various cultivars. J. Hort. Sci. 72:519–530.
- Lespinasse, J.M. and J.F. Delort. 1986. Apple tree management in vertical axis: Appraisal after ten years of experiments. Acta Hort. 160:120–155.
- Lüttge, U., M. Kluge, and G. Bauer. 1992. Botanique. Traité fondamental. 3rd ed., 574 p. Edition Technique et Documentation, Paris.
- MARH (Ministère de l’Agriculture et des Ressources Hydrauliques). 2012. Budget économique 2012. Agriculture et Pêche, Tunis (Tunisie).
- Mars, M., K. Chatti, O. Saddoud, A. Salhi-Hannachi, M. Trifi, and M. Marrakchi. 2008. Fig cultivation and genetic resources in Tunisia: An overview. Acta Hort. 798:27–32.
- Mars, M., B. Gaaliche, I. Ouerfelli, and S. Chouat. 2009. Systèmes de production et ressources génétiques du figuier (Ficus carica L.) à Djebba et Kesra, deux villages de montagne au nord ouest de la Tunisie. Rev. Régions Arides. 22:33–45.
- Renton, M., Y. Guédon, C. Godin, and E. Costes. 2006. Similarities and gradients in growth unit branching patterns during tree ontogeny based on a stochastic approach in ‘Fuji’ apple trees. J. Exp. Bot. 57:3131–3143.
- Sabatier, S. and D. Barthélémy. 1999. Growth dynamics and morphology of annual shoots, according to their architectural position, in young Cedrus atlantica (Endl.) Manetti ex Carrière (Pinaceae). Ann. Bot. 84:387–392.
- Sabatier, S., D. Barthélémy, I. Ducousso, and E. Germain. 1998. Modalités d’allongement et morphologie des pousses annuelles chez le noyer commun, Juglans regia L., ‘Lara’ (Juglandaceae). Can. J. Bot. 76:1253–1264.
- Şahin, B. and B. Balci. 1998. Studies on phenological and pomological properties of Breba fruit producing fig cultivars. Acta Hort. 480:91–99.
- Sansavini, S. and S. Musacchi. 1993. Canopy architecture, training and pruning in the modern orchard. Proc. Sixth Int. Symp. Pear Growing, Medford, Oregon, 12–14 July 1993. (Acta Hort. 367:152–172).
- Scorza, R. 1984. Characterization of four distinct peach tree growth types. J. Amer. Soc. Hort. Sci. 109:455–457.
- Solar, A. and F. Stampar. 2003. Genotypic differences in branching patterns in fruiting habit in common walnut (Juglans regia L.). Annu. Bot. 92:317–322.
- Suzuki, A. 2000. Patterns of vegetative growth and reproduction in relation to branch orders: The plant as a spatially structured population. Trees 14:329–333.